
Different Diet Energy Levels Alter Body Condition, Glucolipid Metabolism, Fecal Microbiota and Metabolites in Adult Beagle Dogs
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Serum Biochemistry Analysis
2.3. Fecal pH Analysis
2.4. Content Analysis of SCFAs
2.5. Content Analysis of BAs
2.6. Microbial Analysis
2.6.1. DNA Extraction, Amplification and Sequencing
2.6.2. Bioinformatics Analysis
2.7. Statistical Analysis
3. Results
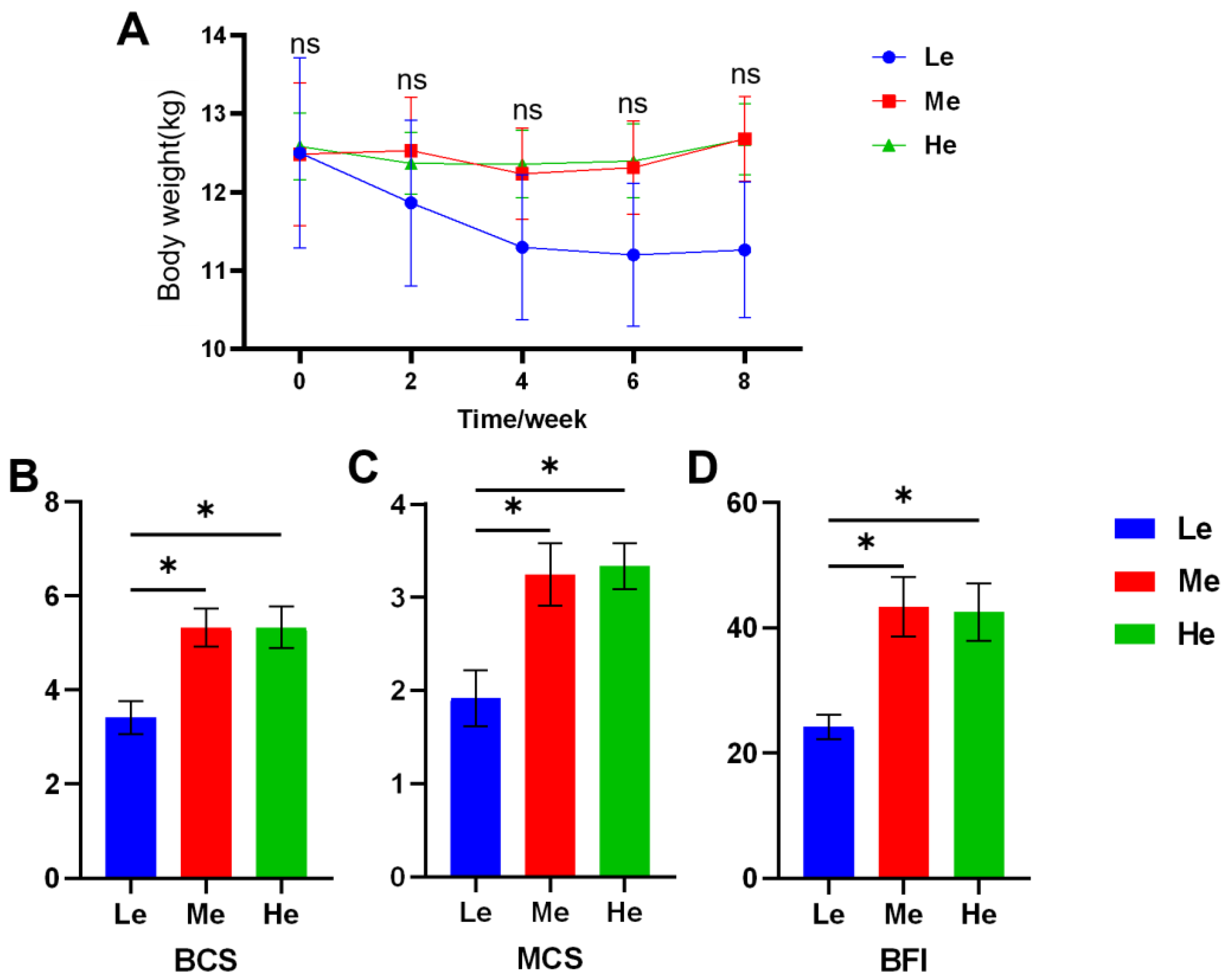
3.1. Changes in Body Weight and Body Condition of Beagles with Different Diet Energy Levels
3.2. Changes in Blood Glucose and Blood Lipid Levels of Beagles with Different Diet Energy Levels
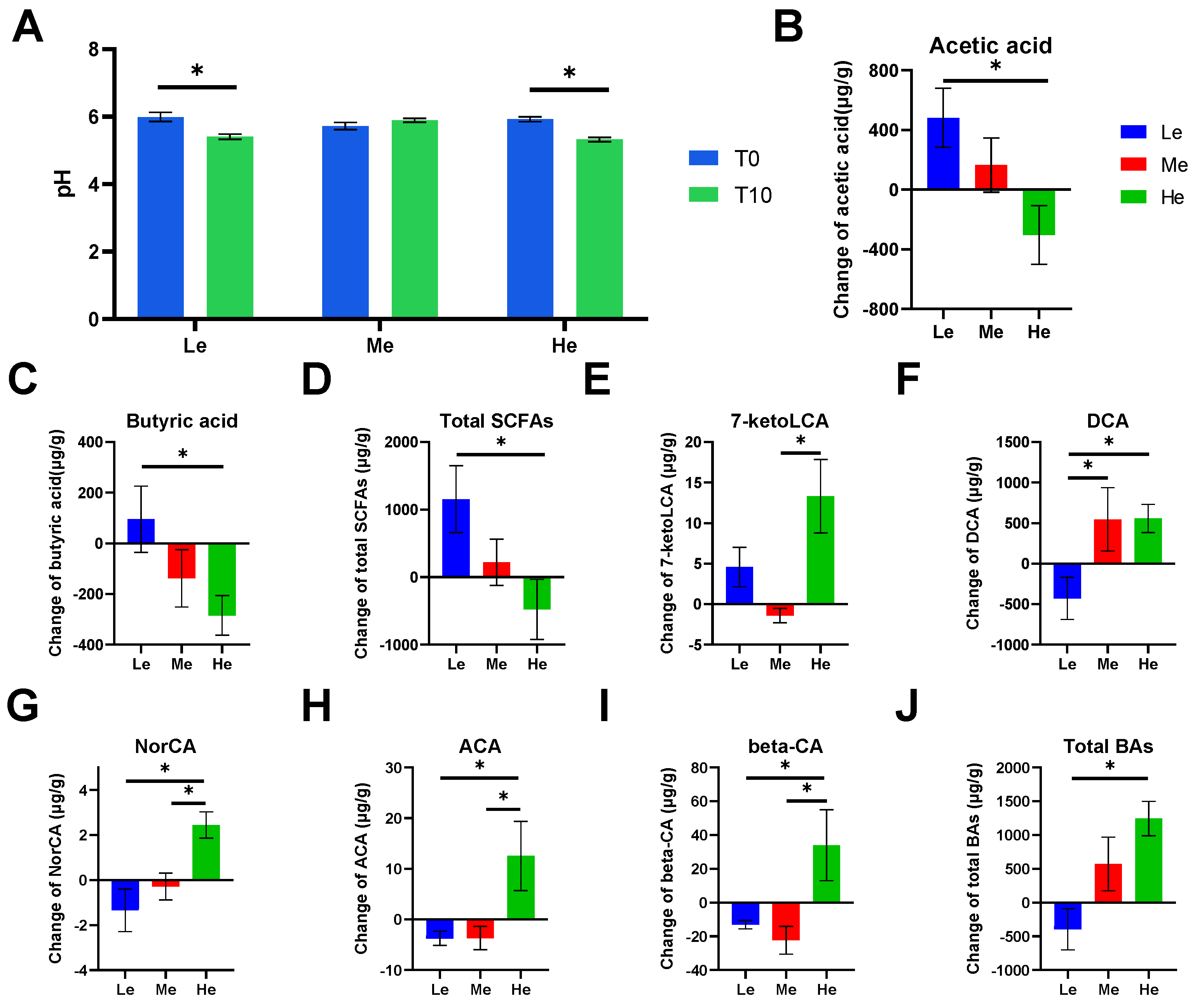
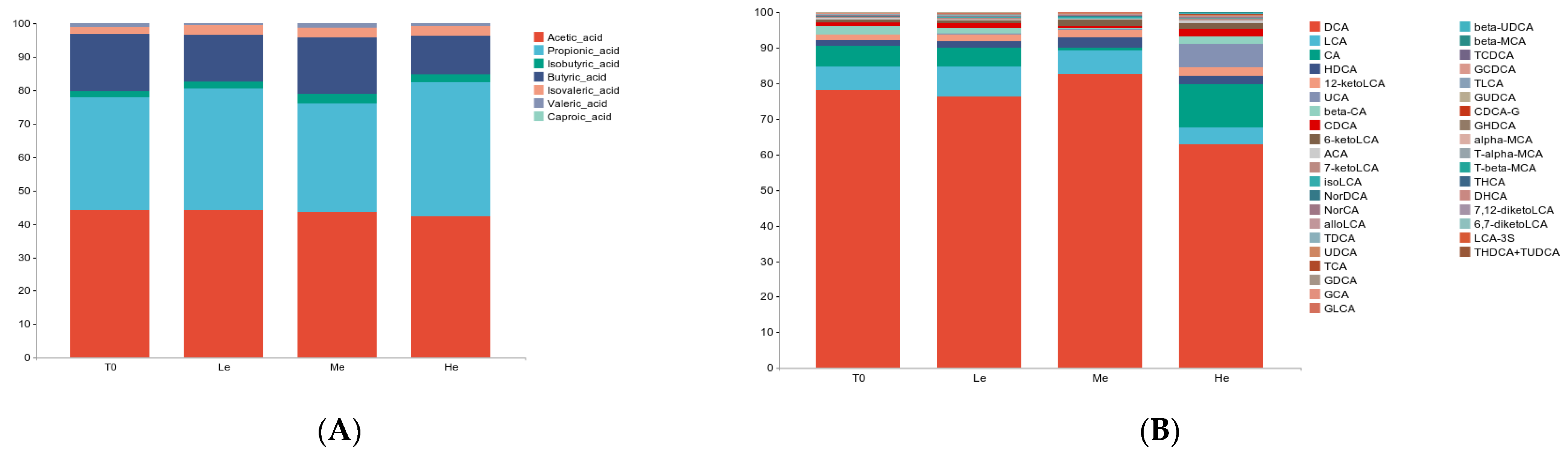
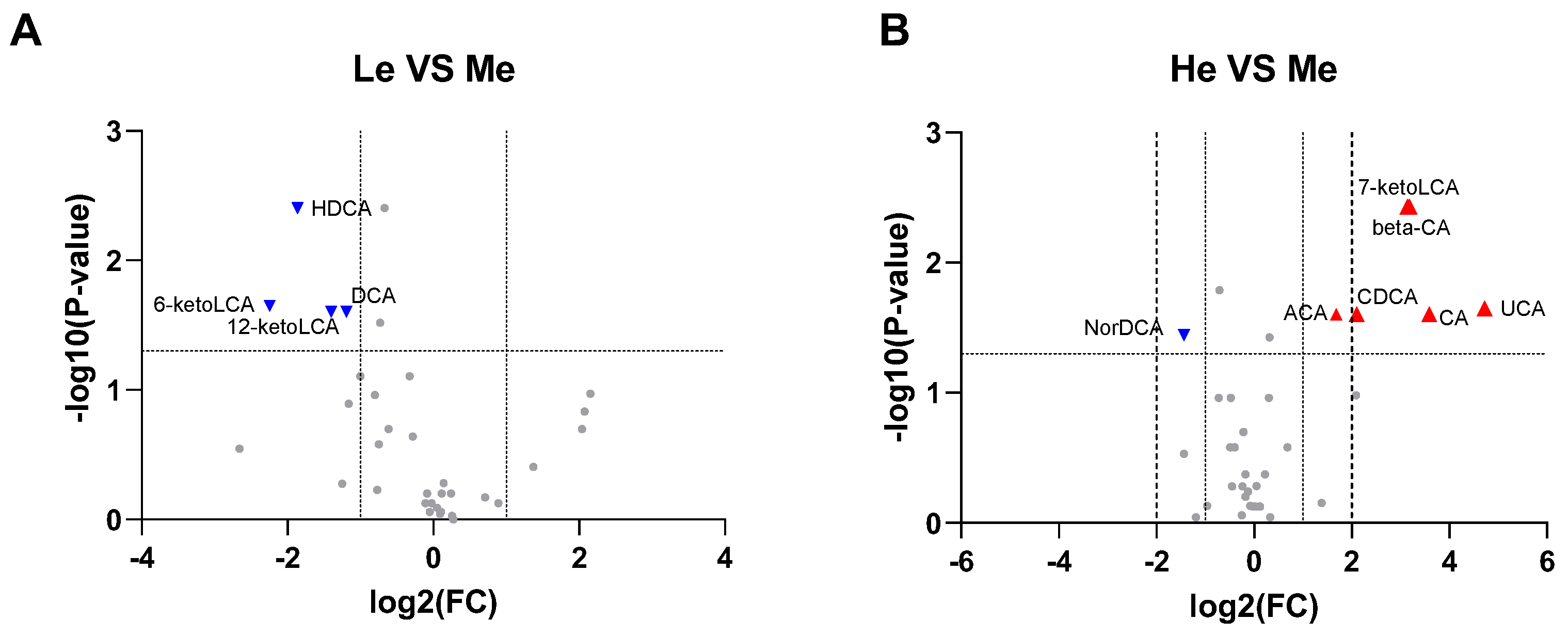
3.3. Changes in Fecal pH, SCFAs and BA Levels of Beagles with Different Diet Energy Levels
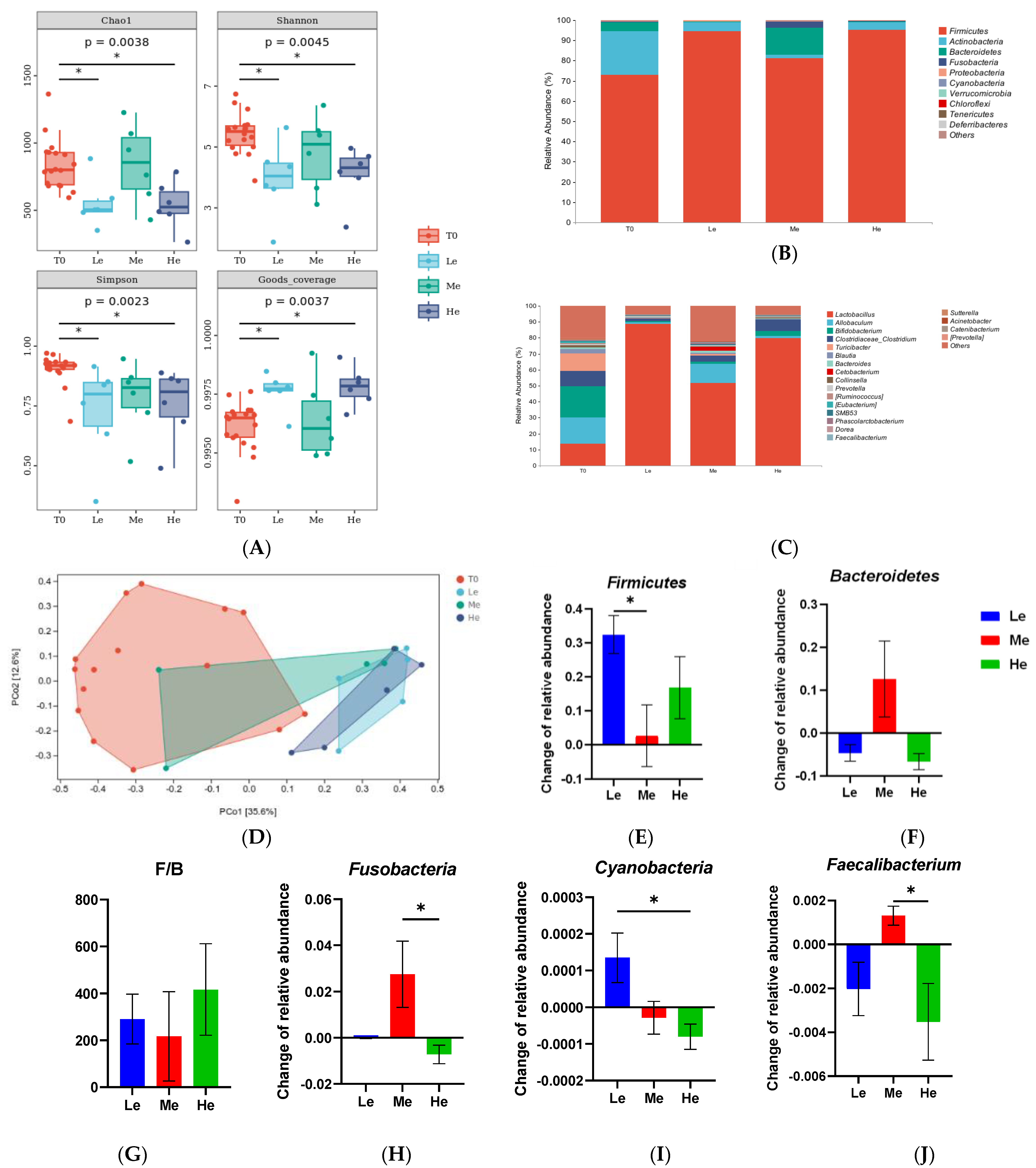
3.4. Changes in the Structure and Composition of Fecal Microbiota of Beagles with Different Diet Energy Levels
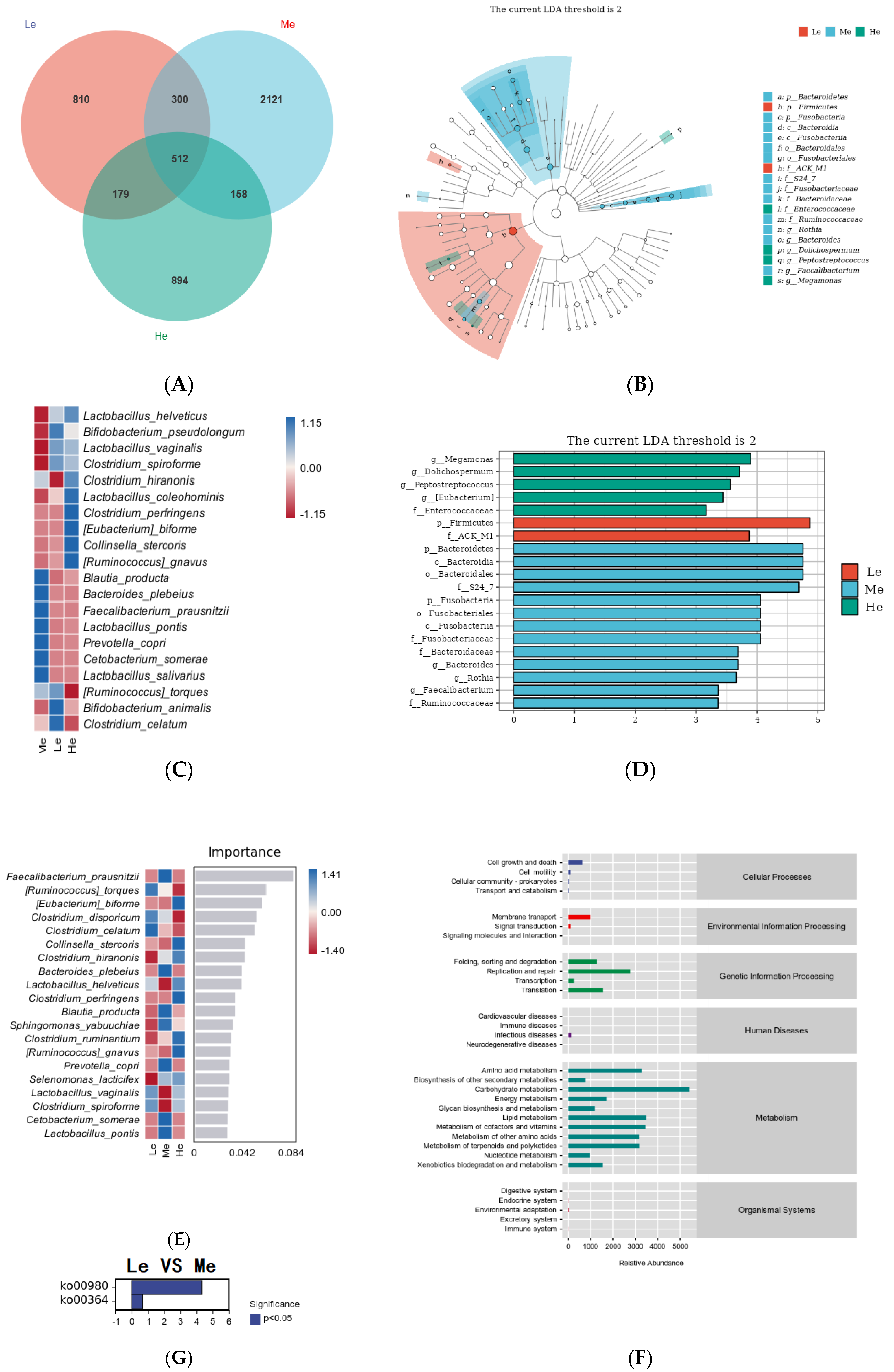
3.5. Effects of Diet Energy Level on Fecal Microbiota and Metabolic Pathways
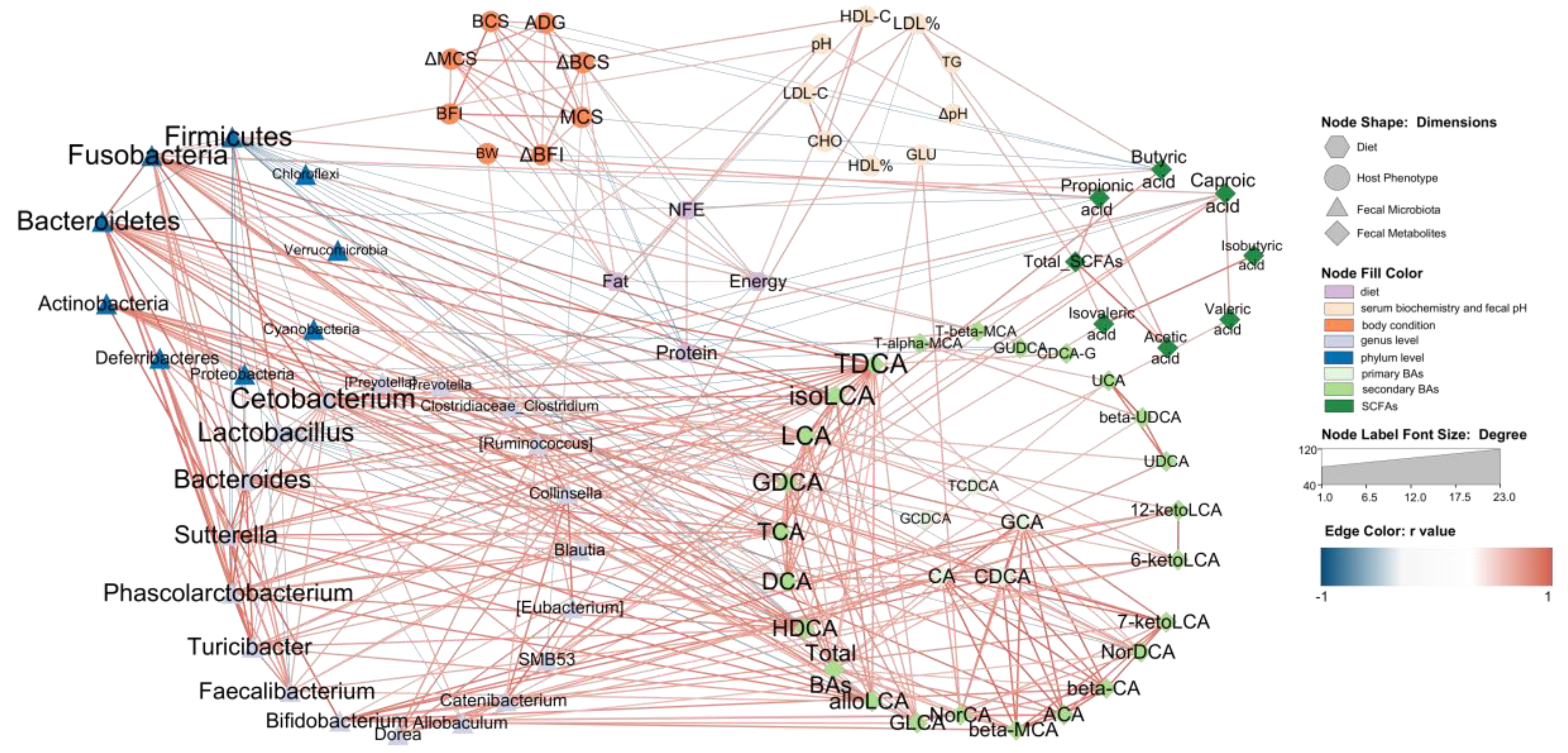
3.6. Network Relation between Diet, Host Microbiota and Fecal Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marchi, P.H.; Vendramini, T.H.A.; Perini, M.P.; Zafalon, R.V.A.; Amaral, A.R.; Ochamotto, V.A.; Da Silveira, J.C.; Dagli, M.L.Z.; Brunetto, M.A. Obesity, Inflammation, and Cancer in Dogs: Review and Perspectives. Front. Vet. Sci. 2022, 9, 1004122. [Google Scholar] [CrossRef] [PubMed]
- Partington, C.; Hodgkiss-Geere, H.; Woods, G.R.T.; Dukes-McEwan, J.; Flanagan, J.; Biourge, V.; German, A.J. The Effect of Obesity and Subsequent Weight Reduction on Cardiac Structure and Function in Dogs. BMC Vet. Res. 2022, 18, 351. [Google Scholar] [CrossRef]
- Ahuja, R.P.; Fletcher, J.M.; Granger, L.A.; Liu, C.-C.; Miessler, B.; Mitchell, M.A. Changes in Glucose Tolerance and Insulin Secretion in a Cohort of Cats with Chronic Obesity. Can. J. Vet. Res. 2022, 86, 181–187. [Google Scholar] [PubMed]
- Linder, D.E.; Freeman, L.M. Evaluation of Calorie Density and Feeding Directions for Commercially Available Diets Designed for Weight Loss in Dogs and Cats. J. Am. Vet. Med. Assoc. 2010, 236, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Olivindo, R.F.G.; Zafalon, R.V.A.; Teixeira, F.A.; Vendramini, T.H.A.; Pedrinelli, V.; Brunetto, M.A. Evaluation of the Nutrients Supplied by Veterinary Diets Commercialized in Brazil for Obese Dogs Undergoing a Weight Loss Program. J. Anim. Physiol. Anim. Nutr. 2022, 106, 355–367. [Google Scholar] [CrossRef]
- Farcas, A.K.; Larsen, J.A.; Owens, T.J.; Nelson, R.W.; Kass, P.H.; Fascetti, A.J. Evaluation of Total Dietary Fiber Concentration and Composition of Commercial Diets Used for Management of Diabetes Mellitus, Obesity, and Dietary Fat-Responsive Disease in Dogs. J. Am. Vet. Med. Assoc. 2015, 247, 501–507. [Google Scholar] [CrossRef]
- Bland, I.M.; Guthrie-Jones, A.; Taylor, R.D.; Hill, J. Dog Obesity: Veterinary Practices’ and Owners’ Opinions on Cause and Management. Prev. Vet. Med. 2010, 94, 310–315. [Google Scholar] [CrossRef]
- Shepherd, M. Canine and Feline Obesity Management. Vet. Clin. N. Am. Small Anim. Pract. 2021, 51, 653–667. [Google Scholar] [CrossRef]
- Grant, C.E.; Shoveller, A.K.; Blois, S.; Bakovic, M.; Monteith, G.; Verbrugghe, A. Dietary Intake of Amino Acids and Vitamins Compared to NRC Requirements in Obese Cats Undergoing Energy Restriction for Weight Loss. BMC Vet. Res. 2020, 16, 426. [Google Scholar] [CrossRef]
- Asaro, N.J.; Seymour, D.J.; Mansilla, W.D.; Cant, J.P.; Zijlstra, R.T.; Berendt, K.D.; Brewer, J.; Shoveller, A.K. Modelling Net Energy of Commercial Cat Diets. PLoS ONE 2019, 14, e0218173. [Google Scholar] [CrossRef] [Green Version]
- Keller, E.; Sagols, E.; Flanagan, J.; Biourge, V.; German, A.J. Use of Reduced-Energy Content Maintenance Diets for Modest Weight Reduction in Overweight Cats and Dogs. Res. Vet. Sci. 2020, 131, 194–205. [Google Scholar] [CrossRef]
