Melatonin’s Impact on Antioxidative and Anti-Inflammatory Reprogramming in Homeostasis and Disease


 ,
,
Abstract
:1. Introduction
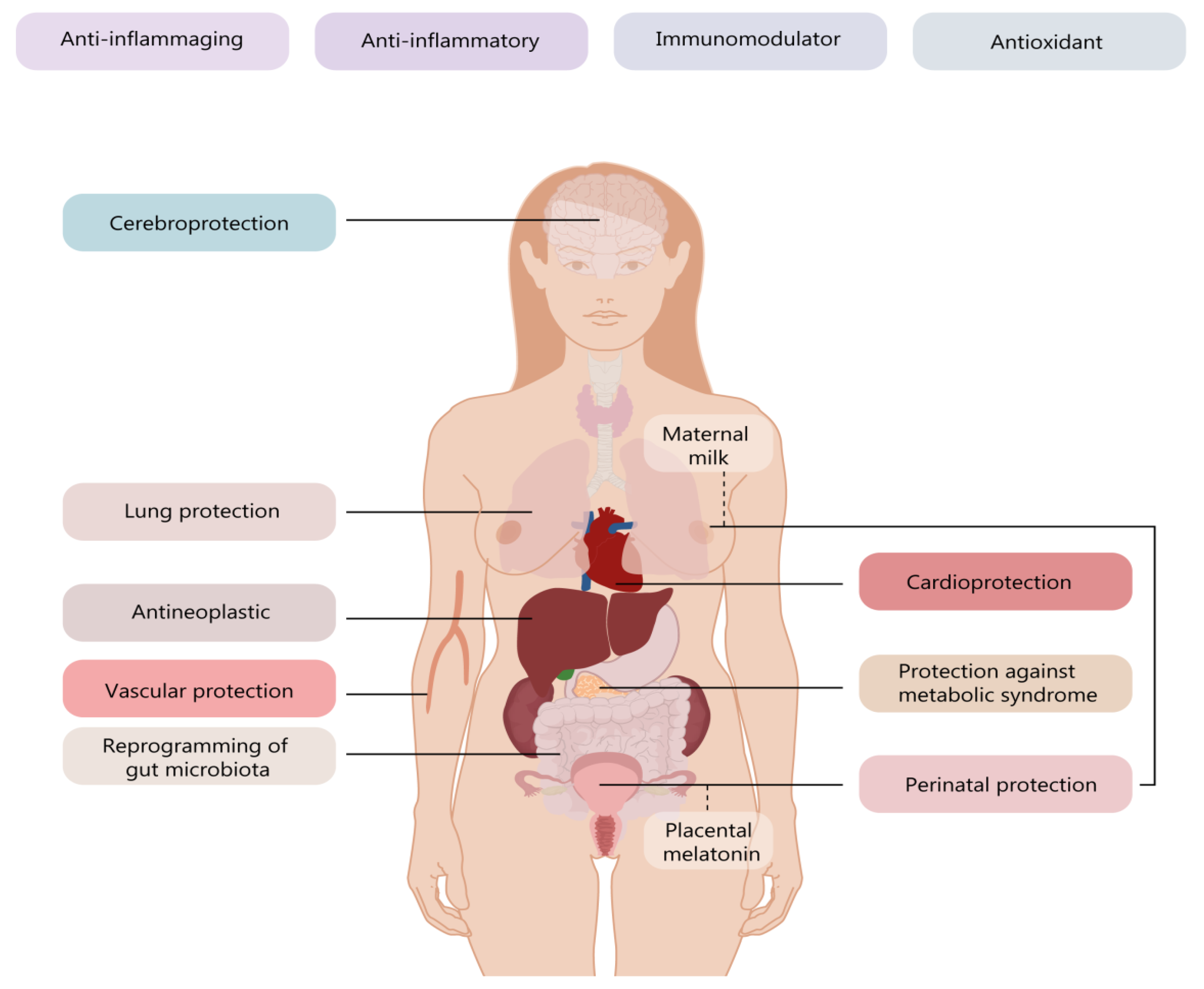
2. Melatonin Involvement in Maternal and Fetal Health
2.1. Melatonin in Maternal-Fetal Signaling
2.2. Melatonin for the Prevention of Maternal Pregnancy Complications
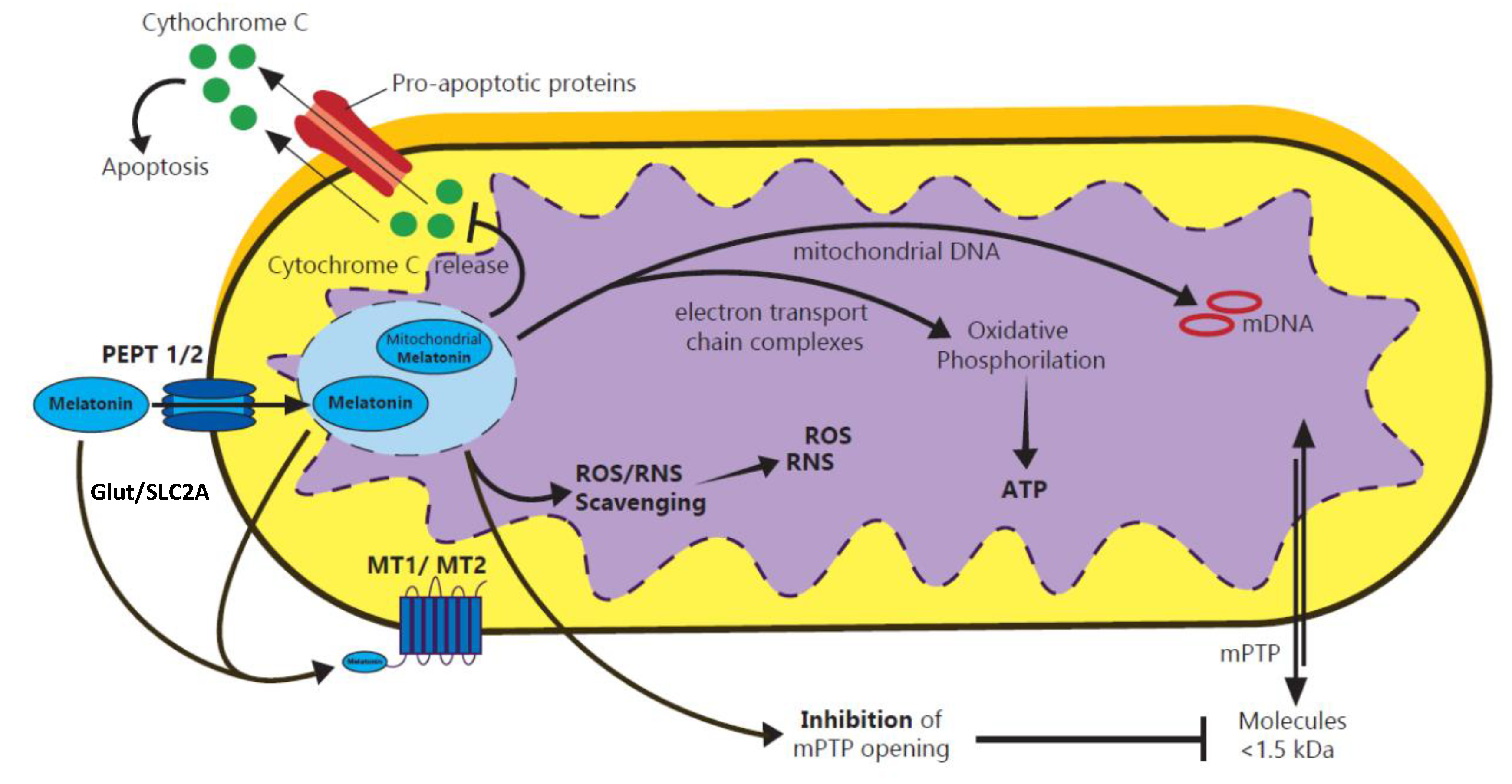
2.3. Antioxidative Effects of Melatonin in Embryo-Fetal Development: Function and Therapeutic Approaches
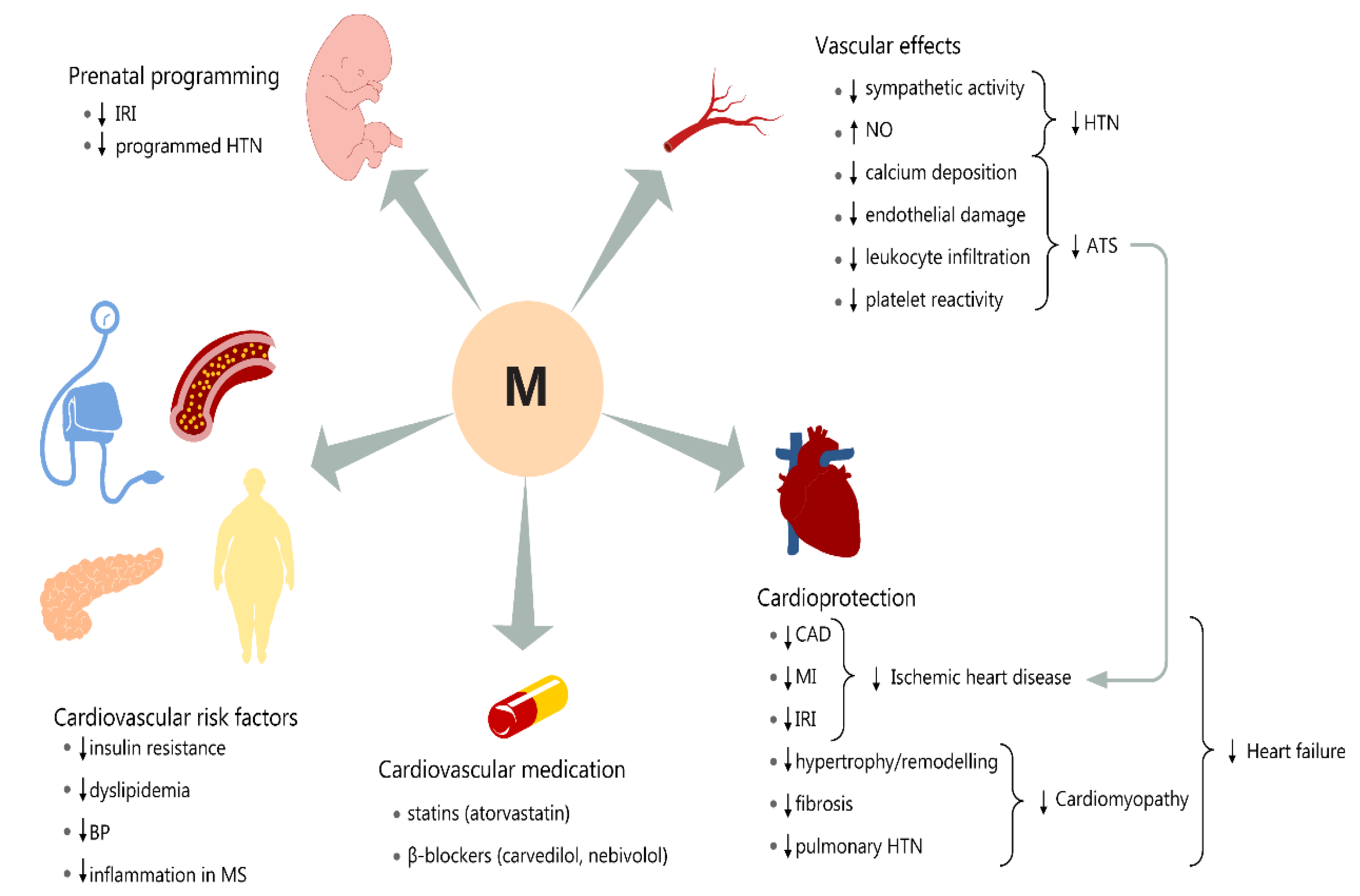
3. Melatonin’s Role in Programming Adult Cardiovascular Homeostasis and Allostasis
3.1. Prenatal and Neonatal Cardiovascular Effects
3.2. Antioxidant and Anti-Inflammatory Roles in Cardiovascular Pathologies
3.3. Effect on Adult Cardiovascular Risk Factors
3.4. Vascular and Myocardial Effects
3.5. Neutral and Deleterious Effects
4. Melatonin Effects in Neuronal Inflammation and Degenerative Neurological Diseases
4.1. Alzheimer’s Disease
4.2. Parkinson’s Disease
5. Melatonin as an Adjuvant in Respiratory Distress Caused by SARS-CoV-2
6. Promising Clinical Targets And Pending Questions
Author Contributions
Funding
Conflicts of Interest
References
- Acuña-Castroviejo, D.; Escames, G.; Venegas, C.; Díaz-Casado, M.E.; Lima-Cabello, E.; López, L.C.; Rosales-Corral, S.; Tan, D.X.; Reiter, R.J. Extrapineal Melatonin: Sources, Regulation, and Potential Functions. Cell. Mol. Life Sci. 2014, 71, 2997–3025. [Google Scholar] [CrossRef] [PubMed]
- Jiki, Z.; Lecour, S.; Nduhirabandi, F. Cardiovascular Benefits of Dietary Melatonin: A Myth or a Reality? Front. Physiol. 2018, 528, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Venegas, C.; García, J.A.; Escames, G.; Ortiz, F.; López, A.; Doerrier, C.; García-Corzo, L.; López, L.C.; Reiter, R.J.; Acuña-Castroviejo, D. Extrapineal Melatonin: Analysis of Its Subcellular Distribution and Daily Fluctuations. J. Pineal Res. 2012, 52, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Scholtens, R.M.; Van Munster, B.C.; Van Kempen, M.F.; De Rooij, S.E.J.A. Physiological Melatonin Levels in Healthy Older People: A Systematic Review. J. Psychosom. Res. 2016, 86, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Hickie, I.B.; Rogers, N.L. Novel Melatonin-Based Therapies: Potential Advances in the Treatment of Major Depression. Lancet 2011, 378, 621–631. [Google Scholar] [CrossRef]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an Antioxidant: Under Promises but over Delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef]
- Poeggeler, B.; Reiter, R.J.; Tan, D.-X.; Chen, L.-D.; Manchester, L.C. Melatonin, Hydroxyl Radical-mediated Oxidative Damage, and Aging: A Hypothesis. J. Pineal Res. 1993, 14, 151–168. [Google Scholar] [CrossRef]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin: A Versatile Protector against Oxidative DNA Damage. Molecules 2018, 23, 530. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Diduk, R.; Galano, A.; Tan, D.X.; Reiter, R.J. N-Acetylserotonin and 6-Hydroxymelatonin against Oxidative Stress: Implications for the Overall Protection Exerted by Melatonin. J. Phys. Chem. B 2015, 119, 8535–8543. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Paredes, S.D.; Korkmaz, A.; Sainz, R.M.; Mayo, J.C.; Fuentes-Broto, L.; Reiter, R.J. The Changing Biological Roles of Melatonin during Evolution: From an Antioxidant to Signals of Darkness, Sexual Selection and Fitness. Biol. Rev. 2010, 85, 607–623. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Qin, L.; Reiter, R.J. Melatonin: A Mitochondrial Targeting Molecule Involving Mitochondrial Protection and Dynamics. Int. J. Mol. Sci. 2016, 17, 2124. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Wang, C.; Yu, Z.; Peng, Y.; Wang, S.; Feng, S.; Zhang, S.; Tian, X.; Sun, C.; Liu, K.; et al. Human Transporters, PEPT1/2, Facilitate Melatonin Transportation into Mitochondria of Cancer Cells: An Implication of the Therapeutic Potential. J. Pineal Res. 2017, 62, e12390. [Google Scholar] [CrossRef] [PubMed]
- Mayo, J.C.; Sainz, R.M.; González-Menéndez, P.; Hevia, D.; Cernuda-Cernuda, R. Melatonin Transport into Mitochondria. Cell. Mol. Life Sci. 2017, 74, 3927–3940. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.M.; Nabavi, S.F.; Sureda, A.; Xiao, J.; Dehpour, A.R.; Shirooie, S.; Silva, A.S.; Baldi, A.; Khan, H.; Daglia, M. Anti-Inflammatory Effects of Melatonin: A Mechanistic Review. Crit. Rev. Food Sci. Nutr. 2018, 59, S4–S16. [Google Scholar] [CrossRef] [PubMed]
- El-Shenawy, S.M.; Abdel-Salam, O.M.E.; Baiuomy, A.R.; El-Batran, S.; Arbid, M.S. Studies on the Anti-Inflammatory and Anti-Nociceptive Effects of Melatonin in the Rat. Pharm. Res. 2002, 46, 235–243. [Google Scholar] [CrossRef]
- Carrasco, C.; Marchena, A.M.; Holguín-Arévalo, M.S.; Martín-Partido, G.; Rodríguez, A.B.; Paredes, S.D.; Pariente, J.A. Anti-Inflammatory Effects of Melatonin in a Rat Model of Caerulein-Induced Acute Pancreatitis. Cell Biochem. Funct. 2013, 31, 585–590. [Google Scholar] [CrossRef]
- Liu, Z.; Gan, L.; Xu, Y.; Luo, D.; Ren, Q.; Wu, S.; Sun, C. Melatonin Alleviates Inflammasome-Induced Pyroptosis through Inhibiting NF-ΚB/GSDMD Signal in Mice Adipose Tissue. J. Pineal Res. 2017, 63, e12414. [Google Scholar] [CrossRef]
- Deng, W.G.; Tang, S.T.; Tseng, H.P.; Wu, K.K. Melatonin Suppresses Macrophage Cyclooxygenase-2 and Inducible Nitric Oxide Synthase Expression by Inhibiting P52 Acetylation and Binding. Blood 2006, 108, 518–524. [Google Scholar] [CrossRef]
- Yang, Y.; Sun, Y.; Yi, W.; Li, Y.; Fan, C.; Xin, Z.; Jiang, S.; Di, S.; Qu, Y.; Reiter, R.J.; et al. A Review of Melatonin as a Suitable Antioxidant against Myocardial Ischemia-Reperfusion Injury and Clinical Heart Diseases. J. Pineal Res. 2014, 57, 357–366. [Google Scholar] [CrossRef]
- Opie, L.H.; Lecour, S. Melatonin Has Multiorgan Effects. Eur. Heart J. Cardiovasc. Pharm. 2016, 2, 258–265. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.H.; Xu, J.H.; Ren, Q.C.; Duan, T.; Mo, F.; Zhang, W. Melatonin Promotes Secondary Hair Follicle Development of Early Postnatal Cashmere Goat and Improves Cashmere Quantity and Quality by Enhancing Antioxidant Capacity and Suppressing Apoptosis. J. Pineal Res. 2019, 67, e12569. [Google Scholar] [CrossRef]
- Carbajo-Pescador, S.; Ordoñez, R.; Benet, M.; Jover, R.; García-Palomo, A.; Mauriz, J.L.; González-Gallego, J. Inhibition of VEGF Expression through Blockade of Hif1α and STAT3 Signalling Mediates the Anti-Angiogenic Effect of Melatonin in HepG2 Liver Cancer Cells. Br. J. Cancer 2013, 109, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, P.; Zheng, X.; Du, X. Therapeutic Strategies of Melatonin in Cancer Patients: A Systematic Review and Meta-Analysis. OncoTargets Ther. 2018, 11, 7895–7908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, C.M.; Figueiro, M.G. Measuring Light at Night and Melatonin Levels in Shift Workers: A Review of the Literature. Biol. Res. Nurs. 2017, 19, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Laudon, M.; Frydman-Marom, A. Therapeutic Effects of Melatonin Receptor Agonists on Sleep and Comorbid Disorders. Int. J. Mol. Sci. 2014, 15, 15924–15950. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Clough, S.J.; Hutchinson, A.J.; Adamah-Biassi, E.B.; Popovska-Gorevski, M.; Dubocovich, M.L. MT 1 and MT 2 Melatonin Receptors: A Therapeutic Perspective. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 361–383. [Google Scholar] [CrossRef] [Green Version]
- Atkin, T.; Comai, S.; Gobbi, G. Drugs for Insomnia beyond Benzodiazepines: Pharmacology, Clinical Applications, and Discovery. Pharmacol. Rev. 2018, 70, 197–245. [Google Scholar] [CrossRef]
