
Serratiopeptidase: An integrated View of Multifaceted Therapeutic Enzyme


Abstract
:1. Introduction
2. The Enzyme and Its Properties
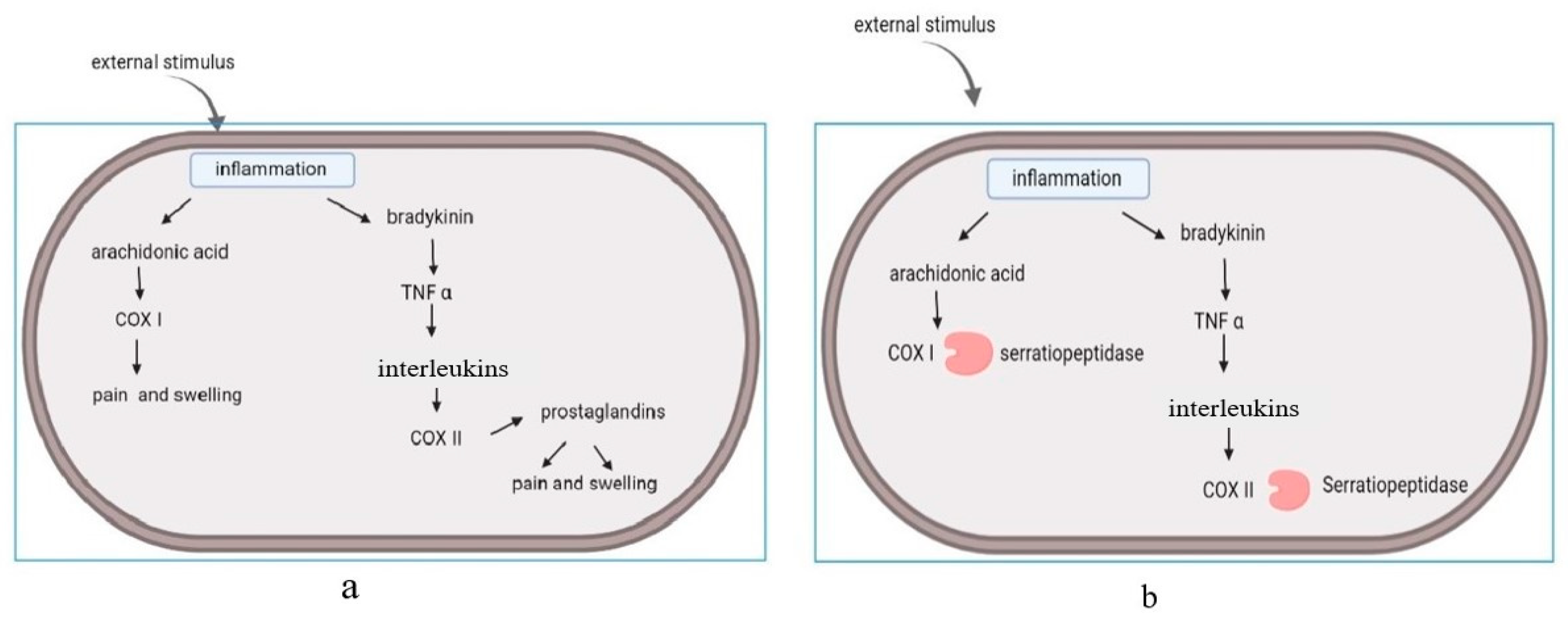
2.1. Anti-Inflammatory Action of Serratiopeptidase
2.2. Wound-Healing Activity of Serratiopeptidase
2.3. Antibiofilm Activity of Serratiopeptidase
2.4. Mucolytic Activity of Serratiopeptidase
2.5. Hemolytic Activity of Serratiopeptidase
2.6. Synergistic Property of Serratiopeptidase
3. Enzyme Production
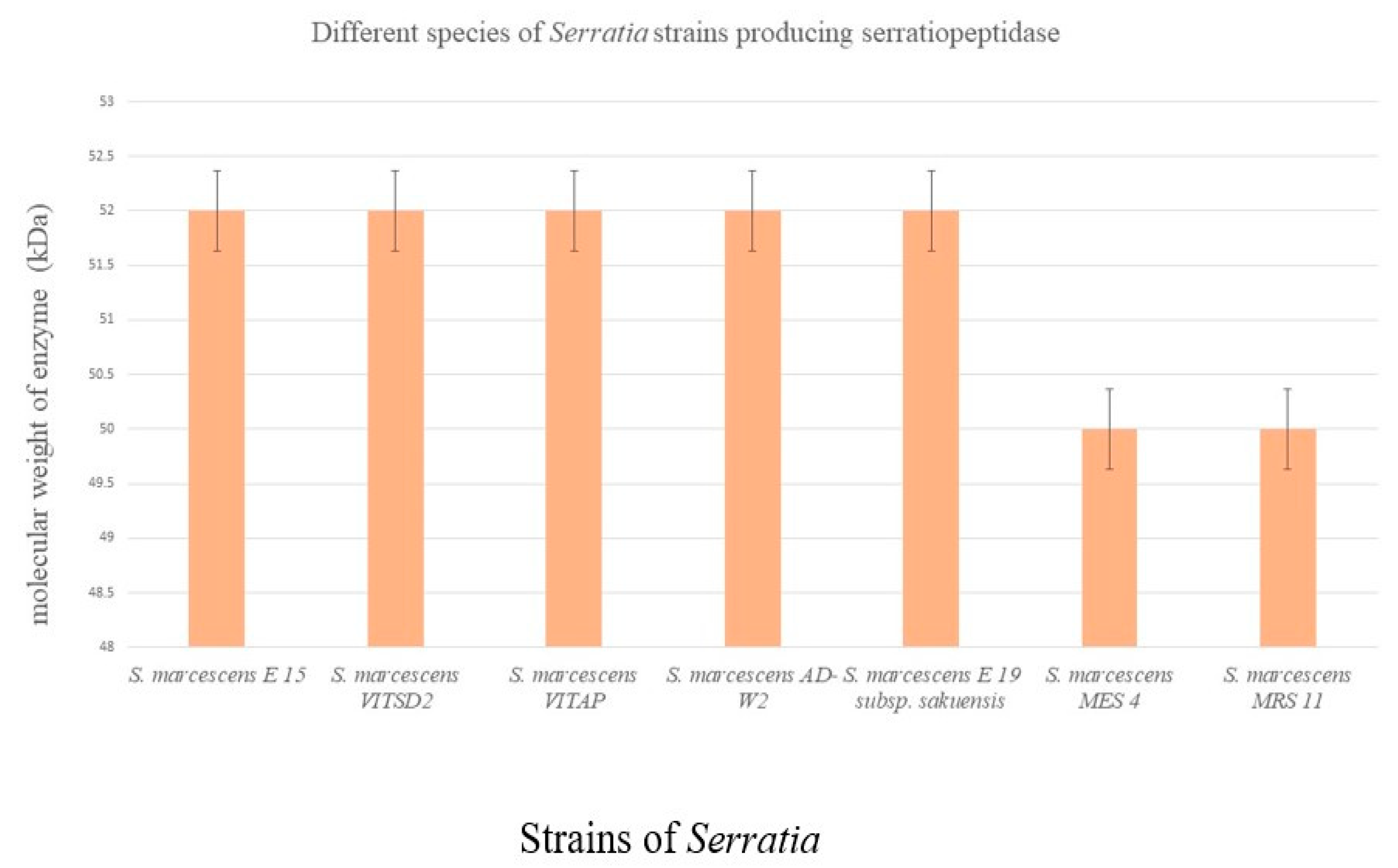
3.1. Serratia Marcescens
3.2. Alternative Species for Production
4. Analytical Approaches of Enzyme
5. Therapeutic Aspects of Serratiopeptidase
6. Clinical Significance
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Duve, C. The significance of lysosomes in pathology and medicine. Proc. Inst. Med. Chic. 1966, 26, 73–76. [Google Scholar] [PubMed]
- Tasaka, K.; Meshi, T.; Akagi, M.; Kakimoto, M.; Saito, R.; Okada, I.; Maki, K. Anti-Inflammatory Activity of a Proteolytic Enzyme, Prozime-10. Pharmacology 1980, 21, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Devi, C.S.; Elizabeth, J.R.; Saravanan, H.; Naine, S.J.; Srinivansan, V.M. Screening and molecular characterization of Serratia mar-cescens VITSD2: A strain producing optimum serratiopeptidase. Front. Biol. 2013, 8, 632–639. [Google Scholar] [CrossRef]
- Mohankumar, A.; Raj, R.K. Production and Characterization of Serratiopeptidase Enzyme from Serratia Marcescens. Int. J. Biol. 2011, 3, 39. [Google Scholar] [CrossRef]
- Steiger, S.; Harper, J.L. Mechanisms of Spontaneous Resolution of Acute Gouty Inflammation. Curr. Rheumatol. Rep. 2013, 16, 392. [Google Scholar] [CrossRef] [PubMed]
