
Identification and Characterization of a Novel, Cold-Adapted d-Xylobiose- and d-Xylose-Releasing Endo-β-1,4-Xylanase from an Antarctic Soil Bacterium, Duganella sp. PAMC 27433

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Isolation and Identification of a Xylanolytic Microorganism
2.3. Molecular Cloning of the Endo-β-1,4-Xylanase (XylR) Gene
2.4. Overproduction and Purification of Recombinant Proteins
2.5. Analysis of Proteins
2.6. Enzyme Assays
2.7. Effects of pH, Temperature, and Chemicals on the rXylR Activity
2.8. Analysis of the Degradation Products
2.9. Binding Assay
3. Results and Discussion
3.1. Identification of a Xylanolytic Antarctic Isolate
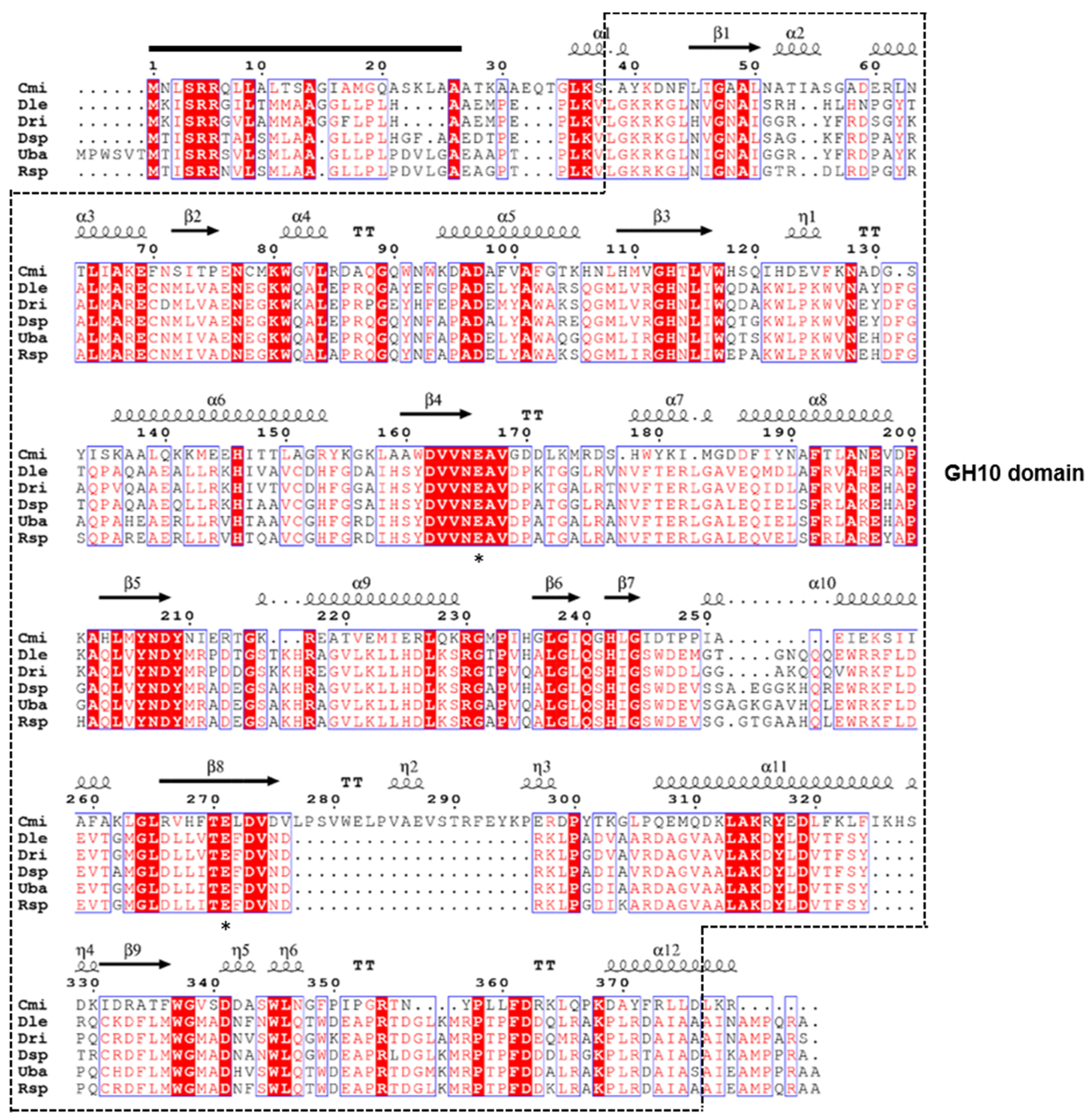
3.2. Molecular Characterization of the GH10 Endo-β-1,4-Xylanase Gene
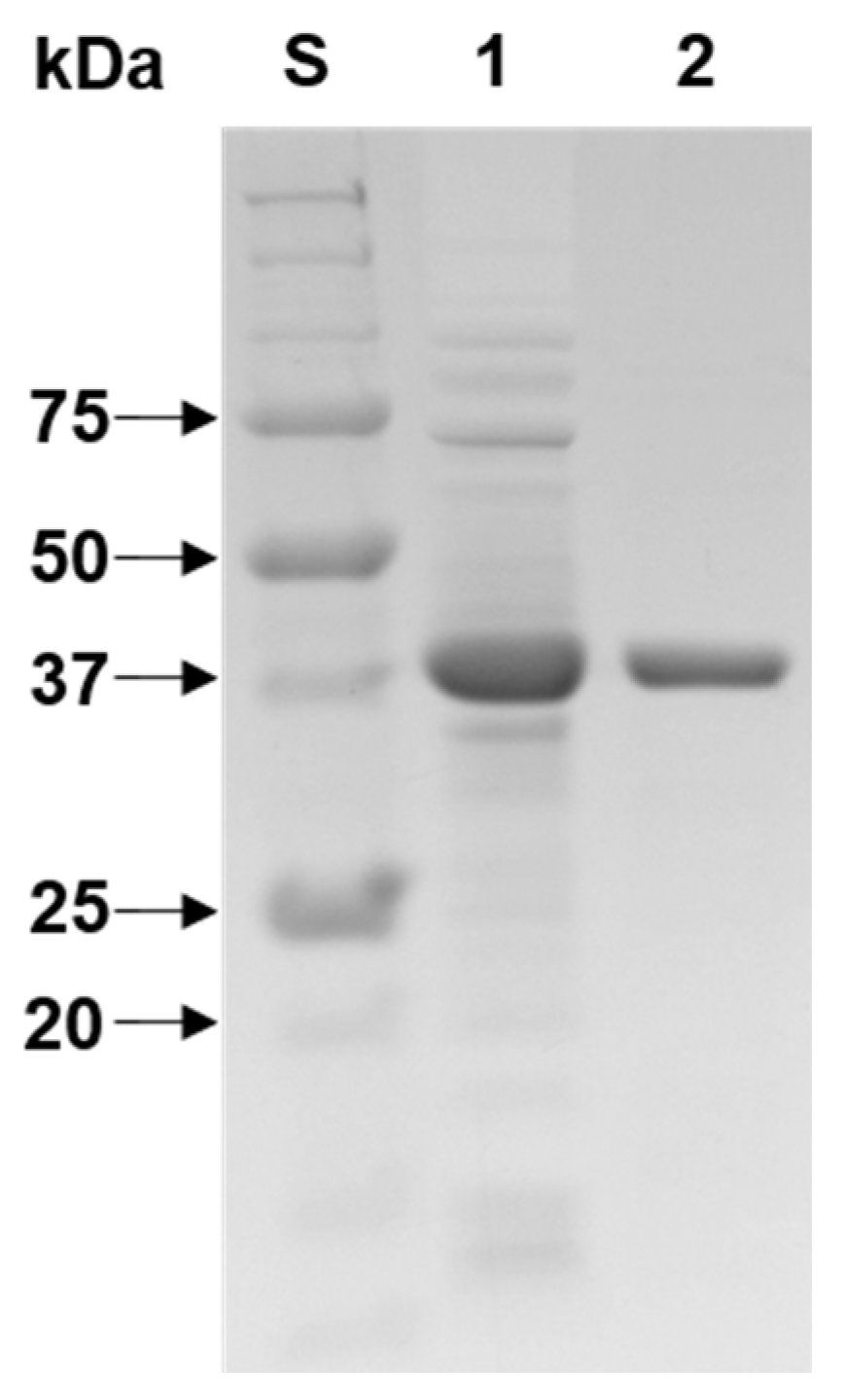
3.3. Purification and SDS-PAGE Analysis of rXylR Proteins
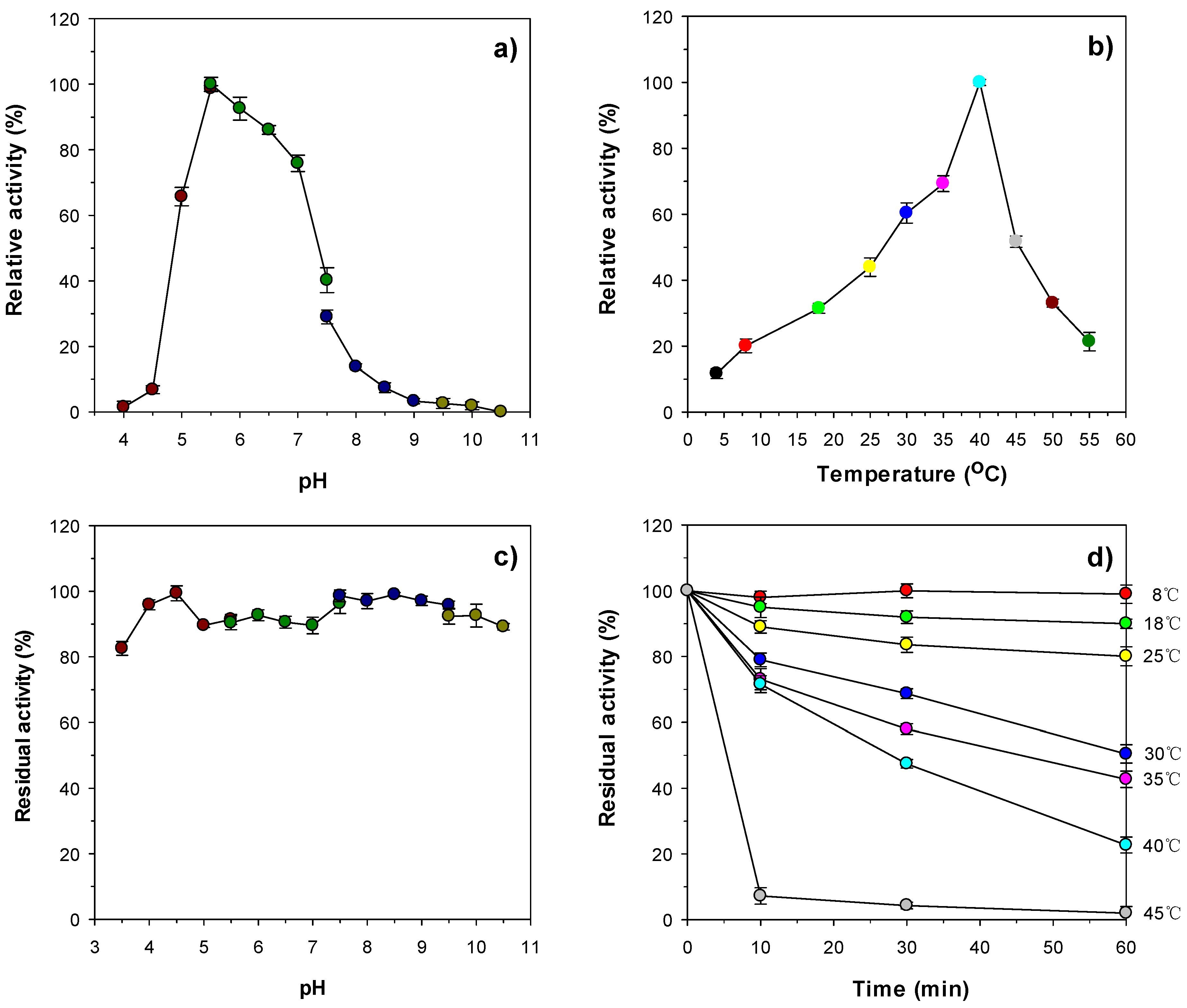
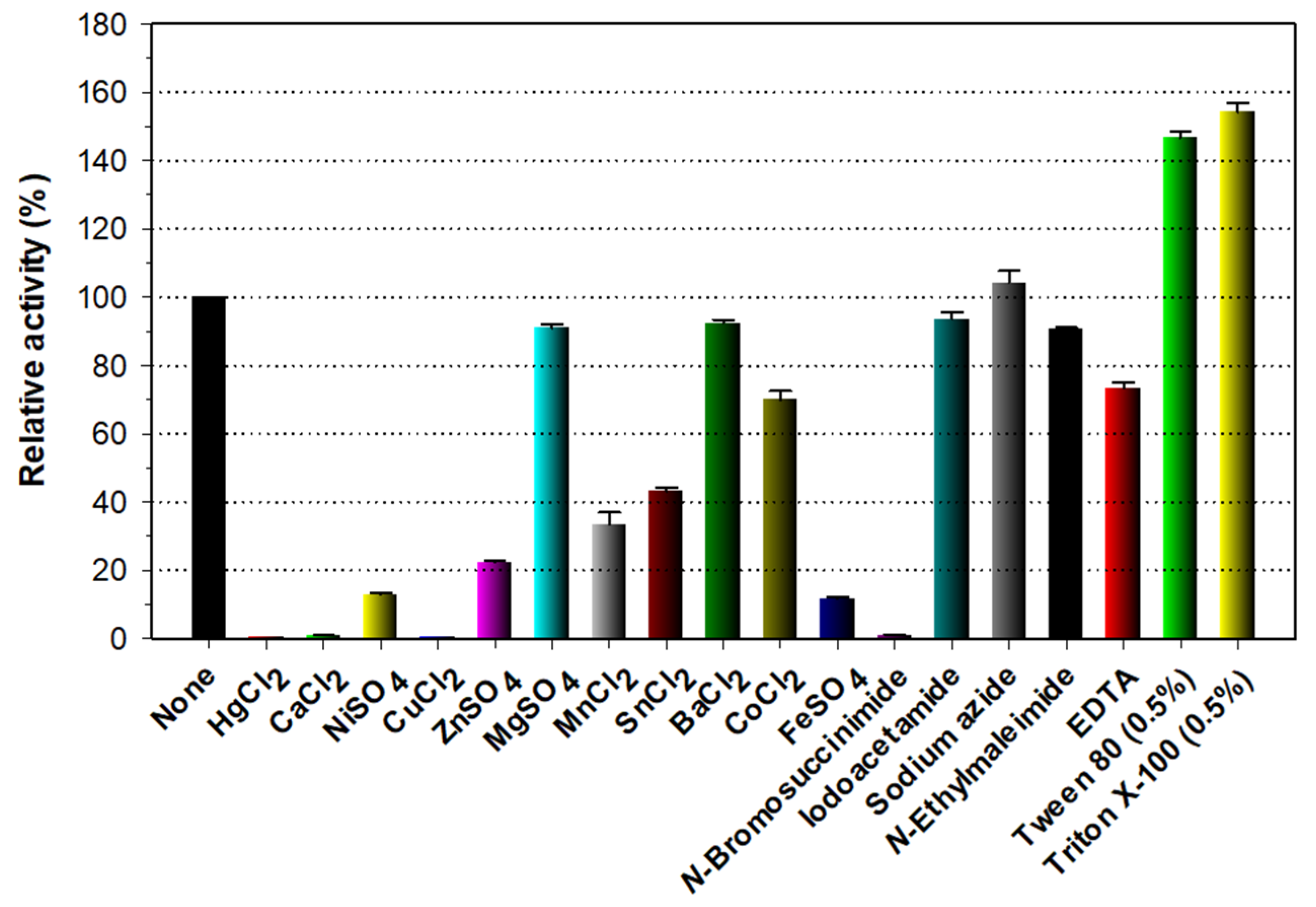
3.4. Biochemical Characterization of Recombinant Enzymes
3.5. Substrate Specificity and Hydrolytic Properties of rXylR
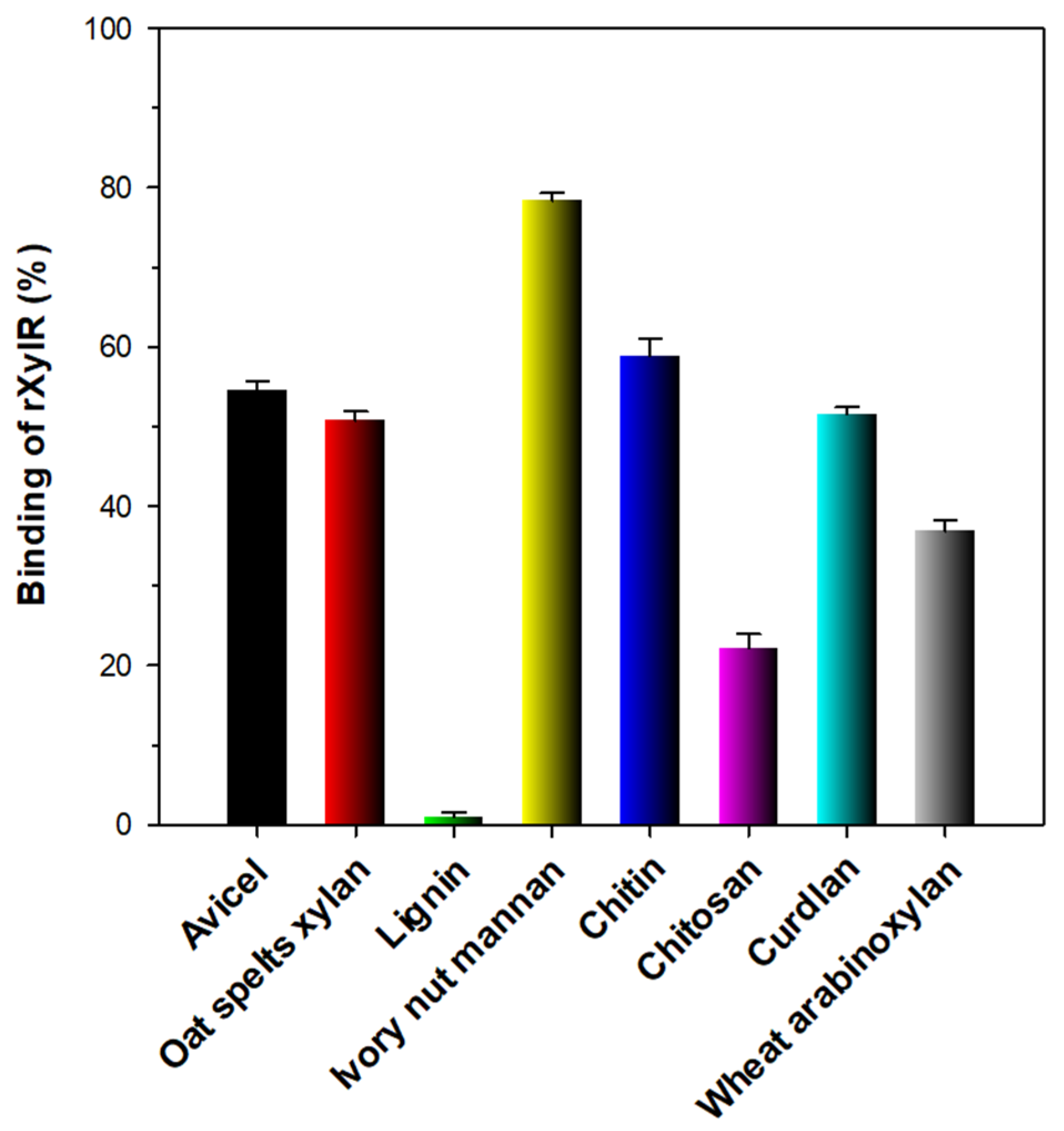
3.6. Binding Affinity of rXylR to Hydrophobic Polymers
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Collins, T.; Gerady, C.; Feller, G. Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiol. Rev. 2005, 29, 3–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shallom, D.; Shoham, Y. Microbial hemicellulases. Curr. Opin. Microbiol. 2003, 6, 219–228. [Google Scholar] [CrossRef]
