
Novel Anti-Fungal d-Laminaripentaose-Releasing Endo-β-1,3-glucanase with a RICIN-like Domain from Cellulosimicrobium funkei HY-13

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cloning of the Endo-β-1,3-glucanase (GluY) Gene
2.3. Production and Purification of Recombinant Proteins
2.4. Analysis of Proteins
2.5. Enzyme Assays
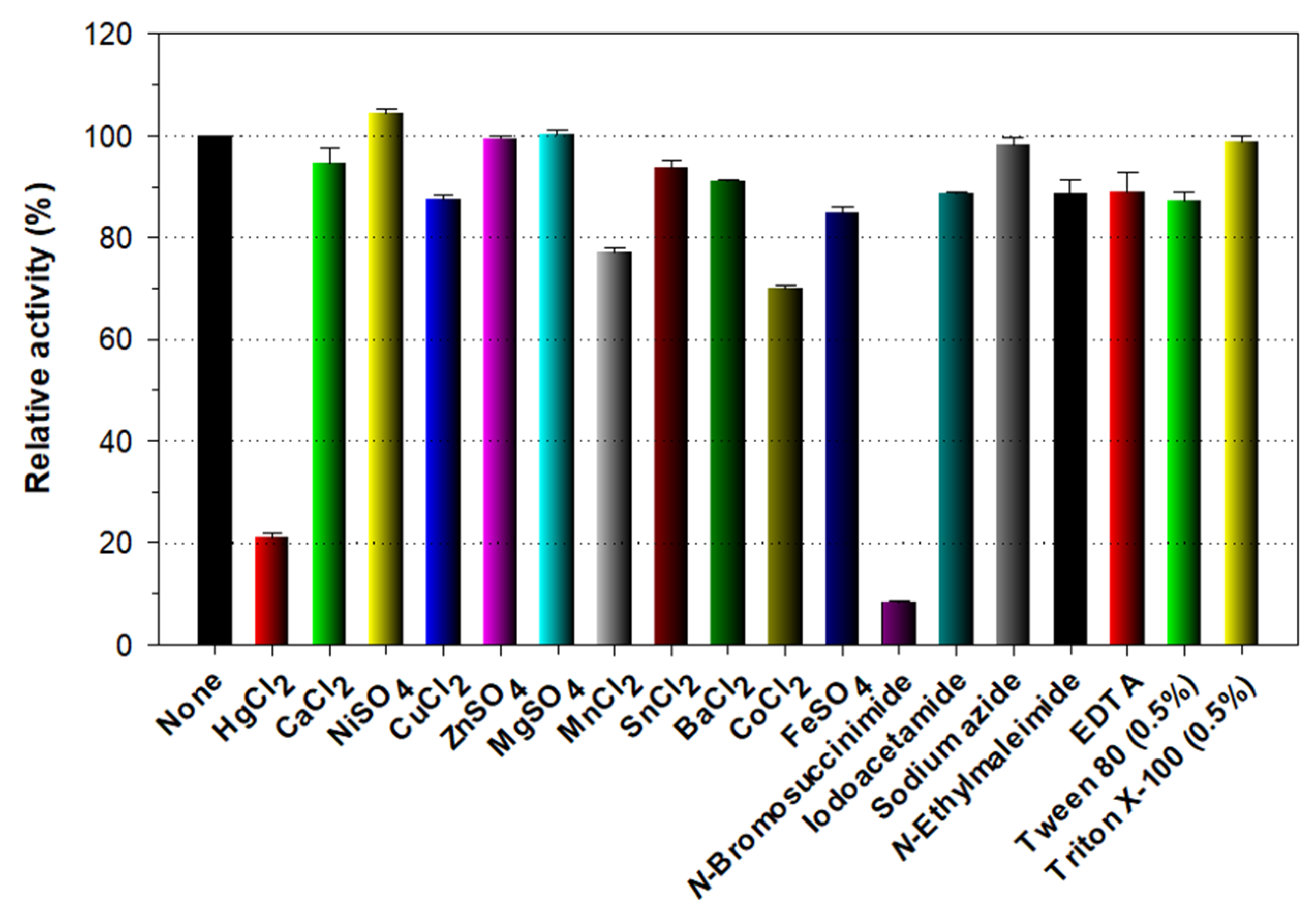
2.6. Effects of pH, Temperature, and Chemicals on the Endo-β-1,3-glucanase Activity
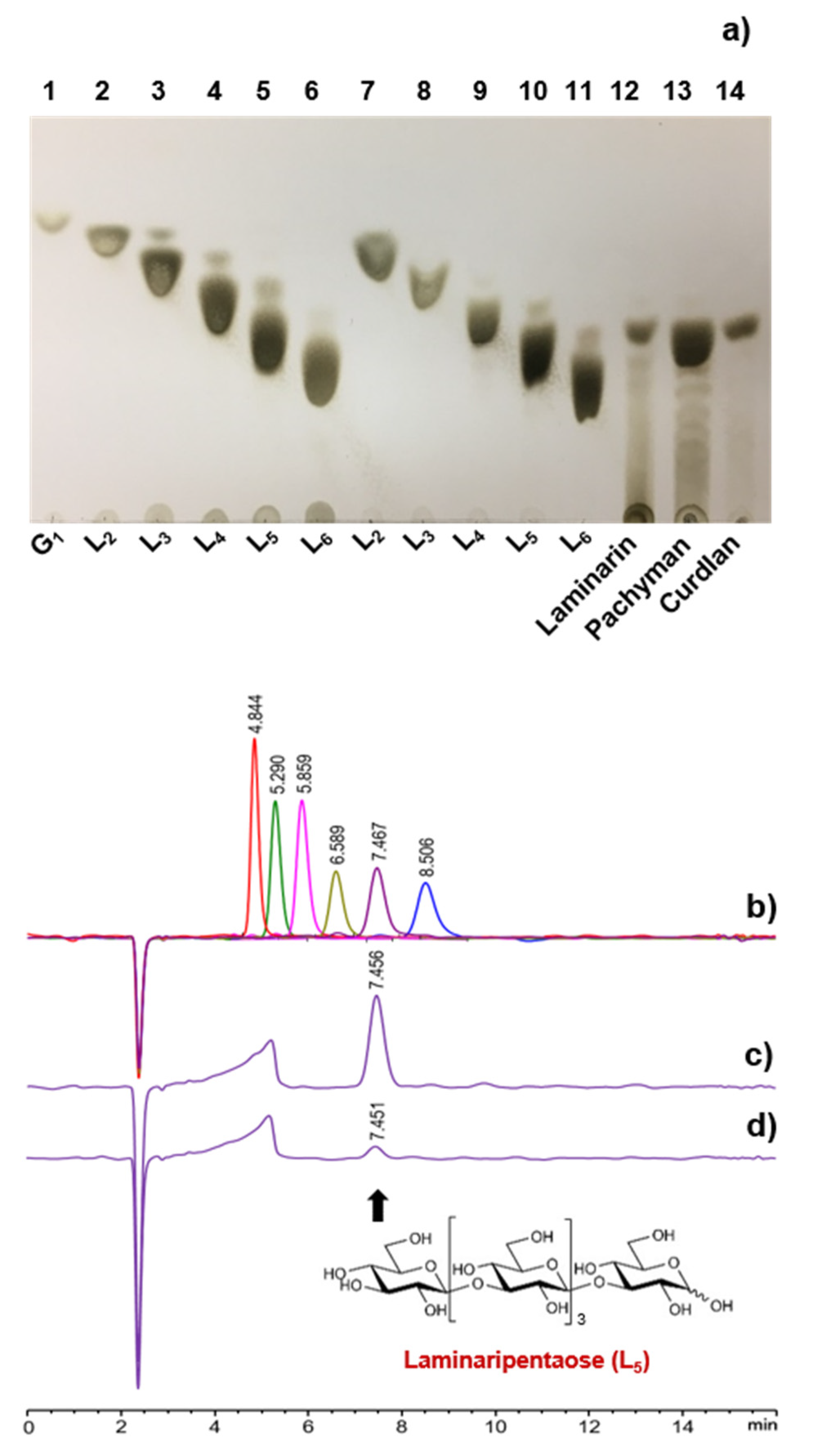
2.7. Analysis of the Degradation Products
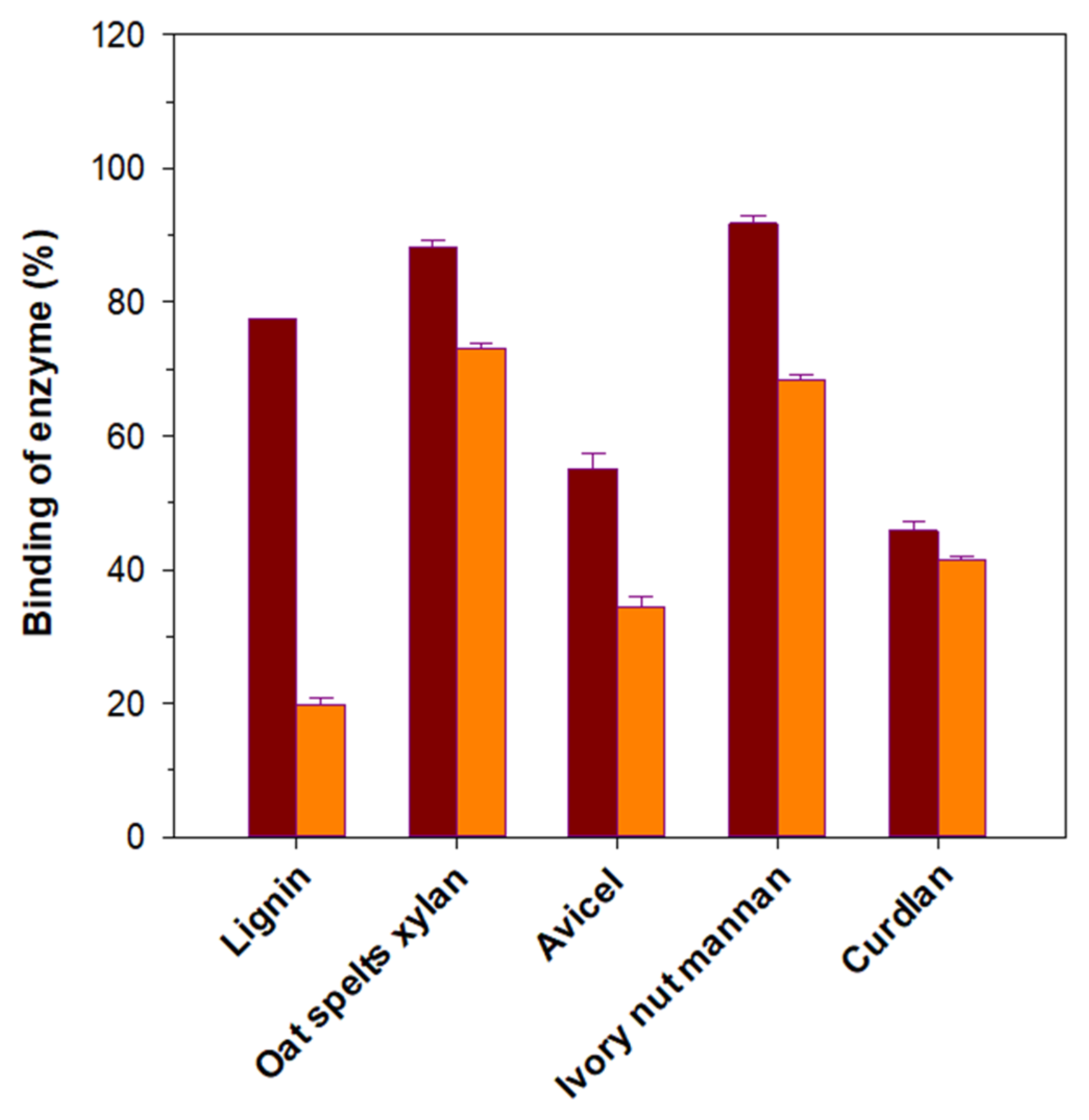
2.8. Binding Assay
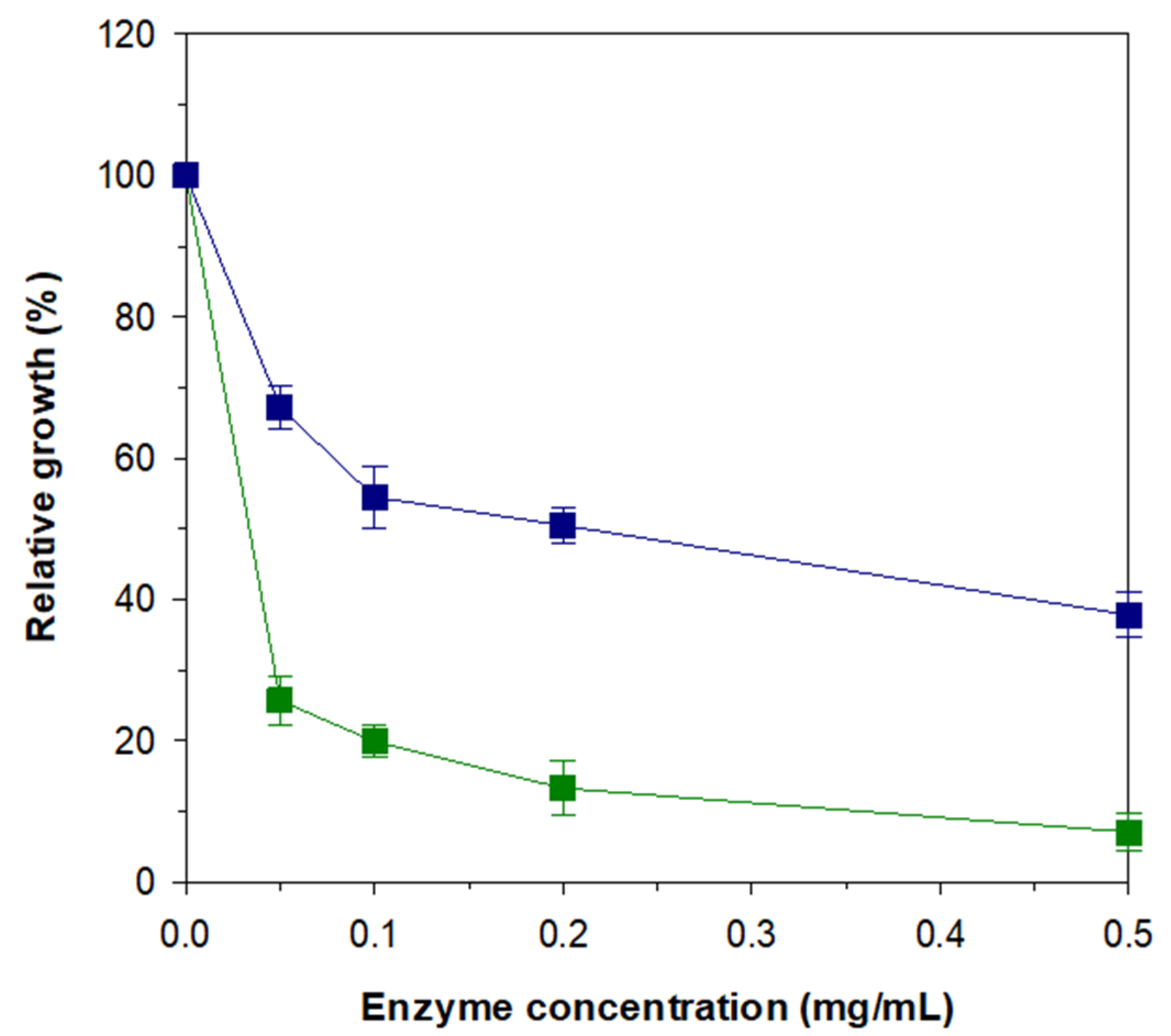
2.9. Anti-Fungal Assay
3. Results and Discussion
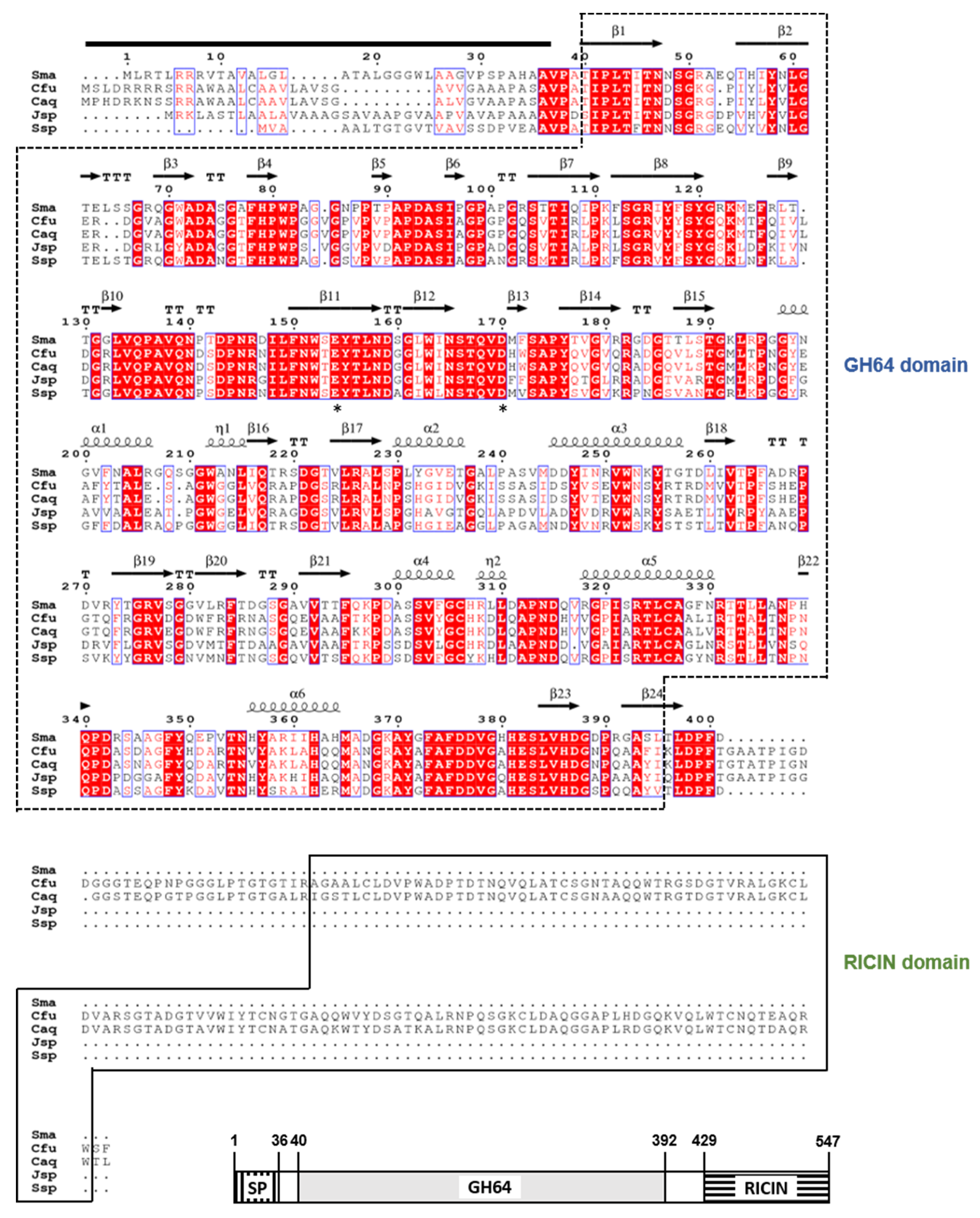
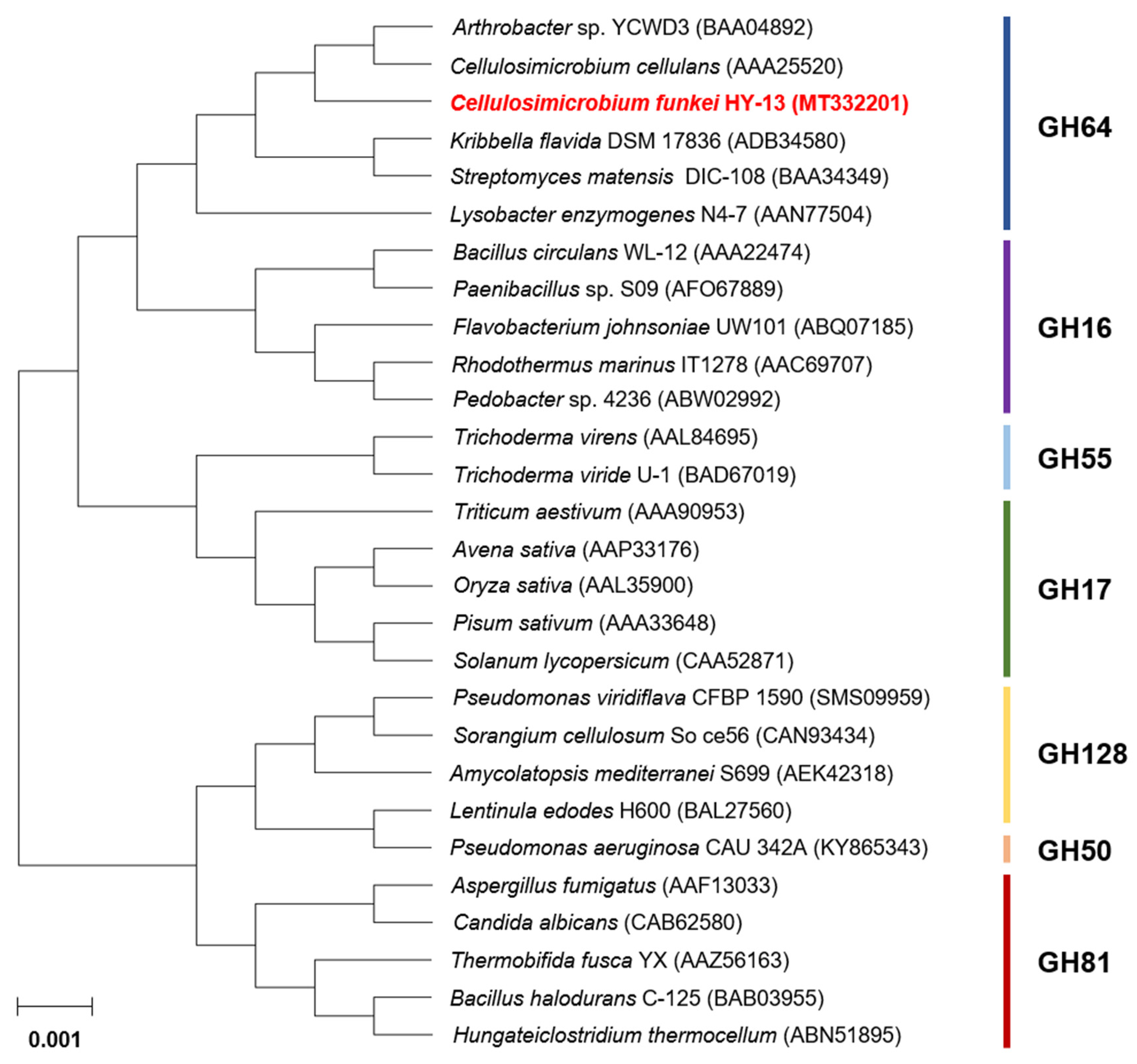
3.1. Molecular Characterization of the GH64 Endo-β-1,3-glucanase Gene
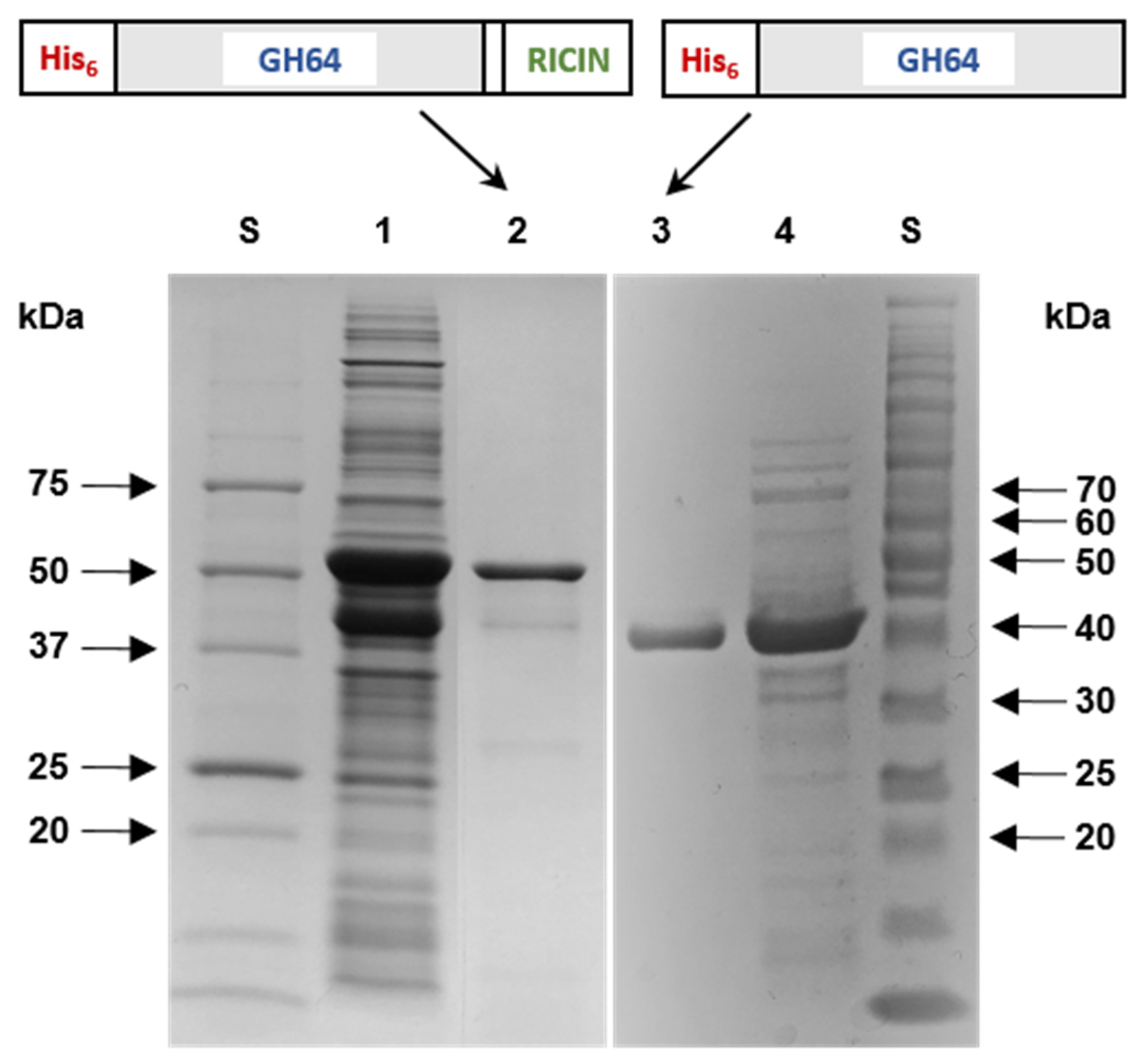
3.2. Purification and SDS-PAGE Analysis of Recombinant Enzymes
3.3. Biocatalytic Characterization of Recombinant Enzymes
3.4. Substrate Specificity
3.5. Binding Affinity of rGluY and rGluYΔRICIN to Insoluble Substrates
3.6. Anti-Fungal Activities of rGluY and rGluYΔRICIN
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barsanti, L.; Passarelli, V.; Evangelista, V.; Frassanito, A.M.; Gualtieri, P. Chemistry, physico-chemistry and applications linked to biological activities of β-glucans. Nat. Prod. Rep. 2011, 28, 457–466. [Google Scholar] [CrossRef]
