
High Glucose Concentrations Impair the Processing and Presentation of Mycobacterium tuberculosis Antigens In Vitro

 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Bacterial Culture
2.2. High-Glucose Model In Vitro and Human Monocyte-Derived Macrophages (MDMs) and Dendritic Cells (DCs)
High Glucose Stimulation
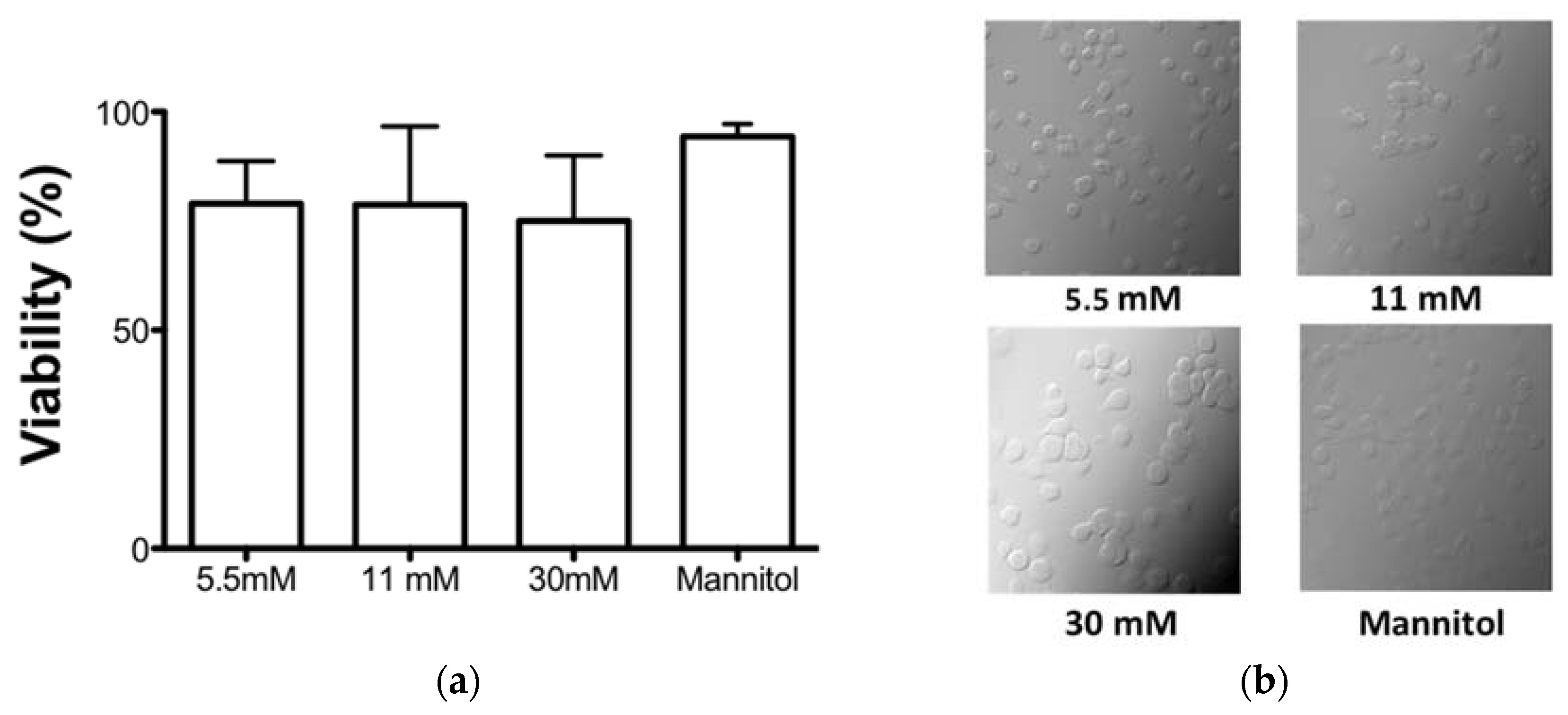
2.3. WST-1 Viability Assay
2.4. Antibodies
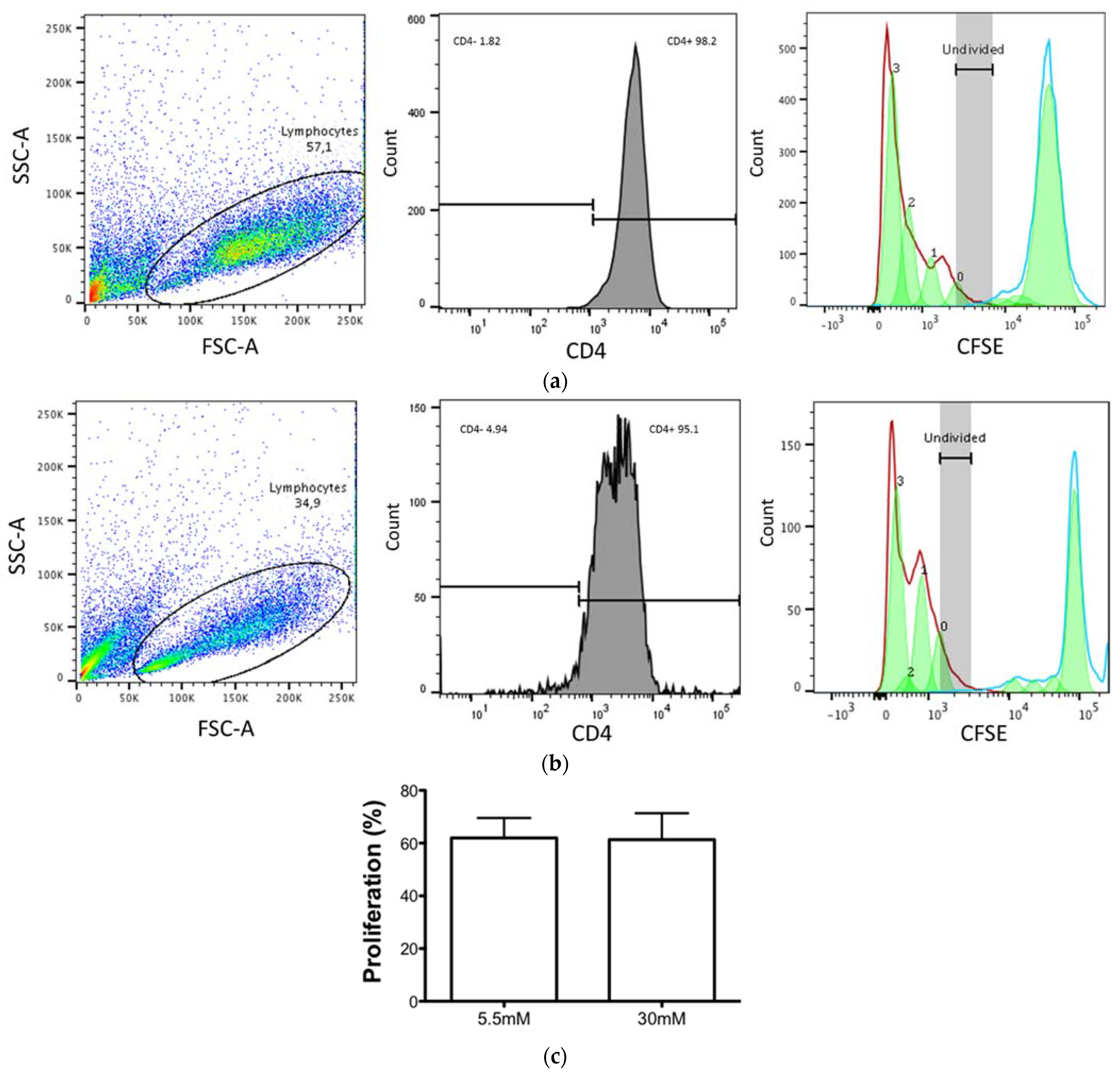
2.5. CD4+ T-Lymphocyte Proliferation Assays
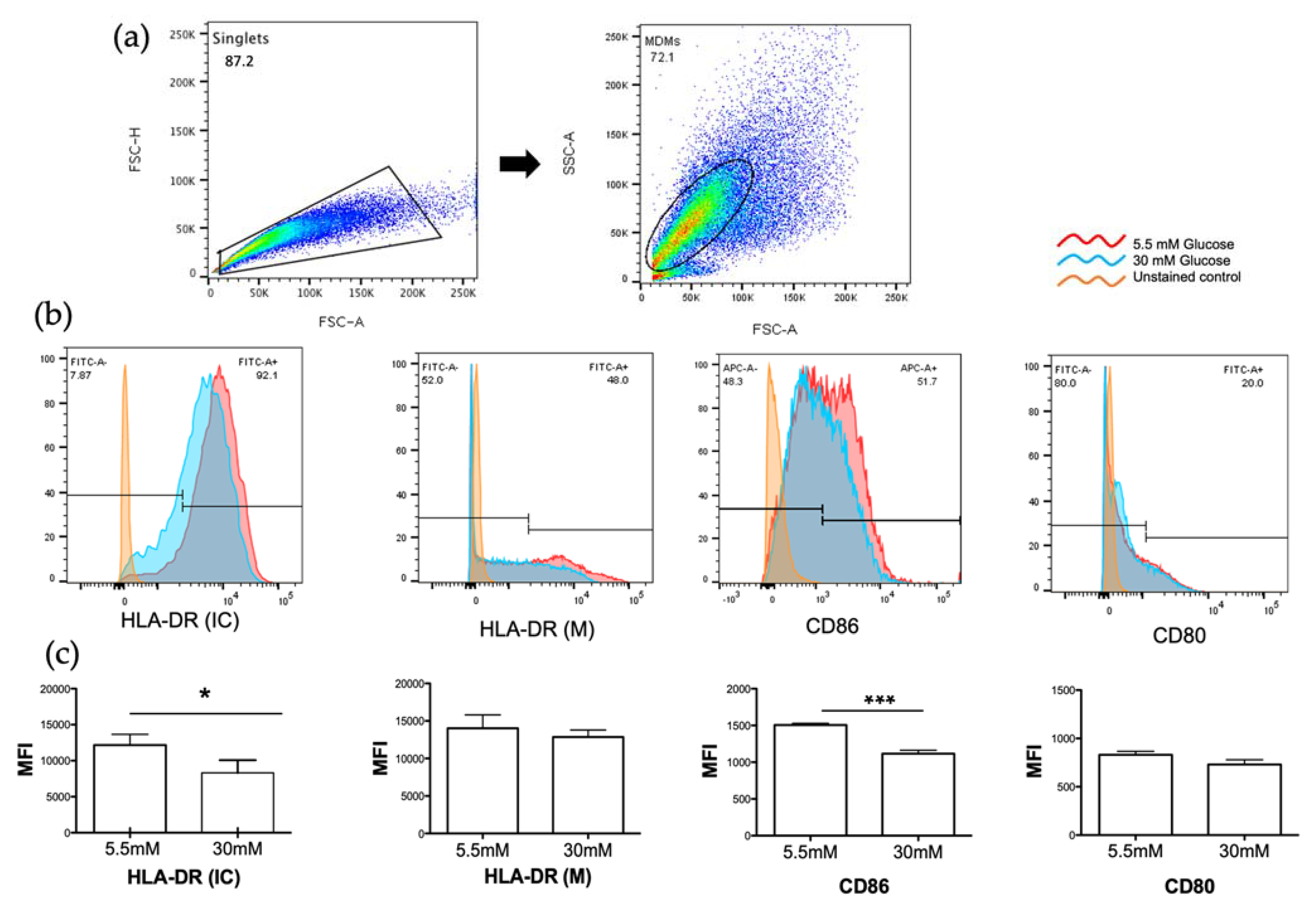
2.6. Detection of HLA-DR, CD80, and CD86 Molecules by Flow Cytometry
2.7. Detection of Proteins of the Phagolysosomal Pathway by Confocal Microscopy
2.8. Amplification of M. tuberculosis-Specific CD4+ T Cells
2.9. Detection of M. tuberculosis Peptide–MHC II Complexes and Determination of M. tuberculosis-Processing Kinetics in Different Concentrations of Glucose
3. Results
3.1. High Glucose Concentrations Did Not Affect the Cell Viability in an In Vitro Model
3.2. CD4+ T Cell Proliferation Was Maintained despite the High Glucose Concentrations
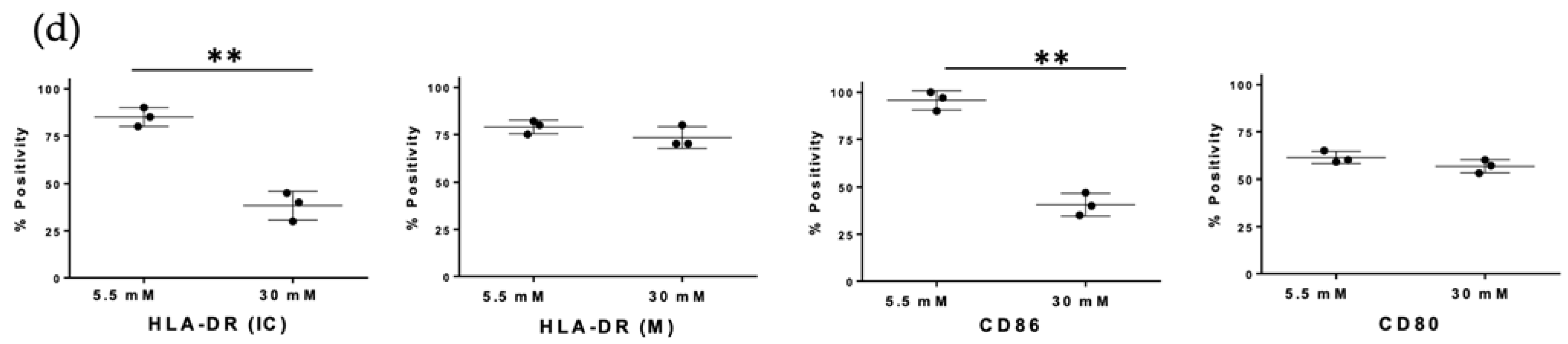
3.3. High Glucose Concentrations Decreased MHC Class II and CD86 Molecules in MDMs
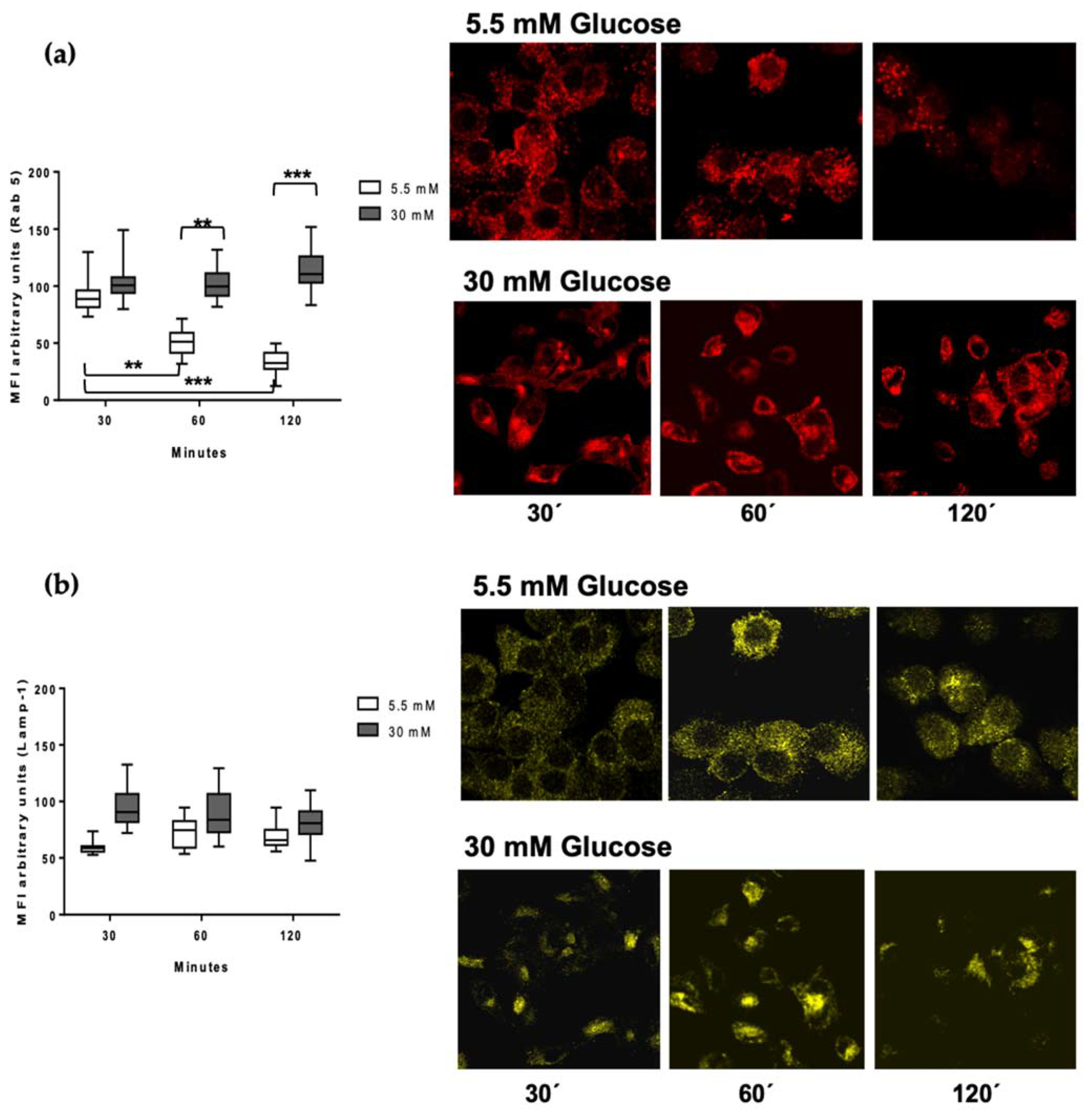
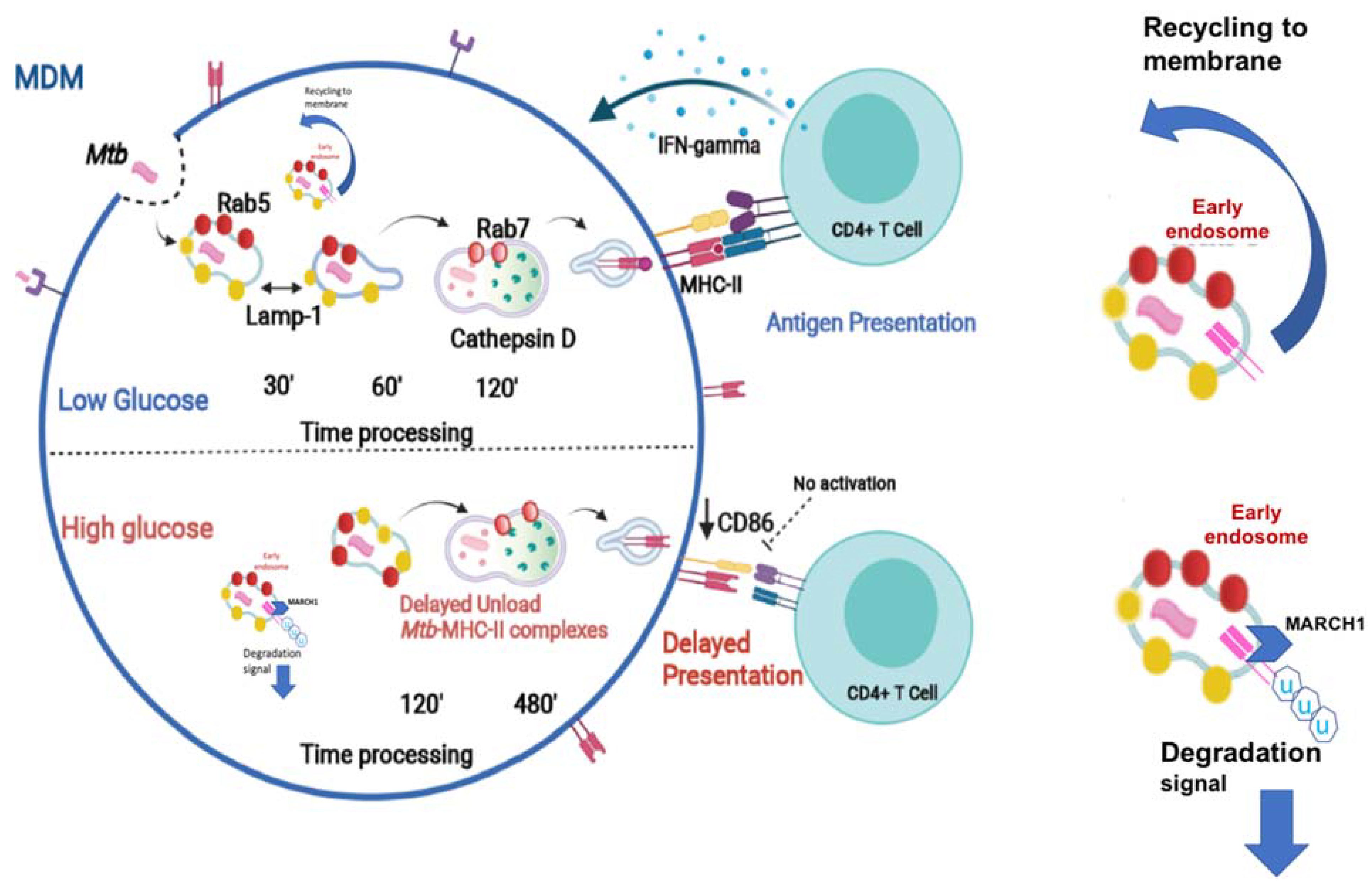
3.4. Rab 5 and Lamp-1 Are Retained in High Glucose Concentrations
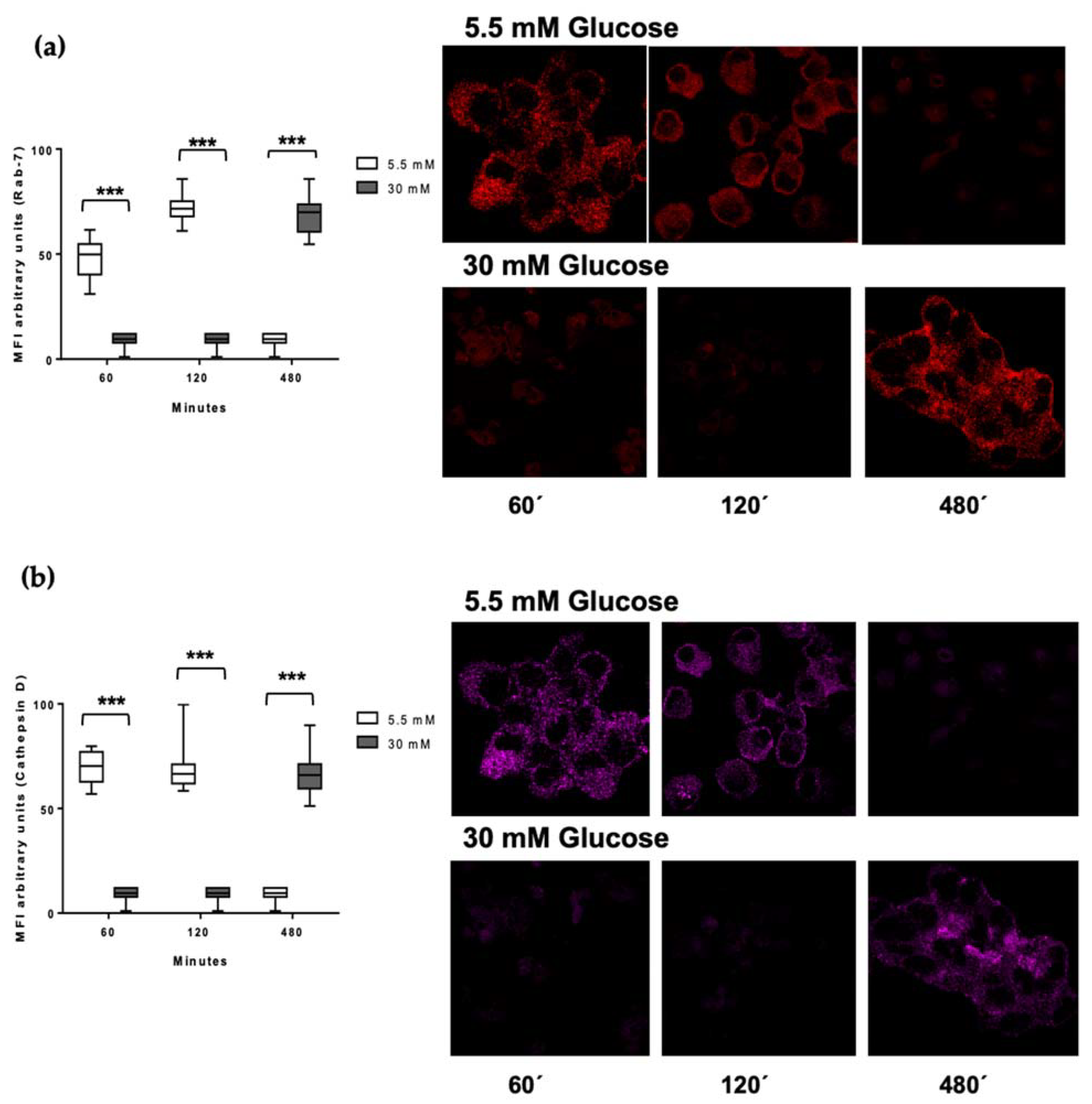
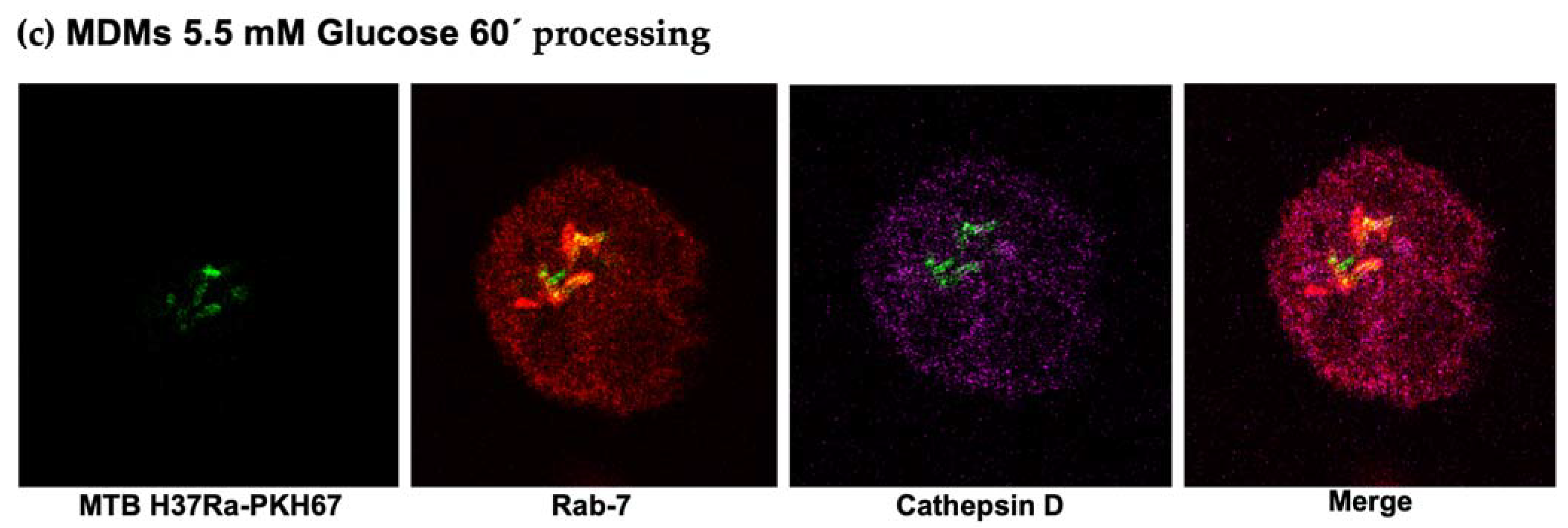
3.5. Delayed Acquisition of Rab 7 and Cathepsin D in High Glucose Concentrations
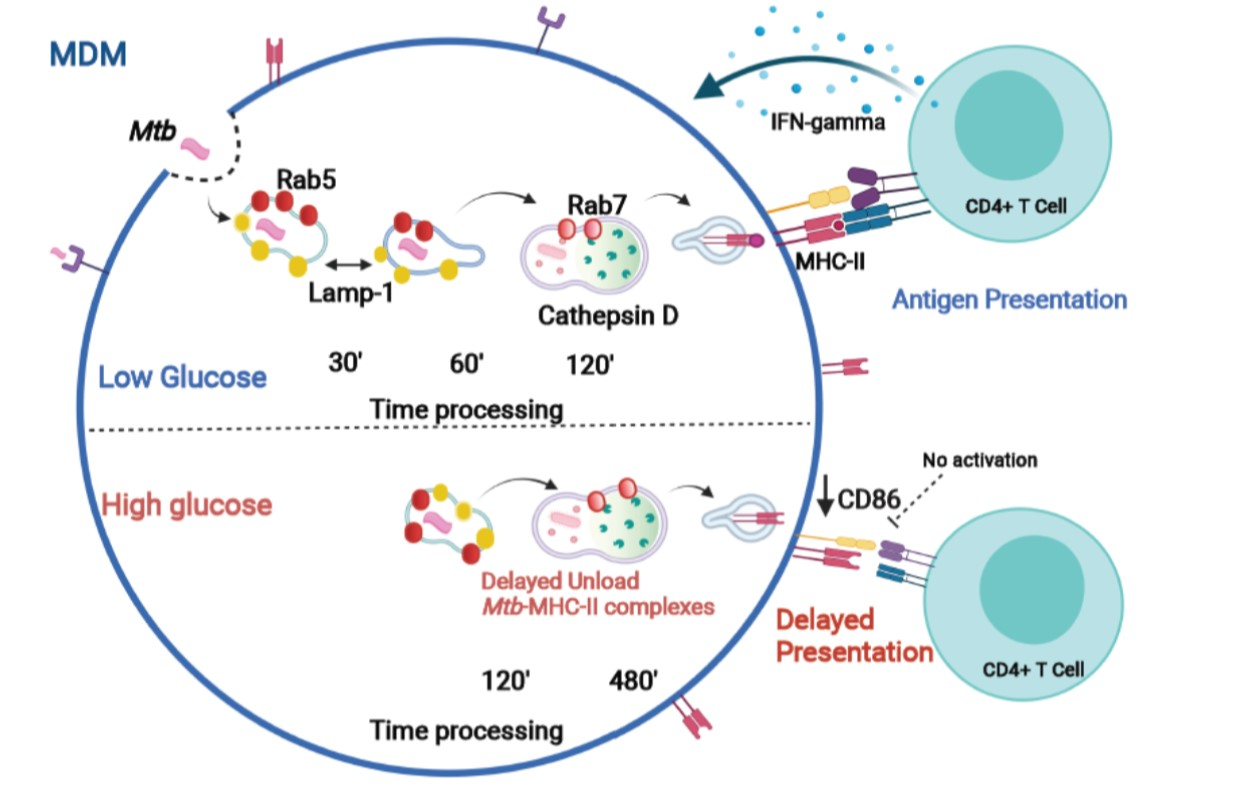
3.6. High Glucose Concentrations Impair M. tuberculosis Antigenic Presentation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Diabetes Association. Erratum. Classification and diagnosis of diabetes. Sec. 2. Standards of Medical Care in Diabetes-2016. Diabetes Care 2016, 39 (Suppl. 1), S13–S22. [Google Scholar]