- Malgas, S.; Mafa, M.S.; Mkabayi, L.; Pletschke, B.I. A mini review of xylanolytic enzymes with regards to their synergistic interactions during hetero-xylan degradation. World J. Microbiol. Biotechnol. 2019, 35, 187. [Google Scholar] [CrossRef] [PubMed]
- Alokika; Singh, B. Production, characteristics, and biotechnological applications of microbial xylanases. Appl. Microbiol. Biotechnol. 2019, 103, 8763–8784. [Google Scholar] [CrossRef]
- Kim, D.Y.; Ham, S.-J.; Kim, H.J.; Kim, J.; Lee, M.-H.; Cho, H.-Y.; Shin, D.-H.; Rhee, Y.H.; Son, K.-H.; Park, H.-Y. Novel modular endo-β-1,4-xylanase with transglycosylation activity from Cellulosimicrobium sp. strain HY-13 that is homologous to inverting GH family 6 enzymes. Bioresour. Technol. 2012, 107, 25–32. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Kumar, B.; Verma, P. A detailed overview of xylanases: An emerging biomolecule for current and future perspective. Bioresour. Bioprocess. 2019, 6, 40. [Google Scholar] [CrossRef] [Green Version]
- Juturu, V.; Wu, J.C. Microbial xylanases: Engineering, production and industrial applications. Biotechnol. Adv. 2012, 30, 1219–1227. [Google Scholar] [CrossRef]
- Polizeli, M.; Rizzatti, A.; Monti, R.; Terenzi, H.; Jorge, J.A.; Amorim, D. Xylanases from fungi: Properties and industrial applications. Appl. Microbiol. Biotechnol. 2005, 67, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Tokuda, G. Lignocellulose-degrading enzymes from termites and their symbiotic microbiota. Biotechnol. Adv. 2013, 31, 838–850. [Google Scholar] [CrossRef]