- Serisier, S.; Weber, M.; Feugier, A.; Fardet, M.-O.; Garnier, F.; Biourge, V.; German, A.J. Maintenance Energy Requirements in Miniature Colony Dogs. J. Anim. Physiol. Anim. Nutr. 2013, 97 (Suppl. S1), 60–67. [Google Scholar] [CrossRef]
- Piantedosi, D.; Palatucci, A.T.; Giovazzino, A.; Ruggiero, G.; Rubino, V.; Musco, N.; Carriero, F.; Farina, F.; Attia, Y.A.E.W.; Terrazzano, G.; et al. Effect of a Weight Loss Program on Biochemical and Immunological Profile, Serum Leptin Levels, and Cardiovascular Parameters in Obese Dogs. Front. Vet. Sci. 2020, 7, 398. [Google Scholar] [CrossRef]
- Xue, J.; Lu, Y.; Zou, T.; Shi, W.; Wang, S.; Cheng, X.; Wan, J.; Chen, Y.; Wang, M.; Wang, Q.; et al. A Protein- and Fiber-Rich Diet with Astaxanthin Alleviates High-Fat Diet-Induced Obesity in Beagles. Front. Nutr. 2022, 9, 1019615. [Google Scholar] [CrossRef]
- Phungviwatnikul, T.; Lee, A.H.; Belchik, S.E.; Suchodolski, J.S.; Swanson, K.S. Weight Loss and High-Protein, High-Fiber Diet Consumption Impact Blood Metabolite Profiles, Body Composition, Voluntary Physical Activity, Fecal Microbiota, and Fecal Metabolites of Adult Dogs. J. Anim. Sci. 2022, 100, skab379. [Google Scholar] [CrossRef]
- Weber, M.; Bissot, T.; Servet, E.; Sergheraert, R.; Biourge, V.; German, A.J. A High-Protein, High-Fiber Diet Designed for Weight Loss Improves Satiety in Dogs. J. Vet. Intern. Med. 2007, 21, 1203–1208. [Google Scholar] [CrossRef]
- Bermudez Sanchez, S.; Pilla, R.; Sarawichitr, B.; Gramenzi, A.; Marsilio, F.; Steiner, J.M.; Lidbury, J.A.; Woods, G.R.T.; Suchodolski, J.S.; German, A.J. Untargeted Fecal Metabolome Analysis in Obese Dogs after Weight Loss Achieved by Feeding a High-Fiber-High-Protein Diet. Metabolomics 2021, 17, 66. [Google Scholar] [CrossRef]
- André, A.; Leriche, I.; Chaix, G.; Thorin, C.; Burger, M.; Nguyen, P. Recovery of Insulin Sensitivity and Optimal Body Composition after Rapid Weight Loss in Obese Dogs Fed a High-Protein Medium-Carbohydrate Diet. J. Anim. Physiol. Anim. Nutr. 2017, 101 (Suppl. S1), 21–30. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.; Bartges, J.W.; Zemel, M.B.; Kirk, C.A.; Witzel-Rollins, A. Effect of a Leucine/Pyridoxine Nutraceutical on Caloric Intake and Body Composition of Obese Dogs Losing Weight. Front. Vet. Sci. 2020, 7, 555. [Google Scholar] [CrossRef]
- Floerchinger, A.M.; Jackson, M.I.; Jewell, D.E.; MacLeay, J.M.; Hahn, K.A.; Paetau-Robinson, I. Effect of Feeding a Weight Loss Food beyond a Caloric Restriction Period on Body Composition and Resistance to Weight Gain in Cats. J. Am. Vet. Med. Assoc. 2015, 247, 365–374. [Google Scholar] [CrossRef]
- Sabchuk, T.T.; Scheraiber, M.; Zanatta, C.P.; Maiorka, A.; Félix, A.P.; Oliveira, S.G. Body Condition of Dogs Fed Diets Containing Soya Hulls. J. Nutr. Sci. 2014, 3, e42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKnight, L.L.; Eyre, R.; Gooding, M.A.; Davenport, G.M.; Shoveller, A.K. Dietary Mannoheptulose Increases Fasting Serum Glucagon like Peptide-1 and Post-Prandial Serum Ghrelin Concentrations in Adult Beagle Dogs. Animals 2015, 5, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Alexander, C.; Cross, T.-W.L.; Devendran, S.; Neumer, F.; Theis, S.; Ridlon, J.M.; Suchodolski, J.S.; de Godoy, M.R.C.; Swanson, K.S. Effects of Prebiotic Inulin-Type Fructans on Blood Metabolite and Hormone Concentrations and Faecal Microbiota and Metabolites in Overweight Dogs. Br. J. Nutr. 2018, 120, 711–720. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, C.S.; Vendramini, T.H.A.; Amaral, A.R.; Rentas, M.F.; Ernandes, M.C.; da Silva, F.L.; Oba, P.M.; de Oliveira Roberti Filho, F.; Brunetto, M.A. Metabolic Variables of Obese Dogs with Insulin Resistance Supplemented with Yeast Beta-Glucan. BMC Vet. Res. 2022, 18, 14. [Google Scholar] [CrossRef] [PubMed]
- Apper, E.; Privet, L.; Taminiau, B.; Le Bourgot, C.; Svilar, L.; Martin, J.-C.; Diez, M. Relationships between Gut Microbiota, Metabolome, Body Weight, and Glucose Homeostasis of Obese Dogs Fed with Diets Differing in Prebiotic and Protein Content. Microorganisms 2020, 8, 513. [Google Scholar] [CrossRef] [Green Version]