- American Diabetes Association. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2020. Diabetes Care 2020, 43, S14–S31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, B.R.; Hux, J.E. Quantifying the Risk of Infectious Diseases for People with Diabetes. Diabetes Care 2003, 26, 510–513. [Google Scholar] [CrossRef] [Green Version]
- Russell, D.G.; Barry, C.E., 3rd; Flynn, J.L. Tuberculosis: What we don’t know can, and does, hurt us. Science 2010, 328, 852–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dooley, K.E.; Chaisson, R.E. Tuberculosis and diabetes mellitus: Convergence of two epidemics. Lancet Infect. Dis. 2009, 9, 737–746. [Google Scholar] [CrossRef] [Green Version]
- Dooley, K.E.; Golub, J.E.; Cronin, W.; Tang, T.; Dorman, S.E. Impact of Diabetes Mellitus on Treatment Outcomes of Patients with Active Tuberculosis. Am. J. Trop. Med. Hyg. 2009, 80, 634–639. [Google Scholar] [CrossRef]
- O’Garra, A.; Redford, P.S.; McNab, F.W.; Bloom, C.I.; Wilkinson, R.J.; Berry, M.P. The immune response in tuberculosis. Annu. Rev. Immunol. 2013, 31, 475–527. [Google Scholar] [CrossRef] [PubMed]
- Verreck, F.A.; de Boer, T.; Langenberg, D.M.; van der Zanden, L.; Ottenhoff, T.H. Phenotypic and functional profiling of human proinflammatory type-1 and anti-inflammatory type-2 macrophages in response to microbial antigens and IFN-gamma- and CD40L-mediated costimulation. J. Leukoc. Biol. 2006, 79, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, M. Biogenesis of phagolysosomes: The ’kiss and run’ hypothesis. Trends Cell Biol. 1995, 5, 183–186. [Google Scholar] [CrossRef]
- Boulais, J.; Trost, M.; Landry, C.R.; Dieckmann, R.; Levy, E.D.; Soldati, T.; Michnick, S.W.; Thibault, P.; Desjardins, M. Molecular characterization of the evolution of phagosomes. Mol. Syst. Biol. 2010, 6, 423. [Google Scholar] [CrossRef]