- Metkar, S.K.; Girigoswami, A.; Vijayashree, R.; Girigoswami, K. Attenuation of subcutaneous insulin induced amyloid mass in vivo using Lumbrokinase and Serratiopeptidase. Int. J. Biol. Macromol. 2020, 163, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Morihara, K. [24] Serralysin and related bacterial proteinases. Methods Enzymol. 1995, 248, 395–413. [Google Scholar] [CrossRef]
- Gupte, V.; Luthra, U. Analytical techniques for serratiopeptidase: A review. J. Pharm. Anal. 2017, 7, 203–207. [Google Scholar] [CrossRef]
- Bhagat, S.; Agarwal, M.; Roy, V. Serratiopeptidase: A systematic review of the existing evidence. Int. J. Surg. 2013, 11, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Suma, K.C.; Manasa, H.; Likhitha, A.; Nagamani, T.S. Isolation, Purification, and Characterization of Serratiopeptidase Enzyme from Serratia marcescens. Int. J. Innov. Sci. Res. Technol. 2020, 5, 156–161. [Google Scholar]
- Srivastava, S.; Singh, D.; Patel, S.; Singh, M.R. Treatment of rheumatoid arthritis by targeting macrophages through folic acid tailored superoxide dismutase and serratiopeptidase. J. Drug Deliv. Sci. Technol. 2017, 41, 431–435. [Google Scholar] [CrossRef]
- Teller, P.; White, T.K. The Physiology of Wound Healing: Injury Through Maturation. Perioper. Nurs. Clin. 2011, 6, 159–170. [Google Scholar] [CrossRef]
- Strodtbeck, F. Physiology of wound healing. Newborn Infant Nurs. Rev. 2001, 1, 43–52. [Google Scholar] [CrossRef]