- Söder, J.; Höglund, K.; Dicksved, J.; Hagman, R.; Eriksson Röhnisch, H.; Moazzami, A.A.; Wernersson, S. Plasma Metabolomics Reveals Lower Carnitine Concentrations in Overweight Labrador Retriever Dogs. Acta Vet. Scand. 2019, 61, 10. [Google Scholar] [CrossRef] [Green Version]
- Kilburn, L.R.; Koester, L.R.; Schmitz-Esser, S.; Serão, N.V.L.; Rossoni Serão, M.C. High-Fat Diets Led to OTU-Level Shifts in Fecal Samples of Healthy Adult Dogs. Front. Microbiol. 2020, 11, 564160. [Google Scholar] [CrossRef]
- Kilburn, L.R.; Allenspach, K.; Jergens, A.E.; Bourgois-Mochel, A.; Mochel, J.P.; Serao, M.C.R. Apparent Total Tract Digestibility, Fecal Characteristics, and Blood Parameters of Healthy Adult Dogs Fed High-Fat Diets. J. Anim. Sci. 2020, 98, skaa043. [Google Scholar] [CrossRef]
- Moinard, A.; Payen, C.; Ouguerram, K.; André, A.; Hernandez, J.; Drut, A.; Biourge, V.C.; Suchodolski, J.S.; Flanagan, J.; Nguyen, P.; et al. Effects of High-Fat Diet at Two Energetic Levels on Fecal Microbiota, Colonic Barrier, and Metabolic Parameters in Dogs. Front. Vet. Sci. 2020, 7, 566282. [Google Scholar] [CrossRef]
- Lyu, Y.; Liu, D.; Nguyen, P.; Peters, I.; Heilmann, R.M.; Fievez, V.; Hemeryck, L.Y.; Hesta, M. Differences in Metabolic Profiles of Healthy Dogs Fed a High-Fat vs. a High-Starch Diet. Front. Vet. Sci. 2022, 9, 801863. [Google Scholar] [CrossRef]
- Schauf, S.; de la Fuente, G.; Newbold, C.J.; Salas-Mani, A.; Torre, C.; Abecia, L.; Castrillo, C. Effect of Dietary Fat to Starch Content on Fecal Microbiota Composition and Activity in Dogs. J. Anim. Sci. 2018, 96, 3684–3698. [Google Scholar] [CrossRef]
- Nutritional Research Council. Nutrient Requirements of Dogs and Cats; The National Academies Press: Washington, DC, USA, 2006. [Google Scholar]
- Laflamme, D.R.P.C. Development and Validation of a Body Condition Score System for Dogs. Canine Pract. 1997, 22, 10–15. [Google Scholar]
- Freeman, L.M.; Michel, K.E.; Zanghi, B.M.; Vester Boler, B.M.; Fages, J. Evaluation of the Use of Muscle Condition Score and Ultrasonographic Measurements for Assessment of Muscle Mass in Dogs. Am. J. Vet. Res. 2019, 80, 595–600. [Google Scholar] [CrossRef]
- Witzel, A.L.; Kirk, C.A.; Henry, G.A.; Toll, P.W.; Brejda, J.J.; Paetau-Robinson, I. Use of a Novel Morphometric Method and Body Fat Index System for Estimation of Body Composition in Overweight and Obese Dogs. J. Am. Vet. Med. Assoc. 2014, 244, 1279–1284. [Google Scholar] [CrossRef]
- Han, X.; Guo, J.; You, Y.; Yin, M.; Ren, C.; Zhan, J.; Huang, W. A Fast and Accurate Way to Determine Short Chain Fatty Acids in Mouse Feces Based on GC-MS. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2018, 1099, 73–82. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, H.; Zhu, M.-J. A Sensitive GC/MS Detection Method for Analyzing Microbial Metabolites Short Chain Fatty Acids in Fecal and Serum Samples. Talanta 2019, 196, 249–254. [Google Scholar] [CrossRef]
- Hsu, Y.-L.; Chen, C.-C.; Lin, Y.-T.; Wu, W.-K.; Chang, L.-C.; Lai, C.-H.; Wu, M.-S.; Kuo, C.-H. Evaluation and Optimization of Sample Handling Methods for Quantification of Short-Chain Fatty Acids in Human Fecal Samples by GC-MS. J. Proteome Res. 2019, 18, 1948–1957. [Google Scholar] [CrossRef]
- Bhargava, P.; Smith, M.D.; Mische, L.; Harrington, E.; Fitzgerald, K.C.; Martin, K.; Kim, S.; Reyes, A.A.; Gonzalez-Cardona, J.; Volsko, C.; et al. Bile Acid Metabolism Is Altered in Multiple Sclerosis and Supplementation Ameliorates Neuroinflammation. J. Clin. Investig. 2020, 130, 3467–3482. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.; An, Z.; Shi, C.; Li, P.; Liu, L. A Sensitive and Efficient Method for Simultaneous Profiling of Bile Acids and Fatty Acids by UPLC-MS/MS. J. Pharm. Biomed. Anal. 2020, 178, 112815. [Google Scholar] [CrossRef]