- Andersen, L.P.H.; Gögenur, I.; Rosenberg, J.; Reiter, R.J. The Safety of Melatonin in Humans. Clin. Drug Investig. 2015, 36, 169–175. [Google Scholar] [CrossRef]
- Zagrean, A.-M.; Chitimus, D.M.; Badiu, C.; Panaitescu, A.M.; Peltecu, G.; Zagrean, L. The Pineal Gland and Its Function in Pregnancy and Lactation. In Maternal-Fetal and Neonatal Endocrinology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 15–37. [Google Scholar] [CrossRef]
- Nakamura, Y.; Tamura, H.; Kashida, S.; Takayama, H.; Yamagata, Y.; Karube, A.; Sugino, N.; Kato, H. Changes of Serum Melatonin Level and Its Relationship to Feto-Placental Unit during Pregnancy. J. Pineal Res. 2001, 30, 29–33. [Google Scholar] [CrossRef]
- Tamura, H.; Takayama, H.; Nakamura, Y.; Reiter, R.J.; Sugino, N. Fetal/Placental Regulation of Maternal Melatonin in Rats. J. Pineal Res. 2008, 44, 335–340. [Google Scholar] [CrossRef]
- Reik, W. Stability and Flexibility of Epigenetic Gene Regulation in Mammalian Development. Nature 2007, 447, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.; Hendrich, B.; Reik, W.; Dean, W. Dynamic Reprogramming of DNA Methylation in the Early Mouse Embryo. Dev. Biol. 2002, 241, 172–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Zhu, P.; Yan, L.; Li, R.; Hu, B.; Lian, Y.; Yan, J.; Ren, X.; Lin, S.; Li, J.; et al. The DNA Methylation Landscape of Human Early Embryos. Nature 2014, 511, 606–610. [Google Scholar] [CrossRef]
- Verma, N.; Pan, H.; Doré, L.C.; Shukla, A.; Li, Q.V.; Pelham-Webb, B.; Teijeiro, V.; González, F.; Krivtsov, A.; Chang, C.J.; et al. TET Proteins Safeguard Bivalent Promoters from de Novo Methylation in Human Embryonic Stem Cells. Nat. Genet. 2018, 50, 83–95. [Google Scholar] [CrossRef]
- Yang, M.; Tao, J.; Wu, H.; Guan, S.; Liu, L.; Zhang, L.; Deng, S.; He, C.; Ji, P.; Liu, J.; et al. Aanat Knockdown and Melatonin Supplementation in Embryo Development: Involvement of Mitochondrial Function and DNA Methylation. Antioxid. Redox Signal. 2019, 30, 2050–2065. [Google Scholar] [CrossRef]
- Koh, P.O. Melatonin Regulates the Calcium-Buffering Proteins, Parvalbumin and Hippocalcin, in Ischemic Brain Injury. J. Pineal Res. 2012, 53, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Cuzzocrea, S.; Costantino, G.; Gitto, E.; Mazzon, E.; Fulia, F.; Serraino, I.; Cordaro, S.; Barberi, I.; De Sarro, A.; Caputi, A.P. Protective Effects of Melatonin in Ischemic Brain Injury. J. Pineal Res. 2000, 29, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Lu, C.; Li, T.; Wang, W.; Ye, W.; Zeng, R.; Ni, L.; Lai, Z.; Wang, X.; Liu, C. The Protective Effect of Melatonin on Brain Ischemia and Reperfusion in Rats and Humans: In Vivo Assessment and a Randomized Controlled Trial. J. Pineal Res. 2018, 65, e12521. [Google Scholar] [CrossRef] [PubMed]
- Phillipson, O.T. Alpha-Synuclein, Epigenetics, Mitochondria, Metabolism, Calcium Traffic, & Circadian Dysfunction in Parkinson’s Disease. An Integrated Strategy for Management. Ageing Res. Rev. 2017, 40, 149–167. [Google Scholar] [CrossRef]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative Stress: The Mitochondria-Dependent and Mitochondria-Independent Pathways of Apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef]
- Chandra, J.; Samali, A.; Orrenius, S. Triggering and Modulation of Apoptosis by Oxidative Stress. Free Radic. Biol. Med. 2000, 29, 323–333. [Google Scholar] [CrossRef]
- Zhang, H.M.; Zhang, Y. Melatonin: A Well-Documented Antioxidant with Conditional pro-Oxidant Actions. J. Pineal Res. 2014, 57, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jiang, S.; Dong, Y.; Fan, C.; Zhao, L.; Yang, X.; Li, J.; Di, S.; Yue, L.; Liang, G.; et al. Melatonin Prevents Cell Death and Mitochondrial Dysfunction via a SIRT1-Dependent Mechanism during Ischemic-Stroke in Mice. J. Pineal Res. 2015, 58, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Giusti, P.; Upartiti, M.; Franceschini, D.; Schiavo, N.; Floreani, M.; Manev, H. Neuroprotection by Melatonin from Kainate-induced Excitotoxicity in Rats. FASEB J. 1996, 10, 891–896. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.L.; Bayraktutan, U. Oxidative Stress and Its Role in the Pathogenesis of Ischaemic Stroke. Int. J. Stroke 2009, 4, 461–470. [Google Scholar] [CrossRef]
- Cazevieille, C.; Safa, R.; Osborne, N.N. Melatonin Protects Primary Cultures of Rat Cortical Neurones from NMDA Excitotoxicity and Hypoxia/Reoxygenation. Brain Res. 1997, 768, 120–124. [Google Scholar] [CrossRef]
- Martín, M.; Macías, M.; Escames, G.; León, J.; Acuña-Castroviejo, D. Melatonin but Not Vitamins C and E Maintains Glutathione Homeostasis in T-butyl Hydroperoxide-induced Mitochondrial Oxidative Stress. FASEB J. 2000, 14, 1677–1679. [Google Scholar] [CrossRef]
- Loren, P.; Sánchez, R.; Arias, M.E.; Felmer, R.; Risopatrón, J.; Cheuquemán, C. Melatonin Scavenger Properties against Oxidative and Nitrosative Stress: Impact on Gamete Handling and In Vitro Embryo Production in Humans and Other Mammals. Int. J. Mol. Sci. 2017, 18, 1119. [Google Scholar] [CrossRef] [Green Version]
- García, J.J.; Lõpez-Pingarrõn, L.; Almeida-Souza, P.; Tres, A.; Escudero, P.; García-Gil, F.A.; Tan, D.X.; Reiter, R.J.; Ramírez, J.M.; Bernal-Pérez, M. Protective Effects of Melatonin in Reducing Oxidative Stress and in Preserving the Fluidity of Biological Membranes: A Review. J. Pineal Res. 2014, 56, 225–237. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Korkmaz, A.; Rosales-Corral, S.A. Melatonin and Stable Circadian Rhythms Optimize Maternal, Placental and Fetal Physiology. Hum. Reprod. Update 2014, 20, 293–307. [Google Scholar] [CrossRef] [Green Version]
- Welin, A.K.; Svedin, P.; Lapatto, R.; Sultan, B.; Hagberg, H.; Gressens, P.; Kjellmer, I.; Mallard, C. Melatonin Reduces Inflammation and Cell Death in White Matter in the Mid-Gestation Fetal Sheep Following Umbilical Cord Occlusion. Pediatr. Res. 2007, 61, 153–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhu, S.; Pei, Z.; Drozda, M.; Stavrovskaya, I.G.; Del Signore, S.J.; Cormier, K.; Shimony, E.M.; Wang, H.; Ferrante, R.J.; et al. Inhibitors of Cytochrome c Release with Therapeutic Potential for Huntington’s Disease. J. Neurosci. 2008, 28, 9473–9485. [Google Scholar] [CrossRef]
- Jan, J.E.; Wasdell, M.B.; Freeman, R.D.; Bax, M. Evidence Supporting the Use of Melatonin in Short Gestation Infants. J. Pineal Res. 2007, 42, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Schernhammer, E.S.; Kroenke, C.H.; Dowsett, M.; Folkerd, E.; Hankinson, S.E. Urinary 6-Sulfatoxymelatonin Levels and Their Correlations with Lifestyle Factors and Steroid Hormone Levels. J. Pineal Res. 2006, 40, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Tucker, J.; McGuire, W. ABC of Preterm Birth Epidemiology of Preterm Birth. BMJ 2004, 329, 675–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zetner, D.; Andersen, L.P.H.; Rosenberg, J. Pharmacokinetics of Alternative Administration Routes of Melatonin: A Systematic Review. Drug Res. 2015, 66, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Robertson, N.J.; Faulkner, S.; Fleiss, B.; Bainbridge, A.; Andorka, C.; Price, D.; Powell, E.; Lecky-Thompson, L.; Thei, L.; Chandrasekaran, M.; et al. Melatonin Augments Hypothermic Neuroprotection in a Perinatal Asphyxia Model. Brain 2013, 136, 90–105. [Google Scholar] [CrossRef]
- Petrosillo, G.; Colantuono, G.; Moro, N.; Ruggiero, F.M.; Tiravanti, E.; Di Venosa, N.; Fiore, T.; Paradies, G. Melatonin Protects against Heart Ischemia-Reperfusion Injury by Inhibiting Mitochondrial Permeability Transition Pore Opening. Am. J. Physiol. Circ. Physiol. 2009, 297, H1487–H1493. [Google Scholar] [CrossRef] [Green Version]