- Wu, D.; Li, A.; Ma, F.; Yang, J.; Xie, Y. Genetic control and regulatory mechanisms of succinoglycan and curdlan biosynthesis in genus Agrobacterium. Appl. Microbiol. Biotechnol. 2016, 100, 6183–6192. [Google Scholar] [CrossRef]
- Zhang, R.; Edgar, K.J. Properties, chemistry, and applications of the bioactive polysaccharide curdlan. Biomacromolecules 2014, 15, 1079–1096. [Google Scholar] [CrossRef]
- Linton, S.M. Review: The structure and function of cellulase (endo-β-1,4-glucanase) and hemicellulase (β-1,3-glucanase and endo-β-1,4-mannanase) enzymes in invertebrates that consume materials ranging from microbes, algae to leaf litter. Comp. Biochem. Physiol. B 2020, 240, 110354. [Google Scholar] [CrossRef]
- Yi, P.; Yan, Q.; Jiang, Z.; Wang, L. A first glycoside hydrolase family 50 endo-β-1,3-d-glucanase from Pseudomonas aeruginosa. Enzyme Microb. Technol. 2018, 108, 34–41. [Google Scholar] [CrossRef]
- Shrestha, K.L.; Liu, S.-W.; Huang, C.-P.; Wu, H.-M.; Wang, W.-C.; Li, Y.K. Characterization and identification of essential residues of the glycoside hydrolase family 64 laminaripentaose-producing-β-1,3-glucanase. Protein Eng. Des. Sel. 2011, 24, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.-M.; Hong, T.-Y.; Liu, C.-C.; Meng, M. Cloning and functional characterization of a complex endo-β-1,3-glucanase from Paenibacillus sp. Appl. Microbiol. Biotechnol. 2009, 81, 1051–1061. [Google Scholar] [CrossRef] [Green Version]
