
Identification, Characterization, and Expression of a β-Galactosidase from Arion Species (Mollusca)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Galactosidase Purification from Arion Lusitanicus
2.3. Mass Spectrometry of the Protein
2.4. Expression of the Full-Length β-Galactosidase Gene from Arion Vulgaris
2.5. Determination of Protein Content
2.6. Determination of β-Galactosidase Activity
3. Results
3.1. Purification of β-Galactosidase from Arion Lusitanicus
3.2. Recombinant β-Galactosidase from A. Vulgaris Expressed as Protein in Sf9 Insect Cells
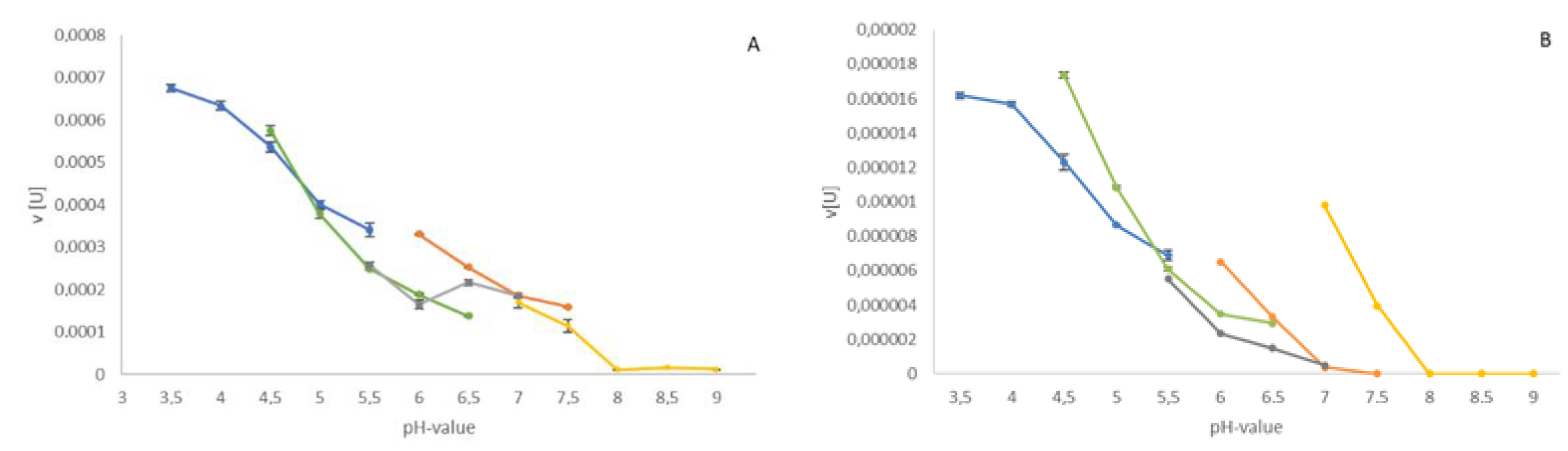
3.3. Biochemical Parameters of Native and Recombinant β-Galactosidases from Two Arion Species
3.4. Determination of Substrate Specificity of β-Galactosidase
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Varki, A. Biological roles of glycans. Glycobiology 2016, 27, 3–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummings, R.D. Stuck on sugars—How carbohydrates regulate cell adhesion, recognition, and signaling. Glycoconj. J. 2019, 36, 241–257. [Google Scholar] [CrossRef]
- Stanley, P.; Moremen, K.W.; Lewis, N.E.; Taniguchi, N.; Aebi, M.; Varki, A.; Cummings, R.D.; Esko, J.D.; Stanley, P.; Hart, G.W.; et al. (Eds.) N-Glycans. In Essentials of Glycobiology, 4th ed.; Cold Spring Harbor Laboratory Press: Long Island, NY, USA, 2022; Chapter 9. [Google Scholar]
- Parodi, A.; Cummings, R.D.; Aebi, M.; Varki, A.; Cummings, R.D.; Esko, J.D.; Stanley, P.; Hart, G.W.; Aebi, M.; Darvill, A.G.; et al. (Eds.) Glycans in Glycoprotein Quality Control. In Essentials of Glycobiology, 3rd ed.; Cold Spring Harbor Laboratory Press: Long Island, NY, USA, 2015; Chapter 39. [Google Scholar]
- Wanninger, A.; Wollesen, T. The evolution of molluscs. Biol. Rev. 2018, 94, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Staudacher, E. Mucin-Type O-Glycosylation in Invertebrates. Molecules 2015, 20, 10622–10640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staudacher, E. Mollusc N-glycosylation: Structures, Functions and Perspectives. Biomolecules 2021, 11, 1820. [Google Scholar] [CrossRef]
- Drula, E.; Garron, M.L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic. Acids. Res. 2022, 50, 571–577. [Google Scholar] [CrossRef]
- Vetere, A.; Paoletti, S. High-yield synthesis of N-acetyllactosamine by regioselective transglycosylation. Biochem. Biophys. Res. Commun. 1996, 219, 6–13. [Google Scholar] [CrossRef]
- Osman, A. Synthesis of Prebiotic Galacto-Oligosaccharides: Science and Technology. In Probiotics, Prebiotics, and Synbiotics: Bioactive Foods in Health Promotion, 1st ed.; Watson, R.R., Preedy, V.R., Levy, N., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; Chapter 9; pp. 135–154. [Google Scholar]
- Usvalampi, A.; Maaheimo, H.; Tossavainen, O.; Frey, A.D. Enzymatic synthesis of fucose-containing galacto-oligosaccharides using β-galactosidase and identification of novel disaccharide structures. Glycoconj. J. 2018, 35, 31–40. [Google Scholar] [CrossRef]
- Jacobson, R.H.; Zhang, X.-J.; DuBose, R.F.; Mattews, B.W. Three-dimensional structure of β-galactosidase from E. coli. Nature 1994, 369, 761–766. [Google Scholar] [CrossRef]
