

Phenylbenzohydrazides Obtained from Isatoic Anhydride Present Anti-Inflammatory Activity In Vivo and In Vitro

 , , and
, , and 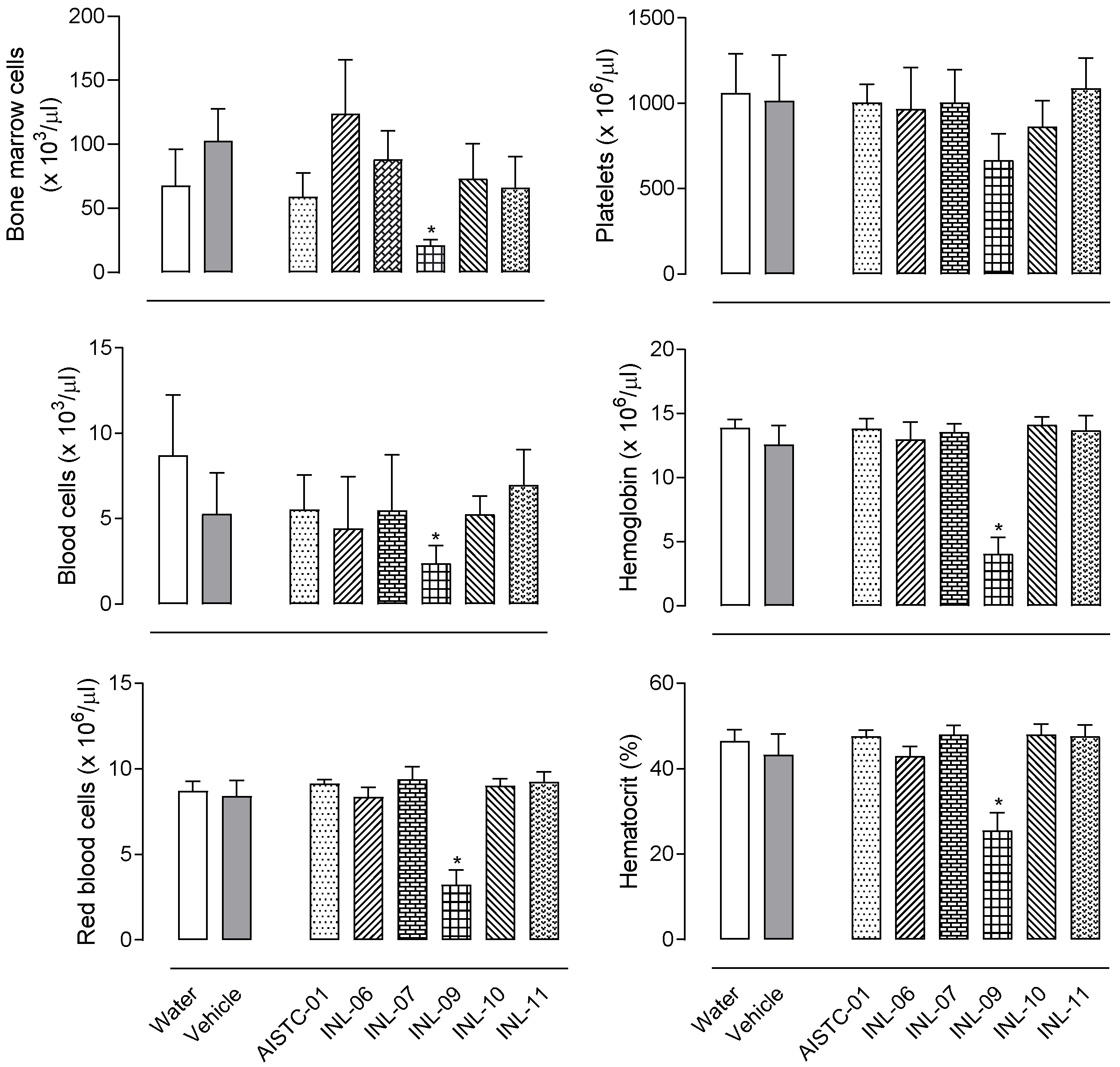{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
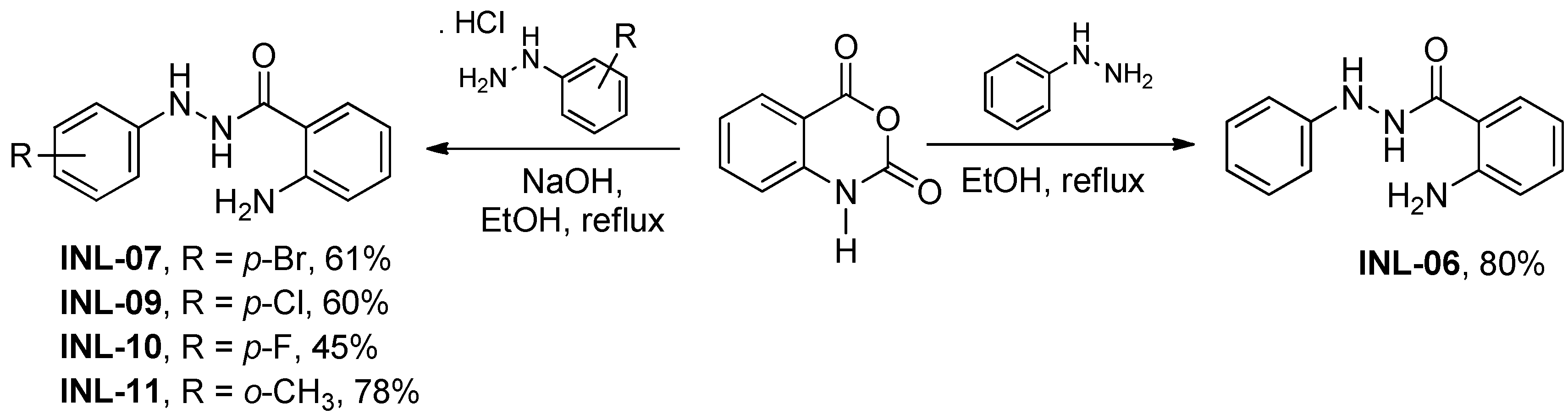
2.1. Synthesis of INLs
2.2. Animals
2.3. Drugs, Reagents, and Treatments
2.4. Cell Culture
2.5. In Vitro Toxicity Test (Cell Viability)
2.6. In Vivo Toxicity Test
2.7. Carrageenan-Induced Inflammation into the Subcutaneous Air Pouch (SAP)
2.8. Quantification of Proteins and Cytokines
2.9. Quantification of Nitric Oxide (NO) Production
2.10. Cell Migration In Vitro
2.11. Statistical Analysis
3. Results
3.1. Chemistry
3.2. None of INLs Did Induce Any Toxicity after Oral Administration
3.3. Leukocytes Migration and Protein Extravasation Were Inhibited by INLs
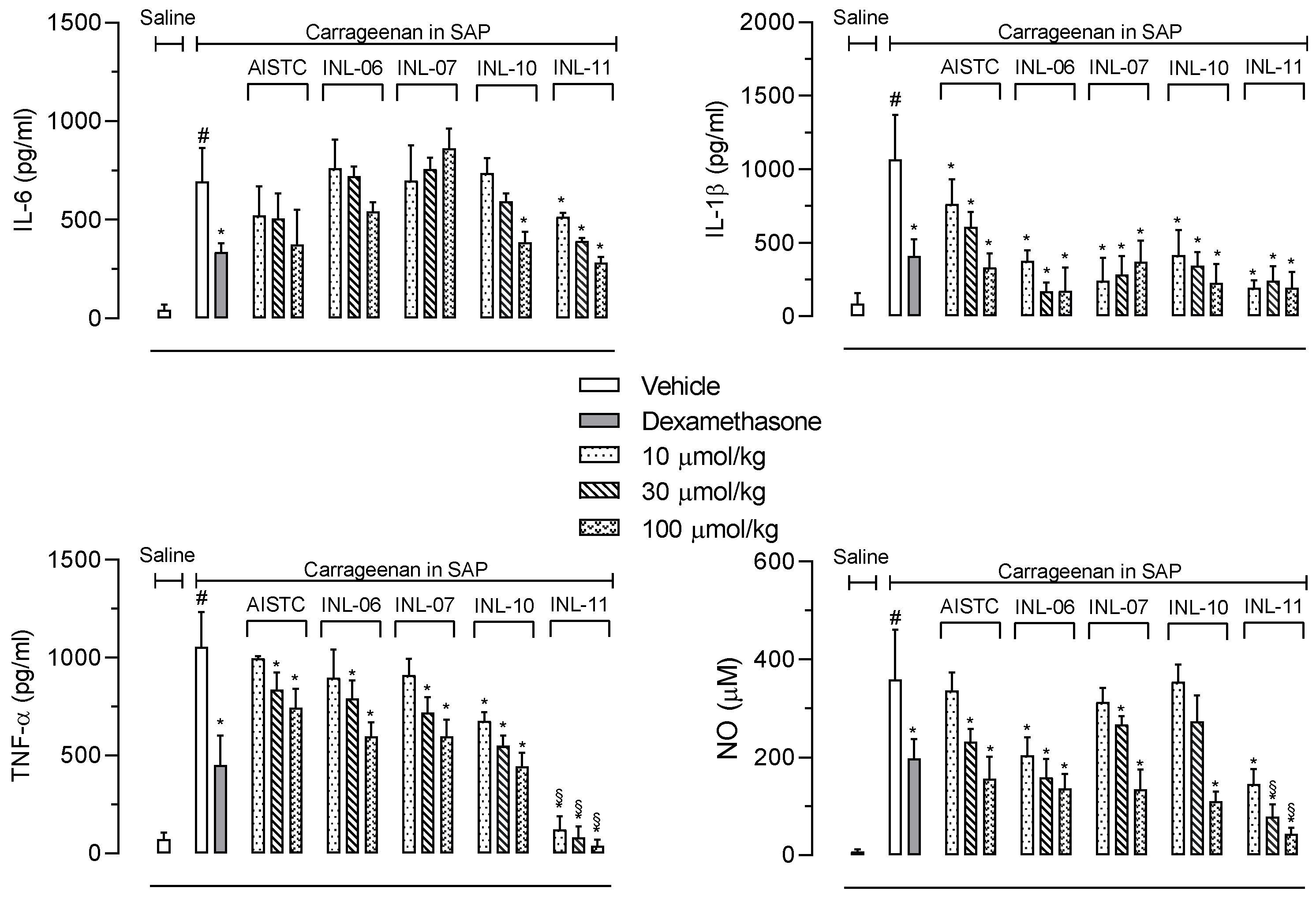
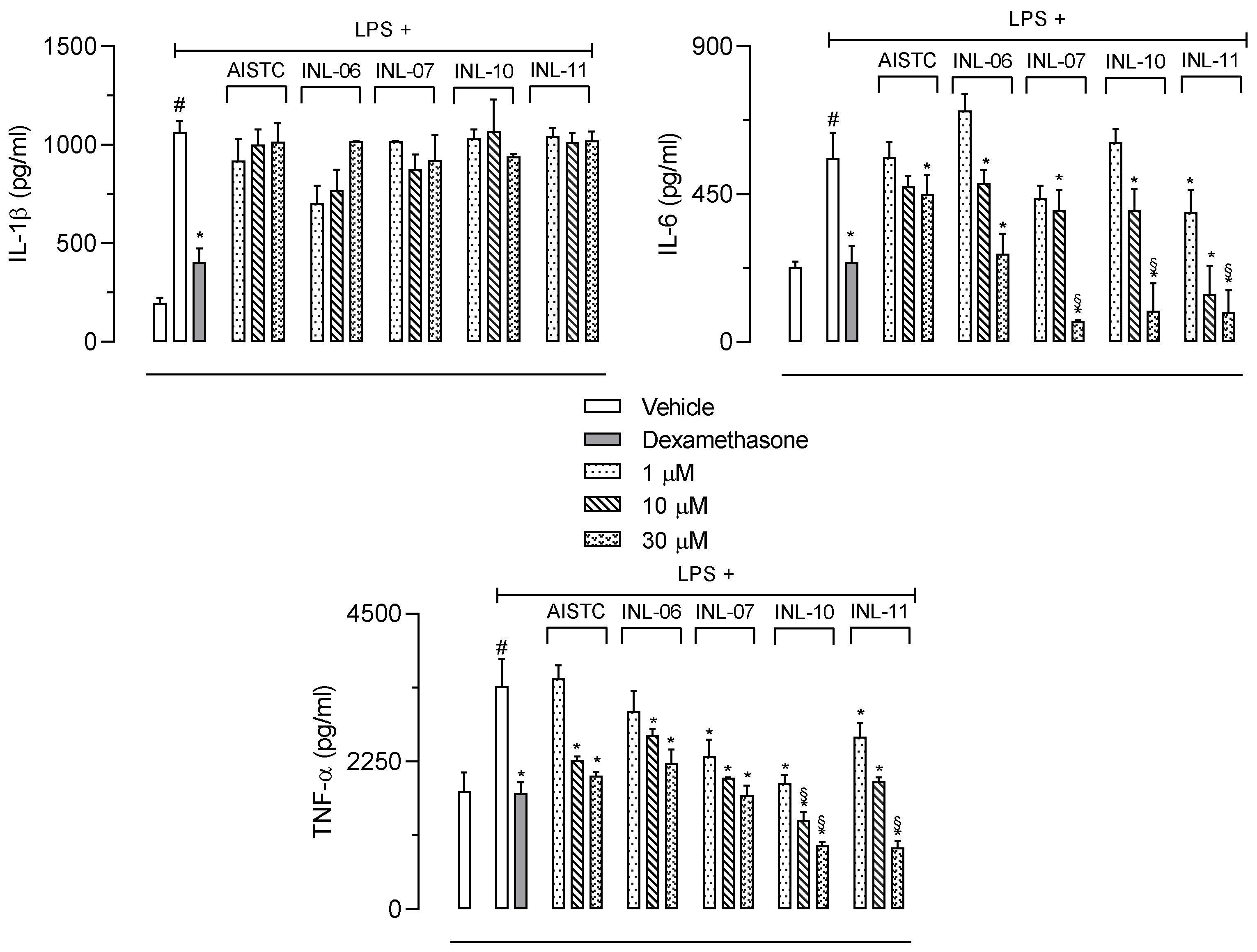
3.4. INLs Inhibited Inflammatory Mediators Produced in the Subcutaneous Air Pouch
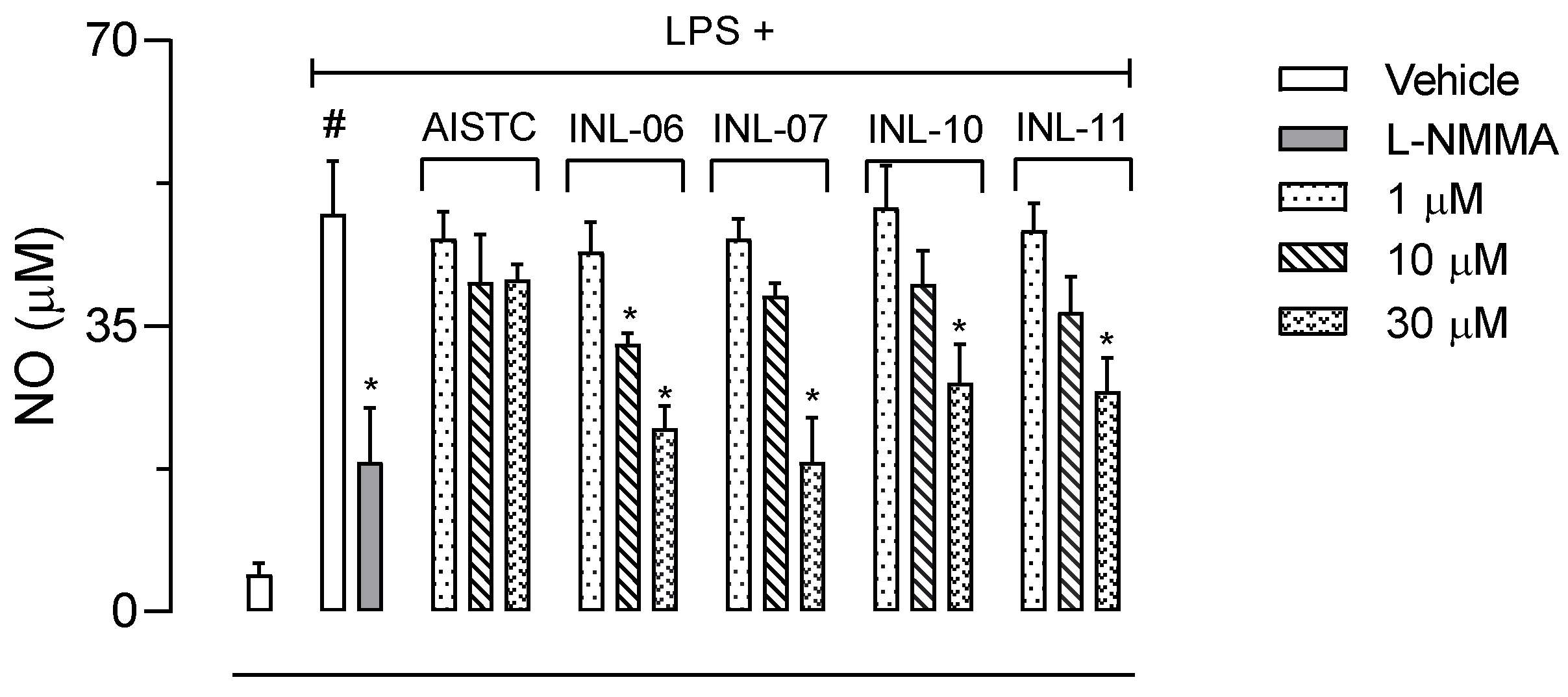
