
Modulation of Canine Melanocortin-3 and -4 Receptors by Melanocortin-2 Receptor Accessory Protein 1 and 2


Department of Anatomy, Physiology, and Pharmacology, College of Veterinary Medicine, Auburn University, Auburn, AL 36849, USA
*
Author to whom correspondence should be addressed.
Biomolecules 2022, 12(11), 1608; https://doi.org/10.3390/biom12111608
Submission received: 12 October 2022
/
Revised: 26 October 2022
/
Accepted: 29 October 2022
/
Published: 1 November 2022
(This article belongs to the Collection Feature Papers in Molecular Genetics)
Abstract
:The neural melanocortin receptors (MCRs), melanocortin-3 and -4 receptors (MC3R and MC4R), have crucial roles in regulating energy homeostasis. The melanocortin-2 receptor accessory proteins (MRAPs, MRAP1 and MRAP2) have been shown to regulate neural MCRs in a species-specific manner. The potential effects of MRAP1 and MRAP2 on canine neural MCRs have not been investigated before. Herein, we cloned canine (c) MC3R and identified one canine MRAP2 splice variant, MRAP2b, with N-terminal extension of cMRAP2a. Canine MC3R showed higher maximal responses to five agonists than those of human MC3R. We further investigated the modulation of cMRAP1, cMRAP2a, and cMRAP2b, on cMC3R and cMC4R pharmacology. For the cMC3R, all MRAPs had no effect on trafficking; cMRAP1 significantly decreased Bmax whereas cMRAP2a and cMRAP2b significantly increased Bmax. Both MRAP1 and MRAP2a decreased Rmaxs in response to α-MSH and ACTH; MRAP2b only decreased α-MSH-stimulated cAMP generation. For the MC4R, MRAP1 and MRAP2a increased cell surface expression, and MRAP1 and MRAP2a increased Bmaxs. All MRAPs had increased affinities to α-MSH and ACTH. MRAP2a increased ACTH-induced cAMP levels, whereas MRAP2b decreased α-MSH- and ACTH-stimulated cAMP production. These findings may lead to a better understanding of the regulation of neural MCRs by MRAP1 and MRAP2s.
1. Introduction
Melanocortin-3 and -4 receptors (MC3R and MC4R), also called neural MCRs, are two melanocortin receptors (MCRs) that are highly expressed in the central nervous system [1,2,3,4], and are essential for regulating energy homeostasis [5,6,7]. Mice lacking Mc3r have a moderate obesity phenotype with normal food intake and metabolism, decreased lean mass and increased fat mass [8,9,10]. Mc4r knockout mice have increased food intake, decreased energy expenditure, and morbid obesity [11,12]. These findings suggest a distinct nonredundant regulation of the energy balance by MC3R and MC4R. Additionally, mutations in MC3R and MC4R are associated with obesity [7,13,14,15,16]. MC3R was shown to be involved in other physiological functions, including the modulation of natriuresis [17], cardiovascular function [18,19], immune response [20,21,22,23,24], and timing of sexual maturation [25]. MC4R also has crucial roles in sexual function and reproduction [7,15].
MC3R and MC4R are activated by the endogenous agonists, including α-, β-, γ-melanocyte-stimulating hormones (MSHs) and adrenocorticotropin (ACTH) [26,27]. As members of the family of G protein-coupled receptors (GPCRs), the neural MCRs primarily couple to the stimulatory G protein (Gαs) to stimulate adenylyl cyclase activity, resulting in increased generation of the intracellular second messenger cyclic adenosine monophosphate (cAMP) to trigger downstream signaling.
MCRs have been shown to interact with small single transmembrane proteins, melanocortin-2 receptor accessory proteins (MRAPs, including MRAP1 and MRAP2) [28,29,30,31] (reviewed in [32,33]). MRAP1 was first identified as the specific chaperone for MC2R, essential for MC2R forward trafficking [28,29,33,34]. Human (h) MRAP1 mutations account for ~20% of familial glucocorticoid deficiency cases [28,35]. Subsequent studies suggest that MRAP1 possesses functions beyond regulating MC2R. Indeed, hMRAP1 has been shown to modulate five human MCRs in distinct ways [30,36,37]. MRAP1 regulates MC3R and MC4R in chicken and frog [25,38,39]. So far, there are only a few studies focusing on MRAP1 regulation of neural MCRs [25,30,37,38,39,40,41,42,43].
MRAP2, a paralog of MRAP1, with high expression in the brain, has essential roles in regulating energy homeostasis. Mice lacking Mrap2 show early onset severe obesity [44,45]. Human MRAP2 mutations are also associated with severe obesity [44,46,47,48]. MRAP2 has been shown to modulate MC3R/MC4R trafficking and signaling in vertebrates [30,31,38,43,44,49,50,51,52,53]. Human MRAP2 has three alternatively spliced variants with different C-termini in the proteins, and these three variants exert different effects on MC3R/MC4R pharmacology [43].
From the studies on the regulation of MC3R and MC4R by MRAPs, it was shown that MRAPs modulate neural MCRs in a receptor- and species-dependent manner. The effects of MRAP1 and MRAP2 on canine (Canis lupus familiaris) (c) MC3R and cMC4R have not been studied before. Herein, we investigated the pharmacology of cMC3R as well as the pharmacological modulation of cMC3R and cMC4R by cMRAP1 and cMRAP2a/cMRAP2b. We have previously reported the pharmacology of cMC4R [54].
2. Materials and Methods
2.1. Ligands and Plasmids
[Nle4,D-Phe7]-α-MSH (NDP-MSH) and D-Trp8-γ-MSH were obtained from Vivitide (Louisville, KY, USA). Human α-MSH and β-MSH were purchased from Pi Proteomics (Huntsville, AL, USA). Human ACTH (1-24) was supplied by Phoenix Pharmaceuticals (Burlingame, CA, USA). Canine α-MSH, β-MSH, and ACTH (1-24), share the same sequences with the corresponding human counterparts. [125I]-cAMP and [125I]-NDP-MSH were iodinated using chloramine T method [55,56]. N-terminal myc-tagged hMC3R, cMC3R, and cMC4R [54]) and N-terminal Flag-tagged MRAPs (cMRAP1, cMRAP2a, and cMRAP2b) were commercially synthesized and subcloned into pcDNA3.1 by GenScript (Piscataway, NJ, USA).
2.2. Cell Culture and Transfection
Human embryonic kidney (HEK) 293T cells (ATCC, Manassas, VA, USA) were cultured at 37 °C in a 5% CO2-humidified incubator [57]. Cells were plated into gelatin-coated 6- or 24-well plates. Cells were co-transfected with 0.25 μg/μL cMC3R or cMC4R with or without MRAP plasmids using calcium phosphate precipitation method [58].
2.3. Ligand Binding Assays
Binding assay was performed as described previously [49,57]. The ligands and their final concentrations used in this study were NDP-MSH (from 10−12 to 10−6 M), α-MSH (from 10−11 to 10−5 M), β-MSH (from 10−11 to 10−5 M), ACTH (1-24) (from 10−12 to 10−6 M), and D-Trp8-γ-MSH (from 10−12 to 10−6 M). To investigate the regulation of MRAPs on the binding properties of cMC3R and cMC4R, cMC3R or cMC4R (0.25 μg/μL) and cMRAP1, cMRAP2a or cMRAP2b plasmids in two ratios (1:0 and 1:5) were co-transfected into 6-well plates, and two ligands, α-MSH and ACTH (1-24), were used for binding and signaling assays.
2.4. Ligand-Stimulated cAMP Assays
Radioimmunoassay (RIA) was used to determine intracellular cAMP levels as described previously [55,57]. Five ligands, including NDP-MSH, α-MSH, β-MSH, ACTH (1-24), and D-Trp8-γ-MSH, were used. To investigate effects of MRAPs on cMC3R or cMC4R signaling, cells were co-transfected with cMC3R or cMC4R (0.25 μg/μL) and cMRAP plasmids (1:5), and two ligands, α-MSH and ACTH (1-24), were used.
2.5. Flow Cytometry Assay
The regulation of cMRAP1 or cMRAP2s on the expression of cMC3R and cMC4R was carried out using flow cytometry (Accuri Cytometers, Ann Arbor, MI, USA) as described previously [59,60]. Cells were co-transfected with cMC3R or cMC4R and cMRAP1, cMRAP2a, or cMRAP2b plasmids (1:5). Fluorescence of cells transfected with pcDNA3.1 was used for background staining. The expression of receptors was calculated as the percentage of the cell transfected with receptors in absence of MRAPs (set as 100%) [59].
2.6. Statistical Analysis
All data were presented as mean ± S.E.M. GraphPad Prism 8.3 software (GraphPad, San Diego, CA, USA) was used to calculate the parameters of ligand binding, cAMP signaling, and flow cytometry assay. The significance in binding and signaling parameters between cMC3R and hMC3R, as well as vehicle and ligand-treated groups, were all determined by Student’s t-test. One-way ANOVA was used to analyze the significant differences in binding, cAMP, and flow cytometry between multiple groups.
3. Results
3.1. Nucleotide and Deduced Amino Acid Sequences of cMC3R, cMRAP1, and cMRAP2s
The canine MC3R (GenBank: NM_001135124.1) had 972 bp open reading frame (ORF), encoding a putative protein of 323 amino acids with 35.79 kDa molecular mass (Figure 1A). cMC3R had seven hydrophobic transmembrane domains (TMDs). Several conserved motifs, including PMY, DRY, and DPxxY, and three potential N-linked glycosylation sites (Asn2, Asn16, and Asn28) in N-terminus, were present at homologous positions with MC3Rs of other species (Figure 1). Canine MC3R shared high identities with other MC3R orthologs, 96% to giant panda, 92% to human, 90% to cat, 89% to pig, 87% to mouse, 76% to chicken, 76% to turtle, 75% to frog, and 72% to zebrafish. The phylogenetic tree showed that cMC3R nested with mammalian MC3Rs (Figure 1B).
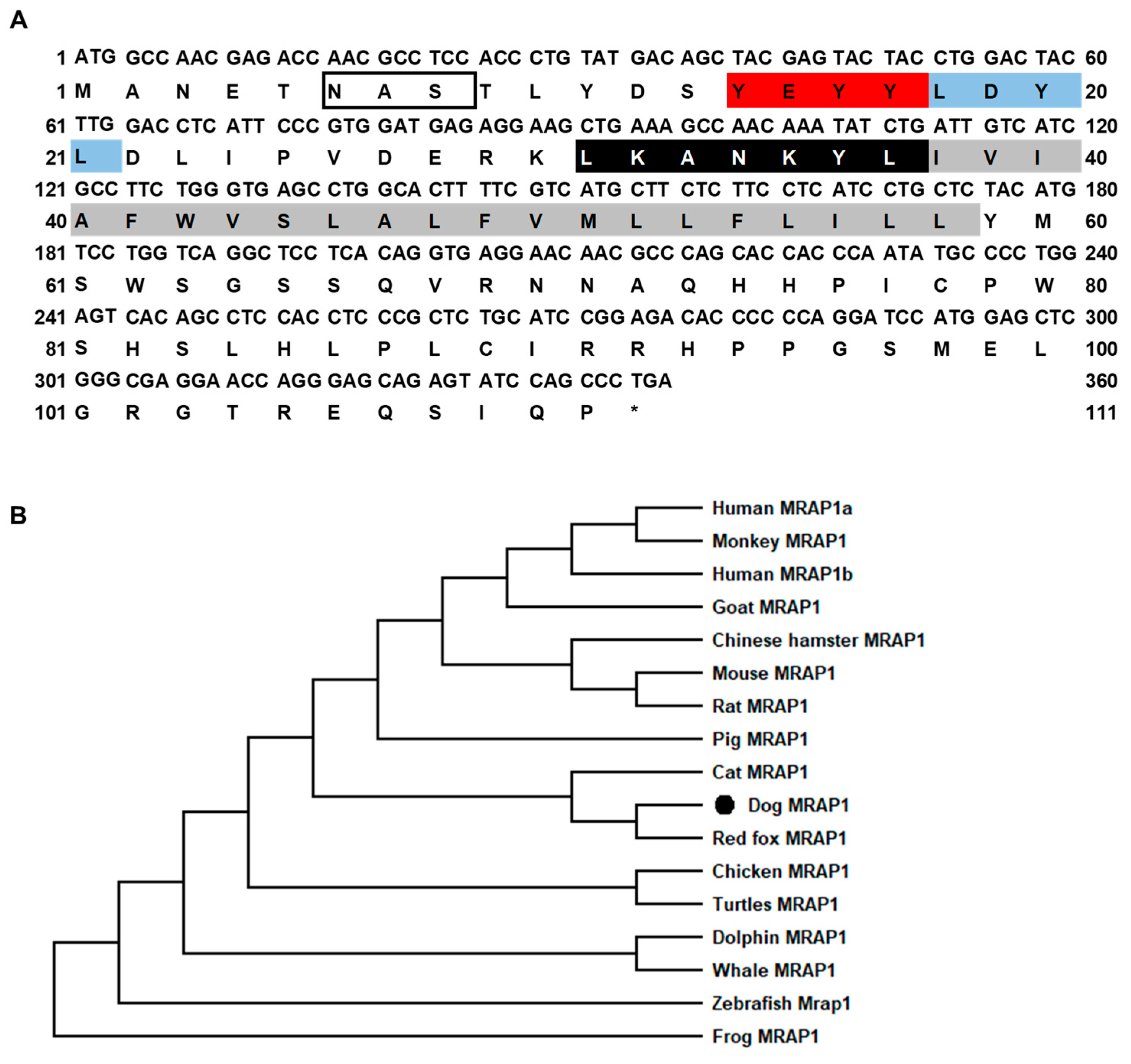
The canine MRAP1 had 336 bp ORF that encoded a putative protein of 111 amino acids with 12.90 kDa molecular mass (Figure 2A). cMRAP1 had the classical characteristic of other MRAP orthologs, including two potential N-linked glycosylation sites (Asn3 and Asn6), YEYY motif, LDYL motif, LKANKYL motif, and a single TMD (Figure 2A). cMRAP1 shared high identities with cat MRAP1 (90%), and lower identities with other MRAP1 (44–79%). The phylogenetic tree showed that cMRAP1 clustered with mammalian MRAP1s and was evolutionarily closer to red fox MRAP1 (Figure 2B).