- Juers, D.H.; Jacobson, R.H.; Wigley, D.; Zhang, X.-J.; Huber, R.E.; Tronrud, D.E.; Mattews, B.W. High resolution refinement of -galactosidase in a new crystal from reveals multiple metal-binding sites and provides a structural basis for α-complementation. Protein Sci. 2000, 9, 1685–1699. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Splechtna, B.; Krasteva, S.; Kneifel, W.; Kulbe, K.D.; Divne, C.; Haltrich, D. Characterization and molecular cloning of a heterodimeric β-galactosidase from the probiotic strain Lactobacillus acidophilus R22. FEMS Microbiol. Lett. 2007, 269, 136–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becerra, M.; Cerdán, E.; Siso, M.I.G. Micro-scale purification of β-galactosidase from Kluyveromyces lactis reveals that dimeric and tetrameric forms are active. Biotechnol. Tech. 1998, 12, 253–256. [Google Scholar] [CrossRef]
- Neustroev, K.N.; De Sousa, E.A.; Golubev, A.M.; Brandao Neto, J.R.; Eneyskaya, E.V.; Kulminskaya, A.A.; Polikarpov, I. Purification, crystallization and preliminary diffraction study of β-galactosidase from Penicillium sp. Acta Crystallogr. Sect. D Struct. Biol. 2000, 56, 1508–1509. [Google Scholar] [CrossRef] [PubMed]
- Pilipenko, O.S.; Atyaksheva, L.F.; Poltorak, O.M.; Chukhrai, E.S. Dissociation and Catalytic Activity of Oligomer Forms of β-Galactosidases. Russ. J. Phys. Chem. A 2006, 81, 990–994. [Google Scholar] [CrossRef]
- Husain, Q. β-galactosidases and their potential applications: A review. Crit. Rev. Biotechnol. 2009, 30, 41–62. [Google Scholar] [CrossRef]
- Saqib, S.; Akram, A.; Halim, S.A.; Tassaduq, R. Sources of β-galactosidase and its applications in food industry. 3 Biotech 2017, 7, 79. [Google Scholar] [CrossRef] [Green Version]
- Skudlarek, M.D.; Tulsiani, D.R.; Orgebin-Crist, M.C. Rat epididymal luminal fluid acid beta-D-galactosidase optimally hydrolyses glycoprotein substrate at neutral pH. Biochem. J. 1992, 286, 907–914. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Nishimura, K. Aggregation-dissociation and stability of acid β-galactosidase purified from porcine spleen. Int. J. Biochem. 1986, 18, 327–335. [Google Scholar] [CrossRef]
- Bretting, H.; Jacobs, G.; Müller, S.; Breitfeld, O. The galactan-degrading enzymes in the snail Biomphalaria glabrata. Comp. Biochem. Physiol. B 1991, 99, 629–636. [Google Scholar] [CrossRef]
- Leparoux, S.; Padrines, M.; Placier, G.; Colas, B. Characterization of a strictly specific acid beta-galactosidase from Achatina achatina. Biochim. Biophys. Acta-Gen. Subj. 1997, 1336, 522–532. [Google Scholar] [CrossRef]
- Oliveira, F.W.; Chavante, S.F.; Santos, E.A.; Dietrich, C.P.; Nader, H.B. Appearance and fate of a beta-galactanase, alpha, beta-galactosidases, heparan sulfate and chondroitin sulfate degrading enzymes during embryonic development of the mollusc Pomacea sp. Biochim. Biophys. Acta 1994, 1200, 241–246. [Google Scholar] [CrossRef]
- Escobar-Correas, S.; Mendoza-Porras, O.; Dellagnola, F.A.; Colgrave, M.L.; Vega, I.A. Integrative Proteomic Analysis of Digestive Tract Glycosidases from the Invasive Golden Apple Snail, Pomacea canaliculate. J. Proteome Res. 2019, 18, 3342–3352. [Google Scholar] [CrossRef] [PubMed]