- Chadha, B.S.; Kaur, B.; Basotra, N.; Tsang, A.; Pandey, A. Thermostable xylanases from thermophilic fungi and bacteria: Current perspective. Bioresour. Technol. 2019, 277, 195–203. [Google Scholar] [CrossRef]
- Petrescu, I.; Lamotte-Brasseur, J.; Chessa, J.-P.; Ntarima, P.; Claeyssens, M.; Devreese, B.; Marino, G.; Gerday, C. Xylanase from the psychrophilic yeast Cryptococcus adeliae. Extremophiles 2000, 4, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.; Meuwis, M.-A.; Stals, I.; Claeyssens, M.; Feller, G. A novel family 8 xylanase, functional and physicochemical characterization. J. Biol. Chem. 2002, 277, 35133–35139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Durán, C.; Ravanal, M.-C.; Ubilla, P.; Vaca, I.; Chávez, R. Heterologous expression, purification and characterization of a highly thermolabile endoxylanase from the Antarctic fungus Cladosporium sp. Fungal Biol. 2018, 122, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, J.P.; Reetz, M.T.; Asenjo, J.A.; Parra, L.P. One-step combined focused epPCR and saturation mutagenesis for thermostability evolution of a cold-active xylanase. Enzym. Microb. Technol. 2017, 100, 60–70. [Google Scholar] [CrossRef]
- Wang, G.; Luo, H.; Wang, Y.; Huang, H.; Shi, P.; Yang, P.; Meng, K.; Bai, Y.; Yao, B. A novel cold-active xylanase gene from the environmental DNA of goat rumen contents: Direct cloning, expression and enzyme characterization. Bioresour. Technol. 2011, 102, 3330–3336. [Google Scholar] [CrossRef]
