
Effects of the Myokine Irisin on Stromal Cells from Swine Adipose Tissue

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Adipose Stromal Cells (ASCs) from Swine Abdominal Subcutaneous Adipose Tissue
2.2. ASC Growth
2.2.1. Cell Proliferation
2.2.2. Metabolic Activity
2.3. ASC Redox Status Parameters
2.3.1. ASCs Superoxide Anion (O2−) Production
2.3.2. ASCs Nitric Oxide (NO) Production
2.3.3. ASCs Superoxide Dismutase (SOD) Activity
2.3.4. ASCs Non-Enzymatic Scavenging Activity
2.4. ASC Adipogenic Differentiation
- Three days with an induction medium: culture medium supplemented with dexamethasone (DEX) 1 μM, insulin (INS) 1.7 μM, 3-iso-butyl-1-methylxanthine 0.5 mM, and indomethacin 250 μM;
- One day with a differentiation maintaining medium: culture medium supplemented with INS 1.7 μM.
2.4.1. Solubilization and Quantification of Oil Red O Production
2.4.2. Oil Red O Staining
2.4.3. Gene Expression Analysis
2.5. Statistical Analysis
3. Results
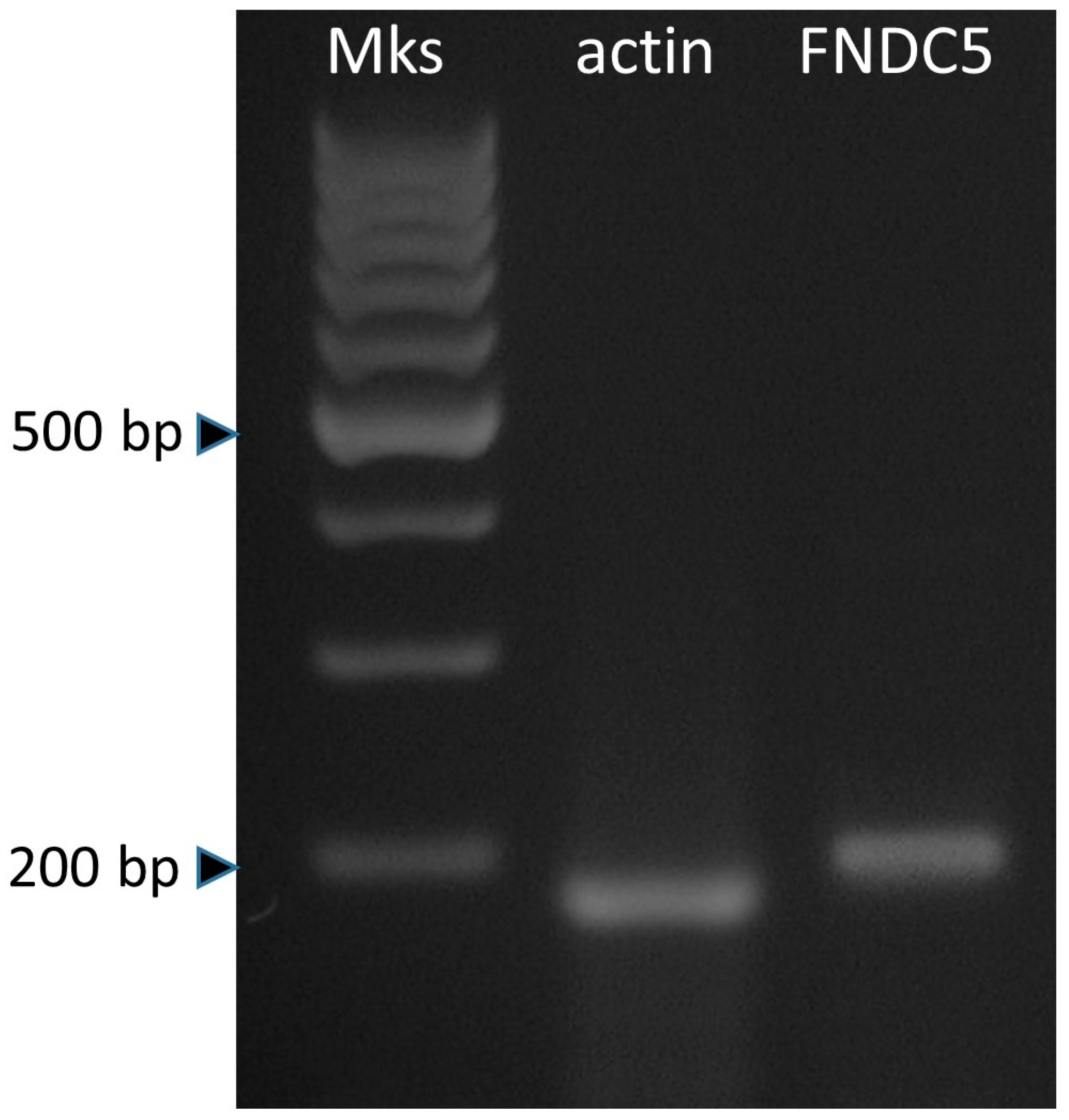
3.1. Expression of Irisin and the Effect on ASC Growth
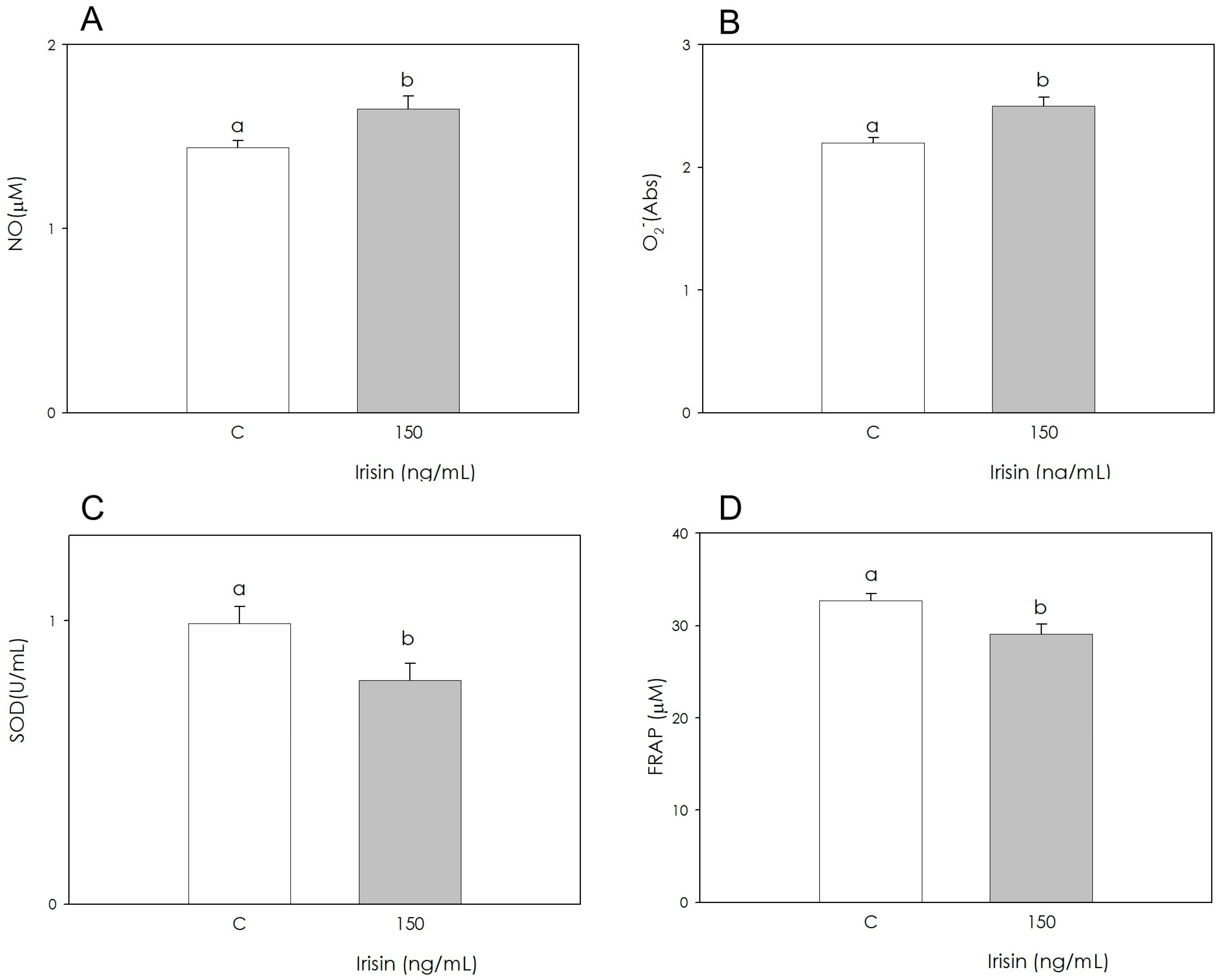
3.2. Irisin Effects on Redox Status Parameters
3.3. Changes in FNDC5, PPARγ and LEPTIN Gene Expression during Adipogenic Differentiation and Irisin Treatment
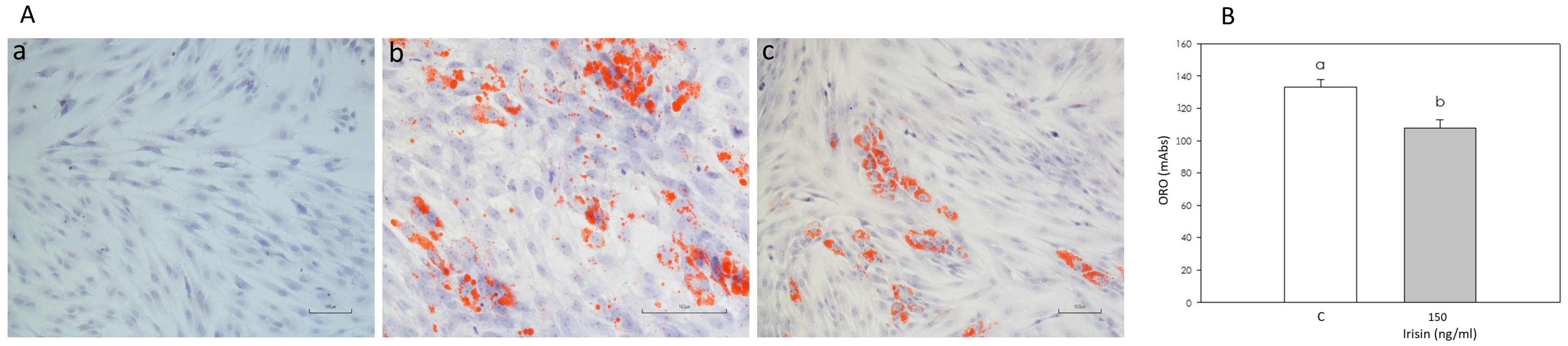
3.4. Effect of Irisin on Adipogenic Differentiation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zunner, B.E.M.; Wachsmuth, N.B.; Eckstein, M.L.; Scherl, L.; Schierbauer, J.R.; Haupt, S.; Stumpf, C.; Reusch, L.; Moser, O. Myokines and Resistance Training: A Narrative Review. Int. J. Mol. Sci. 2022, 23, 3501. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Qi, B.; Gan, L.; Shen, Y.; Zou, Y. A Bibliometric Analysis of the Literature on Irisin from 2012–2021. Int. J. Environ. Res. Public Health 2022, 19, 6153. [Google Scholar] [CrossRef] [PubMed]