- Paradies, G.; Paradies, V.; Ruggiero, F.M.; Petrosillo, G. Protective Role of Melatonin in Mitochondrial Dysfunction and Related Disorders. Arch. Toxicol. 2015, 89, 923–939. [Google Scholar] [CrossRef]
- Petrosillo, G.; Moro, N.; Ruggiero, F.M.; Paradies, G. Melatonin Inhibits Cardiolipin Peroxidation in Mitochondria and Prevents the Mitochondrial Permeability Transition and Cytochrome c Release. Free Radic. Biol. Med. 2009, 47, 969–974. [Google Scholar] [CrossRef]
- Yang, Y.; Duan, W.; Jin, Z.; Yi, W.; Yan, J.; Zhang, S.; Wang, N.; Liang, Z.; Li, Y.; Chen, W.; et al. JAK2/STAT3 Activation by Melatonin Attenuates the Mitochondrial Oxidative Damage Induced by Myocardial Ischemia/Reperfusion Injury. J. Pineal Res. 2013, 55, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Yeung, H.M.; Hung, M.W.; Fung, M.L. Melatonin Ameliorates Calcium Homeostasis in Myocardial and Ischemia-Reperfusion Injury in Chronically Hypoxic Rats. J. Pineal Res. 2008, 45, 373–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Li, L.N.; Guo, S.; Zhao, X.Y.; Liu, Y.Z.; Liang, C.; Tu, S.; Wang, D.; Li, L.; Dong, J.Z.; et al. Melatonin Improves Cardiac Function in a Mouse Model of Heart Failure with Preserved Ejection Fraction. Redox Biol. 2018, 18, 211–221. [Google Scholar] [CrossRef]
- Javanmard, S.H.; Ziaei, A.; Ziaei, S.; Ziaei, E.; Mirmohammad-Sadeghi, M. The Effect of Preoperative Melatonin on Nuclear Erythroid 2-Related Factor 2 Activation in Patients Undergoing Coronary Artery Bypass Grafting Surgery. Oxidative Med. Cell. Longev. 2013, 2013, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, F.; García, J.A.; Acuña-Castroviejo, D.; Doerrier, C.; Lõpez, A.; Venegas, C.; Volt, H.; Luna-Sánchez, M.; Lõpez, L.C.; Escames, G. The Beneficial Effects of Melatonin against Heart Mitochondrial Impairment during Sepsis: Inhibition of i NOS and Preservation of n NOS. J. Pineal Res. 2014, 56, 71–81. [Google Scholar] [CrossRef]
- Veneroso, C.; Tuñón, M.J.; González-Gallego, J.; Collado, P.S. Melatonin Reduces Cardiac Inflammatory Injury Induced by Acute Exercise. J. Pineal Res. 2009, 47, 184–191. [Google Scholar] [CrossRef]
- Oliveira, L.G.R.; Kuehn, C.C.; Dos Santos, C.D.; Miranda, M.A.; Da Costa, C.M.B.; Mendonça, V.J.; Do Prado, J.C. Protective Actions of Melatonin against Heart Damage during Chronic Chagas Disease. Acta Trop. 2013, 128, 652–658. [Google Scholar] [CrossRef]
- Permpoonputtana, K.; Tangweerasing, P.; Mukda, S.; Boontem, P.; Nopparat, C.; Govitrapong, P. Long-Term Administration of Melatonin Attenuates Neuroinflammation in the Aged Mouse Brain. EXCLI J. 2018, 17, 634–646. [Google Scholar] [CrossRef]
- Wu, X.; Ji, H.; Wang, Y.; Gu, C.; Gu, W.; Hu, L.; Zhu, L. Melatonin Alleviates Radiation-Induced Lung Injury via Regulation of MiR-30e/NLRP3 Axis. Oxidative Med. Cell. Longev. 2019, 2019, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.C.; Peng, C.K.; Liao, W.I.; Pao, H.P.; Huang, K.L.; Chu, S.J. Melatonin Receptor Agonist Protects against Acute Lung Injury Induced by Ventilator through Up-Regulation of IL-10 Production. Respir. Res. 2020, 21, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Shin, N.R.; Ko, J.W.; Kim, J.C.; Park, G.; Kim, S.H.; Kim, M.S.; Kim, J.S.; Shin, I.S. Role of Melatonin as an SIRT1 Enhancer in Chronic Obstructive Pulmonary Disease Induced by Cigarette Smoke. J. Cell. Mol. Med. 2020, 24, 1151–1156. [Google Scholar] [CrossRef] [Green Version]
- Zagrean, A.-M.; Chitimus, D.M.; Paslaru, F.G.; Voiculescu, S.E.; Badiu, C.; Peltecu, G.; Panaitescu, A.M. Pineal Gland Disorders and Circadian Rhythm Alterations in Pregnancy and Lactation. In Maternal-Fetal and Neonatal Endocrinology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 241–257. [Google Scholar] [CrossRef]
- Zhu, J.L.; Hjollund, N.H.; Olsen, J. Shift Work, Duration of Pregnancy, and Birth Weight: The National Birth Cohort in Denmark. Am. J. Obstet. Gynecol. 2004, 191, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Wierrani, F.; Grin, W.; Hlawka, B.; Kroiss, A.; Grünberger, W. Elevated Serum Melatonin Levels during Human Late Pregnancy and Labour. J. Obstet. Gynaecol. 1997, 17, 449–451. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tamura, H.; Tan, D.X.; Xu, X.Y. Melatonin and the Circadian System: Contributions to Successful Female Reproduction. Fertil. Steril. 2014, 102, 321–328. [Google Scholar] [CrossRef]
- Okatani, Y.; Hayashi, K.; Sagara, Y. Effect of Estrogen on Melatonin Synthesis in Female Peripubertal Rats as Related to Adenylate Cyclase Activity. J. Pineal Res. 1998, 25, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Kennaway, D.J. Programming of the Fetal Suprachiasmatic Nucleus and Subsequent Adult Rhythmicity. Trends Endocrinol. Metab. 2002, 13, 398–402. [Google Scholar] [CrossRef]
- Conway, S.; Drew, J.E.; Canning, S.J.; Barrett, P.; Jockers, R.; Strosberg, A.D.; Guardiola-Lemaitre, B.; Delagrange, P.; Morgan, P.J. Identification of Mel1a Melatonin Receptors in the Human Embryonic Kidney Cell Line HEK293: Evidence of G Protein-Coupled Melatonin Receptors Which Do Not Mediate the Inhibition of Stimulated Cyclic AMP Levels. FEBS Lett. 1997, 407, 121–126. [Google Scholar] [CrossRef]
- Yu, X.; Li, Z.; Zheng, H.; Ho, J.; Chan, M.T.V.; Wu, W.K.K. Protective Roles of Melatonin in Central Nervous System Diseases by Regulation of Neural Stem Cells. Cell Prolif. 2017, 50, e12323. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Tain, Y.L. Light and Circadian Signaling Pathway in Pregnancy: Programming of Adult Health and Disease. Int. J. Mol. Sci. 2020, 21, 2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahoney, M.M. Shift Work, Jet Lag, and Female Reproduction. Int. J. Endocrinol. 2010, 2010, 813764. [Google Scholar] [CrossRef]
- Hardeland, R.; Madrid, J.A.; Tan, D.-X.; Reiter, R.J. Melatonin, the Circadian Multioscillator System and Health: The Need for Detailed Analyses of Peripheral Melatonin Signaling. J. Pineal Res. 2012, 52, 139–166. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Teoh, A.N.; Shukri, N.H.M.; Shafie, S.R.; Bustami, N.A.; Takahashi, M.; Lim, P.J.; Shibata, S. Circadian Rhythm and Its Association with Birth and Infant Outcomes: Research Protocol of a Prospective Cohort Study. BMC Pregnancy Childbirth 2020, 20, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzer, D.; Pauli, L.; Mueller, A.; Reutter, H.; Reinsberg, J.; Fimmers, R.; Bartmann, P.; Bagci, S. Melatonin Concentrations and Antioxidative Capacity of Human Breast Milk According to Gestational Age and the Time of Day. J. Hum. Lact. 2016, 32, NP105–NP110. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Shi, W.; Zhuang, J.; Liu, Y.; Tang, L.; Bu, J.; Sun, J.; Bei, F. Variations in Melatonin Levels in Preterm and Term Human Breast Milk during the First Month after Delivery. Sci. Rep. 2019, 9, 1–5. [Google Scholar] [CrossRef]
- Cai, C.; Vandermeer, B.; Khurana, R.; Nerenberg, K.; Featherstone, R.; Sebastianski, M.; Davenport, M.H. The Impact of Occupational Shift Work and Working Hours during Pregnancy on Health Outcomes: A Systematic Review and Meta-Analysis. Am. J. Obstet. Gynecol. 2019, 221, 563–576. [Google Scholar] [CrossRef]
- Stevens, R.G.; Schernhammer, E. Epidemiology of Urinary Melatonin in Women and Its Relation to Other Hormones and Night Work. Cancer Epidemiol. Biomark. Prev. 2005, 14, 551. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Song, H.; Dash, O.; Park, M.; Shin, N.E.; McLane, M.W.; Lei, J.; Hwang, J.Y.; Burd, I. Administration of Melatonin for Prevention of Preterm Birth and Fetal Brain Injury Associated with Premature Birth in a Mouse Model. Am. J. Reprod. Immunol. 2019, 82, e13151. [Google Scholar] [CrossRef]