- Hogan, S.; Zapotoczna, M.; Stevens, N.; Humphreys, H.; O’Gara, J.; O’Neill, E. Potential use of targeted enzymatic agents in the treatment of Staphylococcus aureus biofilm-related infections. J. Hosp. Infect. 2017, 96, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Rath, G.; Johal, E.S.; Goyal, A.K. Development of Serratiopeptidase and Metronidazole Based Alginate Microspheres for Wound Healing. Artif. Cells Blood Substit. Biotechnol. 2010, 39, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Mungantiwar, A.; Bhatt, N.; Shrivastava, P.; More, J.; Shaikh, R. A Randomized, Open-Label, Phase IV Clinical Study to Compare the Safety and Efficacy of the Fixed-Dose Combination of Trypsin, Bromelain, and Rutoside versus Serratiopeptidase in Minor Sur-gical Wound. IJRAMT. 2021, 2, 308–313. [Google Scholar]
- Selan, L.; Berlutti, F.; Passariello, C.; Comodi-Ballanti, M.R.; Thaller, M.C. Proteolytic enzymes: A new treatment strategy for prosthetic infections? Antimicrob. Agents Chemother. 1993, 37, 2618–2621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mecikoglu, M.; Saygi, B.; Yildirim, Y.; Karadag-Saygi, E.; Ramadan, S.S.; Esemenli, T. The effect of proteolytic enzyme serratiopep-tidase in the treatment of experimental implant-related infection. JBJS. 2006, 88, 1208–1214. [Google Scholar] [CrossRef]
- Selan, L.; Papa, R.; Tilotta, M.; Vrenna, G.; Carpentieri, A.; Amoresano, A.; Pucci, P.; Artini, M. Serratiopeptidase: A well-known metallo-protease with a new non-proteolytic activity against S. aureus biofilm. BMC Microbiol. 2015, 15, 207. [Google Scholar]
- Selan, L.; Artini, M.; Papa, R. Compounds from natural sources for new diagnostics and drugs against biofilm infections. In Microbial Biofilms—Importance and Applications; Dhanasekaran, D., Ed.; 2016; pp. 487–509. Available online: https://www.intechopen.com/chapters/50474 (accessed on 13 July 2016).