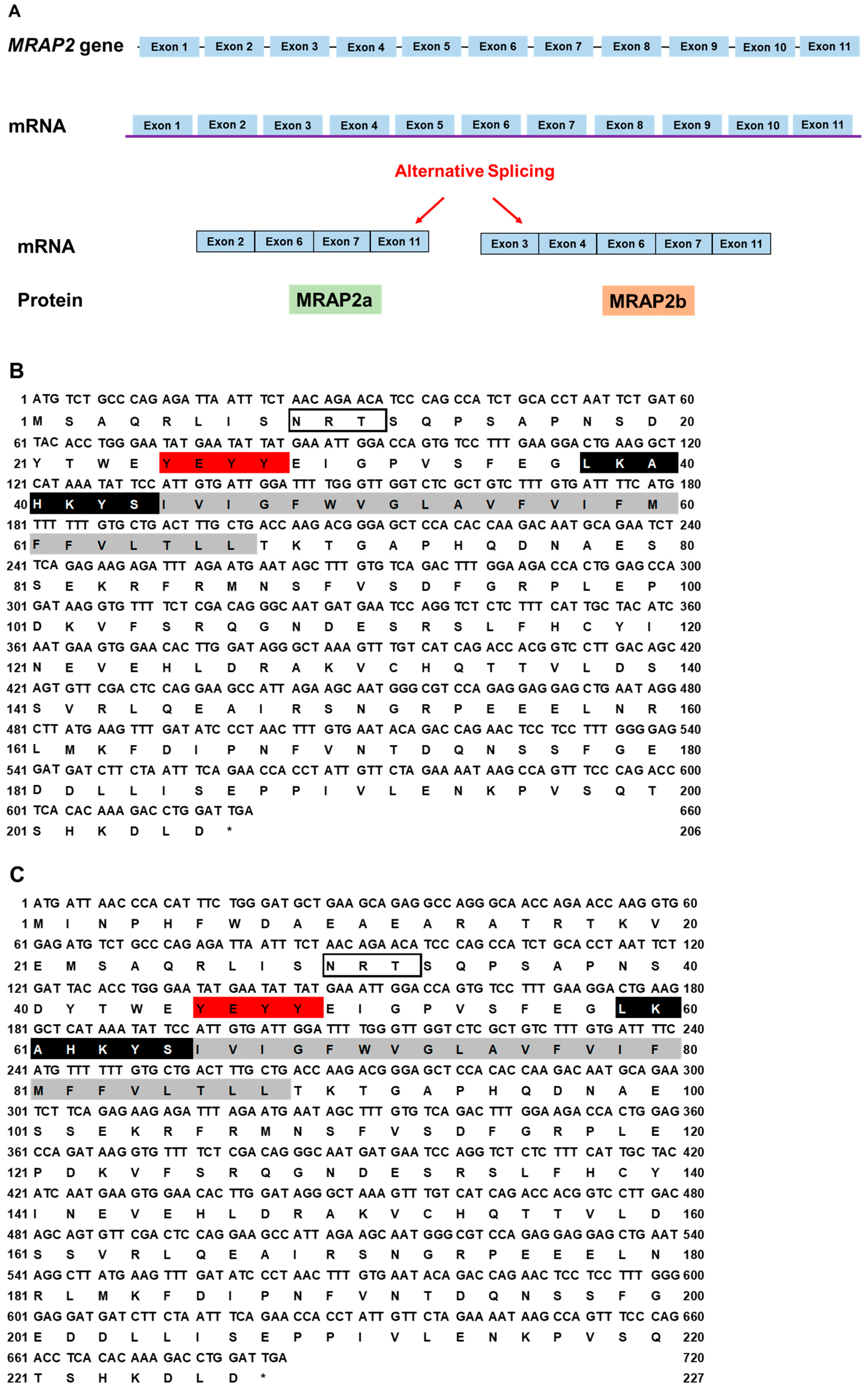
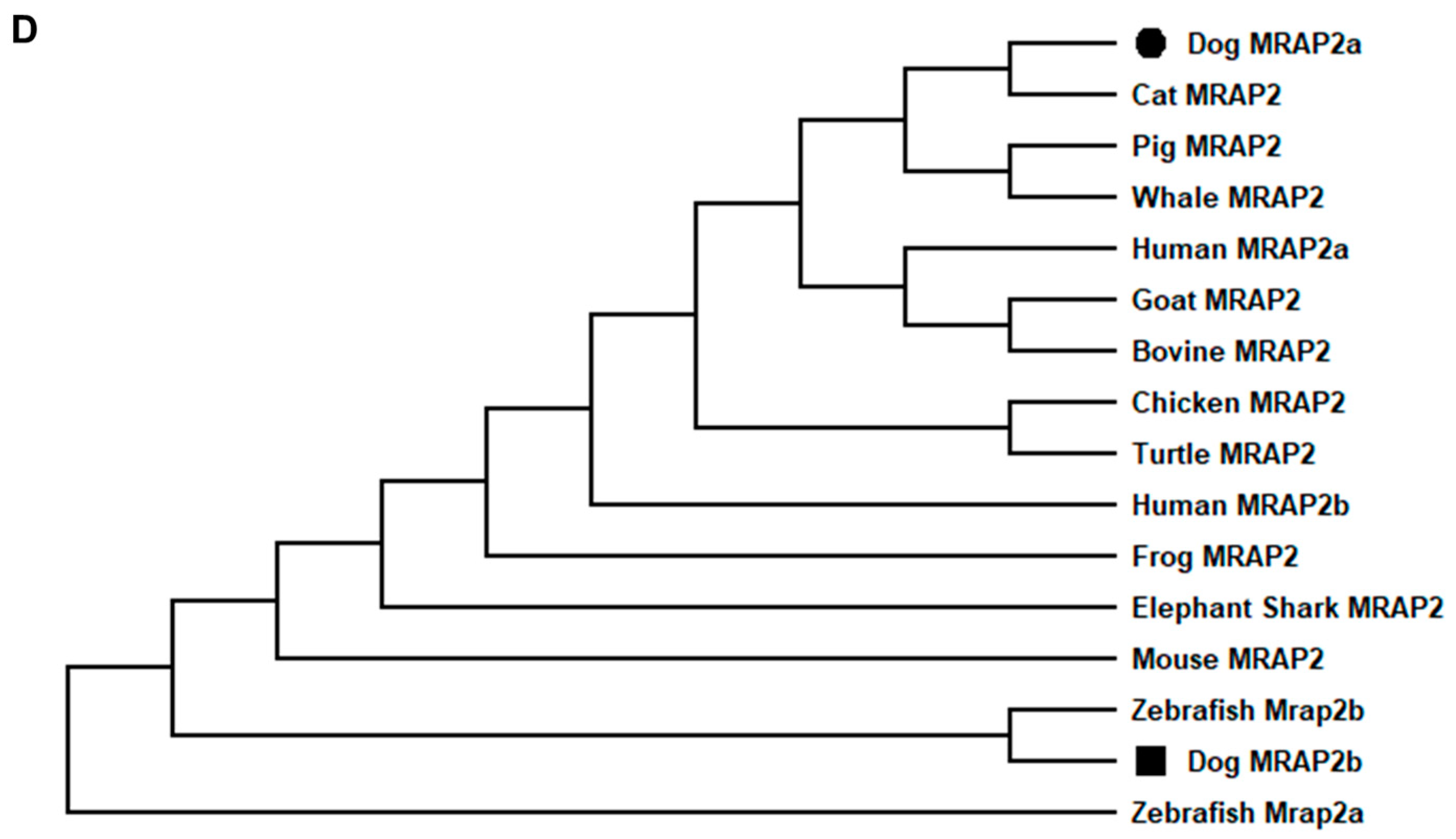
The canine MRAP2 consists of 11 exons. Two MRAP2 splice variants were identified: MRAP2a (XM_038682814.1) derived from four exons (2, 6, 7, and 11) that had 621 bp ORF, encoding a putative protein of 206 amino acids with 23.62 kDa molecular mass (Figure 3A); MRAP2b (XM_038682813.1) derived from five exons (3, 4, 6, 7, and 11) that had 684 bp ORF encoding a putative protein of 232 amino acids with 26.71 kDa molecular mass (Figure 3A). Canine MRAP2b had an extended N-terminus (26 amino acids) compared with cMRAP2a, and they shared the common structure with other MRAP2s, such as one potential N-linked glycosylation sites (Asn9 in MRAP2a and Asn35 in MRAP2b), YEYY motif, LKAHKYS motif, and a single TMD (Figure 3B,C). Multiple sequence alignment analysis showed that dog MRAP2a and MRAP2b shared high identities with mammalian MRAP2s (>81%) and lower identities with MRAP2s from other species (<78%). The two isoforms of cMRAP2 were clustered with different MRAP2s, in which MRAP2a was evolutionarily closer to cat MRAP2, and MRAP2b was nested with Nile tilapia MRAP2 (Figure 3D).