- Jahnke, G.; Marr, M.; Myers, C.; Wilson, R.; Travlos, G.; Price, C. Maternal and Developmental Toxicity Evaluation of Melatonin Administered Orally to Pregnant Sprague-Dawley Rats. Toxicol. Sci. 1999, 50, 271–279. [Google Scholar] [CrossRef]
- Freeman, M.P.; Sosinsky, A.Z.; Moustafa, D.; Viguera, A.C.; Cohen, L.S. Supplement Use by Women during Pregnancy: Data from the Massachusetts General Hospital National Pregnancy Registry for Atypical Antipsychotics. Arch. Womens Ment. Health 2016, 19, 437–441. [Google Scholar] [CrossRef]
- Karumanchi, S.A.; Granger, J.P. Preeclampsia and Pregnancy-Related Hypertensive Disorders. Hypertension 2016, 67, 238–242. [Google Scholar] [CrossRef]
- Zeng, K.; Gao, Y.; Wan, J.; Tong, M.; Lee, A.C.; Zhao, M.; Chen, Q. The Reduction in Circulating Levels of Melatonin May Be Associated with the Development of Preeclampsia. J. Hum. Hypertens. 2016, 30, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Hobson, S.R.; Gurusinghe, S.; Lim, R.; Alers, N.O.; Miller, S.L.; Kingdom, J.C.; Wallace, E.M. Melatonin Improves Endothelial Function in Vitro and Prolongs Pregnancy in Women with Early-Onset Preeclampsia. J. Pineal Res. 2018, 65, e12508. [Google Scholar] [CrossRef] [PubMed]
- Hannan, N.J.; Binder, N.K.; Beard, S.; Nguyen, T.V.; Kaitu’u-Lino, T.J.; Tong, S. Melatonin Enhances Antioxidant Molecules in the Placenta, Reduces Secretion of Soluble Fms-like Tyrosine Kinase 1 (SFLT) from Primary Trophoblast but Does Not Rescue Endothelial Dysfunction: An Evaluation of Its Potential to Treat Preeclampsia. PLoS ONE 2018, 13, e0187082. [Google Scholar] [CrossRef] [PubMed]
- Hobson, S.R.; Lim, R.; Gardiner, E.E.; Alers, N.O.; Wallace, E.M. Phase I Pilot Clinical Trial of Antenatal Maternally Administered Melatonin to Decrease the Level of Oxidative Stress in Human Pregnancies Affected by Pre-Eclampsia (PAMPR): Study Protocol. BMJ Open 2013, 3, e003788. [Google Scholar] [CrossRef] [Green Version]
- Nutt, J.G.; Bohnen, N.I. Non-Dopaminergic Therapies. J. Park. Dis. 2018, 8, S73–S78. [Google Scholar] [CrossRef] [Green Version]
- Cetin, I.; Alvino, G. Intrauterine Growth Restriction: Implications for Placental Metabolism and Transport. A Review. Placenta 2009, 30, 77–82. [Google Scholar] [CrossRef]
- Miller, S.L.; Yan, E.B.; Castillo-Meléndez, M.; Jenkin, G.; Walker, D.W. Melatonin Provides Neuroprotection in the Late-Gestation Fetal Sheep Brain in Response to Umbilical Cord Occlusion. Dev. Neurosci. 2005, 27, 200–210. [Google Scholar] [CrossRef]
- Wang, L.; Feng, C.; Zheng, X.; Guo, Y.; Zhou, F.; Shan, D.; Liu, X.; Kong, J. Plant Mitochondria Synthesize Melatonin and Enhance the Tolerance of Plants to Drought Stress. J. Pineal Res. 2017, 63, e12429. [Google Scholar] [CrossRef]
- Voiculescu, S.E.; Zygouropoulos, N.; Zahiu, C.D.; Zagrean, A.M. Role of Melatonin in Embryo Fetal Development. J. Med. Life. 2014, 7, 488–492. [Google Scholar]
- Rodriguez-Osorio, N.; Kim, I.J.; Wang, H.; Kaya, A.; Memili, E. Melatonin Increases Cleavage Rate of Porcine Preimplantation Embryos in Vitro. J. Pineal Res. 2007, 43, 283–288. [Google Scholar] [CrossRef]
- Sampaio, R.V.; Conceição, D.S.B.; Miranda, M.S.; Sampaio, L.d.F.S.; Ohashi, O.M. MT3 Melatonin Binding Site, MT1 and MT2 Melatonin Receptors Are Present in Oocyte, but Only MT1 Is Present in Bovine Blastocyst Produced In Vitro. Reprod. Biol. Endocrinol. 2012, 10, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Tian, X.; Zhang, L.; Tan, D.; Reiter, R.J.; Liu, G. Melatonin Promotes the in Vitro Development of Pronuclear Embryos and Increases the Efficiency of Blastocyst Implantation in Murine. J. Pineal Res. 2013, 55, 267–274. [Google Scholar] [CrossRef]
- Schwimmer, H.; Metzer, A.; Pilosof, Y.; Szyf, M.; Machnes, Z.M.; Fares, F.; Harel, O.; Haim, A. Light at Night and Melatonin Have Opposite Effects on Breast Cancer Tumors in Mice Assessed by Growth Rates and Global DNA Methylation. Chronobiol. Int. 2014, 31, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Smith, Z.D.; Meissner, A. DNA Methylation: Roles in Mammalian Development. Nat. Rev. Genet. 2013, 14, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, J.; González, B.; Sempere, V.; Mas, A.; Torija, M.J.; Beltran, G. Melatonin Reduces Oxidative Stress Damage Induced by Hydrogen Peroxide in Saccharomyces Cerevisiae. Front. Microbiol. 2017, 8, 1066. [Google Scholar] [CrossRef] [PubMed]
- Gitto, E.; Aversa, S.; Reiter, R.J.; Barberi, I.; Pellegrino, S. Update on the Use of Melatonin in Pediatrics. J. Pineal Res. 2011, 50, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Gitto, E.; Pellegrino, S.; Gitto, P.; Barberi, I.; Reiter, R.J. Oxidative Stress of the Newborn in the Pre- and Postnatal Period and the Clinical Utility of Melatonin. J. Pineal Res. 2009, 46, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L.; Purvis, C.C.; Drew, J.E.; Abramovich, D.R.; Williams, L.M. Melatonin Receptors in Human Fetal Brain: 2-[125I]Iodomelatonin Binding and MT1 Gene Expression. J. Pineal Res. 2002, 33, 218–224. [Google Scholar] [CrossRef]
- McDonald, C.A.; Penny, T.R.; Paton, M.C.B.; Sutherland, A.E.; Nekkanti, L.; Yawno, T.; Castillo-Melendez, M.; Fahey, M.C.; Jones, N.M.; Jenkin, G.; et al. Effects of Umbilical Cord Blood Cells, and Subtypes, to Reduce Neuroinflammation Following Perinatal Hypoxic-Ischemic Brain Injury. J. Neuroinflamm. 2018, 15, 47. [Google Scholar] [CrossRef]
- Cao, W.; Carney, J.M.; Duchon, A.; Floyd, R.A.; Chevion, M. Oxygen Free Radical Involvement in Ischemia and Reperfusion Injury to Brain. Neurosci. Lett. 1988, 88, 233–238. [Google Scholar] [CrossRef]
- Jang, M.H.; Jung, S.B.; Lee, M.H.; Kim, C.J.; Oh, Y.T.; Kang, I.; Kim, J.; Kim, E.H. Melatonin Attenuates Amyloid Beta25-35-Induced Apoptosis in Mouse Microglial BV2 Cells. Neurosci. Lett. 2005, 380, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Ebadi, M.; Sharma, S.K.; Ghafourifar, P.; Brown-Borg, H.; El ReFaey, H. Peroxynitrite in the Pathogenesis of Parkinson’s Disease and the Neuroprotective Role of Metallothioneins. Methods Enzymol. 2005, 396, 276–298. [Google Scholar] [CrossRef] [PubMed]
- Aly, H.; Elmahdy, H.; El-Dib, M.; Rowisha, M.; Awny, M.; El-Gohary, T.; Elbatch, M.; Hamisa, M.; El-Mashad, A.R. Melatonin Use for Neuroprotection in Perinatal Asphyxia: A Randomized Controlled Pilot Study. J. Perinatol. 2015, 35, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Dayoub, J.C.; Ortiz, F.; Lõpez, L.C.; Venegas, C.; Del Pino-Zumaquero, A.; Roda, O.; Sánchez-Montesinos, I.; Acuña-Castroviejo, D.; Escames, G. Synergism between Melatonin and Atorvastatin against Endothelial Cell Damage Induced by Lipopolysaccharide. J. Pineal Res. 2011, 51, 324–330. [Google Scholar] [CrossRef]
- Hu, S.; Zhu, P.; Zhou, H.; Zhang, Y.; Chen, Y. Melatonin-Induced Protective Effects on Cardiomyocytes against Reperfusion Injury Partly through Modulation of IP3R and SERCA2a via Activation of ERK1. Arq. Bras. Cardiol. 2018, 110, 44–51. [Google Scholar] [CrossRef]
- Giacomo, C.G.; Antonio, M. Melatonin in Cardiac Ischemia/Reperfusion-Induced Mitochondrial Adaptive Changes. Cardiovasc. Hematol. Disord. Drug Targets 2007, 7, 163–169. [Google Scholar] [CrossRef]
- Qi, X.; Wang, J. Melatonin Improves Mitochondrial Biogenesis through the AMPK / PGC1α Pathway to Attenuate Ischemia/Reperfusion -Induced Myocardial Damage. Aging 2020, 12, 1–14. [Google Scholar] [CrossRef]