- Longhi, C.; Scoarughi, G.L.; Poggiali, F.; Cellini, A.; Carpentieri, A.; Seganti, L.; Pucci, P.; Amoresano, A.; Cocconcelli, P.S.; Artini, M.; et al. Protease treatment affects both invasion ability and biofilm formation in Listeria monocytogenes. Microb. Pathog. 2008, 45, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Artini, M.; Papa, R.; Scoarughi, G.L.; Galano, E.; Barbato, G.; Pucci, P.; Selan, L. Comparison of the action of different proteases on virulence properties related to the staphylococcal surface. J. Appl. Microbiol. 2012, 114, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M. The role of serratiopeptidase in the resolution of inflammation. Asian J. Pharm. Sci. 2017, 12, 209–215. [Google Scholar] [CrossRef]
- Gioia, M.; Ciaccio, C.; Calligari, P.; De Simone, G.; Sbardella, D.; Tundo, G.; Fasciglione, G.F.; Di Masi, A.; Di Pierro, D.; Bocedi, A.; et al. Role of proteolytic enzymes in the COVID-19 infection and promising therapeutic approaches. Biochem. Pharmacol. 2020, 182, 114225. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.K.; Rahman, A.; Kumar, A.; Gavel, P.; Asia, P. Adjuvant therapy with Serratiopeptidase and Vitamin D for COVID-19 patients: A new perspective. Int. J. Med. Sci. 2021, 4, 282–287. [Google Scholar]
- Kim, G.-U.; Kim, M.-J.; Ra, S.; Lee, J.; Bae, S.; Jung, J.; Kim, S.-H. Clinical characteristics of asymptomatic and symptomatic patients with mild COVID-19. Clin. Microbiol. Infect. 2020, 26, 948.e1–948.e3. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, F.I.; Morgan, R.C.; Dhawan, N.; Dinh, J.; Yatzkan, G.; Michel, G. Airway Hygiene in COVID-19 Pneumonia: Treatment Responses of 3 Critically Ill Cruise Ship Employees. Am. J. Case Rep. 2020, 21, e926596–e926596-8. [Google Scholar] [CrossRef]
- Sharma, C.; Jha, N.K.; Meeran, M.N.; Patil, C.R.; Goyal, S.N.; Ojha, S. Serratiopeptidase, a serine protease anti-inflammatory, fibrinolytic, and mucolytic drug can be a useful adjuvant for management in COVID-19. Front. Pharmacol. 2021, 12, 1188. [Google Scholar] [CrossRef]
- Connors, J.M.; Levy, J.H. COVID-19 and its implications for thrombosis and anticoagulation. Blood 2020, 135, 2033–2040. [Google Scholar] [CrossRef] [PubMed]
- Kase, Y.; Seo, H.; Oyama, Y.; Sakata, M.; Tomoda, K.; Takahama, K.; Hitoshi, T.; Okano, Y.; Miyata, T. A new method for evaluating mucolytic expectorant activity and its application. II. Application to two proteolytic enzymes, serratiopeptidase and seaprose. Arzneim.-Forsch. 1982, 32, 374–378. [Google Scholar]
- Kotb, E. Activity assessment of microbial fibrinolytic enzymes. Appl. Microbiol. Biotechnol. 2013, 97, 6647–6665. [Google Scholar] [CrossRef]
- Krishnamurthy, A.; Belur, P.D. A novel fibrinolytic serine metalloprotease from the marine Serratia marcescens subsp. sakuensis: Purification and characterization. Int. J. Biol. Macromol. 2018, 112, 110–118. [Google Scholar] [CrossRef]
- Shanks, R.M.Q.; Stella, N.A.; Lahr, R.M.; Wang, S.; Veverka, T.I.; Kowalski, R.P.; Liu, X. Serratamolide is a Hemolytic Factor Produced by Serratia marcescens. PLoS ONE 2012, 7, e36398. [Google Scholar] [CrossRef] [Green Version]