- Yang, T.; Shu, T.; Liu, G.; Mei, H.; Zhu, X.; Huang, X.; Zhang, L.; Jiang, Z. Quantitative Profiling of 19 Bile Acids in Rat Plasma, Liver, Bile and Different Intestinal Section Contents to Investigate Bile Acid Homeostasis and the Application of Temporal Variation of Endogenous Bile Acids. J. Steroid Biochem. Mol. Biol. 2017, 172, 69–78. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing Large Minimum Evolution Trees with Profiles Instead of a Distance Matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing Taxonomic Classification of Marker-Gene Amplicon Sequences with QIIME 2′s Q2-Feature-Classifier Plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a Unified Paradigm for Sequence-Based Identification of Fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Zaura, E.; Keijser, B.J.F.; Huse, S.M.; Crielaard, W. Defining the Healthy “Core Microbiome” of Oral Microbial Communities. BMC Microbiol. 2009, 9, 259. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Liaw, A.; Wiener, M. Classification and Regression by RandomForest. Forest 2001, 23, 18–22. [Google Scholar]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Douglas, G.; Maffei, V.; Zaneveld, J.; Yurgel, S.; Brown, J.; Taylor, C.; Huttenhower, C.; Langille, M. PICRUSt2: An Improved and Extensible Approach for Metagenome Inference. BioRxiv 2019, 672295. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, J.; Harvey, W.J.; Bruss, M.L. Clinical Biochemistry of Domestic Animals; Academic Press: Cambridge, MA, USA, 2008. [Google Scholar]
- Minamoto, Y.; Minamoto, T.; Isaiah, A.; Sattasathuchana, P.; Buono, A.; Rangachari, V.R.; McNeely, I.H.; Lidbury, J.; Steiner, J.M.; Suchodolski, J.S. Fecal Short-Chain Fatty Acid Concentrations and Dysbiosis in Dogs with Chronic Enteropathy. J. Vet. Intern. Med. 2019, 33, 1608–1618. [Google Scholar] [CrossRef] [Green Version]
- Vítek, L.; Haluzík, M. The Role of Bile Acids in Metabolic Regulation. J. Endocrinol. 2016, 228, R85–R96. [Google Scholar] [CrossRef]
- Thompson, J.; Johansen, R.; Dunbar, J.; Munsky, B. Machine Learning to Predict Microbial Community Functions: An Analysis of Dissolved Organic Carbon from Litter Decomposition. PLoS ONE 2019, 14, e0215502. [Google Scholar] [CrossRef] [Green Version]
- Jobgen, W.S.; Fried, S.K.; Fu, W.J.; Meininger, C.J.; Wu, G. Regulatory Role for the Arginine-Nitric Oxide Pathway in Metabolism of Energy Substrates. J. Nutr. Biochem. 2006, 17, 571–588. [Google Scholar] [CrossRef]
- Bolesławska, I.; Kowalówka, M.; Dobrzyńska, M.; Karaźniewicz-Łada, M.; Przysławski, J. Differences in the Concentration of Vitamin D Metabolites in Plasma Due to the Low-Carbohydrate-High-Fat Diet and the Eastern European Diet—A Pilot Study. Nutrients 2021, 13, 2774. [Google Scholar] [CrossRef]
- Bütikofer, U.; Burnand, D.; Portmann, R.; Blaser, C.; Schwander, F.; Kopf-Bolanz, K.A.; Laederach, K.; Badertscher, R.; Walther, B.; Vergères, G. Serum Metabolites Responding in a Dose-Dependent Manner to the Intake of a High-Fat Meal in Normal Weight Healthy Men Are Associated with Obesity. Metabolites 2021, 11, 392. [Google Scholar] [CrossRef]
- Yang, J.; Wei, H.; Zhou, Y.; Szeto, C.-H.; Li, C.; Lin, Y.; Coker, O.O.; Lau, H.C.H.; Chan, A.W.H.; Sung, J.J.Y.; et al. High-Fat Diet Promotes Colorectal Tumorigenesis Through Modulating Gut Microbiota and Metabolites. Gastroenterology 2022, 162, 135–149.e2. [Google Scholar] [CrossRef]
- Liu, Y.; Kong, X.; Jiang, G.; Tan, B.; Deng, J.; Yang, X.; Li, F.; Xiong, X.; Yin, Y. Effects of Dietary Protein/Energy Ratio on Growth Performance, Carcass Trait, Meat Quality, and Plasma Metabolites in Pigs of Different Genotypes. J. Anim. Sci. Biotechnol. 2015, 6, 36. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhou, J.; Degen, A.; Liu, H.; Cao, X.; Hao, L.; Shang, Z.; Ran, T.; Long, R. A Comparison of Average Daily Gain, Apparent Digestibilities, Energy Balance, Rumen Fermentation Parameters, and Serum Metabolites between Yaks (Bos grunniens) and Qaidam Cattle (Bos taurus) Consuming Diets Differing in Energy Level. Anim. Nutr. 2023, 12, 77–86. [Google Scholar] [CrossRef]
- Loftus, J.P.; Wakshlag, J.J. Canine and Feline Obesity: A Review of Pathophysiology, Epidemiology, and Clinical Management. Vet. Med. Res. Rep. 2015, 6, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Kleinert, M.; Clemmensen, C.; Hofmann, S.M.; Moore, M.C.; Renner, S.; Woods, S.C.; Huypens, P.; Beckers, J.; de Angelis, M.H.; Schürmann, A.; et al. Animal Models of Obesity and Diabetes Mellitus. Nat. Rev. Endocrinol. 2018, 14, 140–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Mazcorro, J.F.; Minamoto, Y.; Kawas, J.R.; Suchodolski, J.S.; de Vos, W.M. Akkermansia and Microbial Degradation of Mucus in Cats and Dogs: Implications to the Growing Worldwide Epidemic of Pet Obesity. Vet. Sci. 2020, 7, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.-S.; Seeley, R.J.; Sandoval, D.A. Signalling from the Periphery to the Brain That Regulates Energy Homeostasis. Nat. Rev. Neurosci. 2018, 19, 185–196. [Google Scholar] [CrossRef]