- Stroethoff, M.; Goetze, L.; Torregroza, C.; Bunte, S.; Raupach, A.; Heinen, A.; Mathes, A.; Hollmann, M.W.; Huhn, R. The Melatonin Receptor Agonist Ramelteon Induces Cardioprotection That Requires MT2 Receptor Activation and Release of Reactive Oxygen Species. Cardiovasc. Drugs Ther. 2020, 34, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Ma, Q.; Zhu, P.; Ren, J.; Reiter, R.J.; Chen, Y. Protective Role of Melatonin in Cardiac Ischemia-Reperfusion Injury: From Pathogenesis to Targeted Therapy. J. Pineal Res. 2018, 64, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Baker, J.; Kimpinski, K. Role of melatonin in blood pressure regulation: An adjunct anti-hypertensive agent. Clin. Exp. Pharmacol. Physiol. 2018, 45, 755–766. [Google Scholar] [CrossRef]
- Pechanova, O.; Paulis, L.; Simko, F. Peripheral and Central Effects of Melatonin on Blood Pressure Regulation. Int. J. Mol. Sci. 2014, 15, 17920–17937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardinali, D.P.; Hardeland, R. Inflammaging, Metabolic Syndrome and Melatonin: A Call for Treatment Studies. Neuroendocrinology 2017, 104, 382–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P.; De La Torre-Hernandez, J.M.; Gonzalez-Gonzalez, J.; Garcia-Camarero, T.; Consuegra-Sanchez, L.; Garcia-Saiz, M.d.M.; Aldea-Perona, A.; Virgos-Aller, T.; Azpeitia, A.; et al. Effect of Intravenous and Intracoronary Melatonin as an Adjunct to Primary Percutaneous Coronary Intervention for Acute ST-Elevation Myocardial Infarction: Results of the Melatonin Adjunct in the Acute MyocaRdial Infarction Treated with Angioplasty Trial. J. Pineal Res. 2017, 62, e12374. [Google Scholar] [CrossRef] [PubMed]
- Ekeløf, S.V.; Halladin, N.L.; Jensen, S.E.; Zaremba, T.; Aarøe, J.; Kjærgaard, B.; Simonsen, C.W.; Rosenberg, J.; Gögenur, I. Effects of Intracoronary Melatonin on Ischemia–Reperfusion Injury in ST-Elevation Myocardial Infarction. Heart Vessel. 2016, 31, 88–95. [Google Scholar] [CrossRef]
- Virag, J.; Lust, R. Circadian Influences on Myocardial Infarction. Front. Physiol. 2014, 5, 1–10. [Google Scholar] [CrossRef]
- Singhanat, K.; Apaijai, N.; Chattipakorn, S.C.; Chattipakorn, N. Roles of Melatonin and Its Receptors in Cardiac Ischemia—Reperfusion Injury. Cell. Mol. Life Sci. 2018, 75, 4125–4149. [Google Scholar] [CrossRef]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P.; Reiter, R.J. Melatonin for Cardioprotection in ST Elevation Myocardial Infarction: Are We Ready for the Challenge? Heart 2016, 103, 647–648. [Google Scholar] [CrossRef]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P.; Arroyo-Ucar, E.; Reiter, R.J. Decreased Level of Melatonin in Serum Predicts Left Ventricular Remodelling after Acute Myocardial Infarction. J. Pineal Res. 2012, 53, 319–323. [Google Scholar] [CrossRef]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P.; De La Torre-Hernandez, J.M.; Consuegra-Sanchez, L.; Piccolo, R.; Gonzalez-Gonzalez, J.; Garcia-Camarero, T.; Del Mar Garcia-Saiz, M.; Aldea-Perona, A.; Reiter, R.J.; et al. Usefulness of Early Treatment With Melatonin to Reduce Infarct Size in Patients With ST-Segment Elevation Myocardial Infarction Receiving Percutaneous Coronary Intervention (From the Melatonin Adjunct in the Acute Myocardial Infarction Treated With Angioplasty Trial). Am. J. Cardiol. 2017, 120, 522–526. [Google Scholar] [CrossRef]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P.; Piccolo, R.; Galasso, G.; Reiter, R.J. Melatonin Is Associated with Reverse Remodeling after Cardiac Resynchronization Therapy in Patients with Heart Failure and Ventricular Dyssynchrony. Int. J. Cardiol. 2016, 221, 359–363. [Google Scholar] [CrossRef]
- Misaka, T.; Yoshihisa, A.; Yokokawa, T.; Sato, T.; Oikawa, M.; Kobayashi, A.; Yamaki, T.; Sugimoto, K.; Kunii, H.; Nakazato, K.; et al. Plasma Levels of Melatonin in Dilated Cardiomyopathy. J. Pineal Res. 2019, 66, e12564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, H.; Wang, Y.; Li, H.; Li, Y.; Sahil, A.; Lv, J.; Liu, Y.; Yang, Z.; Dong, R.; Xue, H.; et al. Melatonin Alleviates Cardiac Fibrosis via Inhibiting LncRNA MALAT1/MiR-141-Mediated NLRP3 Inflammasome and TGF-Β1/Smads Signaling in Diabetic Cardiomyopathy. FASEB J. 2020, 34, 5282–5298. [Google Scholar] [CrossRef] [PubMed]
- Crnko, S.; Printezi, M.I.; Jansen, T.P.J.; Leiteris, L.; Van Der Meer, M.G.; Schutte, H.; Van Faassen, M.; Du Pré, B.C.; De Jonge, N.; Asselbergs, F.W.; et al. Prognostic Biomarker Soluble ST2 Exhibits Diurnal Variation in Chronic Heart Failure Patients. ESC Heart Fail. 2020, 7, 1224–1233. [Google Scholar] [CrossRef] [PubMed]
- Dwaich, K.H.; Al-Amran, F.G.Y.; AL-Sheibani, B.I.M.; Al-Aubaidy, H.A. Melatonin Effects on Myocardial Ischemia–Reperfusion Injury: Impact on the Outcome in Patients Undergoing Coronary Artery Bypass Grafting Surgery. Int. J. Cardiol. 2016, 221, 977–986. [Google Scholar] [CrossRef]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P. Future Strategies for Acute Cardioprotection: “Melatonin as Promising Therapy.”. Cardiovasc. Res. 2017, 113, 1418. [Google Scholar] [CrossRef]
- Popescu, M.R.; Panaitescu, A.M.; Pavel, B.; Zagrean, L.; Peltecu, G.; Zagrean, A.M. Getting an Early Start in Understanding Perinatal Asphyxia Impact on the Cardiovascular System. Front. Pediatr. 2020, 8, 68. [Google Scholar] [CrossRef]
- Tare, M.; Parkington, H.C.; Wallace, E.M.; Sutherland, A.E.; Lim, R.; Yawno, T.; Coleman, H.A.; Jenkin, G.; Miller, S.L. Maternal Melatonin Administration Mitigates Coronary Stiffness and Endothelial Dysfunction, and Improves Heart Resilience to Insult in Growth Restricted Lambs. J. Physiol. 2014, 592, 2695–2709. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.L.; Huang, L.T.; Hsu, C.N. Developmental Programming of Adult Disease: Reprogramming by Melatonin? Int. J. Mol. Sci. 2017, 18, 426. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.L.; Huang, L.T.; Hsu, C.N.; Lee, C.-T. Melatonin Therapy Prevents Programmed Hypertension and Nitric Oxide Deficiency in Offspring Exposed to Maternal Caloric Restriction. Oxidative Med. Cell. Longev. 2014, 2014, 1–21. [Google Scholar] [CrossRef]
- Tain, Y.L.; Chan, J.Y.H.; Lee, C.-T.; Hsu, C.N. Maternal Melatonin Therapy Attenuates Methyl-Donor Diet-Induced Programmed Hypertension in Male Adult Rat Offspring. Nutrients 2018, 10, 1407. [Google Scholar] [CrossRef] [Green Version]
- Mendez, N.; Halabi, D.; Spichiger, C.; Salazar, E.R.; Vergara, K.; Alonso-Vasquez, P.; Carmona, P.; Sarmiento, J.M.; Richter, H.G.; Seron-Ferre, M.; et al. Gestational Chronodisruption Impairs Circadian Physiology in Rat Male Offspring, Increasing the Risk of Chronic Disease. Endocrinology 2016, 157, 4654–4668. [Google Scholar] [CrossRef] [PubMed]
- Alamili, M.; Bendtzen, K.; Lykkesfeldt, J.; Rosenberg, J.; Gögenur, I. Melatonin Suppresses Markers of Inflammation and Oxidative Damage in a Human Daytime Endotoxemia Model. J. Crit. Care 2014, 29, 184.e9–184.e13. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P.; Garcia, M.; Ferrer, J.; De La Rosa, A.; Vargas, M.; Reiter, R.J. Light/Dark Patterns of Interleukin-6 in Relation to the Pineal Hormone Melatonin in Patients with Acute Myocardial Infarction. Cytokine 2004, 26, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Rodriguez, A.; Garcia-Gonzalez, M.; Abreu-Gonzalez, P.; Ferrer, J.; Kaski, J.C. Relation of Nocturnal Melatonin Levels to C-Reactive Protein Concentration in Patients with ST-Segment Elevation Myocardial Infarction. Am. J. Cardiol. 2006, 97, 10–12. [Google Scholar] [CrossRef]
- Libby, P. Inflammation in Atherosclerosis. Arter. Thromb. Vasc. Biol. 2012, 32, 2045–2051. [Google Scholar] [CrossRef] [Green Version]
- Bäck, M.; Yurdagul, A.; Tabas, I.; Öörni, K.; Kovanen, P.T. Inflammation and Its Resolution in Atherosclerosis: Mediators and Therapeutic Opportunities. Nat. Rev. Cardiol. 2019, 1, 389–406. [Google Scholar] [CrossRef]
- Oxenkrug, G.F.; Summergrad, P. Ramelteon Attenuates Age-Associated Hypertension and Weight Gain in Spontaneously Hypertensive Rats. In Annals of the New York Academy of Sciences; Blackwell Publishing Inc.: New York, NY, USA, 2010; Volume 1199, pp. 114–120. [Google Scholar] [CrossRef]