- Basini, G.; Bussolati, S.; Iannarelli, M.; Ragionieri, L.; Grolli, S.; Ramoni, R.; Dodi, A.; Gazza, F.; Grasselli, F. The myokine irisin: Localization and effects in swine late medium and large antral ovarian follicle. Domest. Anim. Endocrinol. 2021, 74, 106576. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Chen, L.; Lu, X.; Tan, A.; Chen, Y.; Li, Y.; Peng, X.; Yuan, S.; Cai, D.; Yu, Y. Peri-ovarian adipose tissue contributes to intraovarian control during folliculogenesis in mice. Reproduction 2018, 156, 133–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waseem, R.; Shamsi, A.; Mohammad, T.; Hassan, M.I.; Kazim, S.N.; Chaudhary, A.A.; Rudayni, H.A.; Al-Zharani, M.; Ahmad, F.; Islam, A. FNDC5/Irisin: Physiology and Pathophysiology. Molecules 2022, 27, 1118. [Google Scholar] [CrossRef] [PubMed]
- Hepler, C.; Vishvanath, L.; Gupta, R.K. Sorting out adipocyte precursors and their role in physiology and disease. Genes Dev. 2017, 31, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Hepler, C.; Gupta, R.K. The expanding problem of adipose depot remodelling and postnatal adipocyte progenitor recruitment. Mol. Cell. Endocrinol. 2017, 445, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Scherer, P.E. The many secret lives of adipocytes: Implications for diabetes. Diabetes 2019, 62, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berni, M.; Gigante, P.; Bussolati, S.; Grasselli, F.; Grolli, S.; Ramoni, R.; Basini, G. Bisphenol S, a Bisphenol A alternative, impairs swine ovarian and adipose cell functions. Domest. Anim. Endocrinol. 2019, 66, 48–56. [Google Scholar] [CrossRef]
- Basini, G.; Bussolati, S.; Grolli, S.; Ragionieri, L.; Di Lecce, R.; Grasselli, F. Orexin B inhibits viability and differentiation of stromal cells from swine adipose tissue. Domest. Anim. Endocrinol. 2021, 75, 106594. [Google Scholar] [CrossRef] [PubMed]
- Priya, N.; Sarcar, S.; Sen Majumdar, A.; SundarRaj, S. Explant culture: A simple, reproducible, efficient and economic technique for isolation of mesenchymal stromal cells from human adipose tissue and lipoaspirate. J. Tissue Eng. Reg. Med. 2014, 8, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Basini, G.; Bussolati, S.; Ciccimarra, R.; Grasselli, F. Melatonin potentially acts directly on swine ovary by modulating granulosa cell function and angiogenesis. Reprod. Fertil. Dev. 2017, 29, 2305–2312. [Google Scholar] [CrossRef] [PubMed]