3.5. AISTC and INLs Did Not Present Cytotoxicity and Reduced Inflammatory Mediators’ Production and Cell Migration In Vitro
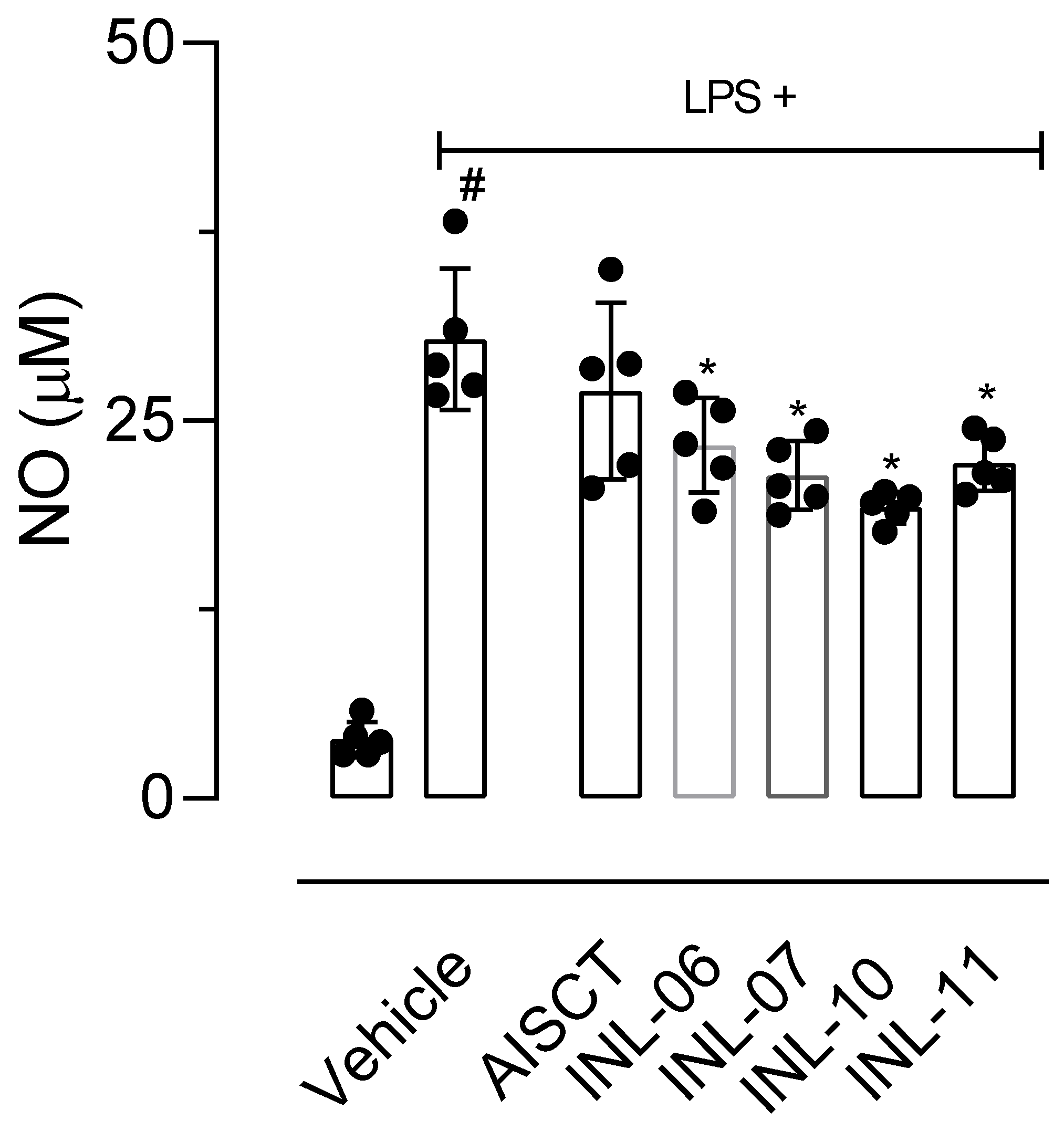
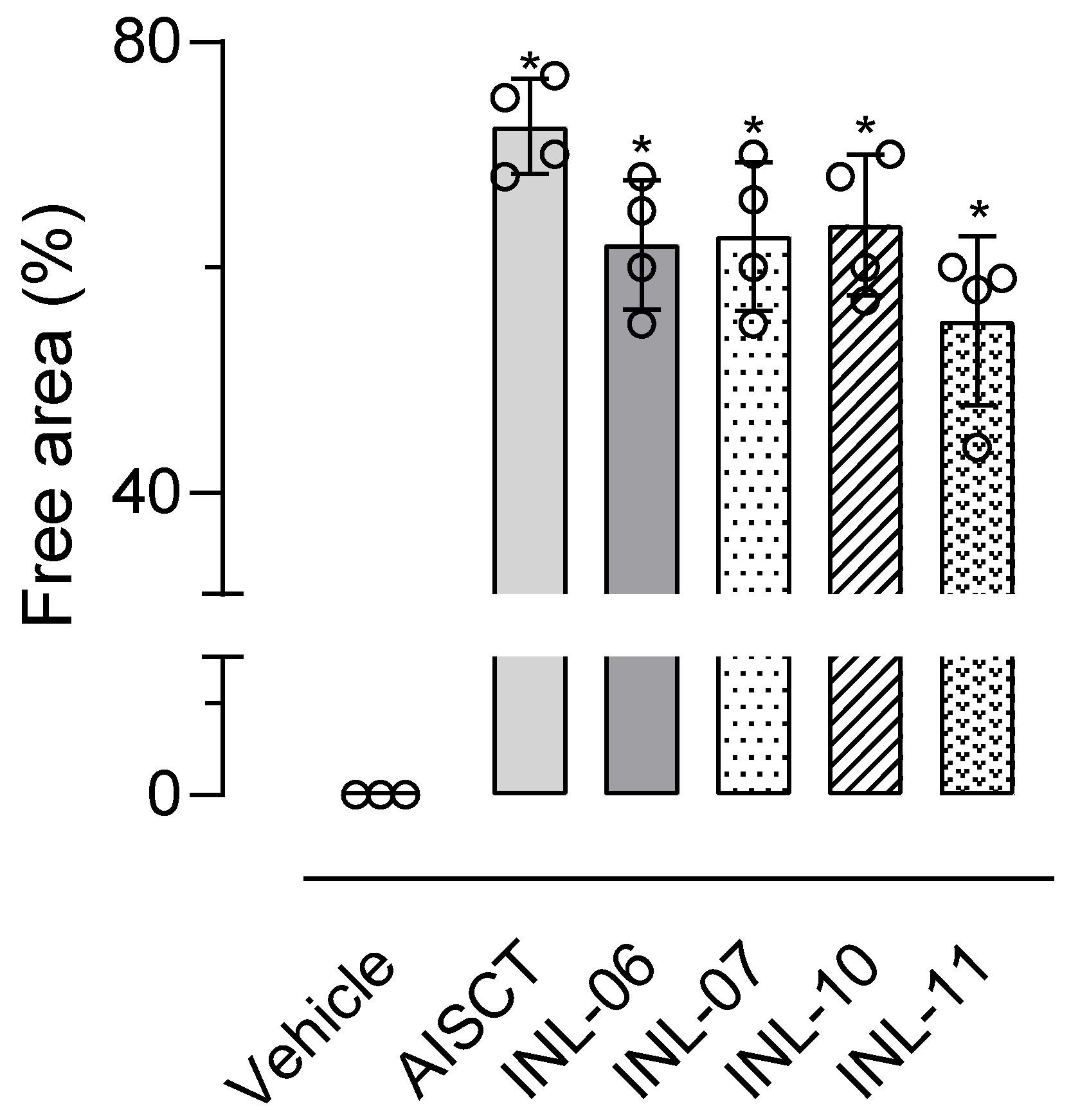
3.6. INLs Reduced Cell Migration In Vitro
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Fioranelli, M.; Roccia, M.G.; Flavin, D.; Cota, L. Regulation of inflammatory reaction in health and disease. Int. J. Mol. Sci. 2021, 22, 5277. [Google Scholar] [CrossRef] [PubMed]
- Timmermans, S.; Souffriau, J.; Libert, C. A General introduction to glucocorticoid biology. Front. Immunol. 2019, 10, 1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, S.M.; Abo-Kul, M.; Soltan, M.K.; Helal, A.S. Synthesis and anti-inflammatory activity of new 2-arylidenehydrazinyl-quinazolinone and 3-amino-triazoloquinazolinone derivatives. Asian J. Res. Chem. Pharmac. Sci. 2013, 1, 59–70. [Google Scholar]
- Giorno, T.B.S.; Ballard, Y.L.L.; Cordeiro, M.S.; Silva, B.V.; Pinto, A.C.; Fernandes, P.D. Central and peripheral antinociceptive activity of 3-(2-oxopropyl)-3-hydroxy-2-oxindoles. Pharmacol. Biochem. Behav. 