- Shanks, R.M.; Lahr, R.M.; Stella, N.A.; Arena, K.E.; Brothers, K.M.; Kwak, D.H.; Liu, X.; Kalivoda, E.J. A Serratia marcescens PigP homolog controls prodigiosin biosynthesis, swarming motility and hemolysis and is regulated by cAMP-CRP and HexS. PLoS ONE 2013, 8, e57634. [Google Scholar] [CrossRef] [Green Version]
- Wasserman, H.H.; Keggi, J.J.; McKeon, J.E. The structure of Serratamolide1-3. J. Am. Chem. Soc. 1962, 84, 2978–2982. [Google Scholar] [CrossRef]
- Maheshwari, M.; Miglani, G.; Mali, A.; Paradkar, A.; Yamamura, S.; Kadam, S. Development of tetracycline-serratiopeptidase-containing periodontal gel: Formulation and preliminary clinical study. AAPS PharmSciTech 2006, 7, E162–E171. [Google Scholar] [CrossRef]
- Alem, M.A.S.; Douglas, L.J. Effects of Aspirin and Other Nonsteroidal Anti-Inflammatory Drugs on Biofilms and Planktonic Cells of Candida albicans. Antimicrob. Agents Chemother. 2004, 48, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Thaller, M.; Selan, L.; Fiorani, P.; Passariello, C.; Rizzo, L.; Speziale, F. A comparative in vitro evaluation of different therapeutic protocols for vascular graft infections. Eur. J. Vasc. Endovasc. Surg. 1997, 14, 35–37. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.V.; Nirwane, A.M.; Belubbi, T.; Nagarsenker, M.S. Pulmonary delivery of synergistic combination of fluoroquinolone antibiotic complemented with proteolytic enzyme: A novel antimicrobial and antibiofilm strategy. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 2371–2384. [Google Scholar] [CrossRef] [PubMed]
- Anil, C.S.; Kashinath, M.A. Production, characterization & optimization of potent protease (serratiopeptidase) from Serratia mar-cescense 15. Int. J. Pharm. Res. Allied Sci. 2013, 3, 95–98. [Google Scholar]
- Acar, J.F. Serratia marcescens Infections. Infect. Control Hosp. Epidemiol. 1986, 7, 273–280. [Google Scholar] [CrossRef]
- Srivastava, V.; Mishra, S.; Chaudhuri, T.K. Enhanced production of recombinant serratiopeptidase in Escherichia coli and its charac-terization as a potential biosimilar to native biotherapeutic counterpart. Microb. Cell Factories 2019, 18, 215. [Google Scholar] [CrossRef]
- Gopinath, S.; Venkataprasad, R.; Rajnish, K.N.; Datta, S.; Selvarajan, E. Enhancement of serrapeptase hyper producing mutant by com-bined chemical and UV mutagenesis and its potential for fibrinolytic activity. J. Pure Appl. Microbiol. 2020, 14, 1295–1303. [Google Scholar] [CrossRef]
- Chander, D.; Khosla, J.K.; Koul, D.; Hossain, M.; Dar, M.J.; Chaubey, A. Purification and characterization of thermoactive serrati-opeptidase from Serratia marcescens AD-W2. AMB Express 2021, 11, 53. [Google Scholar] [CrossRef]
- Barman, S.; Bhattacharya, S.S.; Mandal, N.C. Serratia. Benef. Microbes Agro-Ecol. 2020, 3, 27–36. [Google Scholar]