- Zelck, U.E. Glycosidase activities in plasma of naive and schistosome-infected Biomphalaria glabrata (Gastropoda). Parasitology 1999, 119, 563–568. [Google Scholar] [CrossRef]
- Summers, M.D.; Smith, G.E. A manual of methods for baculovirus vectors and insect cell culture procedures. Tex. Agric. Exp. Stn. Bull. 1987, 1555. [Google Scholar]
- Peyer, C.; Bonay, P.; Staudacher, E. Purification and characterization of a β-xylosidase from potatoas (Solanum tuberosum). Biochim. Biophys. Acta 2004, 1672, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Geisslitz, S.; Islam, S.; Buck, L.; Grunwald-Gruber, C.; Sestili, F.; Camerlengo, F.; Masci, S.; D’Amico, S. Absolute and relative quantitation of amylase/trypsin-inhibitors by LC-MS/MS from wheat lines obtained by CRISPR-Cas9 and RNAi. Front. Plant Sci. 2022, 13, 974881. [Google Scholar] [CrossRef]
- Oomens, A.G.; Blissard, G.W. Requirement for GP64 to Drive Efficient Budding of Autographa californica Multicapsid Nucleopolyhedrovirus. Virology 1999, 254, 297–314. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, D.; Berger, P.; Schaffitzel, C.; Yamada, K.; Richmond, T.J.; Berger, I. Protein complex expression by using multigene baculoviral vectors. Nat. Methods 2006, 3, 1021–1032. [Google Scholar] [CrossRef]
- Gutternigg, M.; Bürgmayr, S.; Pöltl, G.; Rudolf, J.; Staudacher, E. Neutral N-glycan patterns of the gastropods Limax maximus, Cepaea hortensis, Planorbarius corneus, Arianta arbustorum and Achatina fulica. Glycoconj. J. 2007, 24, 475–489. [Google Scholar] [CrossRef]
- Anumula, K.R. Quantitative determination of monosaccharides in glycoproteins by high-performance liquid chromatography with highly sensitive fluorescence detection. Anal. Biochem. 1994, 220, 275–283. [Google Scholar] [CrossRef]
- Kubelka, V.; Altmann, F.; Staudacher, E.; Tretter, V.; März, L.; Hård, K.; Kamerling, J.P.; Vliegenthart, J.F.G. Primary structures of the N-linked carbohydrate chains from honeybee venom phospholipase A2. Eur. J. Biochem. 1993, 213, 1193–1204. [Google Scholar] [CrossRef] [PubMed]
- Gutternigg, M.; Ahrer, K.; Grabher-Meier, H.; Bürgmayr, S.; Staudacher, E. Neutral N-glycans of the gastropod Arion lusitanicus. Eur. J. Biochem. 2004, 271, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Benešová, E.; Šućur, Z.; Těšínský, M.; Spiwok, V.; Lipovová, P. Transglycosylation abilities of β-d-galactosidases from GH family 2. 3 Biotech 2021, 11, 168. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Guimarães, P.M.R.; Domingues, L. Recombinant microbial systems for improved β-galactosidase production and biotechnological applications. Biotechnol. Adv. 2011, 29, 600–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Got, R.; Marnay, A. Isolement, purification et quelques caractéristiques physicochimiques de deux beta-hexosidases du suc digestif d’Helix pomatia [Isolation, purification and some physicochemical characteristics of 2 beta-hexosidases of the digestive juice of Helix pomatia]. Eur. J. Biochem. 1968, 4, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Calvo, P.; Santamaria, M.G.; Melgar, M.J.; Cabezas, J.A. Kinetic evidence for two active sites in beta-D-fucosidase of Helicella ericetorum. Int. J. Biochem. 1983, 15, 685–693. [Google Scholar] [CrossRef]