2015, 135, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Giorno, T.B.S.; da Silva, B.V.; Pinto, A.C.; Fernandes, P.D. Antinociceptive effect and mechanism of action of isatin, N-methyl isatin and oxopropyl isatin in mice. Life Sci. 2016, 151, 189–198. [Google Scholar] [CrossRef]
- Shemchuk, L.A.; Chernykh, V.P.; Krys’kiv, O.S. Reaction of anthranilic acid 2-N-phenylhydrazide with cyclic anhydrides. Russ. J. Org. Chem. 2008, 44, 1006–1008. [Google Scholar] [CrossRef]
- Yang, W.; Qiao, R.; Chen, J.; Huang, X.; Liu, M.; Gao, W.; Ding, J.; Wu, H. Palladium-catalyzed cascade reaction of 2-amino-N-arylbenzohydrazides with triethyl orthobenzoates to construct indazolo[3,2-b]quinazolinones. J. Org. Chem. 2015, 80, 482–489. [Google Scholar] [CrossRef]
- Anand, D.; Patel, O.P.S.; Maurya, R.K.; Kant, R.; Yadav, P. Substrate controlled synthesis of benzisoxazole and benzisothiazole derivatives via PhI(OAc)2-mediated oxidation followed by intramolecular oxidative o−n/s−n bond formation. J. Org. Chem. 2015, 80, 12410–12419. [Google Scholar] [CrossRef]