- Chen, S.; Kaufman, M.G.; Miazgowicz, K.L.; Bagdasarian, M. Molecular characterization of a cold-active recombinant xylanase from Flavobacterium johnsoniae and its applicability in xylan hydrolysis. Bioresour. Technol. 2013, 128, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Huang, Z.; Zhang, X.; Shao, Z.; Liu, Z. Cloning, expression and characterization of a novel cold-active and halophilic xylanase from Zunongwangia profunda. Extremophiles 2014, 18, 441–450. [Google Scholar] [CrossRef]
- Sun, J.; Shi, S.; Wu, J.; Xie, R.; Geng, A.; Zhu, D.C. Characterization of a salt-tolerant and cold-adapted xylanase from Bacillus cellulosilyticus. BioResources 2016, 11, 8875–8889. [Google Scholar] [CrossRef] [Green Version]
- Duarte, A.W.F.; Barato, M.B.; Nobre, F.S.; Polezel, D.A.; de Oliveira, T.B.; dos Santos, J.A.; Rodrigues, A.; Sette, L.D. Production of cold-adapted enzymes by filamentous fungi from King George Island, Antarctica. Polar Biol. 2018, 41, 2511–2521. [Google Scholar] [CrossRef] [Green Version]
- Gerday, C.; Aittaleb, M.; Bentahir, M.; Chessa, J.-P.; Claverie, P.; Collins, T.; D’Amico, S.; Dumont, J.; Garsoux, G.; Georlette, D.; et al. Cold-adapted enzymes: From fundamentals to biotechnology. Trends Biotechnol. 2000, 18, 103–107. [Google Scholar] [CrossRef]
- Kim, D.Y.; Han, M.K.; Oh, H.W.; Bae, K.S.; Jeong, T.-S.; Kim, S.U.; Shin, D.-H.; Kim, I.-H.; Rhee, Y.H.; Son, K.-H.; et al. Novel intracellular GH10 xylanase from Cohnella laeviribosi HY-21: Biocatalytic properties and alterations of substrate specificities by site-directed mutagenesis of Trp residues. Bioresour. Technol. 2010, 101, 8814–8821. [Google Scholar] [CrossRef]