- Levin, R.; Grinstein, S.; Canton, J. The life cycle of phagosomes: Formation, maturation, and resolution. Immunol. Rev. 2016, 273, 156–179. [Google Scholar] [CrossRef]
- Steigbigel, R.T.; Lambert, L.H., Jr.; Remington, J.S. Phagocytic and bacterial properties of normal human monocytes. J. Clin. Investig. 1974, 53, 131–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, M.G. Functional role(s) of phagosomal Rab GTPases. Small GTPases 2013, 4, 148–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawe, D.C.; Chawla, A.; Merithew, E.; Dumas, J.; Carrington, W.; Fogarty, K.; Lifshitz, L.; Tuft, R.; Lambright, D.; Corvera, S. Sequential Roles for Phosphatidylinositol 3-Phosphate and Rab5 in Tethering and Fusion of Early Endosomes via Their Interaction with EEA1. J. Biol. Chem. 2002, 277, 8611–8617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yates, R.M.; Hermetter, A.; Russell, D.G. The Kinetics of Phagosome Maturation as a Function of Phagosome/Lysosome Fusion and Acquisition of Hydrolytic Activity. Traffic 2005, 6, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Jordens, I.; Fernandez-Borja, M.; Marsman, M.; Dusseljee, S.; Janssen, L.; Calafat, J.; Janssen, H.; Wubbolts, R.; Neefjes, J. The Rab7 effector protein RILP controls lysosomal transport by inducing the recruitment of dynein-dynactin motors. Curr. Biol. 2001, 11, 1680–1685. [Google Scholar] [CrossRef] [Green Version]
- Hu, R.; Xia, C.-Q.; Butfiloski, E.; Clare-Salzler, M. Effect of high glucose on cytokine production by human peripheral blood immune cells and type I interferon signaling in monocytes: Implications for the role of hyperglycemia in the diabetes inflammatory process and host defense against infection. Clin. Immunol. 2018, 195, 139–148. [Google Scholar] [CrossRef]
- Vallerskog, T.; Martens, G.W.; Kornfeld, H. Diabetic Mice Display a Delayed Adaptive Immune Response toMycobacterium tuberculosis. J. Immunol. 2010, 184, 6275–6282. [Google Scholar] [CrossRef] [Green Version]