3.2. Ligand Binding Properties of cMC3R
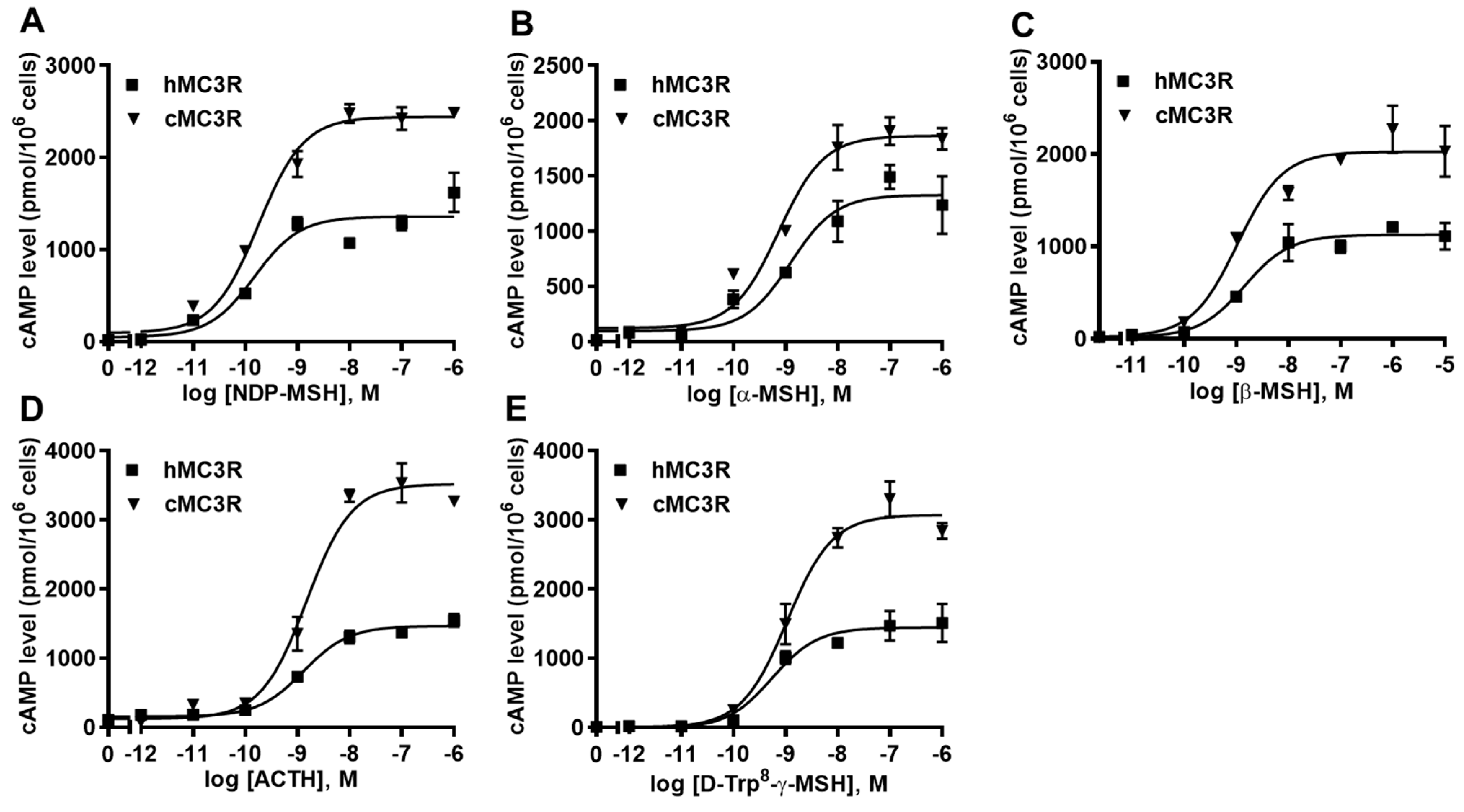
The binding assay was performed using multiple MC3R ligands, including NDP-MSH, α-MSH, β-MSH, ACTH, and D-Trp8-γ-MSH. We included hMC3R for comparison in the same experiments to explore whether cMC3R shows any unique pharmacological characteristics. The maximal binding value (Bmax) of cMC3R was 247.97 ± 13.44% of that of hMC3R (Figure 4 and Table 1). Canine MC3R had significantly lower affinities to NDP-MSH and D-Trp8-γ-MSH than that of hMC3R (Figure 4 and Table 1). The two MC3Rs showed similar IC50s when α-MSH, β-MSH, and ACTH, were used (Figure 4 and Table 1).
3.3. cAMP Signaling Properties of cMC3R
Intracellular cAMP levels were determined to explore whether cMC3R could respond to these agonists. All agonists could dose-dependently stimulate cMC3R and increase cAMP production (Figure 5 and Table 2). Canine MC3R showed higher maximal responses (Rmax) in response to four agonists (NDP-MSH, β-MSH, ACTH, and D-Trp8-γ-MSH) compared to hMC3R (Figure 5 and Table 2). Similar EC50s between the two MC3Rs were observed in response to five agonists (Figure 5 and Table 2). Additionally, cMC3R showed similar basal activity as hMC3R (Table 2).
3.4. Modulation of cMC3R and cMC4R Expression by MRAPs
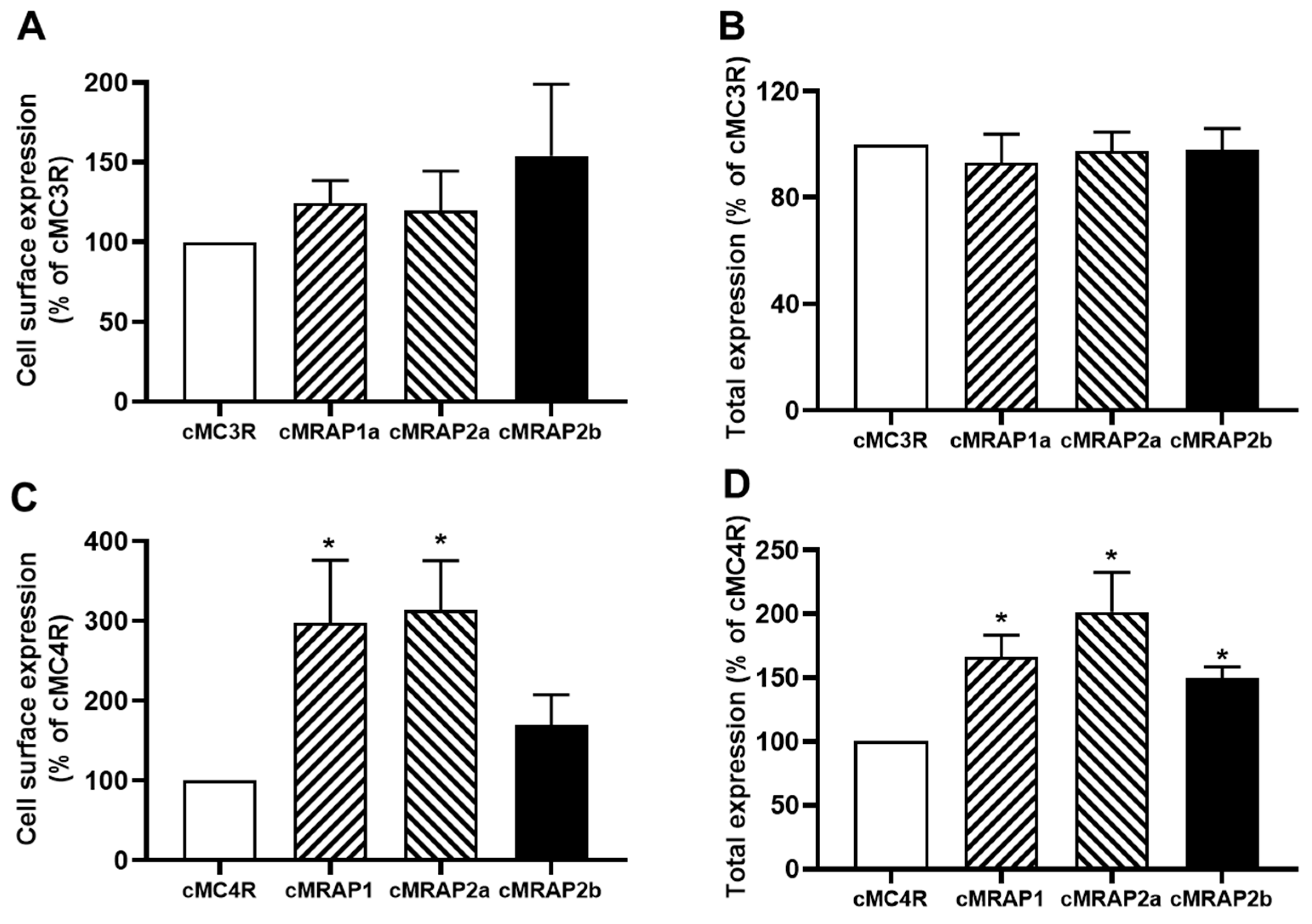
Canine MC3R or MC4R expression regulated by MRAPs was measured using flow cytometry. Results showed that cMRAP1, cMRAP2a, and cMRAP2b had no effect on the cell surface and total expression of cMC3R (Figure 6A,B). For cMC4R, cMRAP1 and cMRAP2a increased cell surface and total expression, and cMMRAP2b only increased total expression of cMC4R (Figure 6C,D).
3.5. Modulation of cMC3R Pharmacology by MRAPs
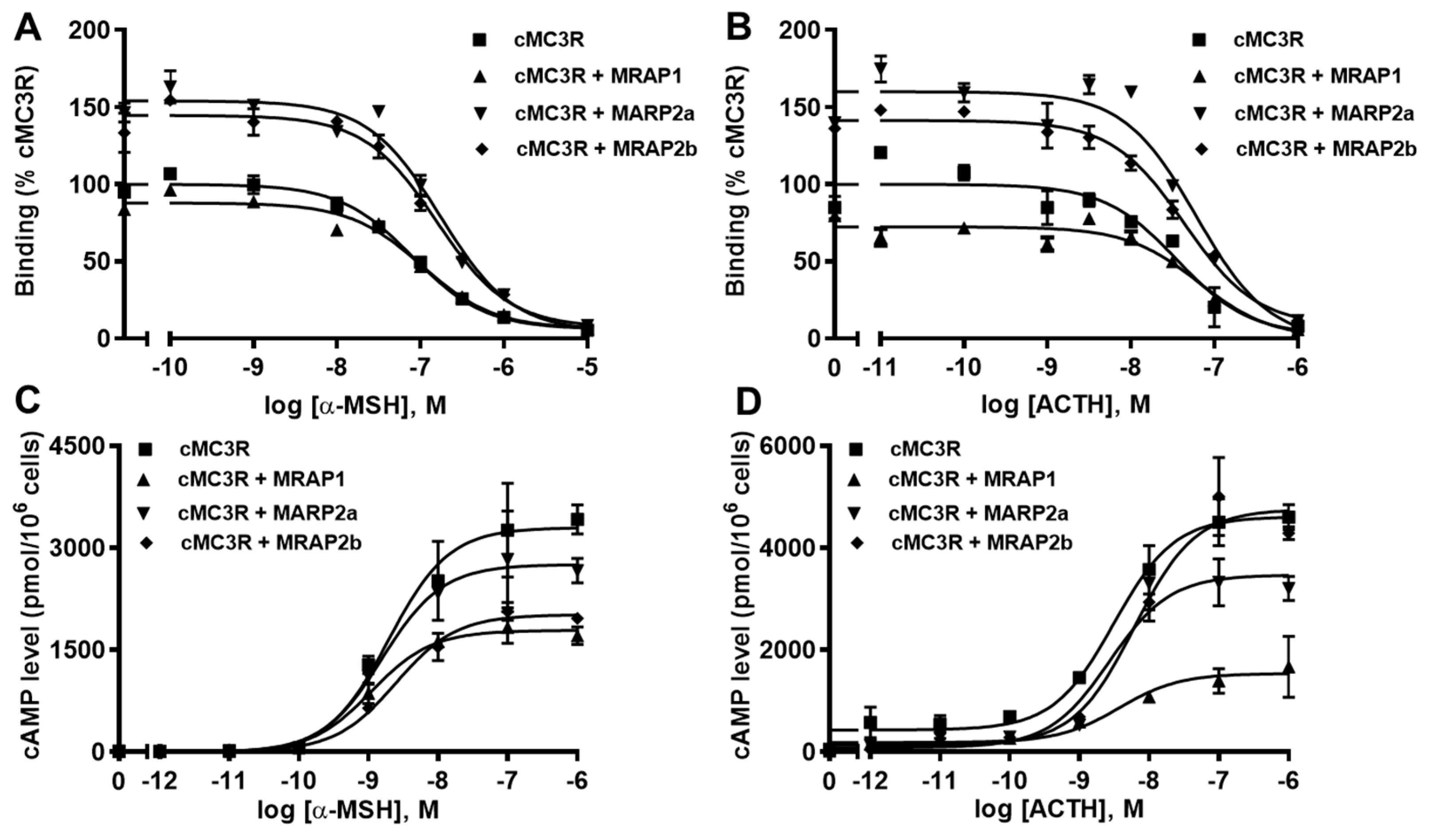
Ligand binding assays with α-MSH and ACTH showed that cMRAP1 decreased the Bmax, whereas cMRAP2a and cMRAP2b increased Bmaxs of cMC3R (Figure 7A,B and Table 3). All MRAPs had no significant effect on IC50s of cMC3R to α-MSH and ACTH (Figure 7A,B and Table 3). Modulation of cMRAPs on cMC3R signaling was also studied. Results showed that all MRAPs did not alter α-MSH and ACTH potencies of cMC3R (Table 4). Both MRAP1 and MRAP2a decreased Rmaxs in response to α-MSH and ACTH; MRAP2b significantly decreased α-MSH-stimulated cAMP generation but showed similar ACTH-induced cAMP production of cMC3R (Figure 7C,D and Table 4). Only MRAP2b decreased the basal activity, and the other MRAPs had no effect on the basal cAMP levels of cMC3R (Table 4).