- Kim, D.Y.; Lee, S.H.; Lee, M.J.; Cho, H.-Y.; Lee, J.S.; Rhee, Y.H.; Shin, D.-H.; Son, K.-H.; Park, H.-Y. Genetic and functional characterization of a novel GH10 endo-β-1,4-xylanase with a ricin-type β-trefoil domain-like domain from Luteimicrobium xylanilyticum HY-24. Int. J. Biol. Macromol. 2018, 106, 620–628. [Google Scholar] [CrossRef]
- Kim, D.Y.; Han, M.K.; Oh, H.-W.; Park, D.-S.; Kim, S.-J.; Lee, S.-G.; Shin, D.-H.; Son, K.-H.; Bae, K.S.; Park, H.-Y. Catalytic properties of a GH10 endo-β-1,4-xylanase from Streptomyces thermocarboxydus HY-15 isolated from the gut of Eisenia fetida. J. Mol. Catal. B Enzym. 2010, 62, 32–39. [Google Scholar] [CrossRef]
- Zolotnitsky, G.; Cogan, U.; Adir, N.; Solomon, V.; Shoham, G.; Shoham, Y. Mapping glycoside hydrolase substrate subsites by isothermal titration calorimetry. Proc. Natl. Acad. Sci. USA 2004, 101, 11275–11280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Upadhyay, V.; Upadhyay, A.K.; Singh, S.M.; Panda, A.K. Protein recovery from inclusion bodies of Escherichia coli using mild solubilization process. Microb. Cell Fact. 2015, 14, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.Y.; Han, M.K.; Park, D.-S.; Lee, J.S.; Oh, H.-W.; Shin, D.-H.; Jeong, T.-S.; Kim, S.U.; Bae, K.S.; Son, K.-H.; et al. Novel GH10 xylanase, with a fibronectin type 3 domain, from Cellulosimicrobium sp. strain HY-13, a bacterium in the gut of Eisenia fetida. Appl. Environ. Microbiol. 2009, 75, 7275–7279. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Liu, L.; Liu, X.; Tang, K. Isolation and characterization of a novel cold-active, halotolerant endoxylanase from Echinicola rosea sp. nov. JL3085T. Mar. Drugs 2020, 18, 245. [Google Scholar] [CrossRef] [PubMed]