- Geerlings, S.E.; Hoepelman, A.I. Immune dysfunction in patients with diabetes mellitus (DM). FEMS Immunol. Med. Microbiol. 1999, 26, 259–265. [Google Scholar] [CrossRef]
- Gomez, D.I.; Twahirwa, M.; Schlesinger, L.S.; Restrepo, B.I. Reduced Mycobacterium tuberculosis association with monocytes from diabetes patients that have poor glucose control. Tuberculosis 2013, 93, 192–197. [Google Scholar] [CrossRef] [Green Version]
- Restrepo, B.I.; Twahirwa, M.; Rahbar, M.H.; Schlesinger, L.S. Phagocytosis via Complement or Fc-Gamma Receptors Is Compromised in Monocytes from Type 2 Diabetes Patients with Chronic Hyperglycemia. PLoS ONE 2014, 9, e92977. [Google Scholar] [CrossRef] [Green Version]
- Ilyas, R.; Wallis, R.; Soilleux, E.J.; Townsend, P.; Zehnder, D.; Tan, B.K.; Sim, R.B.; Lehnert, H.; Randeva, H.S.; Mitchell, D.A. High glucose disrupts oligosaccharide recognition function via competitive inhibition: A potential mechanism for immune dysregulation in diabetes mellitus. Immunobiology 2011, 216, 126–131. [Google Scholar] [CrossRef] [Green Version]
- Via, L.; Deretic, D.; Ulmer, R.J.; Hibler, N.S.; Huber, L.A.; Deretic, V. Arrest of Mycobacterial Phagosome Maturation Is Caused by a Block in Vesicle Fusion between Stages Controlled by rab5 and rab7. J. Biol. Chem. 1997, 272, 13326–13331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rink, J.; Ghigo, E.; Kalaidzidis, Y.; Zerial, M. Rab Conversion as a Mechanism of Progression from Early to Late Endosomes. Cell 2005, 122, 735–749. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.S.; Lee, K.-O.; Low, K.C.; Gamage, A.M.; Liu, Y.; Tan, G.-Y.G.; Koh, H.Q.V.; Alonso, S.; Gan, Y.-H. Glutathione deficiency in type 2 diabetes impairs cytokine responses and control of intracellular bacteria. J. Clin. Investig. 2012, 122, 2289–2300. [Google Scholar] [CrossRef] [PubMed]