3.6. Modulation of cMC4R Pharmacology by MRAPs
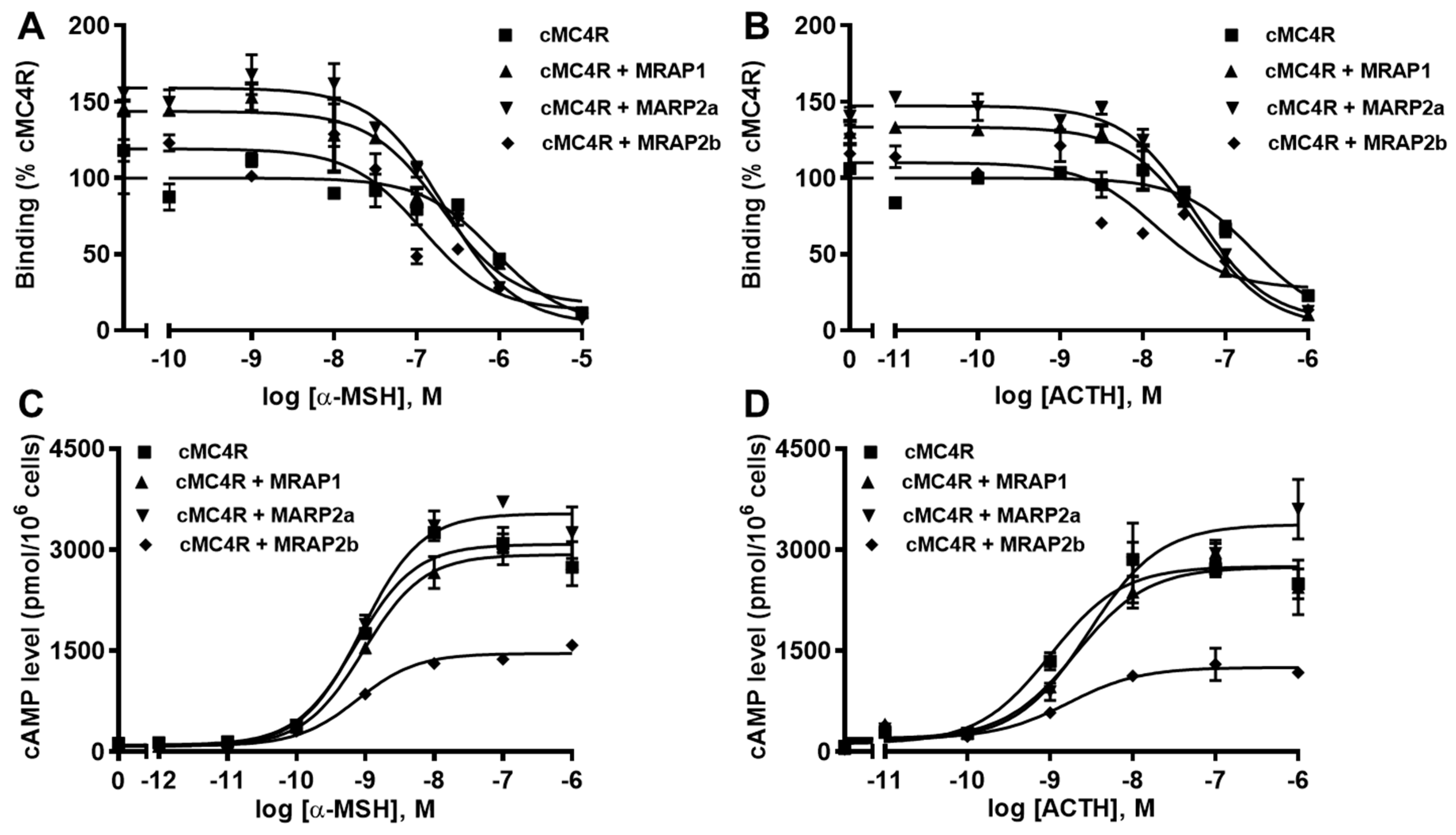
Ligand binding assays showed that MRAP1 and MRAP2a increased Bmaxs of cMC4R and MRAP2b did not affect the Bmax (Figure 8A,B and Table 5). All MRAPs increased the affinities of cMC4R to α-MSH and ACTH (Figure 8A,B and Table 5).
Signaling results showed that all MRAPs did not affect EC50s of cMC4R in response to α-MSH and ACTH (Table 6). MRAP1 had no effect on α-MSH- and ACTH-stimulated cAMP level of cMC4R (Figure 8C,D and Table 6). MRAP2a decreased α-MSH- and ACTH-induced cAMP signaling, whereas MRAP2b increased ACTH-stimulated signaling and did not affect α-MSH-stimulated cAMP signaling (Figure 8C,D and Table 6). Canine MC4R showed higher basal cAMP production than that of hMC4R (2.38 times that of hMC4R), indicating that cMC4R might be constitutively active. In this study, all MRAPs decreased the basal cAMP levels of cMC4R (Table 6).
4. Discussion
In this study, we cloned canine MC3R and investigated its pharmacological properties. We have reported cMC4R pharmacology previously [54,61]. In the current study, we also identified a MRAP2 variant, MRAP2b. The potential regulation of cMRAP1 and two cMRAP2 isoforms on cMC3R/cMC4R pharmacology were further studied.
To investigate the pharmacology of cMC3R, ligand binding and signaling assays were performed. cMC3Rs showed a higher binding capacity than that of hMC3R, consistent with the results of other MC3Rs, including channel catfish [49], topmouth culter [52], and giant panda [62]. For signaling, different from the results of giant panda and pig MC3Rs [62,63], cMC3R had a higher ligand-induced cAMP level than that of hMC3R. In addition, our current results are consistent with mammalian MC3Rs in that MC3R has little or no basal cAMP signaling [62,63,64,65]. Of interest, high constitutive activities were present in several non-mammalian MC3Rs, including teleosts [49,52,66], amphibians (Mexican axolotl) [67], and avian (chicken) [38,68]. The amino acids accounting for the differences in the constitutive activity between these MC3Rs are not clear. N-termini and extracellular loops are essential for the modulation of constitutive activities in hMC4R [69,70], luteinizing hormone receptor [71], and thyroid-stimulating hormone receptor [72,73]. Lower homology is observed in the N-termini and extracellular loops of mammalian and non-mammalian MC3Rs. Further studies are needed to determine the exact molecular determinants.
The potential roles of cMRAPs on cMC3R/cMC4R trafficking were studied. Human MRAP1a decreased [30,74] or increased [43], and MRAP2a decreased [30,74] or had no effect [43] on the cell surface expression of hMC3R. The current study showed that all cMRAPs did not affect the cell surface expression of cMC3R. In other species, frog MRAP1 increased and chicken MRAP1 did not alter the surface expression of MC3Rs [38,39]. MRAP2 decreased the surface expression of clawed frog MC3R [39], increased the surface expression of topmouth culter Mc3r [52], and had no effect on the surface expression of Mexican axolotl and chicken MC3Rs [38,67]. For MC4R, hMRAP1a and hMRAP2a decreased [30,74] or increased [43] the cell surface expression of hMC4R. In this study, cMRAP1 and cMRAP2a increased cell surface expression of cMC4R. MRAP1 had no effect on the surface expression of chicken MC3R [38] and increased frog MC3R expression [42]. MRAP2 has been reported to decrease the surface expression of tilapia and Mexican axolotl MC4Rs [67,75], increase the membrane expression of zebrafish (Mrap2b) [31], topmouth culter (Mrap2a and Mrap2b) [51], and Xenopus MC4Rs [39], or have no effect on the surface expression of chicken and snakehead MC4Rs [38,53]. Collectively, MRAP1 and MRAP2 modulate the MC3R/MC4R trafficking to the plasma membrane in a species- and receptor-specific manner.
Pharmacological studies were further performed on the potential MRAP modulation of cMC3R. Human MRAP1 and MRAP2 decreased α-MSH- and ACTH-induced [43] or increased α-MSH-stimulated [37,74] cAMP production of hMC3R. Xenopus MRAP1 increased α-MSH- and ACTH-induced cAMP signaling, and chicken MRAP1 did not affect agonist-induced signaling of MC3Rs [38,39]. Mrap2-decreased Mc3r signaling was also reported in channel catfish, topmouth culter (Mrap2a) [51], and Mexican axolotl [67], whereas MRAP2-increased MC3R signaling was observed in chicken and Xenopus MC3Rs [39]. Zebrafish Mrap2s did not affect agonist-induced signaling of MC3R [31]. Our results showed that MRAP1 did not alter MC3R trafficking, decreased Bmax, and α-MSH- and ACTH-induced cAMP levels, indicating that interaction between cMRAP1 and cMC3R might inhibit cMC3R bound to ligands, resulting in decreased signaling; MRAP2s did not change cMC3R trafficking, increased Bmaxs, but decreased signaling, probably due to the interaction of cMRAP2s and cMC3R leading to conformation change, further inhibiting G protein-induced intracellular cAMP signaling.
The potential modulation of cMRAPs on cMC4R pharmacology was also studied. Conflicting results were reported previously on hMRAP1a- and hMRAP2a-regulated signaling of hMC4R, where hMRAP1a decreased α-MSH-induced or did not affect α-MSH- and ACTH-stimulated hMC4R signaling [37,43]; MRAP2a had no effect [76], decreased [43] or increased [41,74] α-MSH-stimulated and did not affect ACTH-induced [43,76] signaling of hMC4R. Chicken MRAP1 was shown to decrease α-MSH-stimulated and have no effect on ACTH-induced signaling of MC4R [38]. MRAP1 increased α-MSH- and ACTH-stimulated signaling of Xenopus MC4R [39]. MRAP2-suppressed α-MSH- and/or ACTH-stimulated signaling of MC4Rs were present in several teleosts [31,51,53,75,77]. Our results indicated that MRAP1 had no effect on the efficacy of cMC4R in response to α-MSH and ACTH, MRAP2a increased ACTH-stimulated but had no effect on α-MSH-induced cMC4R signaling, and MRAP2b decreased α-MSH- and ACTH-induced signaling, suggesting that MRAP1 modulated cMC4R trafficking and ligand binding, but did not affect signaling; the interaction of MRAP2a and cMC4R might change MC4R ligand selectivity and sensitivity; MRAP2b might inhibit receptor coupling to G protein, resulting in decreased signaling. Hence, MRAPs might be involved in regulating receptor ligand selectivity and sensitivity in a species-dependent manner. The potential mechanisms of the MRAP regulation of receptor ligand selectivity and sensitivity need further study.
Human MC4R shows modest basal cAMP signaling [65]. The defect in basal activities of MC4R mutations can cause obesity [69,78]. Mrap2- and Agrp-suppressed basal activity of Mc4r play an important role in promoting the growth of zebrafish and culter [31,52,79]. These studies indicate that the basal activity of MC4R plays a pivotal role in the modulation of energy homeostasis [80]. Human MRAP1s increased [37,41,43] or did not affect [30,36] the basal activities of hMC4R, and MRAP2s decreased [43] or had no effect [30,36,41,81] on the basal activities of hMC4R. Our studies showed that cMRAP1 and cMRAP2s decreased the basal cAMP signaling of cMC4R. Decreased MC4R basal activities by MRAP2(s) were also reported in other species [31,51,53,75,77].