- Woo, J.-B.; Kang, H.-N.; Woo, E.-J.; Lee, S.-B. Molecular cloning and functional characterization of an endo-1,3-β-glucanase from Streptomyces matensis ATCC 23935. Food Chem. 2014, 148, 184–187. [Google Scholar] [CrossRef]
- Ryan, E.M.; Ward, O.P. Study of the effect of β-1,3-glucanase from Basidiomycete QM 806 on yeast extract production. Biotechnol. Lett. 1985, 7, 409–412. [Google Scholar] [CrossRef]
- Vetvicka, V. Glucan-immunostimulant, adjuvant, potential drug. World J. Clin. Oncol. 2011, 2, 115–119. [Google Scholar] [CrossRef]
- Ferrer, P. Revisiting the Cellulosimicrobium cellulans yeast-lytic β-1,3-glucanases toolbox: A review. Microb. Cell Fact. 2006, 5, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gacto, M.; Vincente-Soler, J.; Cansado, J.; Villa, T.G. Characterization of an extracellular enzyme system produced by Micromonospora chalcea with lytic activity on yeast cells. J. Appl. Microbiol. 2000, 88, 961–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brune, A. Symbiotic digestion of lignocellulose in termite guts. Nat. Rev. Microbiol. 2014, 12, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Warnecke, F.; Luginbühl, P.; Ivanova, N.; Ghassemian, M.; Richardson, T.H.; Stege, J.T.; Cayouette, M.; McHardy, A.C.; Djordjevic, G.; Aboushadi, N.; et al. Metagenomic and functional analysis of hindgut microbiota of a wood-feeding higher termite. Nature 2007, 450, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Scully, E.D.; Geib, S.M.; Hoover, K.; Tien, M.; Tringe, S.G.; Barry, K.W.; Glavina del Rio, T.; Chovatia, M.; Herr, J.R.; Carlson, J.E. Metagenomic profiling reveals lignocellulose degrading system in a microbial community associated with a wood-feeding beetle. PLoS ONE 2013, 8, e073827. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.Y.; Han, M.K.; Park, D.-S.; Lee, J.S.; Oh, H.-W.; Shin, D.-H.; Jeong, T.-S.; Kim, S.U.; Bae, K.S.; Son, K.-H.; et al. Novel GH10 xylanase, with a fibronectin type 3 domain, from Cellulosimicrobium sp. strain HY-13, a bacterium in the gut of Eisenia fetida. Appl. Environ. Microbiol. 2009, 75, 7275–7279. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.Y.; Ham, S.-J.; Lee, H.J.; Cho, H.-Y.; Kim, J.-H.; Kim, Y.-J.; Shin, D.-H.; Rhee, Y.H.; Son, K.-H.; Park, H.-Y. Cloning and characterization of a modular GH5 β-1,4-mannanase with high specific activity from the fibrolytic bacterium Cellulosimicrobium sp. strain HY-13. Bioresour. Technol. 2011, 102, 9185–9192. [Google Scholar] [CrossRef]
- Kim, D.Y.; Lee, M.J.; Cho, H.-Y.; Lee, J.S.; Lee, M.-H.; Chung, C.W.; Shin, D.-H.; Rhee, Y.H.; Son, K.-H.; Park, H.-Y. Genetic and functional characterization of an extracellular modular GH6 endo-β-1,4-glucanase from an earthworm symbiont, Cellulosimicrobium funkei HY-13. Antonie Leeuwenhoek 2016, 109, 1–12. [Google Scholar] [CrossRef]