- Hall, J.A.; Vondran, J.C.; Vanchina, M.A.; Jewell, D.E. When Fed Foods with Similar Palatability, Healthy Adult Dogs and Cats Choose Different Macronutrient Compositions. J. Exp. Biol. 2018, 221, jeb.173450. [Google Scholar] [CrossRef] [Green Version]
- de Marchi, P.N.; Cardoso, M.J.L.; Fagnani, R.; Calesso, J.R.; Melussi, M.; Machado, L.H.d.A. Metabolic Parameters in Obese Dogs Undergoing to the Diet with Calorie Restriction. Open J. Vet. Med. 2018, 8, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Cline, M.G.; Burns, K.M.; Coe, J.B.; Downing, R.; Durzi, T.; Murphy, M.; Parker, V. 2021 AAHA Nutrition and Weight Management Guidelines for Dogs and Cats. J. Am. Anim. Hosp. Assoc. 2021, 57, 153–178. [Google Scholar] [CrossRef]
- Freeman, L.; Becvarova, I.; Cave, N.; MacKay, C.; Nguyen, P.; Rama, B.; Takashima, G.; Tiffin, R.; Tsjimoto, H.; WSAVA Nutritional Assessment Guidelines Task Force Members. WSAVA Nutritional Assessment Guidelines. J. Small Anim. Pract. 2011, 52, 385–396. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Rhoads, R.P. Effects of Heat Stress on Postabsorptive Metabolism and Energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.; Hosseindoust, A.; Ha, S.; Moturi, J.; Mun, J.; Tajudeen, H.; Kim, J. Metabolic Responses of Dietary Fiber during Heat Stress: Effects on Reproductive Performance and Stress Level of Gestating Sows. Metabolites 2022, 12, 280. [Google Scholar] [CrossRef]
- Kim, K.; Choi, Y.; Hosseindoust, A.; Kim, M.; Hwang, S.; Bu, M.; Lee, J.; Kim, Y.; Chae, B. Evaluation of High Nutrient Diets and Additional Dextrose on Reproductive Performance and Litter Performance of Heat-stressed Lactating Sows. Anim. Sci. J. 2019, 90, 1212–1219. [Google Scholar] [CrossRef]
- Khaki, Z.; Nooshirvani, P.; Shirani, D.; Masoudifard, M. Diagnostic Value of Atrial Natriuretic Peptide (ANP), B-Type Natriuretic Peptide (BNP) and Their Correlation with Lipoproteins in Dogs with Myxomatous Mitral Valve Disease. BMC Vet. Res. 2022, 18, 448. [Google Scholar] [CrossRef]
- Park, M.; Kim, K.H.; Jaiswal, V.; Choi, J.; Chun, J.L.; Seo, K.M.; Lee, M.-J.; Lee, H.-J. Effect of Black Ginseng and Silkworm Supplementation on Obesity, the Transcriptome, and the Gut Microbiome of Diet-Induced Overweight Dogs. Sci. Rep. 2021, 11, 16334. [Google Scholar] [CrossRef]
- do Nascimento, R.C.; Souza, C.M.M.; Bastos, T.S.; Kaelle, G.C.B.; de Oliveira, S.G.; Félix, A.P. Effects of an Herbal Source of Choline on Diet Digestibility and Palatability, Blood Lipid Profile, Liver Morphology, and Cardiac Function in Dogs. Animals 2022, 12, 2658. [Google Scholar] [CrossRef]
- Phungviwatnikul, T.; Alexander, C.; Do, S.; He, F.; Suchodolski, J.; Godoy, M.; Swanson, K. Effects of Dietary Macronutrient Profile on Apparent Total Tract Macronutrient Digestibility and Fecal Microbiota, Fermentative Metabolites, and Bile Acids of Female Dogs after Spay Surgery. J. Anim. Sci. 2021, 99, skab225. [Google Scholar] [CrossRef]
- Geary, E.L.; Oba, P.M.; Applegate, C.C.; Clark, L.V.; Fields, C.J.; Swanson, K.S. Effects of a Mildly Cooked Human-Grade Dog Diet on Gene Expression, Skin and Coat Health Measures, and Fecal Microbiota of Healthy Adult Dogs. J. Anim. Sci. 2022, 100, skac265. [Google Scholar] [CrossRef]
- Wang, L.; Ren, B.; Hui, Y.; Chu, C.; Zhao, Z.; Zhang, Y.; Zhao, B.; Shi, R.; Ren, J.; Dai, X.; et al. Methionine Restriction Regulates Cognitive Function in High-Fat Diet-Fed Mice: Roles of Diurnal Rhythms of SCFAs Producing- and Inflammation-Related Microbes. Mol. Nutr. Food Res. 2020, 64, e2000190. [Google Scholar] [CrossRef]