- Tsukaguchi, K.; Okamura, H.; Matsuzawa, K.; Tamura, M.; Miyazaki, R.; Tamaki, S.; Kimura, H. Longitudinal assessment of IFN-gamma production in patients with pulmonary tuberculosis complicated with diabetes mellitus. Kekkaku 2002, 77, 409–413. [Google Scholar]
- Kitabchi, A.E.; Umpierrez, G.E.; Miles, J.M.; Fisher, J.N. Hyperglycemic Crises in Adult Patients with Diabetes. Diabetes Care 2009, 32, 1335–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif, K.; Ghadir, S.; Jakubowicz, D.; Amital, H.; Bragazzi, N.L.; Watad, A.; Wainstein, J.; Bar-Dayan, Y. Improved outcome of patients with diabetes mellitus with good glycemic control in the cardiac intensive care unit: A retrospective study. Cardiovasc. Diabetol. 2019, 18, 4. [Google Scholar] [CrossRef] [Green Version]
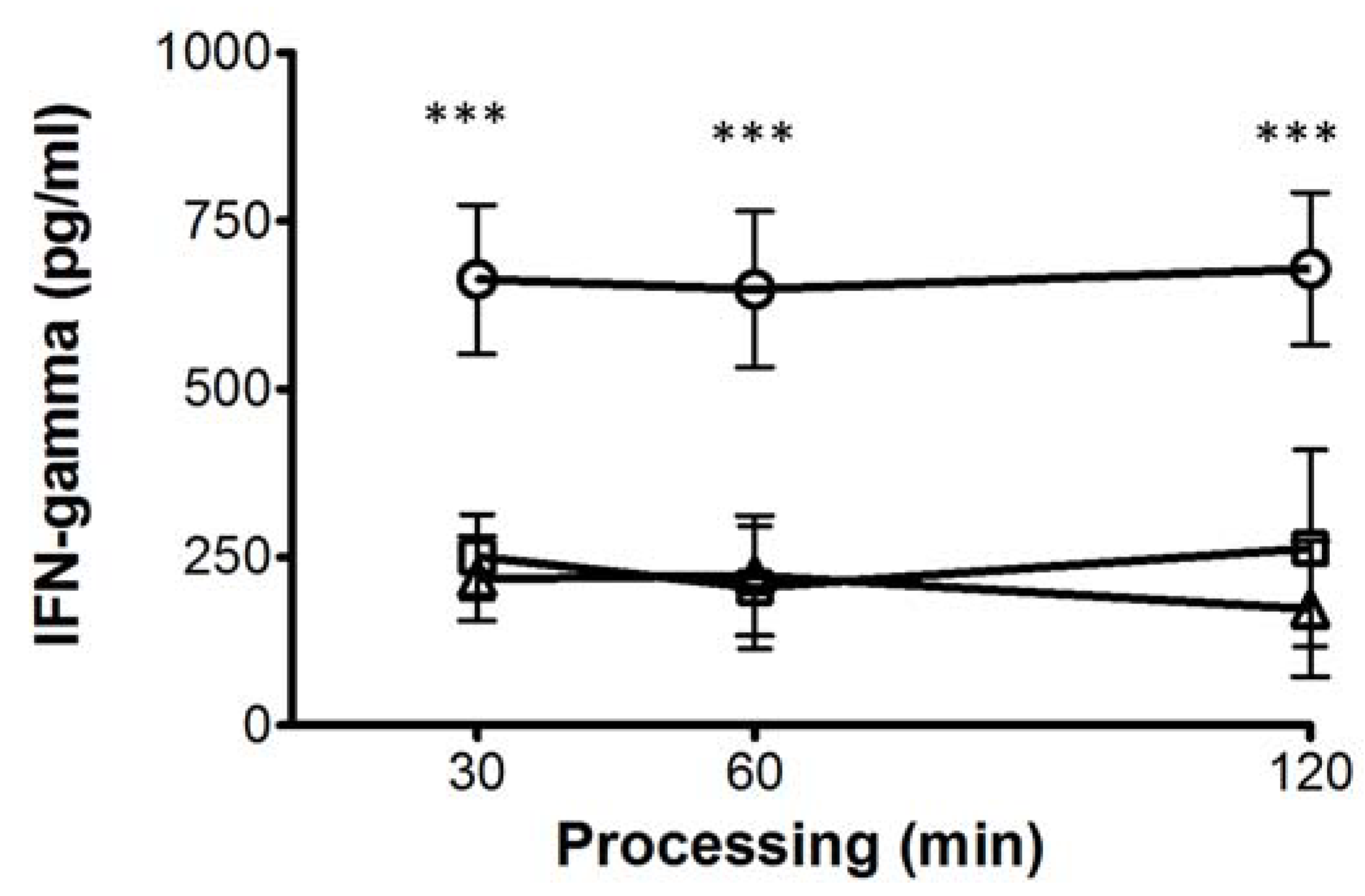
- Bobadilla, K.; Sada, E.; Jaime, M.E.; Gonzalez, Y.; Ramachandra, L.; Rojas, R.E.; Pedraza-Sanchez, S.; Michalak, C.; Gonzalez-Noriega, A.; Torres, M. Human phagosome processing of Mycobacterium tuberculosis antigens is modulated by interferon-gamma and interleukin-10. Immunology 2013, 138, 34–46. [Google Scholar] [CrossRef]
- Tan, J.S.; Canaday, D.H.; Boom, W.H.; Balaji, K.N.; Schwander, S.K.; A Rich, E. Human alveolar T lymphocyte responses to Mycobacterium tuberculosis antigens: Role for CD4+ and CD8+ cytotoxic T cells and relative resistance of alveolar macrophages to lysis. J. Immunol. 1997, 159, 290–297. [Google Scholar]
- Torres, M.; Ramachandra, L.; Rojas, R.E.; Bobadilla, K.; Thomas, J.; Canaday, D.H.; Harding, C.V.; Boom, W.H. Role of Phagosomes and Major Histocompatibility Complex Class II (MHC-II) Compartment in MHC-II Antigen Processing of Mycobacterium tuberculosis in Human Macrophages. Infect. Immun. 