- Basini, G.; Bussolati, S.; Andriani, L.; Grolli, S.; Ramoni, R.; Bertini, S.; Iemmi, T.; Menozzi, A.; Berni, P.; Grasselli, F. Nanoplastics impair in vitro swine granulosa cell functions. Domest. Anim. Endocrinol. 2021, 76, 106611. [Google Scholar] [CrossRef]
- Tan, A.S.; Berridge, M.V. Superoxide produced by activated neutrophils efficiently reduces the tetrazolium salt, WST-1 to produce a soluble formazan: A simple colorimetric assay for measuring respiratory burst activation and for screening anti-inflammatory agents. J. Immunol. Methods 2000, 238, 59–68. [Google Scholar] [CrossRef]
- Xu, C.; Liu, S.; Liu, Z.; Song, F.; Liu, S. Superoxide generated by pyrogallol reduces highly water-soluble tetrazolium salt to produce a soluble formazan: A simple assay for measuring superoxide anion radical scavenging activities of biological and abiological samples. Anal. Chim. Acta 2013, 793, 53–60. [Google Scholar] [CrossRef]
- Basini, G.; Bussolati, S.; Baioni, L.; Grasselli, F. Gossypol, a polyphenolic aldehyde from cotton plant, interferes with swine granulosa cell function. Domest. Anim. Endocrinol. 2009, 37, 30–36. [Google Scholar] [CrossRef]
- Basini, G.; Mainardi, G.L.; Bussolati, S.; Tamanini, C. Steroidogenesis, proliferation and apoptosis in bovine granulosa cells: Role of tumour necrosis factor-alpha and its possible signalling mechanisms. Reprod. Fertil. Dev. 2002, 14, 141–150. [Google Scholar] [CrossRef]
- Bianco, F.; Basini, G.; Grasselli, F. The plant alkaloid Sanguinarine affects swine granulosa cell activity. Reprod. Toxicol. 2006, 21, 335–340. [Google Scholar] [CrossRef]
- Basini, G.; Santini, S.E.; Bussolati, S.; Grasselli, F. The phytoestrogen quercetin impairs steroidogenesis and angiogenesis in swine granulosa cells in vitro. J. Biomed. Biotechnol. 2009, 2009, 419891. [Google Scholar]
- Sengupta, P.; Dutta, S.; Karkada, I.R.; Akhigbe, R.E.; Chinni, S.V. Irisin, Energy Homeostasis and Male Reproduction. Front. Physiol. 2021, 12, 746049. [Google Scholar] [CrossRef] [PubMed]