- Hou, Q.; Huang, J.; Zhao, L.; Pan, X.; Liao, C.; Jiang, Q.; Lei, J.; Guo, F.; Cui, J.; Guo, Y.; et al. Dietary Genistein Increases Microbiota-Derived Short Chain Fatty Acid Levels, Modulates Homeostasis of the Aging Gut, and Extends Healthspan and Lifespan. Pharmacol. Res. 2023, 188, 106676. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Guo, X.; Zhang, M.; Yang, L.; Liu, R.; Yin, J. Anthocyanins in Black Rice, Soybean and Purple Corn Increase Fecal Butyric Acid and Prevent Liver Inflammation in High Fat Diet-Induced Obese Mice. Food Funct. 2017, 8, 3178–3186. [Google Scholar] [CrossRef]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.K.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S.; et al. Effects of Targeted Delivery of Propionate to the Human Colon on Appetite Regulation, Body Weight Maintenance and Adiposity in Overweight Adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef] [Green Version]
- Byrne, C.S.; Chambers, E.S.; Alhabeeb, H.; Chhina, N.; Morrison, D.J.; Preston, T.; Tedford, C.; Fitzpatrick, J.; Irani, C.; Busza, A.; et al. Increased Colonic Propionate Reduces Anticipatory Reward Responses in the Human Striatum to High-Energy Foods. Am. J. Clin. Nutr. 2016, 104, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, Q.; Lian, Y.; Peng, P.; Yang, L.; Zhao, H.; Huang, P.; Ma, H.; Wei, H.; Yin, Y.; Liu, M. Association of Gut Microbiota and SCFAs with Finishing Weight of Diannan Small Ear Pigs. Front. Microbiol. 2023, 14, 1117965. [Google Scholar] [CrossRef] [PubMed]
- Manjarín, R.; Dillard, K.; Coffin, M.; Hernandez, G.V.; Smith, V.A.; Noland-Lidell, T.; Gehani, T.R.; Smart, H.J.; Wheeler, K.; Sprayberry, K.A.; et al. Dietary Fat Composition Shapes Bile Acid Metabolism and Severity of Liver Injury in a Pig Model of Pediatric NAFLD. Am. J. Physiol. Endocrinol. Metab. 2022, 323, E187–E206. [Google Scholar] [CrossRef] [PubMed]
- Breuninger, T.A.; Wawro, N.; Freuer, D.; Reitmeier, S.; Artati, A.; Grallert, H.; Adamski, J.; Meisinger, C.; Peters, A.; Haller, D.; et al. Fecal Bile Acids and Neutral Sterols Are Associated with Latent Microbial Subgroups in the Human Gut. Metabolites 2022, 12, 846. [Google Scholar] [CrossRef]
- Visekruna, A.; Luu, M. The Role of Short-Chain Fatty Acids and Bile Acids in Intestinal and Liver Function, Inflammation, and Carcinogenesis. Front. Cell Dev. Biol. 2021, 9, 703218. [Google Scholar] [CrossRef]
- Liu, L.; Xu, J.; Xu, X.; Mao, T.; Niu, W.; Wu, X.; Lu, L.; Zhou, H. Intestinal Stem Cells Damaged by Deoxycholic Acid via AHR Pathway Contributes to Mucosal Barrier Dysfunction in High-Fat Feeding Mice. Int. J. Mol. Sci. 2022, 23, 15578. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, J.; Wu, X.; Jing, H.; Zhang, S.; Hu, Z.; Rao, L.; Chang, Q.; Wang, L.; Zhang, Z. Gut Microbiota Alteration after Cholecystectomy Contributes to Post-Cholecystectomy Diarrhea via Bile Acids Stimulating Colonic Serotonin. Gut Microbes 2023, 15, 2168101. [Google Scholar] [CrossRef]
- He, Y.; Brouwers, B.; Liu, H.; Liu, H.; Lawler, K.; Mendes de Oliveira, E.; Lee, D.-K.; Yang, Y.; Cox, A.R.; Keogh, J.M.; et al. Human Loss-of-Function Variants in the Serotonin 2C Receptor Associated with Obesity and Maladaptive Behavior. Nat. Med. 2022, 28, 2537–2546. [Google Scholar] [CrossRef]
- Pan, J.; Chui, L.; Liu, T.; Zheng, Q.; Liu, X.; Liu, L.; Zhao, Y.; Zhang, L.; Song, M.; Han, J.; et al. Fecal Microbiota Was Reshaped in UCP1 Knock-In Pigs via the Adipose-Liver-Gut Axis and Contributed to Less Fat Deposition. Microbiol. Spectr. 2023, 11, e0354022. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, K.; Jian, S.; Xin, Z.; Wen, C.; Zhang, L.; Huang, J.; Deng, B.; Deng, J. Effects of Softening Dry Food with Water on Stress Response, Intestinal Microbiome, and Metabolic Profile in Beagle Dogs. Metabolites 2022, 12, 1124. [Google Scholar] [CrossRef]
- Xu, J.; Becker, A.A.M.J.; Luo, Y.; Zhang, W.; Ge, B.; Leng, C.; Wang, G.; Ding, L.; Wang, J.; Fu, X.; et al. The Fecal Microbiota of Dogs Switching to a Raw Diet Only Partially Converges to That of Wolves. Front. Microbiol. 2021, 12, 701439. [Google Scholar] [CrossRef]
- Yang, K.; Jian, S.; Guo, D.; Wen, C.; Xin, Z.; Zhang, L.; Kuang, T.; Wen, J.; Yin, Y.; Deng, B. Fecal Microbiota and Metabolomics Revealed the Effect of Long-Term Consumption of Gallic Acid on Canine Lipid Metabolism and Gut Health. Food Chem. X 2022, 15, 100377. [Google Scholar] [CrossRef]