2006, 74, 1621–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, V.A.; Schorey, J.S. Mycobacterium’s arrest of phagosome maturation in macrophages requires Rab5 activity and accessibility to iron. Mol. Biol. Cell 2003, 14, 3366–3377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restrepo, B.I. Diabetes and Tuberculosis. Microbiol. Spectr. 2016, 4, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadena, J.; Rathinavelu, S.; Lopez-Alvarenga, J.C.; Restrepo, B.I. The re-emerging association between tuberculosis and diabetes: Lessons from past centuries. Tuberculosis 2019, 116, S89–S97. [Google Scholar] [CrossRef]
- Martens, G.W.; Arikan, M.C.; Lee, J.; Ren, F.; Greiner, D.; Kornfeld, H. Tuberculosis Susceptibility of Diabetic Mice. Am. J. Respir. Cell Mol. Biol. 2007, 37, 518–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohashi, P.S.; Oehen, S.; Aichele, P.; Pircher, H.; Odermatt, B.; Herrera, P.; Higuchi, Y.; Buerki, K.; Hengartner, H.; Zinkernagel, R.M. Induction of diabetes is influenced by the infectious virus and local expression of MHC class I and tumor necrosis factor-alpha. J. Immunol. 1993, 150, 5185–5194. [Google Scholar]
- Marroqui, L.; dos Santos, R.S.; De Beeck, A.O.; De Brachène, A.C.; Marselli, L.; Marchetti, P.; Eizirik, D.L. Interferon-α mediates human beta cell HLA class I overexpression, endoplasmic reticulum stress and apoptosis, three hallmarks of early human type 1 diabetes. Diabetologia 2017, 60, 656–667. [Google Scholar] [CrossRef] [Green Version]
- Matsuki, Y.; Ohmura-Hoshino, M.; Goto, E.; Aoki, M.; Mito-Yoshida, M.; Uematsu, M.; Hasegawa, T.; Koseki, H.; Ohara, O.; Nakayama, M.; et al. Novel regulation of MHC class II function in B cells. EMBO J. 2007, 26, 846–854. [Google Scholar] [CrossRef] [Green Version]
- Baravalle, G.; Park, H.; McSweeney, M.; Ohmura-Hoshino, M.; Matsuki, Y.; Ishido, S.; Shin, J.-S. Ubiquitination of CD86 Is a Key Mechanism in Regulating Antigen Presentation by Dendritic Cells. J. Immunol. 2011, 187, 2966–2973. [Google Scholar] [CrossRef] [Green Version]