- She, M.; Hu, X.; Su, Z.; Zhang, C.; Yang, S.; Ding, L.; Laudon, M.; Yin, W. Piromelatine, a Novel Melatonin Receptor Agonist, Stabilizes Metabolic Profiles and Ameliorates Insulin Resistance in Chronic Sleep Restricted Rats. Eur. J. Pharmacol. 2014, 727, 60–65. [Google Scholar] [CrossRef]
- Obayashi, K.; Saeki, K.; Kurumatani, N. Higher Melatonin Secretion Is Associated with Lower Leukocyte and Platelet Counts in the General Elderly Population: The Kyo-Heijo Cohort. J. Pineal Res. 2015, 58, 227–233. [Google Scholar] [CrossRef]
- Paredes, S.D.; Forman, K.A.; García, C.; Vara, E.; Escames, G.; Tresguerres, J.A.F. Protective Actions of Melatonin and Growth Hormone on the Aged Cardiovascular System. Horm. Mol. Biol. Clin. Investig. 2014, 18, 79–88. [Google Scholar] [CrossRef]
- Favero, G.; Franceschetti, L.; Buffoli, B.; Moghadasian, M.H.; Reiter, R.J.; Rodella, L.F.; Rezzani, R. Melatonin: Protection against Age-Related Cardiac Pathology. Ageing Res. Rev. 2017, 35, 336–349. [Google Scholar] [CrossRef]
- Obayashi, K.; Saeki, K.; Tone, N.; Kurumatani, N. Relationship between Melatonin Secretion and Nighttime Blood Pressure in Elderly Individuals with and without Antihypertensive Treatment: A Cross-Sectional Study of the HEIJO-KYO Cohort. Hypertens. Res. 2014, 37, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Korkmaz, A. The Circadian Melatonin Rhythm and Its Modulation: Possible Impact on Hypertension. J. Hypertens. 2009, 27, S17–S20. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Vgontzas, A.N.; Fernandez-Mendoza, J.; Bixler, E.O.; Sun, Y.; Zhou, J.; Ren, R.; Li, T.; Tang, X. Insomnia with Physiological Hyperarousal Is Associated with Hypertension. Hypertension 2015, 65, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Akbari, M.; Ostadmohammadi, V.; Tabrizi, R.; Lankarani, K.B.; Heydari, S.T.; Amirani, E.; Reiter, R.J.; Asemi, Z. The Effects of Melatonin Supplementation on Inflammatory Markers among Patients with Metabolic Syndrome or Related Disorders: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Inflammopharmacology 2018, 26, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Zephy, D.; Ahmad, J. Type 2 Diabetes Mellitus: Role of Melatonin and Oxidative Stress. Diabetes Metab. Syndr. Clin. Res. Rev. 2015, 9, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, A.; Ma, S.; Topal, T.; Rosales-Corral, S.; Tan, D.X.; Reiter, R.J. Glucose: A Vital Toxin and Potential Utility of Melatonin in Protecting against the Diabetic State. Mol. Cell. Endocrinol. 2012, 349, 128–137. [Google Scholar] [CrossRef]
- Rezzani, R.; Favero, G.; Stacchiotti, A.; Rodella, L.F. Endothelial and Vascular Smooth Muscle Cell Dysfunction Mediated by Cyclophylin A and the Atheroprotective Effects of Melatonin. Life Sci. 2013, 92, 875–882. [Google Scholar] [CrossRef]
- Mesri Alamdari, N.; Mahdavi, R.; Roshanravan, N.; Lotfi Yaghin, N.; Ostadrahimi, A.R.; Faramarzi, E. A Double-Blind, Placebo-Controlled Trial Related to the Effects of Melatonin on Oxidative Stress and Inflammatory Parameters of Obese Women. Horm. Metab. Res. 2014, 47, 504–508. [Google Scholar] [CrossRef] [Green Version]
- Favero, G.; Franco, C.; Stacchiotti, A.; Rodella, L.F.; Rezzani, R. Sirtuin1 Role in the Melatonin Protective Effects Against Obesity-Related Heart Injury. Front. Physiol. 2020, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- McMullan, C.J.; Rimm, E.B.; Schernhammer, E.S.; Forman, J.P. A Nested Case-Control Study of the Association between Melatonin Secretion and Incident Myocardial Infarction. Heart 2016, 103, 694–701. [Google Scholar] [CrossRef]
- Talafi Noghani, M.; Namdar, H. Migraine Associated with Gastrointestinal Disorders: A Pathophysiological Explanation. Med. Hypotheses 2019, 125, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Li, Y.; Han, H.; Chen, S.; Gao, J.; Liu, G.; Wu, X.; Deng, J.; Yu, Q.; Huang, X.; et al. Melatonin Reprogramming of Gut Microbiota Improves Lipid Dysmetabolism in High-Fat Diet-Fed Mice. J. Pineal Res. 2018, 65, e12524. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.P.; Fang, X.L.; Fang, N.; Wang, X.B.; Qian, H.Y.; Cao, Z.; Cheng, Y.; Wang, B.N.; Wang, Y. Melatonin Ameliorates Vascular Endothelial Dysfunction, Inflammation, and Atherosclerosis by Suppressing the TLR4/NF-ΚB System in High-Fat-Fed Rabbits. J. Pineal Res. 2013, 55, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.R.; Zhou, Y.J.; Yang, J.Q.; Liu, F.; Wu, X.P.; Sha, Y. Melatonin Attenuates Calcium Deposition from Vascular Smooth Muscle Cells by Activating Mitochondrial Fusion and Mitophagy via an AMPK/OPA1 Signaling Pathway. Oxidative Med. Cell. Longev. 2020, 2020, 1–23. [Google Scholar] [CrossRef]
- Xu, F.; Zhong, J.Y.; Lin, X.; Shan, S.K.; Guo, B.; Zheng, M.H.; Wang, Y.; Li, F.; Cui, R.R.; Wu, F.; et al. Melatonin Alleviates Vascular Calcification and Ageing through Exosomal MiR-204/MiR-211 Cluster in a Paracrine Manner. J. Pineal Res. 2020, 68, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Sallinen, P.; Mänttäri, S.; Leskinen, H.; Ilves, M.; Vakkuri, O.; Ruskoaho, H.; Saarela, S. The Effect of Myocardial Infarction on the Synthesis, Concentration and Receptor Expression of Endogenous Melatonin. J. Pineal Res. 2007, 42, 254–260. [Google Scholar] [CrossRef]
- Altun, A.; Yaprak, M.; Aktoz, M.; Vardar, A.; Betul, U.A.; Ozbay, G. Impaired Nocturnal Synthesis of Melatonin in Patients with Cardiac Syndrome X. Neurosci. Lett. 2002, 327, 143–145. [Google Scholar] [CrossRef]
- Domínguez-Rodríguez, A.; Abreu-González, P.; García, M.J.; Sanchez, J.; Marrero, F.; De Armas-Trujillo, D. Decreased Nocturnal Melatonin Levels during Acute Myocardial Infarction. J. Pineal Res. 2002, 33, 248–252. [Google Scholar] [CrossRef]
- Fernandes, P.A.; Tamura, E.K.; D’Argenio-Garcia, L.; Muxel, S.M.; Da Silveira Cruz-Machado, S.; Marçola, M.; Carvalho-Sousa, C.E.; Cecon, E.; Ferreira, Z.S.; Markus, R.P. Dual Effect of Catecholamines and Corticosterone Crosstalk on Pineal Gland Melatonin Synthesis. Neuroendocrinology 2016, 104, 126–134. [Google Scholar] [CrossRef]
- Yang, J.B.; Kang, Y.M.; Zhang, C.; Yu, X.J.; Chen, W.S. Infusion of Melatonin into the Paraventricular Nucleus Ameliorates Myocardial Ischemia-Reperfusion Injury by Regulating Oxidative Stress and Inflammatory Cytokines. J. Cardiovasc. Pharmacol. 2019, 74, 336–347. [Google Scholar] [CrossRef]
- Nduhirabandi, F.; Du Toit, E.F.; Blackhurst, D.; Marais, D.; Lochner, A. Chronic Melatonin Consumption Prevents Obesity-Related Metabolic Abnormalities and Protects the Heart against Myocardial Ischemia and Reperfusion Injury in a Prediabetic Model of Diet-Induced Obesity. J. Pineal Res. 2011, 50, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Li, H.R.; Wang, C.; Sun, P.; Liu, D.; Du, G.; Tian, J.W. Melatonin Attenuates Doxorubicin-Induced Cardiotoxicity through Preservation of YAP Expression. J. Cell. Mol. Med. 2020, 24, 3634–3646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoschitzky, K.; Stoschitzky, G.; Brussee, H.; Bonell, C.; Dobnig, H. Comparing Beta-Blocking Effects of Bisoprolol, Carvedilol and Nebivolol. Cardiology 2006, 106, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Stoschitzky, K.; Sakotnik, A.; Lercher, P.; Zweiker, R.; Maier, R.; Liebmann, P.; Lindner, W. Influence of Beta-Blockers on Melatonin Release. Eur. J. Clin. Pharmacol. 1999, 55, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Drobnik, J.; Owczarek, K.; Piera, L.; Tosik, D.; Olczak, S.; Ciosek, J.; Hrabec, E. Melatonin-Induced Augmentation of Collagen Deposition in Cultures of Fibroblasts and Myofibroblasts Is Blocked by Luzindole—A Melatonin Membrane Receptors Inhibitor. Pharmacol. Rep. 2013, 65, 642–649. [Google Scholar] [CrossRef]