- Butowski, C.F.; Moon, C.D.; Thomas, D.G.; Young, W.; Bermingham, E.N. The Effects of Raw-Meat Diets on the Gastrointestinal Microbiota of the Cat and Dog: A Review. N. Z. Vet. J. 2022, 70, 1–9. [Google Scholar] [CrossRef]
- Munukka, E.; Rintala, A.; Toivonen, R.; Nylund, M.; Yang, B.; Takanen, A.; Hänninen, A.; Vuopio, J.; Huovinen, P.; Jalkanen, S.; et al. Faecalibacterium Prausnitzii Treatment Improves Hepatic Health and Reduces Adipose Tissue Inflammation in High-Fat Fed Mice. ISME J. 2017, 11, 1667–1679. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Christophersen, C.T.; Sorich, M.J.; Gerber, J.P.; Angley, M.T.; Conlon, M.A. Increased Abundance of Sutterella Spp. and Ruminococcus Torques in Feces of Children with Autism Spectrum Disorder. Mol. Autism 2013, 4, 42. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Li, X.; Wang, X.; Zhang, C.; Zou, L.; Ren, H.; Liu, W. Characterization and Genomic Analysis of a Novel Lytic Phage DCp1 against Clostridium Perfringens Biofilms. Int. J. Mol. Sci. 2023, 24, 4191. [Google Scholar] [CrossRef]
- Paul, G.C.; Friend, D.G. Clostridial Enterotoxemia and Coccidiosis in Weanling Cottontail Rabbits (Sylvilagus audubonii, Sylvilagus floridanus, Sylvilagus nuttallii) from Colorado, USA. J. Wildl. Dis. 2019, 55, 189–195. [Google Scholar] [CrossRef]
- Pei, T.; Zhu, D.; Yang, S.; Hu, R.; Wang, F.; Zhang, J.; Yan, S.; Ju, L.; He, Z.; Han, Z.; et al. Bacteroides Plebeius Improves Muscle Wasting in Chronic Kidney Disease by Modulating the Gut-Renal Muscle Axis. J. Cell. Mol. Med. 2022, 26, 6066–6078. [Google Scholar] [CrossRef]
- Mao, B.; Guo, W.; Liu, X.; Cui, S.; Zhang, Q.; Zhao, J.; Tang, X.; Zhang, H. Potential Probiotic Properties of Blautia Producta Against Lipopolysaccharide-Induced Acute Liver Injury. Probiotics Antimicrob. Proteins 2023. [Google Scholar] [CrossRef]
- Yang, Y.-N.; Wang, Q.-C.; Xu, W.; Yu, J.; Zhang, H.; Wu, C. The Berberine-Enriched Gut Commensal Blautia Producta Ameliorates High-Fat Diet (HFD)-Induced Hyperlipidemia and Stimulates Liver LDLR Expression. Biomed. Pharmacother. 2022, 155, 113749. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Zhang, Z.; Ding, Q.; Yang, Y.; Bindelle, J.; Ran, C.; Zhou, Z. Intestinal Cetobacterium and Acetate Modify Glucose Homeostasis via Parasympathetic Activation in Zebrafish. Gut Microbes 2021, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Hao, Q.; Xia, R.; Olsen, R.E.; Ringø, E.; Yang, Y.; Zhang, Z.; Ran, C.; Zhou, Z. Nuclease-Treated Stabilized Fermentation Product of Cetobacterium Somerae Improves Growth, Non-Specific Immunity, and Liver Health of Zebrafish (Danio Rerio). Front. Nutr. 2022, 9, 918327. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Derous, D.; Huang, X.; Mitchell, S.E.; Douglas, A.; Lusseau, D.; Wang, Y.; Speakman, J.R. The Effects of Graded Levels of Calorie Restriction: XIX. Impact of Graded Calorie Restriction on Protein Expression in the Liver. J. Gerontol. Ser. A 2023, glad017. [Google Scholar] [CrossRef]
- Zhao, M.; Liu, L.; Liu, F.; Liu, L.; Liu, Z.; Gao, Y.; Cao, J. Traditional Chinese Medicine Improves Myasthenia Gravis by Regulating the Symbiotic Homeostasis of the Intestinal Microbiota and Host. Front. Microbiol. 2022, 13, 1082565. [Google Scholar] [CrossRef]
- Montassier, E.; Gastinne, T.; Vangay, P.; Al-Ghalith, G.A.; Bruley des Varannes, S.; Massart, S.; Moreau, P.; Potel, G.; de La Cochetière, M.F.; Batard, E.; et al. Chemotherapy-Driven Dysbiosis in the Intestinal Microbiome. Aliment. Pharmacol. Ther. 2015, 42, 515–528. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Ma, Y.; Yang, S.; Zhang, S.; Liu, S.; Xiao, J.; Wang, Y.; Wang, W.; Yang, H.; Li, S.; et al. Gut Microbiota-Derived Ursodeoxycholic Acid from Neonatal Dairy Calves Improves Intestinal Homeostasis and Colitis to Attenuate Extended-Spectrum β-Lactamase-Producing Enteroaggregative Escherichia Coli Infection. Microbiome 2022, 10, 79. [Google Scholar] [CrossRef]