Alternative splicing is prevalent in eukaryotes and isoforms generated by alternative splicing might have different functions [82,83,84,85]. Splicing variants provide a nature-made chance to investigate the roles of specific domains. Human MRAP and MRAP2 have two and three alternatively spliced forms, respectively, and they show different effects on the hMC3R/hMC4R pharmacology [28,43]. Canine MRAP2 also had two alternatively spliced variants, MRAP2a and MRAP2b (MRAP2b with extension at N-terminus compared with MRAP2a). This extension sequence at the N-terminus of MRAP2b is not found in other MRAP2s. Our results showed that MRAP2a and MRAP2b had different effects on cMC3R/cMC4R pharmacology. The N-termini of MRAP1 and MRAP2, with several conserved motifs, have important roles in modulating GPCR pharmacology [43,86,87,88]. We speculate that the extension sequences at the N-termini of MRAP2 might have important roles in MC3R/MC4R pharmacology.
5. Conclusions
In summary, we cloned canine MC3R and investigated its pharmacology, as well as modulation of MC3R and MC4R pharmacology by MRAPs. MRAP1 did not affect the MC3R trafficking and decreased α-MSH- and ACTH-induced signaling, whereas MRAP1 increased the cell surface expression and decreased the basal activity of cMC4R. The two MRAP2 isoforms exerted different effects on cMC3R or MC4R pharmacology. MRAP2a decreased α-MSH- and ACTH-induced signaling, whereas MRAP2b only the decreased α-MSH-stimulated signaling of cMC3R. MRAP2a increased the cell surface expression and ACTH-induced signaling, decreased the basal activity of cMC4R, whereas MRAP2b had no effect on trafficking, and decreased basal and α-MSH- and ACTH-induced signaling. This study contributes to a better understanding of cMC3R/cMC4R.
Author Contributions
R.-L.J.: methodology, software, formal analysis, validation, manuscript writing and editing; S.-S.J.: methodology and formal analysis; Y.-X.T.: conceptualization, data curation, supervision, manuscript revision and editing. All authors have read and agreed to the published version of the manuscript.
Funding
R.-L.J. and S.-S.J. received fellowships from China Scholarship Council, People’s Republic of China.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors upon request, without undue reservation.
Conflicts of Interest
The authors declare that there is no conflict of interest that would prejudice the impartiality of this study.
References
- Gantz, I.; Miwa, H.; Konda, Y.; Shimoto, Y.; Tashiro, T.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular cloning, expression, and gene localization of a fourth melanocortin receptor. J. Biol. Chem. 1993, 268, 15174–15179. [Google Scholar] [CrossRef]
- Gantz, I.; Konda, Y.; Tashiro, T.; Shimoto, Y.; Miwa, H.; Munzert, G.; Watson, S.J.; DelValle, J.; Yamada, T. Molecular cloning of a novel melanocortin receptor. J. Biol. Chem. 1993, 268, 8246–8250. [Google Scholar] [CrossRef]
- Roselli-Rehfuss, L.; Mountjoy, K.G.; Robbins, L.S.; Mortrud, M.T.; Low, M.J.; Tatro, J.B.; Entwistle, M.L.; Simerly, R.B.; Cone, R.D. Identification of a receptor for γ melanotropin and other proopiomelanocortin peptides in the hypothalamus and limbic system. Proc. Natl. Acad. Sci. USA 1993, 90, 8856–8860. [Google Scholar] [CrossRef] [Green Version]
- Mountjoy, K.G.; Mortrud, M.T.; Low, M.J.; Simerly, R.B.; Cone, R.D. Localization of the melanocortin-4 receptor (MC4-R) in neuroendocrine and autonomic control circuits in the brain. Mol. Endocrinol. 1994, 8, 1298–1308. [Google Scholar]
- Cone, R.D. Anatomy and regulation of the central melanocortin system. Nat. Neurosci. 2005, 8, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.X. Molecular mechanisms of the neural melanocortin receptor dysfunction in severe early onset obesity. Mol. Cell. Endocrinol. 2005, 239, 1–14. [Google Scholar] [CrossRef]
- Liu, T.; Ji, R.L.; Tao, Y.X. Naturally occurring mutations in G protein-coupled receptors associated with obesity and type 2 diabetes mellitus. Pharmacol. Ther. 2022, 234, 108044. [Google Scholar] [CrossRef]
- Chen, A.S.; Marsh, D.J.; Trumbauer, M.E.; Frazier, E.G.; Guan, X.M.; Yu, H.; Rosenblum, C.I.; Vongs, A.; Feng, Y.; Cao, L.; et al. Inactivation of the mouse melanocortin-3 receptor results in increased fat mass and reduced lean body mass. Nat. Genet. 2000, 26, 97–102. [Google Scholar] [CrossRef]
- Butler, A.A.; Kesterson, R.A.; Khong, K.; Cullen, M.J.; Pelleymounter, M.A.; Dekoning, J.; Baetscher, M.; Cone, R.D. A unique metabolic syndrome causes obesity in the melanocortin-3 receptor-deficient mouse. Endocrinology 2000, 141, 3518–3521. [Google Scholar] [CrossRef]
- Zhang, Y.; Kilroy, G.E.; Henagan, T.M.; Prpic-Uhing, V.; Richards, W.G.; Bannon, A.W.; Mynatt, R.L.; Gettys, T.W. Targeted deletion of melanocortin receptor subtypes 3 and 4, but not CART, alters nutrient partitioning and compromises behavioral and metabolic responses to leptin. FASEB J. 2005, 19, 1482–1491. [Google Scholar] [CrossRef]
- Huszar, D.; Lynch, C.A.; Fairchild-Huntress, V.; Dunmore, J.H.; Fang, Q.; Berkemeier, L.R.; Gu, W.; Kesterson, R.A.; Boston, B.A.; Cone, R.D.; et al. Targeted disruption of the melanocortin-4 receptor results in obesity in mice. Cell 1997, 88, 131–141. [Google Scholar] [CrossRef]
- Balthasar, N.; Dalgaard, L.T.; Lee, C.E.; Yu, J.; Funahashi, H.; Williams, T.; Ferreira, M.; Tang, V.; McGovern, R.A.; Kenny, C.D.; et al. Divergence of melanocortin pathways in the control of food intake and energy expenditure. Cell 2005, 123, 493–505. [Google Scholar] [CrossRef]
- Tao, Y.X. Mutations in melanocortin-4 receptor and human obesity. Prog. Mol. Biol. Transl. Sci. 2009, 88, 173–204. [Google Scholar]
- Tao, Y.X. Mutations in the melanocortin-3 receptor (MC3R) gene: Impact on human obesity or adiposity. Curr. Opin. Investig. Drugs 2010, 11, 1092–1096. [Google Scholar] [PubMed]
- Tao, Y.X. The melanocortin-4 receptor: Physiology, pharmacology, and pathophysiology. Endocr. Rev. 2010, 31, 506–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Tao, Y.X. Mutations in melanocortin-3 receptor gene and human obesity. Prog. Mol. Biol. Transl. Sci. 2016, 140, 97–129. [Google Scholar] [CrossRef]
- Chandramohan, G.; Durham, N.; Sinha, S.; Norris, K.; Vaziri, N.D. Role of γ melanocyte-stimulating hormone-renal melanocortin 3 receptor system in blood pressure regulation in salt-resistant and salt-sensitive rats. Metabolism 2009, 58, 1424–1429. [Google Scholar] [CrossRef]
- Versteeg, D.H.; Van Bergen, P.; Adan, R.A.; De Wildt, D.J. Melanocortins and cardiovascular regulation. Eur. J. Pharmacol. 1998, 360, 1–14. [Google Scholar] [CrossRef]
- Mioni, C.; Giuliani, D.; Cainazzo, M.M.; Leone, S.; Bazzani, C.; Grieco, P.; Novellino, E.; Tomasi, A.; Bertolini, A.; Guarini, S. Further evidence that melanocortins prevent myocardial reperfusion injury by activating melanocortin MC3 receptors. Eur. J. Pharmacol. 2003, 477, 227–234. [Google Scholar] [CrossRef]
- Getting, S.J.; Christian, H.C.; Lam, C.W.; Gavins, F.N.; Flower, R.J.; Schioth, H.B.; Perretti, M. Redundancy of a functional melanocortin 1 receptor in the anti-inflammatory actions of melanocortin peptides: Studies in the recessive yellow (e/e) mouse suggest an important role for melanocortin 3 receptor. J. Immunol. 2003, 170, 3323–3330. [Google Scholar] [CrossRef] [Green Version]
- Catania, A.; Gatti, S.; Colombo, G.; Lipton, J.M. Targeting melanocortin receptors as a novel strategy to control inflammation. Pharmacol. Rev. 2004, 56, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Getting, S.J.; Riffo-Vasquez, Y.; Pitchford, S.; Kaneva, M.; Grieco, P.; Page, C.P.; Perretti, M.; Spina, D. A role for MC3R in modulating lung inflammation. Pulm. Pharmacol. Ther. 2008, 21, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.B.; Montero-Melendez, T.; Greco, K.V.; Perretti, M. Melanocortin receptors as novel effectors of macrophage responses in inflammation. Front. Immunol. 2011, 2, 41. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Guo, D.Y.; Lin, Y.J.; Tao, Y.X. Melanocortin regulation of inflammation. Front. Endocrinol. 2019, 10, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, B.Y.H.; Williamson, A.; Finer, S.; Day, F.R.; Tadross, J.A.; Goncalves Soares, A.; Wade, K.; Sweeney, P.; Bedenbaugh, M.N.; Porter, D.T.; et al. MC3R links nutritional state to childhood growth and the timing of puberty. Nature 2021, 599, 436–441. [Google Scholar] [CrossRef]
- Smith, A.I.; Funder, J.W. Proopiomelanocortin processing in the pituitary, central nervous system, and peripheral tissues. Endocr. Rev. 1988, 9, 159–179. [Google Scholar] [CrossRef]
- Dores, R.M.; Lecaude, S. Trends in the evolution of the proopiomelanocortin gene. Gen. Comp. Endocrinol. 2005, 142, 81–93. [Google Scholar] [CrossRef]
- Metherell, L.A.; Chapple, J.P.; Cooray, S.; David, A.; Becker, C.; Ruschendorf, F.; Naville, D.; Begeot, M.; Khoo, B.; Nurnberg, P.; et al. Mutations in MRAP, encoding a new interacting partner of the ACTH receptor, cause familial glucocorticoid deficiency type 2. Nat. Genet. 2005, 37, 166–170. [Google Scholar] [CrossRef]
- Sebag, J.A.; Hinkle, P.M. Melanocortin-2 receptor accessory protein MRAP forms antiparallel homodimers. Proc. Natl. Acad. Sci. USA 2007, 104, 20244–20249. [Google Scholar] [CrossRef] [Green Version]
- Chan, L.F.; Webb, T.R.; Chung, T.T.; Meimaridou, E.; Cooray, S.N.; Guasti, L.; Chapple, J.P.; Egertova, M.; Elphick, M.R.; Cheetham, M.E.; et al. MRAP and MRAP2 are bidirectional regulators of the melanocortin receptor family. Proc. Natl. Acad. Sci. USA 2009, 106, 6146–6151. [Google Scholar] [CrossRef] [Green Version]
- Sebag, J.A.; Zhang, C.; Hinkle, P.M.; Bradshaw, A.M.; Cone, R.D. Developmental control of the melanocortin-4 receptor by MRAP2 proteins in zebrafish. Science 2013, 341, 278–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouault, A.A.J.; Srinivasan, D.K.; Yin, T.C.; Lee, A.A.; Sebag, J.A. Melanocortin receptor accessory proteins (MRAPs): Functions in the melanocortin system and beyond. Biochim. Biophys. Acta 2017, 1864, 2322–2329. [Google Scholar] [CrossRef]
- Tao, Y.X. Molecular chaperones and G protein-coupled receptor maturation and pharmacology. Mol. Cell. Endocrinol. 2020, 511, 110862. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Rached, M.; Gallo-Payet, N. Differential regulation of the human adrenocorticotropin receptor [melanocortin-2 receptor (MC2R)] by human MC2R accessory protein isoforms alpha and beta in isogenic human embryonic kidney 293 cells. Mol. Endocrinol. 2007, 21, 1656–1669. [Google Scholar] [CrossRef] [Green Version]
- Novoselova, T.; King, P.; Guasti, L.; Metherell, L.A.; Clark, A.J.L.; Chan, L.F. ACTH signalling and adrenal development: Lessons from mouse models. Endocr. Connect. 2019, 8, R122–R130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinkle, P.M.; Serasinghe, M.N.; Jakabowski, A.; Sebag, J.A.; Wilson, K.R.; Haskell-Luevano, C. Use of chimeric melanocortin-2 and -4 receptors to identify regions responsible for ligand specificity and dependence on melanocortin 2 receptor accessory protein. Eur. J. Pharmacol. 2011, 660, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Kay, E.I.; Botha, R.; Montgomery, J.M.; Mountjoy, K.G. hMRAPa increases αMSH-induced hMC1R and hMC3R functional coupling and hMC4R constitutive activity. J. Mol. Endocrinol. 2013, 50, 203–215. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, X.; Zhou, Y.; Cui, L.; Li, J.; Wu, C.; Wan, Y.; Li, J.; Wang, Y. The interaction of MC3R and MC4R with MRAP2, ACTH, α-MSH and AgRP in chickens. J. Endocrinol. 2017, 234, 155–174. [Google Scholar] [CrossRef]
- Tai, X.; Xue, S.; Zhang, C.; Liu, Y.; Chen, J.; Han, Y.; Lin, G.; Zhang, C. Pharmacological evaluation of MRAP proteins on Xenopus neural melanocortin signaling. J. Cell. Physiol. 2021, 236, 6344–6361. [Google Scholar] [CrossRef]