- Drobnik, J.; Karbownik-Lewińska, M.; Szczepanowska, A.; Słotwińska, D.; Olczak, S.; Jakubowski, L.; Da̧browski, R. Regulatory Influence of Melatonin on Collagen Accumulation in the Infarcted Heart Scar. J. Pineal Res. 2008, 45, 285–290. [Google Scholar] [CrossRef]
- Drobnik, J.; Tosik, D.; Piera, L.; Szczepanowska, A.; Olczak, S.; Zielinska, A.; Liberski, P.P.; Ciosek, J. Melatonin-Induced Glycosaminoglycans Augmentation in Myocardium Remote to Infarction. J. Physiol. Pharm. 2013, 64, 737–744. [Google Scholar]
- Feigin, V.L.; Nichols, E.; Alam, T.; Bannick, M.S.; Beghi, E.; Blake, N.; Culpepper, W.J.; Dorsey, E.R.; Elbaz, A.; Ellenbogen, R.G.; et al. Global, Regional, and National Burden of Neurological Disorders, 1990–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 459–480. [Google Scholar] [CrossRef] [Green Version]
- Hurtado-Alvarado, G.; Domínguez-Salazar, E.; Pavon, L.; Velázquez-Moctezuma, J.; Gómez-González, B. Blood-Brain Barrier Disruption Induced by Chronic Sleep Loss: Low-Grade Inflammation May Be the Link. J. Immunol. Res. 2016, 2016, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Ali, T.; Rehman, S.U.; Shah, F.A.; Kim, M.O. Acute Dose of Melatonin via Nrf2 Dependently Prevents Acute Ethanol-Induced Neurotoxicity in the Developing Rodent Brain. J. Neuroinflamm. 2018, 15, 119. [Google Scholar] [CrossRef]
- Tyagi, E.; Agrawal, R.; Nath, C.; Shukla, R. Effect of Melatonin on Neuroinflammation and Acetylcholinesterase Activity Induced by LPS in Rat Brain. Eur. J. Pharmacol. 2010, 640, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Guzman-Martinez, L.; Maccioni, R.B.; Andrade, V.; Navarrete, L.P.; Pastor, M.G.; Ramos-Escobar, N. Neuroinflammation as a Common Feature of Neurodegenerative Disorders. Front. Pharmacol. 2019, 10, 1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.H.; Karri, V.; Tay, N.W.R.; Chang, K.H.; Ah, H.Y.; Ng, P.Q.; Ho, H.S.; Keh, H.W.; Candasamy, M. Emerging Pathways to Neurodegeneration: Dissecting the Critical Molecular Mechanisms in Alzheimer’s Disease, Parkinson’s Disease. Biomed. Pharmacother. 2019, 111, 765–777. [Google Scholar] [CrossRef] [PubMed]
- Rudnitskaya, E.A.; Maksimova, K.Y.; Muraleva, N.A.; Logvinov, S.V.; Yanshole, L.V.; Kolosova, N.G.; Stefanova, N.A. Beneficial Effects of Melatonin in a Rat Model of Sporadic Alzheimer’s Disease. Biogerontology 2015, 16, 303–316. [Google Scholar] [CrossRef]
- Cardinali, D.P. Melatonin: Clinical Perspectives in Neurodegeneration. Front. Endocrinol. 2019, 10, 480. [Google Scholar] [CrossRef]
- Alghamdi, B.S. The Neuroprotective Role of Melatonin in Neurological Disorders. J. Neurosci. Res. 2018, 96, 1136–1149. [Google Scholar] [CrossRef]
- Sayre, L.; Smith, M.; Perry, G. Chemistry and Biochemistry of Oxidative Stress in Neurodegenerative Disease. Curr. Med. Chem. 2012, 8, 721–738. [Google Scholar] [CrossRef]
- Maurizi, C.P. Alzheimer’s Disease: Roles for Mitochondrial Damage, the Hydroxyl Radical, and Cerebrospinal Fluid Deficiency of Melatonin. Med. Hypotheses 2001, 57, 156–160. [Google Scholar] [CrossRef]
- Wongprayoon, P.; Govitrapong, P. Melatonin Attenuates Methamphetamine-Induced Neurotoxicity. Curr. Pharm. Des. 2016, 22, 1022–1032. [Google Scholar] [CrossRef]
- Şirin, F.B.; Kumbul Doğuc, D.; Vural, H.; Eren, İ.; İnanli, İ.; Sütçü, R.; Delibaş, N. Plasma 8-IsoPGF2α and Serum Melatonin Levels in Patients with Minimal Cognitive Impairment and Alzheimer Disease. Turk. J. Med. Sci. 2015, 45, 1073–1077. [Google Scholar] [CrossRef]
- Menegardo, C.S.; Friggi, F.A.; Scardini, J.B.; Rossi, T.S.; Vieira, T.D.S.; Tieppo, A.; Morelato, R.L. Sundown Syndrome in Patients with Alzheimer’s Disease Dementia. Dement. Neuropsychol. 2019, 13, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Paul, R.; Phukan, B.C.; Justin Thenmozhi, A.; Manivasagam, T.; Bhattacharya, P.; Borah, A. Melatonin Protects against Behavioral Deficits, Dopamine Loss and Oxidative Stress in Homocysteine Model of Parkinson’s Disease. Life Sci. 2018, 192, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Dowling, G.A.; Mastick, J.; Colling, E.; Carter, J.H.; Singer, C.M.; Aminoff, M.J. Melatonin for Sleep Disturbances in Parkinson’s Disease. Sleep Med. 2005, 6, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.F.; Huang, H.J.; Lee, H.C.; Hung, K.C.; Wu, R.T.; Lin, A.M.Y. Melatonin Attenuates Kainic Acid-Induced Neurotoxicity in Mouse Hippocampus via Inhibition of Autophagy and α-Synuclein Aggregation. J. Pineal Res. 2012, 52, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.M.Y.; Fang, S.F.; Chao, P.L.; Yang, C.H. Melatonin Attenuates Arsenite-Induced Apoptosis in Rat Brain: Involvement of Mitochondrial and Endoplasmic Reticulum Pathways and Aggregation of α-Synuclein. J. Pineal Res. 2007, 43, 163–171. [Google Scholar] [CrossRef]
- Brito-Armas, J.M.; Baekelandt, V.; Castro-Hernández, J.R.; González-Hernández, T.; Rodríguez, M.; Fuentes, R.C. Melatonin Prevents Dopaminergic Cell Loss Induced by Lentiviral Vectors Expressing A30P Mutant Alpha-Synuclein. Histol. Histopathol. 2013, 28, 999–1006. [Google Scholar] [CrossRef]
- Hu, W.; Ma, Z.; Jiang, S.; Fan, C.; Deng, C.; Yan, X.; Di, S.; Lv, J.; Reiter, R.J.; Yang, Y. Melatonin: The Dawning of a Treatment for Fibrosis? J. Pineal Res. 2016, 60, 121–131. [Google Scholar] [CrossRef]
- Zhao, X.; Sun, J.; Su, W.; Shan, H.; Zhang, B.; Wang, Y.; Shabanova, A.; Shan, H.; Liang, H. Melatonin Protects against Lung Fibrosis by Regulating the Hippo/YAP Pathway. Int. J. Mol. Sci. 2018, 19, 1118. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, Z.; Kotuk, M.; Erdogan, H.; Iraz, M.; Yagmurca, M.; Kuku, I.; Fadillioglu, E. Preventive Effect of Melatonin on Bleomycin-Induced Lung Fibrosis in Rats. J. Pineal Res. 2006, 40, 27–33. [Google Scholar] [CrossRef]
- Wang, R.; Zhou, S.; Wu, P.; Li, M.; Ding, X.; Sun, L.; Xu, X.; Zhou, X.; Zhou, L.; Cao, C.; et al. Identifying Involvement of H19-MiR-675-3p-IGF1R and H19-MiR-200a-PDCD4 in Treating Pulmonary Hypertension with Melatonin. Mol. Ther. Nucleic Acids 2018, 13, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Candia, A.; Veliz, M.; Carrasco-Pozo, C.; Castillo, R.L.; Cárdenas, J.C.; Ebensperger, G.; Reyes, R.V.; Llanos, A.J.; Herrera, E.A. Antenatal Melatonin Modulates an Enhanced Antioxidant/pro-Oxidant Ratio in Pulmonary Hypertensive Newborn Sheep. Redox Biol. 2019, 22, 101128. [Google Scholar] [CrossRef] [PubMed]
- Hung, M.W.; Yeung, H.M.; Lau, C.F.; Poon, A.M.S.; Tipoe, G.L.; Fung, M.L. Melatonin Attenuates Pulmonary Hypertension in Chronically Hypoxic Rats. Int. J. Mol. Sci. 2017, 18, 1125. [Google Scholar] [CrossRef] [PubMed]
- Maarman, G.; Blackhurst, D.; Thienemann, F.; Blauwet, L.; Butrous, G.; Davies, N.; Sliwa, K.; Lecour, S. Melatonin as a Preventive and Curative Therapy against Pulmonary Hypertension. J. Pineal Res. 2015, 59, 343–353. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Zhang, W.; Qiao, J.; Peng, Z.; Chai, X. Melatonin Protects against COPD by Attenuating Apoptosis and Endoplasmic Reticulum Stress via Upregulating SIRT1 Expression in Rats. Can. J. Physiol. Pharmacol. 2019, 97, 386–392. [Google Scholar] [CrossRef]
- De Matos Cavalcante, A.G.; De Bruin, P.F.C.; De Bruin, V.M.S.; Nunes, D.M.; Pereira, E.D.B.; Cavalcante, M.M.; Andrade, G.M. Melatonin Reduces Lung Oxidative Stress in Patients with Chronic Obstructive Pulmonary Disease: A Randomized, Double-Blind, Placebo-Controlled Study. J. Pineal Res. 2012, 53, 238–244. [Google Scholar] [CrossRef]