- Colaianni, G.; Sanesi, L.; Storlino, G.; Brunetti, G.; Colucci, S.; Grano, M. Irisin and Bone: From Preclinical Studies to the Evaluation of Its Circulating Levels in Different Populations of Human Subjects. Cells 2019, 8, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colaianni, G.; Cuscito, C.; Mongelli, T.; Oranger, A.; Mori, G.; Brunetti, G.; Colucci, S.; Cinti, S.; Grano, M. Irisin enhances osteoblast differentiation in vitro. Int. J. Endocrinol. 2014, 2014, 902186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colaianni, G.; Errede, M.; Sanesi, L.; Notarnicola, A.; Celi, M.; Zerlotin, R.; Storlino, G.; Pignataro, P.; Oranger, A.; Pesce, V.; et al. In Reply to the Letter to the Editor: Involvement of Irisin in Age-Related Osteoporosis and Its Inhibitory Effect on the Senescent Marker p21 in Osteoblasts. J. Bone Miner. Res. 2021, 36, 1420–1421. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Song, F.; Xu, K.; Liu, Z.; Han, S.; Li, F.; Sun, Y. Irisin Attenuates Neuroinflammation and Prevents the Memory and Cognitive Deterioration in Streptozotocin-Induced Diabetic Mice. Mediat. Inflamm. 2019, 2019, 1567179. [Google Scholar] [CrossRef] [Green Version]
- Arhire, L.; Mihalache, L.; Covasa, M. Irisin: A Hope in Understanding and Managing Obesity and Metabolic Syndrome. Front. Endocrinol. 2019, 10, 524. [Google Scholar] [CrossRef] [Green Version]
- Merrick, D.; Sakers, A.; Irgebay, Z.; Okada, C.; Calvert, C.; Morley, M.P.; Percec, I.; Seale, P. Identification of a mesenchymal progenitor cell hierarchy in adipose tissue. Science 2019, 364, eaav2501. [Google Scholar] [CrossRef]
- Xue, C.; Li, X.; Ba, L.; Shen, Y.; Sun, Z.; Gu, J.; Yang, Y.; Han, Q.; Zhao, R.C. Irisin mediates beiging of adipose-derived mesenchymal stem cells through binding to TRPC3. BMC Biol. 2022, 20, 95. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Jian, Z.; Wang, H.; Xu, L.; Zhang, T.; Song, J. Irisin Promotes Osteogenesis by Modulating Oxidative Stress and Mitophagy through SIRT3 Signaling under Diabetic Conditions. Oxid. Med. Cell. Longev. 2022, 10, 3319056. [Google Scholar] [CrossRef]