- Kay, E.I.; Botha, R.; Montgomery, J.M.; Mountjoy, K.G. hMRAPa specifically alters hMC4R molecular mass and N-linked complex glycosylation in HEK293 cells. J. Mol. Endocrinol. 2013, 50, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Kay, E.I.; Botha, R.; Montgomery, J.M.; Mountjoy, K.G. hMRAPα, but not hMRAP2, enhances hMC4R constitutive activity in HEK293 cells and this is not dependent on hMRAPα induced changes in hMC4R complex N-linked glycosylation. PLoS ONE 2015, 10, e0140320. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xu, Y.; Zheng, J.; Kuang, Z.; Zhang, C.; Li, N.; Lin, G.; Zhang, C. Pharmacological modulation of dual melanocortin-4 receptor signaling by melanocortin receptor accessory proteins in the Xenopus laevis. J. Cell. Physiol. 2021, 236, 5980–5993. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.L.; Tao, Y.X. Regulation of melanocortin-3 and -4 receptors by isoforms of melanocortin-2 receptor accessory protein 1 and 2. Biomolecules 2022, 12, 244. [Google Scholar] [CrossRef]
- Asai, M.; Ramachandrappa, S.; Joachim, M.; Shen, Y.; Zhang, R.; Nuthalapati, N.; Ramanathan, V.; Strochlic, D.E.; Ferket, P.; Linhart, K.; et al. Loss of function of the melanocortin 2 receptor accessory protein 2 is associated with mammalian obesity. Science 2013, 341, 275–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novoselova, T.V.; Larder, R.; Rimmington, D.; Lelliott, C.; Wynn, E.H.; Gorrigan, R.J.; Tate, P.H.; Guasti, L.; O’Rahilly, S.; Clark, A.J.L. Loss of Mrap2 is associated with Sim1 deficiency and increased circulating cholesterol. J. Endocrinol. 2016, 230, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Geets, E.; Zegers, D.; Beckers, S.; Verrijken, A.; Massa, G.; Van Hoorenbeeck, K.; Verhulst, S.; Van Gaal, L.; Van Hul, W. Copy number variation (CNV) analysis and mutation analysis of the 6q14.1-6q16.3 genes SIM1 and MRAP2 in Prader Willi like patients. Mol. Genet. Metab. 2016, 117, 383–388. [Google Scholar] [CrossRef]
- Baron, M.; Maillet, J.; Huyvaert, M.; Dechaume, A.; Boutry, R.; Loiselle, H.; Durand, E.; Toussaint, B.; Vaillant, E.; Philippe, J.; et al. Loss-of-function mutations in MRAP2 are pathogenic in hyperphagic obesity with hyperglycemia and hypertension. Nat. Med. 2019, 25, 1733–1738. [Google Scholar] [CrossRef]
- da Fonseca, A.C.P.; Abreu, G.M.; Zembrzuski, V.M.; Campos Junior, M.; Carneiro, J.R.I.; Nogueira Neto, J.F.; Magno, F.; Rosado, E.L.; Bozza, P.T.; de Cabello, G.M.K.; et al. Study of LEP, MRAP2 and POMC genes as potential causes of severe obesity in Brazilian patients. Eat. Weight Disord. 2021, 26, 1399–1408. [Google Scholar] [CrossRef]
- Yang, L.K.; Zhang, Z.R.; Wen, H.S.; Tao, Y.X. Characterization of channel catfish (Ictalurus punctatus) melanocortin-3 receptor reveals a potential network in regulation of energy homeostasis. Gen. Comp. Endocrinol. 2019, 277, 90–103. [Google Scholar] [CrossRef]
- Zhang, J.; Li, J.; Wu, C.; Hu, Z.; An, L.; Wan, Y.; Fang, C.; Zhang, X.; Li, J.; Wang, Y. The Asp298Asn polymorphism of melanocortin-4 receptor (MC4R) in pigs: Evidence for its potential effects on MC4R constitutive activity and cell surface expression. Anim. Genet. 2020, 51, 694–706. [Google Scholar] [CrossRef]
- Tao, M.; Ji, R.L.; Huang, L.; Fan, S.Y.; Liu, T.; Liu, S.J.; Tao, Y.X. Regulation of melanocortin-4 receptor pharmacology by two isoforms of melanocortin receptor accessory protein 2 in topmouth culter (Culter alburnus). Front. Endocrinol. 2020, 11, 538. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.L.; Huang, L.; Wang, Y.; Liu, T.; Fan, S.Y.; Tao, M.; Tao, Y.X. Topmouth culter melanocortin-3 receptor: Regulation by two isoforms of melanocortin-2 receptor accessory protein 2. Endocr. Connect. 2021, 10, 1489–1501. [Google Scholar] [CrossRef]
- Wen, Z.Y.; Liu, T.; Qin, C.J.; Zou, Y.C.; Wang, J.; Li, R.; Tao, Y.X. MRAP2 interaction with melanocortin-4 receptor in snakehead (Channa argus). Biomolecules 2021, 11, 481. [Google Scholar] [CrossRef]
- Yan, J.; Tao, Y.X. Pharmacological characterization of canine melancortin-4 receptor and its natural variant V213F. Domest. Anim. Endocrinol. 2011, 41, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Steiner, A.L.; Kipnis, D.M.; Utiger, R.; Parker, C. Radioimmunoassay for the measurement of adenosine 3′,5′-cyclic phosphate. Proc. Natl. Acad. Sci. USA 1969, 64, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Mo, X.L.; Yang, R.; Tao, Y.X. Functions of transmembrane domain 3 of human melanocortin-4 receptor. J. Mol. Endocrinol. 2012, 49, 221–235. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.X.; Segaloff, D.L. Functional characterization of melanocortin-4 receptor mutations associated with childhood obesity. Endocrinology 2003, 144, 4544–4551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Okayama, H. High-efficiency transformation of mammalian cells by plasmid DNA. Mol. Cell Biol. 1987, 7, 2745–2752. [Google Scholar] [PubMed] [Green Version]
- Wang, S.X.; Fan, Z.C.; Tao, Y.X. Functions of acidic transmembrane residues in human melanocortin-3 receptor binding and activation. Biochem Pharmacol 2008, 76, 520–530. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.Q.; Hou, Z.S.; Wen, H.S.; Li, Y.; Qi, X.; Li, W.J.; Tao, Y.X. Melanocortin-4 receptor in spotted sea bass, Lateolabrax maculatus: Cloning, tissue distribution, physiology, and pharmacology. Front. Endocrinol. 2019, 10, 705. [Google Scholar] [CrossRef] [Green Version]
- Gruber, K.A.; Ji, R.L.; Gallazzi, F.; Jiang, S.; Van Doren, S.R.; Tao, Y.X.; Newton Northup, J. Development of a therapeutic peptide for cachexia suggests a platform approach for drug-like peptides. ACS Pharmacol. Transl. Sci. 2022, 5, 344–361. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.J.; Xie, H.J.; Wang, W.; Wang, Z.Q.; Tao, Y.X. Pharmacology of the giant panda (Ailuropoda melanoleuca) melanocortin-3 receptor. Gen. Comp. Endocrinol. 2019, 277, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.C.; Sartin, J.L.; Tao, Y.X. Molecular cloning and pharmacological characterization of porcine melanocortin-3 receptor. J. Endocrinol. 2008, 196, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.X. Functional characterization of novel melanocortin-3 receptor mutations identified from obese subjects. Biochim. Biophys. Acta 2007, 1772, 1167–1174. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.X.; Huang, H.; Wang, Z.Q.; Yang, F.; Williams, J.N.; Nikiforovich, G.V. Constitutive activity of neural melanocortin receptors. Methods Enzymol. 2010, 484, 267–279. [Google Scholar]
- Renquist, B.J.; Zhang, C.; Williams, S.Y.; Cone, R.D. Development of an assay for high-throughput energy expenditure monitoring in the zebrafish. Zebrafish 2013, 10, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xue, S.; Lei, X.; Song, W.; Li, L.; Li, X.; Fu, Y.; Zhang, C.; Zhang, H.; Luo, Y.; et al. Pharmacological evaluation of melanocortin 2 receptor accessory protein 2 on axolotl neural melanocortin signaling. Front. Endocrinol. 2022, 13, 820896. [Google Scholar] [CrossRef]
- Zhang, H.J.; Cui, Z.H.; Liu, M.; Min, T.Q.; Xiao, X.; Wang, Z.Q.; Tao, Y.X. Pharmacological characterization of three chicken melanocortin-3 receptor mutants. Domest. Anim. Endocrinol. 2021, 74, 106507. [Google Scholar] [CrossRef]
- Srinivasan, S.; Lubrano-Berthelier, C.; Govaerts, C.; Picard, F.; Santiago, P.; Conklin, B.R.; Vaisse, C. Constitutive activity of the melanocortin-4 receptor is maintained by its N-terminal domain and plays a role in energy homeostasis in humans. J. Clin. Investig. 2004, 114, 1158–1164. [Google Scholar] [CrossRef] [Green Version]
- Ersoy, B.A.; Pardo, L.; Zhang, S.; Thompson, D.A.; Millhauser, G.; Govaerts, C.; Vaisse, C. Mechanism of N-terminal modulation of activity at the melanocortin-4 receptor GPCR. Nat. Chem. Biol. 2012, 8, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Nishi, S.; Nakabayashi, K.; Kobilka, B.; Hsueh, A.J.W. The ectodomain of the luteinizing hormone receptor interacts with exoloop 2 to constrain the transmembrane region. Studies using chimeric human and fly receptors. J. Biol. Chem. 2002, 277, 3958–3964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parma, J.; Van Sande, J.; Swillens, S.; Tonacchera, M.; Dumont, J.; Vassart, G. Somatic mutations causing constitutive activity of the thyrotropin receptor are the major cause of hyperfunctioning thyroid adenomas: Identification of additional mutations activating both the cyclic adenosine 3′,5′-monophosphate and inositol phosphate-Ca2+ cascades. Mol. Endocrinol. 1995, 9, 725–733. [Google Scholar] [PubMed] [Green Version]
- Zhang, M.; Tong, K.P.; Fremont, V.; Chen, J.; Narayan, P.; Puett, D.; Weintraub, B.D.; Szkudlinski, M.W. The extracellular domain suppresses constitutive activity of the transmembrane domain of the human TSH receptor: Implications for hormone-receptor interaction and antagonist design. Endocrinology 2000, 141, 3514–3517. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Li, L.; Jin, X.; Xu, B.; Pi, L.; Liu, S.; Zhu, W.; Zhang, C.; Luan, B.; Gong, L.; et al. Pharmacological effect of human melanocortin-2 receptor accessory protein 2 variants on hypothalamic melanocortin receptors. Endocrine 2018, 61, 94–104. [Google Scholar] [CrossRef]
- Wang, M.; Chen, Y.J.; Zhu, M.; Xu, B.X.; Guo, W.X.; Lyu, Y.S.; Zhang, C. Pharmacological modulation of melanocortin-4 receptor by melanocortin receptor accessory protein 2 in Nile tilapia. Gen. Comp. Endocrinol. 2019, 282, 113219. [Google Scholar] [CrossRef]
- Soletto, L.; Hernandez-Balfago, S.; Rocha, A.; Scheerer, P.; Kleinau, G.; Cerdá-Reverter, J.M. Melanocortin receptor accessory protein 2-induced adrenocorticotropic hormone response of human melanocortin 4 receptor. J. Endocr. Soc. 2019, 3, 314–323. [Google Scholar] [CrossRef] [Green Version]
- Rao, Y.Z.; Chen, R.; Zhang, Y.; Tao, Y.X. Orange-spotted grouper melanocortin-4 receptor: Modulation of signaling by MRAP2. Gen. Comp. Endocrinol. 2019, 284, 113234. [Google Scholar] [CrossRef]
- Tao, Y.X. Constitutive activation of G protein-coupled receptors and diseases: Insights into mechanism of activation and therapeutics. Pharmacol. Ther. 2008, 120, 129–148. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Forlano, P.M.; Cone, R.D. AgRP and POMC neurons are hypophysiotropic and coordinately regulate multiple endocrine axes in a larval teleost. Cell Metab. 2012, 15, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.X. Constitutive activity in melanocortin-4 receptor: Biased signaling of inverse agonists. Adv. Pharmacol. 2014, 70, 135–154. [Google Scholar] [CrossRef]
- Gillyard, T.; Fowler, K.; Williams, S.Y.; Cone, R.D. Obesity-associated mutant melanocortin-4 receptors with normal Gαs coupling frequently exhibit other discoverable pharmacological and biochemical defects. J. Neuroendocrinol. 2019, 31, e12795. [Google Scholar] [CrossRef] [PubMed]
- Black, D.L. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem. 2003, 72, 291–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008, 40, 1413–1415. [Google Scholar] [CrossRef]
- Wahl, M.C.; Will, C.L.; Luhrmann, R. The spliceosome: Design principles of a dynamic RNP machine. Cell 2009, 136, 701–718. [Google Scholar] [CrossRef] [Green Version]
- Liang, Q.; Wu, N.; Zaneveld, S.; Liu, H.; Fu, S.; Wang, K.; Bertrand, R.; Wang, J.; Li, Y.; Chen, R. Transcript isoforms of Reep6 have distinct functions in the retina. Hum. Mol. Genet. 2021, 30, 1907–1918. [Google Scholar] [CrossRef] [PubMed]
- Webb, T.R.; Chan, L.; Cooray, S.N.; Cheetham, M.E.; Chapple, J.P.; Clark, A.J. Distinct melanocortin 2 receptor accessory protein domains are required for melanocortin 2 receptor interaction and promotion of receptor trafficking. Endocrinology 2009, 150, 720–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouault, A.A.J.; Lee, A.A.; Sebag, J.A. Regions of MRAP2 required for the inhibition of orexin and prokineticin receptor signaling. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 2322–2329. [Google Scholar] [CrossRef]
- Rouault, A.A.J.; Rosselli-Murai, L.K.; Hernandez, C.C.; Gimenez, L.E.; Tall, G.G.; Sebag, J.A. The GPCR accessory protein MRAP2 regulates both biased signaling and constitutive activity of the ghrelin receptor GHSR1a. Sci. Signal. 2020, 13, eaax4569. [Google Scholar] [CrossRef]
Figure 1.