- Kim, D.Y.; Kim, J.; Lee, S.H.; Chung, C.; Shin, D.-H.; Ku, B.-H.; Son, K.-H.; Park, H.-Y. A d-glucose- and d-xylose-tolerant GH1 β-glucosidase from Cellulosimicrobium funkei HY-13, a fibrolytic gut bacterium of Eisenia fetida. Proc. Biochem. 2020, 94, 282–288. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Kim, D.Y.; Lee, S.H.; Lee, M.J.; Cho, H.-Y.; Lee, J.S.; Rhee, Y.H.; Shin, D.-H.; Son, K.-H.; Park, H.-Y. Genetic and functional characterization of a novel GH10 endo-β-1,4-xylanase with a ricin-type β-trefoil domain-like domain from Luteimicrobium xylanilyticum HY-24. Int. J. Biol. Macromol. 2018, 106, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Doi, A. Cloning and expression in Escherichia coli of the gene for an Arthrobacter β-(1→3)-glucanase. J. Bacteriol. 1986, 168, 1272–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, S.-H.; Chrétien, P.; Bastien, L.; Slilaty, S.N. Primary sequence of the glucanase gene from Oerskovia xanthineolytica. J. Biol. Chem. 1991, 266, 1058–1063. [Google Scholar] [CrossRef]
- Nakabayashi, M.; Nishijima, T.; Ehara, G.; Nikaidou, N.; Nishihashi, H.; Watanabe, T. Structure of the gene encoding laminaripentaose-producing β-1,3-glucanase (LPHase) of Streptomyces matensis DIC-108. J. Ferment. Bioeng. 1998, 85, 459–464. [Google Scholar] [CrossRef]
- Palumbo, J.D.; Sullivan, R.F.; Kobayashi, D.Y. Molecular characterization and expression in Escherichia coli of three β-1,3-glucanase genes from Lysobacter enzymogenes strain N4-7. J. Bacteriol. 2003, 185, 4362–4370. [Google Scholar] [CrossRef] [Green Version]