- Zimmermann, M. Ethical guidelines for investigation of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Romano, M.; Faggioni, R.; Sironi, M.; Sacco, S.; Echtenacher, B.; Di Santo, E.; Salmona, M.; Ghezzi, P. Carrageenan-induced acute inflammation in the mouse air pouch synovial model. Role of tumour necrosis factor. Mediat. Inflamm. 1997, 6, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Raymundo, L.J.R.P.; Guilhon, C.C.; Alviano, D.S.; Matheus, M.E.; Antoniolli, A.R.; Cavalcanti, S.C.H.; Alves, P.B.; Alviano, C.S.; Fernandes, P.D. Characterisation of the anti-inflammatory and antinociceptive activities of the Hyptis pectinata (L.) Poit essential oil. J. Ethnopharmacol. 2011, 134, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Bartholomew, B. A rapid method for the assay of nitrate in urine using the nitrate reductase enzyme of Escherichia coli. Food Chem. Toxicol. 1984, 22, 541–543. [Google Scholar] [CrossRef] [PubMed]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wisnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [5N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Sen, I.; Azizoglu, A. Computational study on the structure of N-(2-aminobenzoyl)-N’-phenyl hydrazine. In Proceedings of the 17th International Electronic Conference on Synthetic Organic Chemistry, online, 1–30 November 2013; Volume 17. [Google Scholar]
- Mason, T.J. Use of ultrasound in chemical synthesis. Ultrasonics 1986, 24, 245. [Google Scholar] [CrossRef]
- Einhorn, C.; Einhorn, J.; Luche, J.L. Sonochemistry—The use of ultrasonic waves in Synthetic Organic Chemistry. Synthesis 1989, 41, 787. [Google Scholar] [CrossRef]
- Luche, J.-L. Effect of ultrasound on heterogeneous systems. Ultrason. Sonochem. 1994, 1, S111–S118. [Google Scholar] [CrossRef]
- Bindu, S.; Mazumber, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharmacol. 2020, 180, 114147. [Google Scholar] [CrossRef]
- Weyand, C.M.; Goronzy, J.J. The immunology of rheumatoid arthritis. Nat. Immunol. 2021, 22, 10–12. [Google Scholar] [CrossRef]
- Wang, J. Neutrophils in tissue injury and repair. Cell Tissue Res. 2018, 371, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Zindel, J.; Kubes, P. DAMPs, PAMPs, and LAMPs in immunity and sterile inflammation. Annu. Rev. Pathol. 2020, 15, 493–518. [Google Scholar] [CrossRef] [PubMed]