- Gattinoni, L.; Chiumello, D.; Caironi, P.; Busana, M.; Romitti, F.; Brazzi, L.; Camporota, L. COVID-19 Pneumonia: Different Respiratory Treatments for Different Phenotypes? Intensive Care Med. 2020, 46, 1099–1102. [Google Scholar] [CrossRef]
- Wichmann, D.; Sperhake, J.-P.; Lütgehetmann, M.; Steurer, S.; Edler, C.; Heinemann, A.; Heinrich, F.; Mushumba, H.; Kniep, I.; Schröder, A.S.; et al. Autopsy Findings and Venous Thromboembolism in Patients With COVID-19. Ann. Intern. Med. 2020, 173, 268–277. [Google Scholar] [CrossRef]
- Ye, Q.; Wang, B.; Mao, J. The Pathogenesis and Treatment of the ‘Cytokine Storm’ in COVID-19. J. Infect. 2020, 80, 607–613. [Google Scholar] [CrossRef]
- Shneider, A.; Kudriavtsev, A.; Vakhrusheva, A. Can Melatonin Reduce the Severity of COVID-19 Pandemic? Int. Rev. Immunol. 2020, 39, 153–162. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, X.; Ni, L.; Di, X.; Ma, B.; Niu, S.; Liu, C.; Reiter, R.J. COVID-19: Melatonin as a Potential Adjuvant Treatment. Life Sci. 2020, 250, 117583. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.; Grailer, J.J.; Wang, N.; Wang, M.; Yao, J.; Zhong, R.; Gao, G.F.; Ward, P.A.; Tan, D.-X.; et al. Melatonin Alleviates Acute Lung Injury through Inhibiting the NLRP3 Inflammasome. J. Pineal Res. 2016, 60, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, P.H.; Bärtschi, C.; Spillmann, M.; Ehlert, U.; Von Känel, R. Effect of Oral Melatonin on the Procoagulant Response to Acute Psychosocial Stress in Healthy Men: A Randomized Placebo-Controlled Study. J. Pineal Res. 2008, 44, 358–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bekyarova, G.; Tancheva, S.; Hristova, M. The Effects of Melatonin on Burn-Induced Inflammatory Responses and Coagulation Disorders in Rats. Methods Find. Exp. Clin. Pharmacol. 2010, 32, 299–303. [Google Scholar] [CrossRef]
- Baumgartner, L.; Lam, K.; Lai, J.; Barnett, M.; Thompson, A.; Gross, K.; Morris, A. Effectiveness of Melatonin for the Prevention of Intensive Care Unit Delirium. Pharmacotherapy 2019, 39, 280–287. [Google Scholar] [CrossRef]
- Martínez-González, M.A.; Gea, A.; Ruiz-Canela, M. The Mediterranean Diet and Cardiovascular Health: A Critical Review. Circ. Res. 2019, 124, 779–798. [Google Scholar] [CrossRef]
- Guerrini, S.; Mangani, S.; Romboli, Y.; Luti, S.; Pazzagli, L.; Granchi, L. Impact of Saccharomyces Cerevisiae Strains on Health-Promoting Compounds in Wine. Fermentation 2018, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Papada, E.; Forbes, A.; Amerikanou, C.; Torović, L.; Kalogeropoulos, N.; Tzavara, C.; Triantafillidis, J.; Kaliora, A. Antioxidative Efficacy of a Pistacia Lentiscus Supplement and Its Effect on the Plasma Amino Acid Profile in Inflammatory Bowel Disease: A Randomised, Double-Blind, Placebo-Controlled Trial. Nutrients 2018, 10, 1779. [Google Scholar] [CrossRef] [Green Version]
- Treatment Protocol—Frontline COVID-19 Critical Care Alliance. Available online: https://covid19criticalcare.com/treatment-protocol/ (accessed on 24 July 2020).




{kind=link}
{kind=link}
{kind=link}
| Mechanism | Effect | Study Type | |||
|---|---|---|---|---|---|
| Maternal & Fetal Health | Improves mitochondrial function, reduces mitochondrial oxidative stress [48] | Promotes embryo development | In vivo | ||
| TET genes function [36] | Regulation of genomic DNA methylation levels | In vitro | |||
| Counteracts the effects of Aanat knockdown [33] | Protects fertilization and tissue differentiation | In vitro | |||
| Direct scavenger of hydroxyl groups and nitrogen reactive species [49] | Detoxification of superoxide anions | In vitro | |||
| Mitochondrial complexes I and IV [50] | Protects ATP synthesis | In vitro | |||
| Prevents pathological opening of mPTP [51] | Protects mitochondrial DNA | In vitro | |||
| Inhibits anti-inflammatory molecules, like 8-isoprostanes [52] | Reduces hypoxic-ischemic inflammation | In vivo | |||
| Inhibits Cytochrome C release [53] | Anti-apoptotic | In vitro | |||
| Increases Bax and Bad pro-apoptosis molecules production [54] | Anti-apoptotic Prevention of DNA disruption | In vitro | |||
| Enhances glutathione peroxidase levels [55] | Antioxidant | In vitro | |||
| Not known [56] | Increases offspring survival in a model of lipopolysaccharide-induced inflammation | In vivo | |||
| Not known [57] | Prevents intrauterine growth retardation associated with continuous light exposure during pregnancy | In vivo | |||
| Increases plasma NO and SOD levels [58] | Beneficial in hypoxic-ischemic encephalopathy of the newborn | In vivo | |||
| Cardiovascular | Immediate | Intermediate | Final | ||
| Inhibits MPTP opening [59,60] | Decrease apoptosis | Decrease cell death | Prevent/reduce myocardial IRI | In vitro | |
| Inhibit Cyt C release [61] | Decrease apoptosis | Decrease cell death | In vitro | ||
| Activates JAK/STAT3 [62] | Decrease Bax, Increase Bcl | Decrease apoptosis | In vitro | ||
| Improves TAC [60,61] | Scavenger activity Increase endogenous antioxidant capacity Stop cardiolipin peroxidation | Decrease oxidative damage | In vitro | ||
| Improves calcium handling [63] | Ca2+-calmodulin modulation | Decrease cell death, reduce apoptosis | In vivo | ||
| Nrf 2 [64,65] | Transactivate HO-1 | Decrease inflammation and oxidation | In vivo | ||
| iNOS inhibition [66,67] | Lower NO levels | Decrease oxidative stress | In vivo In vitro | ||
| Inhibits inflammatory cytokine release [67] | Decrease of TNF-α, IL-1β, IL-6 | Decrease inflammation | In vitro | ||
| Stimulates anti-inflammatory cytokines [68] | Increase of IL-10 | Decrease inflammation | In vivo | ||
| Neuroinflammation | Maintains the levels of parvalbumin and hippocalcin [37] | Prevents neuronal death in cerebral ischemia | In vivo In vitro | ||
| Decreases NO, peroxynitrite formation [38] | Reduces hyperactivity linked to neurodegeneration induced by cerebral ischemia and reperfusion; Reduces PARS and brain edema | In vivo In vitro | |||
| Activation of SIRT1 signaling [44] | Reduced infarct volume, lowered brain edema, increased neurological scores in IRI | In vivo In vitro | |||
| Prevents accumulation of free radicals [47] | Counteracts the destructive effects of NMDA or hypoxia/reperfusion | In vitro | |||
| Decreases pro-inflammatory cytokines IL-1β, IL-6, and TNF-α in PFC [69] | Attenuates neuroinflammation in the aged mouse brain | In vivo In vitro | |||
| Respiratory | Suppresses the NLRP3 inflammasome [70] | Protectects against radiation-induced lung injury | In vivo In vitro | ||
| Decreasies lung edema and reduces NF-κB activation, enhances the secretion of IL-10 [71] | Averts volume induced lung injury | In vivo In vitro | |||
| Sirtuin-1 upregulation [72] | Diminishes TGF-β1 activity, apoptosis, and endoplasmic reticulum stress | In vivo In vitro | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chitimus, D.M.; Popescu, M.R.; Voiculescu, S.E.; Panaitescu, A.M.; Pavel, B.; Zagrean, L.; Zagrean, A.-M. Melatonin’s Impact on Antioxidative and Anti-Inflammatory Reprogramming in Homeostasis and Disease. Biomolecules 2020, 10, 1211. https://doi.org/10.3390/biom10091211
Chitimus DM, Popescu MR, Voiculescu SE, Panaitescu AM, Pavel B, Zagrean L, Zagrean A-M. Melatonin’s Impact on Antioxidative and Anti-Inflammatory Reprogramming in Homeostasis and Disease. Biomolecules. 2020; 10(9):1211. https://doi.org/10.3390/biom10091211
Chicago/Turabian StyleChitimus, Diana Maria, Mihaela Roxana Popescu, Suzana Elena Voiculescu, Anca Maria Panaitescu, Bogdan Pavel, Leon Zagrean, and Ana-Maria Zagrean. 2020. "Melatonin’s Impact on Antioxidative and Anti-Inflammatory Reprogramming in Homeostasis and Disease" Biomolecules 10, no. 9: 1211. https://doi.org/10.3390/biom10091211