- Moreno-Navarrete, J.M.; Ortega, F.; Serrano, M.; Guerra, E.; Pardo, G.; Tinahones, F.; Ricart, W.; Fernández-Real, J.M. Irisin Is Expressed and Produced by Human Muscle and Adipose Tissue in Association with Obesity and Insulin Resistance. J. Clin. Endocr. Metab. 2013, 98, E769–E778. [Google Scholar] [CrossRef]
- Varela-Rodríguez, B.M.; Pena-Bello, L.; Juiz-Valiña, P.; Vidal-Bretal, B.; Cordido, F.; Sangiao-Alvarellos, S. FNDC5 expression and circulating irisin levels are modified by diet and hormonal conditions in hypothalamus, adipose tissue and muscle. Sci. Rep. 2016, 6, 29898. [Google Scholar] [CrossRef] [PubMed]
- Komolka, K.; Albrecht, E.; Schering, L.; Brenmoehl, J.; Hoeflich, A.; Maak, S. Locus characterization and gene expression of bovine FNDC5: Is the myokine irisin relevant in cattle? PLoS ONE 2014, 9, e88060. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Zhang, Y.; Tran, T.D.N.; Wang, H.; Li, S.; George, E.V.; Zhuang, H.; Zhang, P.; Kandel, A.; Lai, Y.; et al. Irisin Controls Growth, Intracellular Ca2+ Signals, and Mitochondrial Thermogenesis in Cardiomyoblasts. PLoS ONE 2015, 10, e0136816. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Song, N.; Huang, Y.; Chen, Y. Irisin inhibits pancreatic cancer cell growth via the AMPK-mTOR pathway. Sci. Rep. 2018, 8, 15247. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Song, H.; Zhang, Y.; Wu, F.; Mu, Q.; Jiang, M.; Wang, F.; Zhang, W.; Li, L.; Shao, L.; et al. Irisin Inhibits Atherosclerosis by Promoting Endothelial Proliferation through microRNA126-5p. J. Am. Heart Assoc. 2016, 5, e004031. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Du, F.; Li, X.; Wang, M.; Duan, R.; Zhang, J.; Wu, Y.; Zhang, Q. Effects and underlying mechanisms of irisin on the proliferation and apoptosis of pancreatic β cells. PLoS ONE 2017, 12, e0175498. [Google Scholar] [CrossRef] [Green Version]
- Mazur-Bialy, A.; Bilski, J.; Pochec, E.; Brzozowski, T. New insight into the direct anti-inflammatory activity of a myokine irisin against proinflammatory activation of adipocytes. Implication for exercise in obesity. J. Physiol. Pharmacol. 2017, 68, 243–251. [Google Scholar]