- Colas, B. Kinetic studies on β-fucosidases of Achatina Balteata. Biochim. Biophys. Acta—Enzymol. 1979, 613, 448–458. [Google Scholar] [CrossRef]
- El-Gindy, A.; Ibrahim, Z.; Aziz, H. Improvement of Extracellular β-Galactosidase Production by Thermophilic Fungi Chaetomium thermophile and Thermomyces lanuginosus. Aust. J. Basic Appl. Sci. 2009, 3, 1925–1932. [Google Scholar]
- Alpers, D.H. Separation and Isolation of Rat and Human Intestinal β-Galactosidases. J. Biol. Chem. 1968, 244, 1238–1246. [Google Scholar] [CrossRef]
- Martinez-Bilbao, M.; Gaunt, M.T.; Huber, R.E. E461H-beta-galactosidase (Escherichia coli): Altered divalent metal specificity and slow but reversible metal inactivation. Biochemistry 1995, 34, 13437–13442. [Google Scholar] [CrossRef]
- Poltorak, O.M.; Chukhrai, E.S.; Pilipenko, O.S.; Atyaksheva, L.F.; Beregalov, A.E. A Comparative Study of the Structure and Properties of β-galactosidases. Biophys. Chem. 2007, 81, 808–812. [Google Scholar] [CrossRef]
- Tenu, J.-P.; Viratelle, O.M.; Yon, J. Kinetic Study of the Activation Process of β-Galactosidase from Escherichia coli by Mg2+. Eur. J. Biochem. 1972, 26, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Baccino, F.M.; Zuretti, M.F.; Pernigotti, L. Stimulation of Rat Liver β-Galactosidase Activity by Ions. Biochem. J. 1975, 151, 567–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, S.; Geueke, B.; Delgado, O.; Coleman, J.; Hatti-Kaul, R. Beta-galactosidase from a cold-adapted bacterium: Purification, characterization and application for lactose hydrolysis. Appl. Microbiol. Biotechnol. 2002, 58, 313–321. [Google Scholar] [CrossRef]
- Xavier, J.R.; Ramana, K.V.; Sharma, R.K. β-galactosidase: Biotechnological applications in food processing. J. Food Biochem. 2018, 42, e12564. [Google Scholar] [CrossRef]
- Kuby, S.A.; Lardy, H.A. Purification and Kinetics of b-D-Galactosidase from Escherichia coli, Strain K-12. J. Am. Chem. Soc. 1953, 75, 890–896. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Protein [mg] | Total Enzyme [U] | Specific Activity [U/mg] | Purification Fold | Yield [%] | |
|---|---|---|---|---|---|
| After precipitation | 72.74 | 2.82 | 0.04 | 1.00 | 100.00 |
| After HIC | 2.89 | 1.62 | 0.56 | 14.40 | 57.31 |
| After S200 | 0.63 | 0.53 | 0.84 | 21.76 | 18.83 |
| After Con A | 0.22 | 0.21 | 0.99 | 25.64 | 7.62 |
| After Gal-affinity chromatography | 0.04 | 0.10 | 2.44 | 62.83 | 3.51 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thoma, J.; Stenitzer, D.; Grabherr, R.; Staudacher, E. Identification, Characterization, and Expression of a β-Galactosidase from Arion Species (Mollusca). Biomolecules 2022, 12, 1578. https://doi.org/10.3390/biom12111578
Thoma J, Stenitzer D, Grabherr R, Staudacher E. Identification, Characterization, and Expression of a β-Galactosidase from Arion Species (Mollusca). Biomolecules. 2022; 12(11):1578. https://doi.org/10.3390/biom12111578
Chicago/Turabian StyleThoma, Julia, David Stenitzer, Reingard Grabherr, and Erika Staudacher. 2022. "Identification, Characterization, and Expression of a β-Galactosidase from Arion Species (Mollusca)" Biomolecules 12, no. 11: 1578. https://doi.org/10.3390/biom12111578