- Gomes, J.; Gomes, I.; Steiner, W. Thermolabile xylanase of the Antarctic yeast Cryptococcus adeliae: Production and properties. Extremophiles 2000, 4, 227–235. [Google Scholar] [CrossRef]
- Bai, W.; Xue, Y.; Zhou, C.; Ma, Y. Cloning, expression and characterization of a novel salt-tolerant xylanase from Bacillus sp. SN5. Biotechnol. Lett. 2012, 34, 2093–2099. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Chen, X.-L.; Sun, C.-Y.; Zhou, B.-C.; Zhang, Y.-Z. Gene cloning, expression and characterization of a new cold-active and salt-tolerant endo-β-1,4-xylanase from marine Glaciecola mesophila KMM 241. Appl. Microbiol. Biotechnol. 2009, 84, 1107–1115. [Google Scholar] [CrossRef]
- Kim, D.Y.; Shin, D.-H.; Jung, S.; Lee, J.S.; Cho, H.Y.; Bae, K.S.; Sung, C.-K.; Rhee, Y.H.; Son, K.-H.; Park, H.-Y. Biocatalytic properties and substrate-binding ability of a modular GH10 β-1,4-xylanase from an insect-symbiotic bacterium, Streptomyces mexicanus HY-14. J. Microbiol. 2014, 52, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Park, I.; Cho, J. Partial characterization of extracellular xylanolytic activity derived from Paenibacillus sp. KIJ1. Afr. J. Microbiol. Res. 2010, 4, 1257–1264. [Google Scholar]