- Koul, D.; Chander, D.; Manhas, R.S.; Chaubey, A. Isolation and Characterization of Serratiopeptidase Producing Bacteria from Mulberry Phyllosphere. Curr. Microbiol. 2020, 78, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Doshi, P.; Dantroliya, S.; Modi, A.; Shukla, A.; Patel, D.; Joshi, C.; Joshi, M. Enhanced Production Process of Recombinant Mature Serra-ti-opeptidase in Escherichia coli Using Fed-Batch Culture by Self-Proteolytic Activity of Fusion Protein. Fermentation 2022, 8, 307. [Google Scholar] [CrossRef]
- Kyoko, T.; Keiko, M.; Kayoko, S. Quality tests of high-molecular-weight substances by chromatography. Jpn. J. Relig. Stud. 1981, 6, 233–237. [Google Scholar]
- AAnanthakrishnan, B.; Ramesh, M.S. Muthuraman, Optimization studies in the production and purification of serratiopeptidase from Serratia marcescens UV mutant SM3. Int. J. Pharm. Pharm. Sci. 2013, 5, 748–752. [Google Scholar]
- El-Hawa, M. Abou, Purification and characterization of protease produced by Pseudomonas aeruginosa. Egypt. J. Microbiol. 1997, 31, 323–335. [Google Scholar]
- Salamone, P.R.; Wodzinski, R.J. Production, purification and characterization of a 50-kDa extracellular metalloprotease from Serratia marcescens. Appl. Microbiol. Biotechnol. 1997, 48, 317–324. [Google Scholar] [CrossRef]
- Louis, D.; Bernillon, J.; Païsse, J.O.; Wallach, J.M. Use of liquid chromatography-mass spectrometry coupling for monitoring the serra-lysin-catalyzed hydrolysis of a peptide library. J. Chromatogr. 1999, 932, 271–276. [Google Scholar] [CrossRef]
- Sandhya, K.V.; Devi, S.G.; Mathew, S.T. Quantitation of serrapeptase in formulations by UV method in the microplate format. Curr. Drug Deliv. 2008, 5, 303–305. [Google Scholar] [CrossRef] [PubMed]
- Pakhale, S.V.; Bhagwat, S.S. Purification of serratiopeptidase from Serratia marcescens NRRL B 23112 using ultrasound assisted three phase partitioning. Ultrason. Sonochemistry 2016, 31, 532–538. [Google Scholar] [CrossRef]
- Fuchs, R.L.; McPherson, S.A.; Drahos, D.J. Cloning of a Serratia marcescens Gene Encoding Chitinase. Appl. Environ. Microbiol. 1986, 51, 504–509. [Google Scholar] [CrossRef] [Green Version]
- Swamy, A.V.; Patil, P. Effect of some clinically used proteolytic enzymes on inflammation in rats. Indian J. Pharm. Sci. 2008, 70, 114–117. [Google Scholar] [CrossRef] [Green Version]
- Rajinikanth, B.; Venkatachalam, V.V.; Manavalan, R. Investigations on the potential of serratiopeptidase—A proteolytic enzyme, on acetic acid induced ulcerative colitis in mice. Int. J. Pharm. Pharm. Sci. 2014, 6, 525–531. [Google Scholar]
- Jadhav, S.B.; Shah, N.; Rathi, A.; Rathi, V.; Rathi, A. Serratiopeptidase: Insights into the therapeutic applications. Biotechnol. Rep. 2020, 28, e00544. [Google Scholar] [CrossRef]
- Al-Khateeb, T.; Nusair, Y. Effect of the proteolytic enzyme serrapeptase on swelling, pain and trismus after surgical extraction of mandibular third molars. Int. J. Oral Maxillofac. Surg. 2008, 37, 264–268. [Google Scholar] [CrossRef]
- Luthra, U.; Babu, P.; Patel, Y.; Ramesh, J.V.; Sharma, M.; Majeed, I.; Subbiah, S.K.; Pandiyan, R. Serratiopeptidase: A statistical approach towards enhancement of fermentation and biomass product recovery. Biomass-Convers. Biorefinery 2022, 1–8. [Google Scholar] [CrossRef]