Nucleotide and deduced amino acid sequences (A) and phylogenetic tree (B) of cMC3R. Positions of nucleotide and amino acid sequences are indicated on both sides. N-linked glycosylation sites are present in open boxes. Shaded boxes show putative TMD1-7. The conserved motifs (PMY, DRY and DPxxY) are underlined. Asterisk (*) shows stop codon. The tree was constructed by the neighbor-joining (NJ) method. Numbers at nodes indicate the bootstrap value, as percentages, obtained for 1000 replicates. Black dot denotes canine MC3R. MC3Rs: Canis lupus familiaris (dog, NM_001135124.1), Capra hircus (goat, XP_005688382.1), Xenopus tropicalis (frog, XP_002935436.1), Culter alburnus (topmouth culter, MT419813), Carassius auratus (goldfish, BAJ83473.1), Danio rerio (zebrafish, AAO24744.1), Homo sapiens (human, NP_063941.3), Sus scrofa (pig, AFK25142.1), Mus musculus (mouse, AAI03670.1), Gallus gallus (chicken, XP_004947293.1), Rattus norvegicus (rat, NP_001020441.3), Pangasianodon hypophthalmus (iridescent shark, XP_026770221.1), Equus caballus (horse, NP_001243901.1), Pteropus vampyrus (large flying fox, XP_011368476.1), Pteropus alecto (black flying fox, XP_006921991.1), Felis catus (cat, XP_023106851.1), Loxodonta africana (African bush elephant, XP_003419952.1), Salvelinus alpinus (Arctic char, XP_023994975.1), Pelodiscus sinensis (Chinese softshell turtle, XP_006129463.1), and Alligator sinensis (Chinese alligator, XP_006018246.1).
Figure 1.
Nucleotide and deduced amino acid sequences (A) and phylogenetic tree (B) of cMC3R. Positions of nucleotide and amino acid sequences are indicated on both sides. N-linked glycosylation sites are present in open boxes. Shaded boxes show putative TMD1-7. The conserved motifs (PMY, DRY and DPxxY) are underlined. Asterisk (*) shows stop codon. The tree was constructed by the neighbor-joining (NJ) method. Numbers at nodes indicate the bootstrap value, as percentages, obtained for 1000 replicates. Black dot denotes canine MC3R. MC3Rs: Canis lupus familiaris (dog, NM_001135124.1), Capra hircus (goat, XP_005688382.1), Xenopus tropicalis (frog, XP_002935436.1), Culter alburnus (topmouth culter, MT419813), Carassius auratus (goldfish, BAJ83473.1), Danio rerio (zebrafish, AAO24744.1), Homo sapiens (human, NP_063941.3), Sus scrofa (pig, AFK25142.1), Mus musculus (mouse, AAI03670.1), Gallus gallus (chicken, XP_004947293.1), Rattus norvegicus (rat, NP_001020441.3), Pangasianodon hypophthalmus (iridescent shark, XP_026770221.1), Equus caballus (horse, NP_001243901.1), Pteropus vampyrus (large flying fox, XP_011368476.1), Pteropus alecto (black flying fox, XP_006921991.1), Felis catus (cat, XP_023106851.1), Loxodonta africana (African bush elephant, XP_003419952.1), Salvelinus alpinus (Arctic char, XP_023994975.1), Pelodiscus sinensis (Chinese softshell turtle, XP_006129463.1), and Alligator sinensis (Chinese alligator, XP_006018246.1).

Figure 2.
Nucleotide and deduced amino acid sequences (A) and phylogenetic tree (B) of cMRAP1. Positions of nucleotide and amino acid sequences are indicated on both sides. N-linked glycosylation sites are present in open boxes. Red box denotes YEYY motif. Light blue box indicates LDYL motif. Black box is LKANKYL motif. Shaded box shows putative TMD. Asterisk (*) shows stop codon. The tree was constructed by the neighbor-joining (NJ) method. Numbers at nodes indicate the bootstrap value, as percentages, obtained for 1000 replicates. Black dot denotes canine MRAP1. MRAP1s: Canis lupus familiaris (dog, XP_005638887.1), Homo sapiens (human MRAP1a, AAH62721.1; human MRAP1b, NP_996781.1), Mus musculus (mouse, NP_084120.1); Macaca mulatta (monkey, XP_001096328.3), Gallus gallus (chicken, XR_001470382.2), Chrysemys picta bellii (turtle, XP_005283970.1), Xenopus tropicalis (frog, XP_002938489.2), Danio rerio (zebrafish, ENSDART00000148193.3), Sus scrofa (pig, XP_020926573.1), Cricetulus griseus (chinese hamster, XP_003495626.1), Rattus norvegicus (rat, NP_001129306.1), Lagenorhynchus obliquidens (dolphin, XP_026957114.1), Vulpes vulpes (red fox, XP_025840964.1), Delphinapterus leucas (whale, XP_022408665.2), and Capra hircus (goat, XP_005674803.2).
Figure 2.
Nucleotide and deduced amino acid sequences (A) and phylogenetic tree (B) of cMRAP1. Positions of nucleotide and amino acid sequences are indicated on both sides. N-linked glycosylation sites are present in open boxes. Red box denotes YEYY motif. Light blue box indicates LDYL motif. Black box is LKANKYL motif. Shaded box shows putative TMD. Asterisk (*) shows stop codon. The tree was constructed by the neighbor-joining (NJ) method. Numbers at nodes indicate the bootstrap value, as percentages, obtained for 1000 replicates. Black dot denotes canine MRAP1. MRAP1s: Canis lupus familiaris (dog, XP_005638887.1), Homo sapiens (human MRAP1a, AAH62721.1; human MRAP1b, NP_996781.1), Mus musculus (mouse, NP_084120.1); Macaca mulatta (monkey, XP_001096328.3), Gallus gallus (chicken, XR_001470382.2), Chrysemys picta bellii (turtle, XP_005283970.1), Xenopus tropicalis (frog, XP_002938489.2), Danio rerio (zebrafish, ENSDART00000148193.3), Sus scrofa (pig, XP_020926573.1), Cricetulus griseus (chinese hamster, XP_003495626.1), Rattus norvegicus (rat, NP_001129306.1), Lagenorhynchus obliquidens (dolphin, XP_026957114.1), Vulpes vulpes (red fox, XP_025840964.1), Delphinapterus leucas (whale, XP_022408665.2), and Capra hircus (goat, XP_005674803.2).

Figure 3.
Schematic diagram (A), nucleotide and deduced amino acid sequences (B, cMRAP2a, C, cMRAP2b) and phylogenetic tree (D) of MRAP2s. Positions of nucleotide and amino acid sequences are indicated on both sides. N-linked glycosylation sites are present in open boxes. Red box denotes YEYY motif. Light blue box indicates LDYL motif. Black box is LKANKYL motif. Shaded box shows putative TMD. Asterisk (*) shows stop codon. The tree was constructed by the neighbor-joining (NJ) method. Numbers at nodes indicate the bootstrap value, as percentages, obtained for 1000 replicates. Black dot denotes canine MRAP1. MRAP1s: MRAP2s: Canis lupus familiaris (dog MRAP2a, XM_038682814.1; MRAP2b, XM_038682813.1), Mus musculus (mouse NP_001171202.1), Sus scrofa (pig, XP_003353296.2), Capra hircus (goat, XP_017908670.1), Callorhinchus milii (elephant shark, XP_007906624.1), Balaenoptera musculus (whale, XP_036727732.1), Bos taurus (bovine, NP_001092863.1), Danio rerio (zebrafish, MRAP2a: F8W4H9.1, MRAP2b: F8W4H9.1), Gallus gallus (chicken, ALO81626.1), Mus caroli (Ryukyu mouse, XP_021029091.1), and Homo sapiens (human, AAH10003.2).
Figure 3.
Schematic diagram (A), nucleotide and deduced amino acid sequences (B, cMRAP2a, C, cMRAP2b) and phylogenetic tree (D) of MRAP2s. Positions of nucleotide and amino acid sequences are indicated on both sides. N-linked glycosylation sites are present in open boxes. Red box denotes YEYY motif. Light blue box indicates LDYL motif. Black box is LKANKYL motif. Shaded box shows putative TMD. Asterisk (*) shows stop codon. The tree was constructed by the neighbor-joining (NJ) method. Numbers at nodes indicate the bootstrap value, as percentages, obtained for 1000 replicates. Black dot denotes canine MRAP1. MRAP1s: MRAP2s: Canis lupus familiaris (dog MRAP2a, XM_038682814.1; MRAP2b, XM_038682813.1), Mus musculus (mouse NP_001171202.1), Sus scrofa (pig, XP_003353296.2), Capra hircus (goat, XP_017908670.1), Callorhinchus milii (elephant shark, XP_007906624.1), Balaenoptera musculus (whale, XP_036727732.1), Bos taurus (bovine, NP_001092863.1), Danio rerio (zebrafish, MRAP2a: F8W4H9.1, MRAP2b: F8W4H9.1), Gallus gallus (chicken, ALO81626.1), Mus caroli (Ryukyu mouse, XP_021029091.1), and Homo sapiens (human, AAH10003.2).