- Kumagai, Y.; Okuyama, M.; Kimura, A. Heat treatment of curdlan enhances the enzymatic production of biologically active β-(1,3)-glucan oligosaccharides. Carbohyd. Polym. 2016, 146, 396–401. [Google Scholar] [CrossRef]
- Kobayashi, T.; Uchimura, K.; Kubota, T.; Nunoura, T.; Deguchi, S. Biochemical and genetic characterization of β-1,3-glucanase from a deep subseafloor Laceyella putida. Appl. Microbiol. Biotechnol. 2016, 100, 203–214. [Google Scholar] [CrossRef]
- Roberge, M.; Shareck, F.; Morosoli, R.; Kluepfel, D.; Dupont, C. Characterization of active-site aromatic residues in xylanase A from Streptomyces lividans. Protein Eng. 1999, 12, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Zolotnitsky, G.; Cogan, U.; Adir, N.; Solomon, V.; Shoham, G.; Shoham, Y. Mapping glycoside hydrolase substrate subsites by isothermal titration calorimetry. Proc. Natl. Acad. Sci. USA 2004, 101, 11275–11280. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, B.; Srivastava, S.S.; Sankeshi, V.; Garg, R.; Srivastava, S.; Sankaranarayanan, R.; Sharma, Y. βγ-Crystallination endows a novel bacterial glycoside hydrolase 64 with Ca2+-dependent activity modulation. J. Bacteriol. 2019, 201, e00392-19. [Google Scholar] [CrossRef]
- Hong, T.-Y.; Meng, M. Biochemical characterization and antifungal activity of an endo-1,3-β-glucanase of Paenibacillus sp. isolated from garden soil. Appl. Microbiol. Biotechnol. 2003, 61, 472–478. [Google Scholar] [CrossRef]