- Kim, D.Y.; Shin, D.-H.; Jung, S.; Kim, H.; Lee, J.S.; Cho, H.Y.; Bae, K.S.; Sung, C.-K.; Rhee, Y.H.; Son, K.-H.; et al. Novel alkali-tolerant GH10 endo-β-1,4-xylanase with broad substrate specificity from Microbacterium trichothecenolyticum HY-17, a gut bacterium of the mole cricket Gryllotalpa orientalis. J. Microbiol. Biotechnol. 2014, 24, 943–953. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | GH Family | Enzyme | Mr (kDa) | Optimum pH | Optimum Temp. (°C) | kcat/Km (mg−1 s−1 mL) | Reference |
|---|---|---|---|---|---|---|---|
| Duganella sp. PAMC 27433 | 10 | rXylR | 42.0 | 5.5 | 40 | 185.30 a, 52.52 b | This study |
| Psychrobacter sp. strain 2-17 | 10 | Xyl-L | 90.0 | 8.0 | 35 | 35.40 b | [14] |
| Cladosporium sp. | 10 | XynA | 37.0 | 6.0 | 50 | NI c | [13] |
| Cryptococcus adeliae | 10 | Xylanase | 43.0 | 5.0–5.5 | 45–50 | NI | [11,28] |
| Pseudoalteromonas haloplanktis | 8 | Xylanase | 45.9 | 5.3–8.0 | 35 | 44.53 b | [12] |
| Substrate | Specific Activity (U/mg) a |
|---|---|
| Birchwood xylan | 103.2 ± 2.2 |
| Beechwood xylan | 274.7 ± 1.8 |
| Oat spelts xylan | 35.6 ± 0.6 |
| Wheat arabinoxylan | 38.9 ± 1.4 |
| Xyloglucan | ND b |
| Locust bean gum | ND |
| Carboxymethylcellulose | ND |
| pNP-β-d-cellobioside | 365.1 ± 2.6 |
| pNP-β-d-glucopyranoside | ND |
| pNP-β-d-xylopyranoside | 6.7 ± 0.2 |
| pNP-β-d-mannopyranoside | ND |
| pNP-β-d-galactopyranoside | ND |
| Substrate | Vmax (U mg−1) | Km (mg mL−1) | Km (mM) | kcat (s−1) | kcat/Km (mg−1 s−1 mL) | kcat/Km (mM−1 s−1) |
|---|---|---|---|---|---|---|
| Birchwood xylan | 181.60 | 2.42 | 127.12 | 52.52 | ||
| Beechwood xylan | 510.92 | 1.93 | 357.63 | 185.30 | ||
| Oat spelts xylan | 55.51 | 2.85 | 38.85 | 13.63 | ||