- Jadav, S.P.; Patel, N.H.; Shah, T.G.; Gajera, M.V.; Trivedi, H.R.; Shah, B.K. Comparison of anti-inflammatory activity of serratiopeptidase and diclofenac in albino rats. J. Pharmacol. Pharmacother. 2010, 1, 116–117. [Google Scholar] [CrossRef]
- Ateia, Y.A.; Al-Edanni, M.S.; Al-Qurtas, M.I. Impact of metformin and serratiopeptidase in obese patients with knee osteoarthritis. Int. J. Pharm. Pharm. Sci. 2018, 10, 37. [Google Scholar] [CrossRef]
- Araghi, A.; Hashemi, S.; Sepahi, A.A.; Faramarzi, M.A.; Amin, M. Purification and study of anti-cancer effects of Serratia mar-cescens ser-ralysin. Iran J. Microbiol. 2019, 11, 320. [Google Scholar]
- Panthi, V.K.; Jha, S.K.; Chaubey, R.; Pangeni, R. Formulation and development of Serratiopeptidase enteric coated tablets and analytical method validation by UV Spectroscopy. Int. J. Anal. Chem. 2021, 2021, 9749474. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.H.; Paradkar, A. Cubic liquid crystalline glyceryl monooleate matrices for oral delivery of enzyme. Int. J. Pharm. 2005, 294, 161–171. [Google Scholar] [CrossRef]
- Chappi, D.M.; Suresh, K.V.; Patil, M.R.; Desai, R.; Tauro, D.P.; Bharani, K.N.S.S.; Parkar, M.I.; Babaji, H.V. Comparison of clinical efficacy of methylprednisolone and serratiopeptidase for reduction of postoperative sequelae after lower third molar surgery. J. Clin. Exp. Dent. 2015, 7, e197–e202. [Google Scholar] [CrossRef] [Green Version]
- Chanalia, P.; Gandhi, D.; Jodha, D.; Singh, J. Applications of microbial proteases in pharmaceutical industry: An overview. RRMM. 2011, 22, 96–101. [Google Scholar] [CrossRef]
- Shi, J.; Kantoff, P.W.; Wooster, R.; Farokhzad, O.C. Cancer nanomedicine: Progress, challenges and opportunities. Nat. Rev. Cancer 2017, 17, 20–37. [Google Scholar] [CrossRef]
- Jaiswal, S.; Mishra, P. Co-delivery of curcumin and serratiopeptidase in HeLa and MCF-7 cells through nanoparticles show improved anti-cancer activity. Mater. Sci. Eng. C 2018, 92, 673–684. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sl No | Antibiotics | Effect of Enzymes | References |
|---|---|---|---|
| 1. | Ofloxacin | Enhanced the activity of ofloxacin and inhibited biofilm formation. | [17] |
| 2. | Azithromycin | Effective against different strains of biofilm forming Staphylococcus sp. | [38] |
| 3. | Levofloxacin | Eradicated > 90% of the preformed biofilm. | [39] |
| 4. | Vancomycin and rifampicin | Effective in dispersing most of the biofilm forming bacteria | [14] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nair, S.R.; C, S.D. Serratiopeptidase: An integrated View of Multifaceted Therapeutic Enzyme. Biomolecules 2022, 12, 1468. https://doi.org/10.3390/biom12101468
Nair SR, C SD. Serratiopeptidase: An integrated View of Multifaceted Therapeutic Enzyme. Biomolecules. 2022; 12(10):1468. https://doi.org/10.3390/biom12101468
Chicago/Turabian StyleNair, Sreelakshmi R., and Subathra Devi. C. 2022. "Serratiopeptidase: An integrated View of Multifaceted Therapeutic Enzyme" Biomolecules 12, no. 10: 1468. https://doi.org/10.3390/biom12101468