- Jiang, X.; Cai, S.; Jin, Y.; Wu, F.; He, J.; Wu, X.; Tan, Y.; Wang, Y. Irisin Attenuates Oxidative Stress, Mitochondrial Dysfunction, and Apoptosis in the H9C2 Cellular Model of Septic Cardiomyopathy through Augmenting Fundc1-Dependent Mitophagy. Oxid. Med. Cell. Longev. 2021, 18, 2989974. [Google Scholar] [CrossRef]
- Peng, H.; Wang, Q.; Lou, T.; Qin, J.; Jung, S.; Shetty, V.; Li, F.; Wang, Y.; Feng, X.H.; Mitch, W.E.; et al. Myokine mediated muscle-kidney crosstalk suppresses metabolic reprogramming and fibrosis in damaged kidneys. Nat. Commun. 2017, 8, 1493. [Google Scholar] [CrossRef] [Green Version]
- Kuloglu, T.; Aydin, S.; Eren, M.; Yilmaz, M.; Sahin, I.; Kalayci, M.; Sarman, E.; Kaya, N.; Yilmaz, O.; Turk, A.; et al. Irisin: A potentially candidate marker for myocardial infarction. Peptides 2014, 55, 85–91. [Google Scholar] [CrossRef]
- Fu, J.; Han, Y.; Wang, J.; Liu, Y.; Zheng, S.; Zhou, L.; Jose, P.A.; Zeng, C. Irisin Lowers Blood Pressure by Improvement of Endothelial Dysfunction via AMPK-Akt-eNOS-NO Pathway in the Spontaneously Hypertensive Rat. J. Am. Heart Assoc. 2016, 5, e003433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, X.; Huang, W.; Peng, J.; Zhu, T.T.; Sun, X.L.; Zhou, X.Y.; Yang, H.; Xiong, J.F.; He, H.Q.; Xu, Y.H.; et al. Irisin Alleviates Advanced Glycation End Products-Induced Inflammation and Endothelial Dysfunction via Inhibiting ROS-NLRP3 Inflammasome Signaling. Inflammation 2018, 41, 260–275. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhao, Y.T.; Zhang, S.; Dubielecka, P.M.; Du, J.; Yano, N.; Chin, Y.E.; Zhuang, S.; Qin, G.; Zhao, T.C. Irisin plays a pivotal role to protect the heart against ischemia and reperfusion injury. J. Cell. Physiol. 2017, 232, 3775–3785. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Han, T.; Gao, L.; Wang, L.; Wang, S.; Yang, L.; Zhang, J.; Guan, Y.; Yan, N.; Yu, H.; et al. Effect of aerobic exercise and resistance exercise in improving non-alcoholic fatty liver disease: A randomized controlled trial. Chin. J. Hepatol. 2018, 26, 34–41. [Google Scholar]