| Wheat arabinoxylan | 62.21 | 3.22 | 43.54 | 13.52 | ||
| pNP-β-d-cellobioside | 649.80 | 1.21 | 454.86 | 375.91 |
| Substrate | Composition (%) a of Products Formed by Degradation | |
|---|---|---|
| d-Xylose (X1) | d-Xylobiose (X2) | |
| d-Xylobiose (X2) | 1.8 | 98.2 |
| d-Xylotriose (X3) | 19.3 | 80.7 |
| d-Xylotetraose (X4) | 22.3 | 77.7 |
| d-Xylopentaose (X5) | 25.8 | 74.2 |
| d-Xylohexaose (X6) | 28.1 | 71.9 |
| Birchwood xylan | 37.1 | 62.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.Y.; Kim, J.; Lee, Y.M.; Lee, J.S.; Shin, D.-H.; Ku, B.-H.; Son, K.-H.; Park, H.-Y. Identification and Characterization of a Novel, Cold-Adapted d-Xylobiose- and d-Xylose-Releasing Endo-β-1,4-Xylanase from an Antarctic Soil Bacterium, Duganella sp. PAMC 27433. Biomolecules 2021, 11, 680. https://doi.org/10.3390/biom11050680
Kim DY, Kim J, Lee YM, Lee JS, Shin D-H, Ku B-H, Son K-H, Park H-Y. Identification and Characterization of a Novel, Cold-Adapted d-Xylobiose- and d-Xylose-Releasing Endo-β-1,4-Xylanase from an Antarctic Soil Bacterium, Duganella sp. PAMC 27433. Biomolecules. 2021; 11(5):680. https://doi.org/10.3390/biom11050680
Chicago/Turabian StyleKim, Do Young, Jonghoon Kim, Yung Mi Lee, Jong Suk Lee, Dong-Ha Shin, Bon-Hwan Ku, Kwang-Hee Son, and Ho-Yong Park. 2021. "Identification and Characterization of a Novel, Cold-Adapted d-Xylobiose- and d-Xylose-Releasing Endo-β-1,4-Xylanase from an Antarctic Soil Bacterium, Duganella sp. PAMC 27433" Biomolecules 11, no. 5: 680. https://doi.org/10.3390/biom11050680