Figure 4.
Ligand binding properties of cMC3R. Different concentrations of unlabeled NDP-MSH (A), α-MSH (B), β-MSH (C), ACTH (1-24) (D), and D-Trp8-γ-MSH (E) were used to displace the binding of 125I-NDP-MSH. Results are expressed as % of hMC3R binding ± range from duplicate determinations within one experiment. All experiments were repeated at least three independent times.
Figure 4.
Ligand binding properties of cMC3R. Different concentrations of unlabeled NDP-MSH (A), α-MSH (B), β-MSH (C), ACTH (1-24) (D), and D-Trp8-γ-MSH (E) were used to displace the binding of 125I-NDP-MSH. Results are expressed as % of hMC3R binding ± range from duplicate determinations within one experiment. All experiments were repeated at least three independent times.

Figure 5.
Signaling properties of cMC3R. HEK293T cells were transiently transfected with MC3R plasmids. Different concentrations of NDP-MSH (A), α-MSH (B), β-MSH (C), ACTH (1-24) (D), and D-Trp8-γ-MSH (E) were used to stimulate the cells. Data are means ± SEM from triplicate measurements within one experiment. All experiments were performed at least three times independently.
Figure 5.
Signaling properties of cMC3R. HEK293T cells were transiently transfected with MC3R plasmids. Different concentrations of NDP-MSH (A), α-MSH (B), β-MSH (C), ACTH (1-24) (D), and D-Trp8-γ-MSH (E) were used to stimulate the cells. Data are means ± SEM from triplicate measurements within one experiment. All experiments were performed at least three times independently.

Figure 6.
Regulation of cMC3R (A,B) and cMC4R (C,D) expression by MRAPs. Cell surface and total expression of cMC3R and MC4R was measured by flow cytometry. HEK293T cells were co-transfected with cMC3R or cMC4R and cMRAPs (1:5). Fluorescence in cells transfected with empty vector pcDNA3.1 was used for background staining. The results were calculated as % of 1:0 group. Each data point represented the mean ± SEM (n = 3–4). * Indicates significantly different from 1:0 group (p < 0.05) (one-way ANOVA followed by Tukey test).
Figure 6.
Regulation of cMC3R (A,B) and cMC4R (C,D) expression by MRAPs. Cell surface and total expression of cMC3R and MC4R was measured by flow cytometry. HEK293T cells were co-transfected with cMC3R or cMC4R and cMRAPs (1:5). Fluorescence in cells transfected with empty vector pcDNA3.1 was used for background staining. The results were calculated as % of 1:0 group. Each data point represented the mean ± SEM (n = 3–4). * Indicates significantly different from 1:0 group (p < 0.05) (one-way ANOVA followed by Tukey test).

Figure 7.
Modulation of cMC3R pharmacology by MRAPs. Ligand binding (A,B) and signaling (C,D) properties of cMC4R to α-MSH or ACTH (1-24) upon co-expression of cMC4R with cMRAP1, cMRAP2a or cMRAP2b were measured. Results of binding properties were calculated as % of cMC4R without MRAPs, from duplicate determinations within one experiment. All experiments were performed at least three independent times.
Figure 7.
Modulation of cMC3R pharmacology by MRAPs. Ligand binding (A,B) and signaling (C,D) properties of cMC4R to α-MSH or ACTH (1-24) upon co-expression of cMC4R with cMRAP1, cMRAP2a or cMRAP2b were measured. Results of binding properties were calculated as % of cMC4R without MRAPs, from duplicate determinations within one experiment. All experiments were performed at least three independent times.

Figure 8.
Modulation of cMC4R pharmacology by MRAPs. Ligand binding (A,B) and signaling (C,D) properties of cMC4R to α-MSH or ACTH (1-24) upon co-expression of cMC4R with cMRAP1, cMRAP2a or cMRAP2b were measured. Results of binding properties were calculated as % of cMC4R without MRAPs, from duplicate determinations within one experiment. All experiments were performed at least three independent times.
Figure 8.
Modulation of cMC4R pharmacology by MRAPs. Ligand binding (A,B) and signaling (C,D) properties of cMC4R to α-MSH or ACTH (1-24) upon co-expression of cMC4R with cMRAP1, cMRAP2a or cMRAP2b were measured. Results of binding properties were calculated as % of cMC4R without MRAPs, from duplicate determinations within one experiment. All experiments were performed at least three independent times.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The ligand binding properties of cMC3R.
| MC3R | IC50 (nM) | cMC3R | hMC3R |
|---|---|---|---|
| Bmax (%) | 247.97 ± 13.44 b | 100 | |
| NDP-MSH | IC50 (nM) | 4.37 ± 0.72 a | 1.98 ± 0.43 |
| α-MSH | IC50 (nM) | 240.42 ± 37.34 | 221.57 ± 30.23 |
| β-MSH | IC50 (nM) | 140.18 ± 31.37 | 161.56 ± 31.45 |
| ACTH | IC50 (nM) | 58.52 ± 8.67 | 45.71 ± 5.23 |
| D-Trp8-γ-MSH | IC50 (nM) | 39.30 ± 0.30 b | 24.62 ± 1.25 |
Values are expressed as the mean ± SEM of at least three independent experiments. a Significant difference from the parameter of hMC3R, p < 0.05. b Significant difference from the parameter of hMC3R, p < 0.001.
Table 2.
The signaling properties of cMC3R.
| MC3R | EC50/Rmax | cMC3R | hMC3R |
|---|---|---|---|
| Basal (%) | 100.72 ± 7.69 | 100 | |
| NDP-MSH | EC50 (nM) | 0.40 ± 0.16 | 0.24 ± 0.11 |
| Rmax (%) | 199.38 ± 28.06 a | 100 | |
| α-MSH | EC50 (nM) | 0.97 ± 0.35 | 1.49 ± 0.22 |
| Rmax (%) | 145.69 ± 14.30 | 100 | |
| β-MSH | EC50 (nM) | 1.04 ± 0.23 | 1.46 ± 0.55 |
| Rmax (%) | 229.77 ± 46.45 a | 100 | |
| ACTH | EC50 (nM) | 1.46 ± 0.30 | 2.12 ± 0.69 |
| Rmax (%) | 182.83 ± 25.21 a | 100 | |
| D-Trp8-γ-MSH | EC50 (nM) | 1.08 ± 0.15 | 0.71 ± 0.19 |
| Rmax (%) | 188.25 ± 19.56 a | 100 |
Values are expressed as the mean ± SEM of at least three independent experiments. a Significant difference from the parameter of hMC3R, p < 0.05.
Table 3.
The effect of MRAPs on ligand binding properties of cMC3R.
| cMC3R/cMRAPs | Bmax (%) | α-MSH | ACTH |
|---|---|---|---|
| IC50 (nM) | IC50 (nM) | ||
| cMC3R | 100 | 106.68 ± 13.80 | 55.56 ± 11.64 |
| cMC3R/cMRAP1 | 81.81 ± 5.01 a | 127.76 ± 30.47 | 68.59 ± 9.31 |
| cMC3R/cMRAP2a | 148.43 ± 14.94 a | 118.61 ± 22.25 | 57.58 ± 4.09 |
| cMC3R/cMRAP2b | 128.42 ± 9.06 a | 148.03 ± 30.16 | 49.59 ± 6.77 |
Values are expressed as the mean ± SEM of at least three independent experiments. a Significant difference from the parameter of cMC3R, p < 0.05.
Table 4.
The effect of MRAPs on cAMP signaling of cMC3R.
| cMC3R/cMRAPs | Basal (%) | α-MSH | ACTH | ||
|---|---|---|---|---|---|
| EC50 (nM) | Rmax (%) | EC50 (nM) | Rmax (%) | ||
| cMC3R | 100 | 3.08 ± 1.26 | 100 | 2.02 ± 0.39 | 100 |
| cMC3R/cMRAP1 | 100.91 ± 16.74 | 0.88 ± 0.18 | 46.67 ± 8.88 b | 2.11 ± 0.56 | 46.61 ± 12.37 a |
| cMC3R/cMRAP2a | 84.71 ± 19.03 | 1.37 ± 0.26 | 73.02 ± 5.81 a | 2.18 ± 0.39 | 73.79 ± 4.94 a |
| cMC3R/cMRAP2b | 68.88 ± 9.35 a | 3.64 ± 1.11 | 66.93 ± 9.64 a | 3.41 ± 0.99 | 85.02 ± 11.04 |
Values are expressed as the mean ± SEM of at least three independent experiments. a Significant difference from the parameter of cMC3R, p < 0.05. b Significant difference from the parameter of cMC3R, p < 0.001.
Table 5.
The effect of MRAPs on ligand binding properties of cMC4R.
| cMC4R/cMRAPs | Bmax (%) | α-MSH | ACTH |
|---|---|---|---|
| IC50 (nM) | IC50 (nM) | ||
| cMC4R | 100 | 504.66 ± 124.33 | 348.29 ± 94.51 |
| cMC4R/cMRAP1 | 151.36 ± 10.67 a | 186.62 ± 13.60 a | 37.63 ± 6.84 b |
| cMC4R/cMRAP2a | 167.03 ± 11.83 c | 228.40 ± 40.74 a | 35.63 ± 8.10 b |
| cMC4R/cMRAP2b | 121.60 ± 10.06 | 120.19 ± 22.41 b | 16.93 ± 5.69 c |
Values are expressed as the mean ± SEM of at least three independent experiments. a Significant difference from the parameter of cMC4R, p < 0.05. b Significant difference from the parameter of cMC4R, p < 0.01. c Significant difference from the parameter of cMC4R, p < 0.001.
Table 6.
The effect of MRAPs on signaling properties of cMC4R.
| cMC4R/cMRAPs | Basal (%) | α-MSH | ACTH | ||
|---|---|---|---|---|---|
| EC50 (nM) | Rmax (%) | EC50 (nM) | Rmax (%) | ||
| cMC4R | 100 | 1.18 ± 0.21 | 100 | 0.92 ± 0.41 | 100 |
| cMC4R/cMRAP1 | 53.89 ± 4.06 b | 1.37 ± 0.24 | 99.80 ± 3.80 | 1.92 ± 0.40 | 76.21 ± 13.29 |
| cMC4R/cMRAP2a | 74.70 ± 4.28 b | 0.90 ± 0.10 | 87.00 ± 10.28 | 3.82 ± 1.25 | 144.41 ± 13.36 a |
| cMC4R/cMRAP2b | 55.82 ± 3.43 b | 0.63 ± 0.10 | 52.62 ± 7.93 b | 1.20 ± 0.44 | 55.87 ± 9.82 a |
Values are expressed as the mean ± SEM of at least three independent experiments. a Significant difference from the parameter of cMC4R, p < 0.05. b Significant difference from the parameter of cMC3R, p < 0.001.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ji, R.-L.; Jiang, S.-S.; Tao, Y.-X. Modulation of Canine Melanocortin-3 and -4 Receptors by Melanocortin-2 Receptor Accessory Protein 1 and 2. Biomolecules 2022, 12, 1608. https://doi.org/10.3390/biom12111608
AMA Style
Ji R-L, Jiang S-S, Tao Y-X. Modulation of Canine Melanocortin-3 and -4 Receptors by Melanocortin-2 Receptor Accessory Protein 1 and 2. Biomolecules. 2022; 12(11):1608. https://doi.org/10.3390/biom12111608
Chicago/Turabian StyleJi, Ren-Lei, Shan-Shan Jiang, and Ya-Xiong Tao. 2022. "Modulation of Canine Melanocortin-3 and -4 Receptors by Melanocortin-2 Receptor Accessory Protein 1 and 2" Biomolecules 12, no. 11: 1608. https://doi.org/10.3390/biom12111608
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.