- Chen, Y.; Shen, X.; Ma, T.; Yu, X.; Kwok, L.-Y.; Li, Y.; Sun, Z.; Li, D.; Zhang, H. Adjunctive Probio-X Treatment Enhances the Therapeutic Effect of a Conventional Drug in Managing Type 2 Diabetes Mellitus by Promoting Short-Chain Fatty Acid-Producing Bacteria and Bile Acid Pathways. mSystems 2023, 8, e0130022. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Treatment | ||
|---|---|---|---|
| Le | Me | He | |
| Ingredient | % as-is | ||
| Corn | 39.2 | 31.8 | 21.0 |
| Corn gluten meal | 7.0 | 8.1 | 9.2 |
| Chicken fat | - | 6.3 | 16.0 |
| Broken rice | 11 | 11 | 11 |
| Beef meal | 6.5 | 6.5 | 6.5 |
| Bean pulp | 6 | 6 | 6 |
| Wheat shorts | 5 | 5 | 5 |
| Beet pulp | 5 | 5 | 5 |
| Poultry meal | 5 | 5 | 5 |
| Beef and bone meal | 3 | 3 | 3 |
| Vitamin and mineral premix | 3 | 3 | 3 |
| Liquid palatant | 3 | 3 | 3 |
| Hydrolyzed spray-dried chicken blood cells | 2 | 2 | 2 |
| Beer yeast powder | 2 | 2 | 2 |
| CaHPO₄ | 1.3 | 1.3 | 1.3 |
| Chicken liver meal | 1 | 1 | 1 |
| Analyzed composition | |||
| Dry matter (DM), % | 93.85 | 93.89 | 94.25 |
| % DM | |||
| Crude protein | 29.99 | 28.93 | 29.06 |
| Crude fat | 4.69 | 10.22 | 19.74 |
| Nitrogen-free extracts 1 | 53.38 | 48.91 | 39.40 |
| Crude fiber | 4.18 | 4.10 | 4.55 |
| Ash | 7.76 | 7.83 | 7.26 |
| Calcium | 1.04 | 1.08 | 1.02 |
| Phosphorus | 0.81 | 0.78 | 0.80 |
| Lysine | 1.29 | 1.18 | 1.21 |
| GE, MJ/kg 2 | 18.28 | 19.46 | 21.23 |
| ME, MJ/kg 2 | 13.88 | 15.04 | 17.05 |
| Time | Le | Me | He | SEM | p-Value |
|---|---|---|---|---|---|
| T0 | 12.50 | 12.48 | 12.58 | 0.49 | 0.997 |
| T2 | 11.87 | 12.53 | 12.37 | 0.42 | 0.815 |
| T4 | 11.30 | 12.24 | 12.36 | 0.39 | 0.500 |
| T6 | 11.20 | 12.32 | 12.40 | 0.39 | 0.409 |
| T8 | 11.27 | 12.68 | 12.68 | 0.38 | 0.233 |
| Item 1 | Energy Levels | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Le | Me | He | Energy | Liner | Quadratic | ||
| ADG (g/d) | −44.05 a | 7.14 b | 3.27 b | 8.68 | 0.016 | 0.015 | 0.084 |
| ΔBCS 2 | −0.25 a | 1.00 b | 1.67 b | 1.20 | 0.014 | 0.004 | 0.565 |
| ΔMCS 2 | −0.58 a | 0.25 ab | 0.83 b | 0.93 | 0.024 | 0.007 | 0.757 |
| ΔBFI 2 | −0.83 a | 6.67 ab | 12.50 b | 8.43 | 0.017 | 0.005 | 0.816 |
| Item | GLU 1 | CHO 1 | TG 1 | LDL-C 1 | HDL-C 1 | LDL-C% 1 | HDL-C% 1 | |
|---|---|---|---|---|---|---|---|---|
| Unit | mmol/L | mmol/L | mmol/L | mmol/L | mmol/L | % | % | |
| Reference Values [53] | 3.61–6.55 | 3.50–6.99 | 0.20–1.30 | - | - | - | - | |
| T0 | Le | 4.89 AB | 5.44 | 0.62 A | 0.39 | 4.18 | 7.13 A | 77.03 B |
| Me | 4.96 | 5.84 | 0.67 | 0.43 | 4.36 | 7.15 A | 75.80 | |
| He | 5.34 A | 5.44 A | 0.62 | 0.38 A | 4.28 | 6.82 A | 79.27 B | |
| T4 | Le | 5.59 B | 6.09 | 0.70 AB | 0.54 | 4.13 | 8.88 AB | 68.34 A |
| Me | 5.64 | 7.01 | 0.69 | 0.66 | 4.60 | 9.28 B | 66.40 | |
| He | 6.41 B | 6.97 B | 0.84 | 0.62 B | 4.82 | 8.89 B | 69.45 A | |
| T8 | Le | 4.45 A | 5.43 | 0.88 B | 0.56 | 3.74 a | 10.50 B | 69.28 A |
| Me | 4.69 | 6.85 | 0.65 | 0.73 | 4.28 ab | 10.17 B | 65.02 | |
| He | 4.95 A | 6.70 B | 0.72 | 0.62 B | 4.82 b | 9.21 B | 72.58 AB | |
| SEM | 0.11 | 0.18 | 0.03 | 0.03 | 0.08 | 0.28 | 1.05 | |
| p-value | Time | <0.001 | 0.030 | 0.142 | 0.001 | 0.392 | <0.001 | <0.001 |
| Energy | 0.058 | 0.085 | 0.591 | 0.317 | 0.005 | 0.662 | 0.192 | |
| Time × Energy | 0.840 | 0.755 | 0.217 | 0.935 | 0.286 | 0.915 | 0.923 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, H.; Zhang, Q.; Xu, C.; Mao, A.; Zhao, H.; Chen, M.; Sun, W.; Li, G.; Zhang, T. Different Diet Energy Levels Alter Body Condition, Glucolipid Metabolism, Fecal Microbiota and Metabolites in Adult Beagle Dogs. Metabolites 2023, 13, 554. https://doi.org/10.3390/metabo13040554
Sun H, Zhang Q, Xu C, Mao A, Zhao H, Chen M, Sun W, Li G, Zhang T. Different Diet Energy Levels Alter Body Condition, Glucolipid Metabolism, Fecal Microbiota and Metabolites in Adult Beagle Dogs. Metabolites. 2023; 13(4):554. https://doi.org/10.3390/metabo13040554
Chicago/Turabian StyleSun, Haoran, Qiaoru Zhang, Chao Xu, Aipeng Mao, Hui Zhao, Miao Chen, Weili Sun, Guangyu Li, and Tietao Zhang. 2023. "Different Diet Energy Levels Alter Body Condition, Glucolipid Metabolism, Fecal Microbiota and Metabolites in Adult Beagle Dogs" Metabolites 13, no. 4: 554. https://doi.org/10.3390/metabo13040554