- Hong, T.-Y.; Cheng, C.-W.; Huang, J.-W.; Meng, M. Isolation and biochemical characterization of an endo-1,3-β-glucanase from Streptomyces sioyaensis containing a C-terminal family 6 carbohydrate-binding module that binds to 1,3-β-glucan. Microbiology 2002, 148, 1151–1159. [Google Scholar] [CrossRef] [Green Version]
- Jaafar, N.R.; Khoiri, N.M.; Ismail, N.F.; Mahmood, N.A.N.; Murad, A.M.A.; Bakar, F.D.A.; Yajit, N.L.M.; Illias, R.M. Functional characterization and product specificity of endo-β-1,3-glucanase from alkalophilic bacterium, Bacillus lehensis G1. Enzyme Microb. Technol. 2020, 140, 109625. [Google Scholar] [CrossRef] [PubMed]
- Woo, C.-B.; Kang, H.-N.; Lee, S.-B. Molecular cloning and anti-fungal effect of endo-1,3-β-glucanase from Thermotoga marimata. Food Sci. Biotechnol. 2014, 23, 1243–1246. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Enzyme | Mr (kDa) | Optimum pH | Optimum Temp. (°C) | Specific Activity (U/mg) | Reference |
|---|---|---|---|---|---|---|
| Cellulosimicrobium funkei HY-13 | rGluY | 57.5 | 4.5 | 40 | 26.0 a, 18.8 b, 2.3 c | This study |
| Cellulosimicrobium funkei HY-13 | rGluYΔRICIN | 42.0 | 5.5 | 45 | 9.8 a, 5.4 b, 0.4 c | This study |
| Streptomyces matensis DIC-108 | LPHase | 39.4 | 7.5–8.5 | 55 | 4.2 a, 7.1 c | [6] |
| Lysobacter enzymogenes N4-7 | GluB | 41.0 | 4.5–5.0 | 41 | 2.1 a, 0.6 b, 1.2 c | [25] |
| Streptomyces matensis ATCC 23935 | SmβG | 43.0 | 6.0 | 55 | NI d | [8] |