- Kawanami, H.; Nomura, S.; Sakurai, T.; Sakurai, T.; Yamagishi, H.; Komabayashi, T.; Izawa, T. Possible role of nitric oxide on adipocyte lipolysis in exercisetrained rats. Jpn. J. Physiol. 2002, 52, 343–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathew, H.; Castracane, V.D.; Mantzoros, C. Adipose tissue and reproductive health. Metabolism 2018, 86, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, Y.; Wang, F.; Donelan, W.; Christine Zona, M.; Li, S.; Reeves, W.; Ding, Y.; Tang, D.; Yang, L. Effects of irisin on the differentiation and browning of human visceral white adipocytes. Am. J. Transl. Res. 2019, 11, 7410–7421. [Google Scholar]
- Zhang, Y.; Xie, C.; Wang, H.; Foss, R.; Clare, M.; George, E.; Li, S.; Katz, A.; Cheng, H.; Ding, Y.; et al. Irisin exerts dual effects on browning and adipogenesis of human white adipocytes. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E530–E541. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Qiao, X.; Ma, Y.; Deng, H.; Xu, C.; Xu, L. Disordered metabolism in mice lacking irisin. Sci. Rep. 2020, 10, 17368. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Primer Sequences | Reference |
|---|---|---|---|
| FNDC5 | XM_02109583.1 | F: 5′-TGCAGGCCATCTCCATTCAG-3′ R: 5′-ATATTGGCGGCAGAAGAGGG-3′ | [4] |
| Actin | XM_003124280 | F: 5′-ATGGATGACGATATTGCTGC-3′ R: 5′-CCCACGTAGGAGTCCTTCTG-3′ | [4] |
| PPARγ | XM_013981982.1 | F:5′-GCCCTTCACCACTGTTGATT-3′ R:5′GAGTTGGAAGGCTCTTCGTG-3′ | [4] |
| Leptin | NM_213840.1 | F:5′-TGGCCCTATCTGTCCTACG-3′ R:5′-TTTCTGGAAGGCAGACTGGT-3′ | [10] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basini, G.; Bussolati, S.; Grolli, S.; Berni, P.; Di Lecce, R.; Grasselli, F. Effects of the Myokine Irisin on Stromal Cells from Swine Adipose Tissue. Biomolecules 2022, 12, 1895. https://doi.org/10.3390/biom12121895
Basini G, Bussolati S, Grolli S, Berni P, Di Lecce R, Grasselli F. Effects of the Myokine Irisin on Stromal Cells from Swine Adipose Tissue. Biomolecules. 2022; 12(12):1895. https://doi.org/10.3390/biom12121895
Chicago/Turabian StyleBasini, Giuseppina, Simona Bussolati, Stefano Grolli, Priscilla Berni, Rosanna Di Lecce, and Francesca Grasselli. 2022. "Effects of the Myokine Irisin on Stromal Cells from Swine Adipose Tissue" Biomolecules 12, no. 12: 1895. https://doi.org/10.3390/biom12121895