- Ebrahim, R.; Thilo, L. Kinetic evidence that newly-synthesized endogenous lysosome-associated membrane protein-1 (LAMP-1) first transits early endosomes before it is delivered to lysosomes. Mol. Membr. Biol. 2011, 28, 227–242. [Google Scholar] [CrossRef]
- Nakagawa, T.; Roth, W.; Wong, P.; Nelson, A.; Farr, A.; Deussing, J.; Villadangos, J.A.; Ploegh, H.; Peters, C.; Rudensky, A.Y. Cathepsin L: Critical Role in Ii Degradation and CD4 T Cell Selection in the Thymus. Science 1998, 280, 450–453. [Google Scholar] [CrossRef] [PubMed]
- Riese, R.J.; Chapman, H.A. Cathepsins and compartmentalization in antigen presentation. Curr. Opin. Immunol. 2000, 12, 107–113. [Google Scholar] [CrossRef]
- Stalenhoef, J.E.; Alisjahbana, B.; Nelwan, E.J.; Ven-Jongekrijg, J.; Ottenhoff, T.H.M.; Meer, J.W.M.; Nelwan, R.H.; Netea, M.G.; Crevel, R. The role of interferon-gamma in the increased tuberculosis risk in type 2 diabetes mellitus. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 27, 97–103. [Google Scholar] [CrossRef]
- Kumar, N.P.; Sridhar, R.; Banurekha, V.V.; Jawahar, M.S.; Nutman, T.B.; Babu, S. Expansion of Pathogen-Specific T-Helper 1 and T-Helper 17 Cells in Pulmonary Tuberculosis with Coincident Type 2 Diabetes Mellitus. J. Infect. Dis. 2013, 208, 739–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellgaard, L.; Molinari, M.; Helenius, A. Setting the Standards: Quality Control in the Secretory Pathway. Science 1999, 286, 1882–1888. [Google Scholar] [CrossRef] [PubMed]
- Irshad, Z.; Xue, M.; Ashour, A.; Larkin, J.R.; Thornalley, P.J.; Rabbani, N. Activation of the unfolded protein response in high glucose treated endothelial cells is mediated by methylglyoxal. Sci. Rep. 2019, 9, 7889. [Google Scholar] [CrossRef] [Green Version]