- Selders, G.S.; Fetz, A.E.; Radic, M.Z.; Bowlin, G.L. An overview of the role of neutrophils in innate immunity, inflammation and host-biomaterial integration. Regen Biomater. 2017, 4, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.L.; Mogildea, M.; Moreno, I.; Lopes, A. Acute inflammation and metabolism. Inflammation 2018, 41, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Cinelli, M.A.; Do, H.T.; Miley, G.P.; Silverman, R.B. Inducible nitric oxide synthase: Regulation, structure, and inhibition. Med. Res. Rev. 2020, 40, 158–189. [Google Scholar] [CrossRef]
- Salim, T.; Sershen, C.L.; May, E.E. Investigating the role of TNF-α and IFN-γ activation on the dynamics of iNOS gene expression in LPS stimulated macrophages. PLoS ONE 2016, 11, e0153289. [Google Scholar] [CrossRef] [Green Version]
- Kiss, A. L Inflammation in Focus: The Beginning and the End. Pathol. Oncol. Res. 2022, 27, 1610136. [Google Scholar] [CrossRef]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.J.; Liu, J.X.; Xie, Y.; Luo, P.; Liu, Z.Q.; Liu, L.; Zhou, H. Suppression of macrophage migration by down-regulating Src/FAK/P130Cas activation contributed to the anti-inflammatory activity of sinomenine. Pharmacol. Res. 2021, 167, 105513. [Google Scholar] [CrossRef]
- Cui, S.; Wu, Q.; Wang, J.; Li, M.; Qian, J.; Li, S. Quercetin inhibits LPS-induced macrophage migration by suppressing the iNOS/FAK/paxillin pathway and modulating the cytoskeleton. Cell Adhes. Migr. 2019, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Arasteh-Fard, Z.; Dilmaghani, K.A.; Saeedi, M.; Mahdavi, M.; Shafiee, A. Synthesis of novel phthalazino[1,2-b]quinazolinedione derivatives: Efficient and practical reaction of 2-amino-N’-arylbenzohydrazides and 2-formylbenzoic acids. Helv. Chim. Acta 2016, 99, 539–542. [Google Scholar] [CrossRef]
- Kesternich, V.; Gahona, P.; Pérez-Fehrmann, M.; Brito, I.; López-Rodríguez, M. 2-Amino-N’-phenylbenzohydrazide. Acta Cryst. E 2012, 68, o1847. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paiva, J.P.B.; Cordeiro, M.S.; França, P.R.C.; Branco, L.O.P.; Santos, I.S.; Reis, N.F.; Pimentel, P.P.; Giorno, T.B.S.; Lima, E.C.; Fernandes, P.D. Phenylbenzohydrazides Obtained from Isatoic Anhydride Present Anti-Inflammatory Activity In Vivo and In Vitro. Biomolecules 2022, 12, 1901. https://doi.org/10.3390/biom12121901
Paiva JPB, Cordeiro MS, França PRC, Branco LOP, Santos IS, Reis NF, Pimentel PP, Giorno TBS, Lima EC, Fernandes PD. Phenylbenzohydrazides Obtained from Isatoic Anhydride Present Anti-Inflammatory Activity In Vivo and In Vitro. Biomolecules. 2022; 12(12):1901. https://doi.org/10.3390/biom12121901
Chicago/Turabian StylePaiva, João Pedro Barros, Millena Santos Cordeiro, Patricia Ribeiro Carvalho França, Luiz Octavio Pereira Branco, Isabela Souza Santos, Nanashara Figueiredo Reis, Patrick Pedro Pimentel, Thais Biondino Sardella Giorno, Evanoel Crizanto Lima, and Patricia Dias Fernandes. 2022. "Phenylbenzohydrazides Obtained from Isatoic Anhydride Present Anti-Inflammatory Activity In Vivo and In Vitro" Biomolecules 12, no. 12: 1901. https://doi.org/10.3390/biom12121901