| Arthrobacter sp. strain YCWD3 | endo-β-1,3-glucanase | 55.0 | 5.8 | NI | 11–13 b | [22] |
| Kribbella flavida NBRC 14399 | KfGH64 | 43.0 | 5.5 | 45 | NI | [26] |
| Substrate | Specific Activity (U/mg) a of rGluY | Specific Activity (U/mg) of rGluYΔRICIN |
|---|---|---|
| Laminarin | 26.0 ± 1.2 | 9.8 ± 0.1 |
| Pachyman | 18.8 ± 1.4 | 5.4 ± 0.1 |
| Curdlan | 2.3 ± 0.2 | 0.4 ± 0.1 |
| Carboxymethylcellulose | ND b | ND |
| Barley β-1,3-1,4-d-glucan | ND | ND |
| Beechwood xylan | ND | ND |
| Locust bean gum | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, L.; Kim, J.; Son, K.-H.; Shin, D.-H.; Ku, B.-H.; Kim, D.Y.; Park, H.-Y. Novel Anti-Fungal d-Laminaripentaose-Releasing Endo-β-1,3-glucanase with a RICIN-like Domain from Cellulosimicrobium funkei HY-13. Biomolecules 2021, 11, 1080. https://doi.org/10.3390/biom11081080
Bai L, Kim J, Son K-H, Shin D-H, Ku B-H, Kim DY, Park H-Y. Novel Anti-Fungal d-Laminaripentaose-Releasing Endo-β-1,3-glucanase with a RICIN-like Domain from Cellulosimicrobium funkei HY-13. Biomolecules. 2021; 11(8):1080. https://doi.org/10.3390/biom11081080
Chicago/Turabian StyleBai, Lu, Jonghoon Kim, Kwang-Hee Son, Dong-Ha Shin, Bon-Hwan Ku, Do Young Kim, and Ho-Yong Park. 2021. "Novel Anti-Fungal d-Laminaripentaose-Releasing Endo-β-1,3-glucanase with a RICIN-like Domain from Cellulosimicrobium funkei HY-13" Biomolecules 11, no. 8: 1080. https://doi.org/10.3390/biom11081080