- Harding, H.; Zhang, Y.; Bertolotti, A.; Zeng, H.; Ron, D. Perk Is Essential for Translational Regulation and Cell Survival during the Unfolded Protein Response. Mol. Cell 2000, 5, 897–904. [Google Scholar] [CrossRef]
- Harding, H.; Zhang, Y.; Zeng, H.; Novoa, I.; Lu, P.D.; Calfon, M.; Sadri, N.; Yun, C.; Popko, B.; Paules, R.S.; et al. An Integrated Stress Response Regulates Amino Acid Metabolism and Resistance to Oxidative Stress. Mol. Cell 2003, 11, 619–633. [Google Scholar] [CrossRef]
- Subramanian, A.; Capalbo, A.; Iyengar, N.R.; Rizzo, R.; di Campli, A.; Di Martino, R.; Monte, M.L.; Beccari, A.R.; Yerudkar, A.; Del Vecchio, C.; et al. Auto-regulation of Secretory Flux by Sensing and Responding to the Folded Cargo Protein Load in the Endoplasmic Reticulum. Cell 2019, 176, 1461–1476.e23. [Google Scholar] [CrossRef] [Green Version]
- Granados, D.P.; Tanguay, P.-L.; Hardy, M.-P.; Caron, É.; de Verteuil, D.; Meloche, S.; Perreault, C. ER stress affects processing of MHC class I-associated peptides. BMC Immunol. 2009, 10, 10. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monroy-Mérida, G.; Guzmán-Beltrán, S.; Hernández, F.; Santos-Mendoza, T.; Bobadilla, K. High Glucose Concentrations Impair the Processing and Presentation of Mycobacterium tuberculosis Antigens In Vitro. Biomolecules 2021, 11, 1763. https://doi.org/10.3390/biom11121763
Monroy-Mérida G, Guzmán-Beltrán S, Hernández F, Santos-Mendoza T, Bobadilla K. High Glucose Concentrations Impair the Processing and Presentation of Mycobacterium tuberculosis Antigens In Vitro. Biomolecules. 2021; 11(12):1763. https://doi.org/10.3390/biom11121763
Chicago/Turabian StyleMonroy-Mérida, Guadalupe, Silvia Guzmán-Beltrán, Fernando Hernández, Teresa Santos-Mendoza, and Karen Bobadilla. 2021. "High Glucose Concentrations Impair the Processing and Presentation of Mycobacterium tuberculosis Antigens In Vitro" Biomolecules 11, no. 12: 1763. https://doi.org/10.3390/biom11121763