
Lipid Differences and Related Metabolism Present on the Hand Skin Surface of Different-Aged Asiatic Females—An Untargeted Metabolomics Study
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Study Participants
2.3. Sample Collection and Preparation
2.4. Ultraperformance Liquid Chromatography Analysis
2.5. Mass Spectrometry Analysis
2.6. Data Extraction and Analysis
3. Results
3.1. Differences in Right-Hand SSL in Females of Different Age Groups
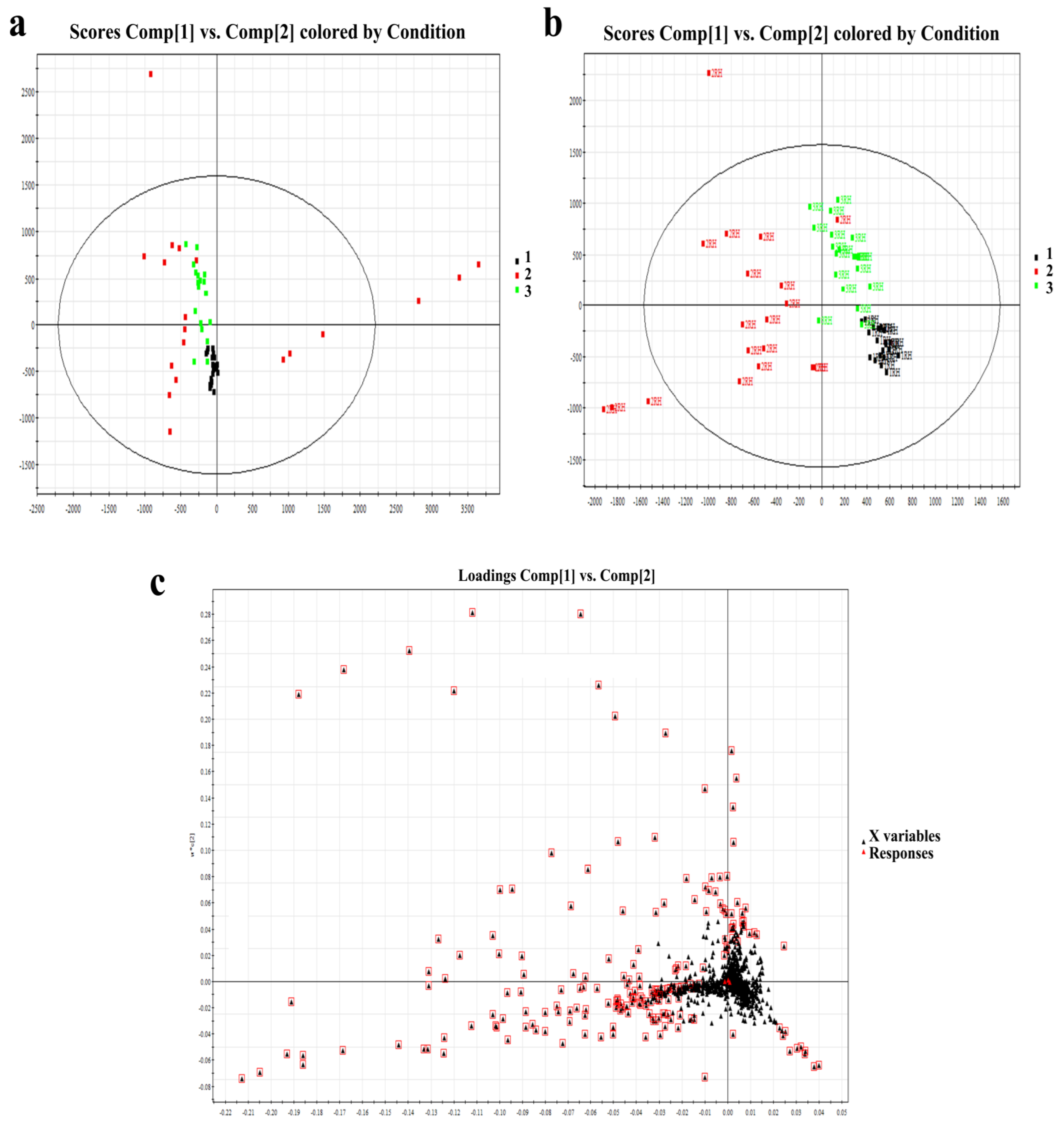
3.2. Multivariate Data Analysis of Right-Hand SSL in Females of Different Age Groups
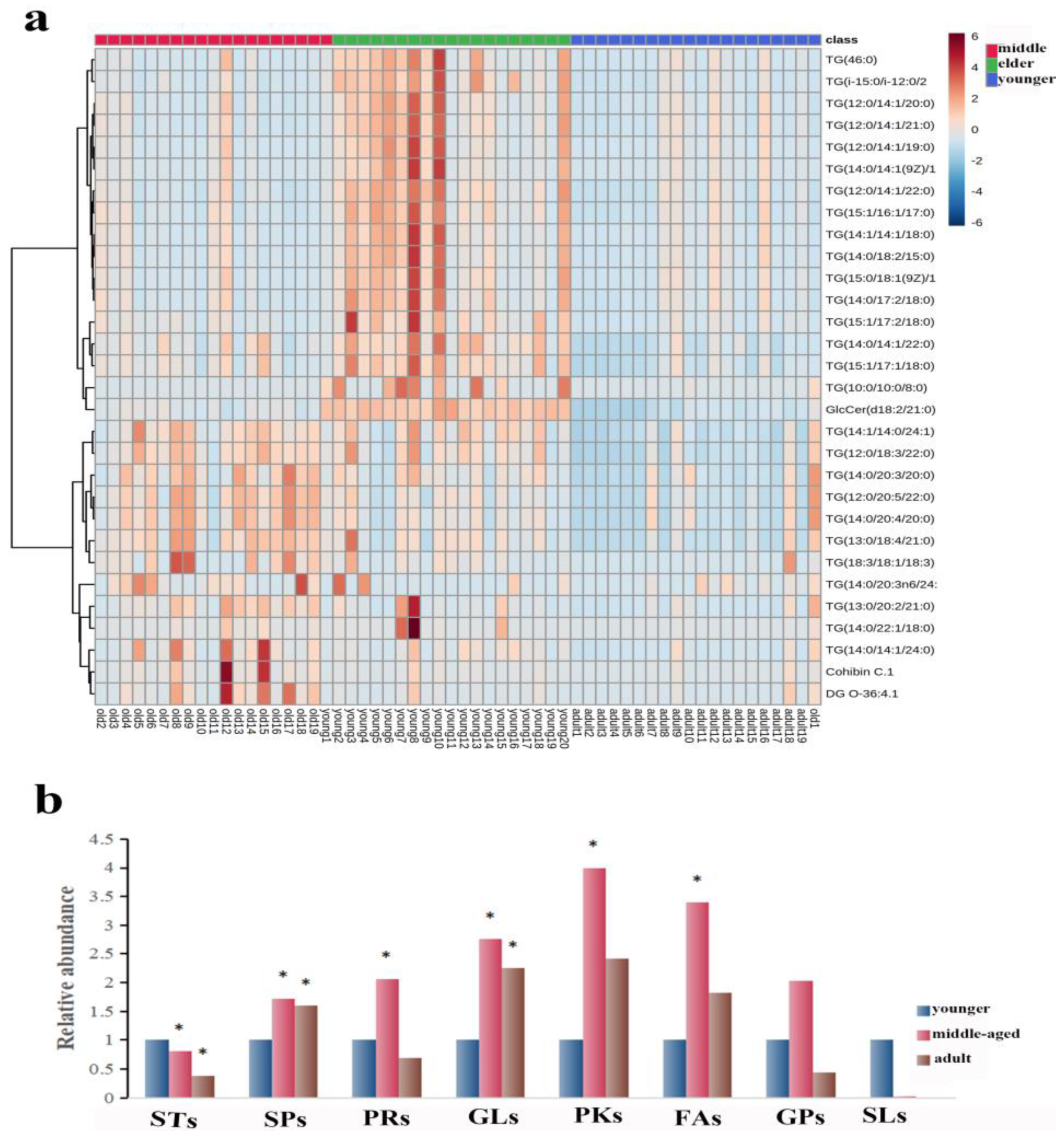
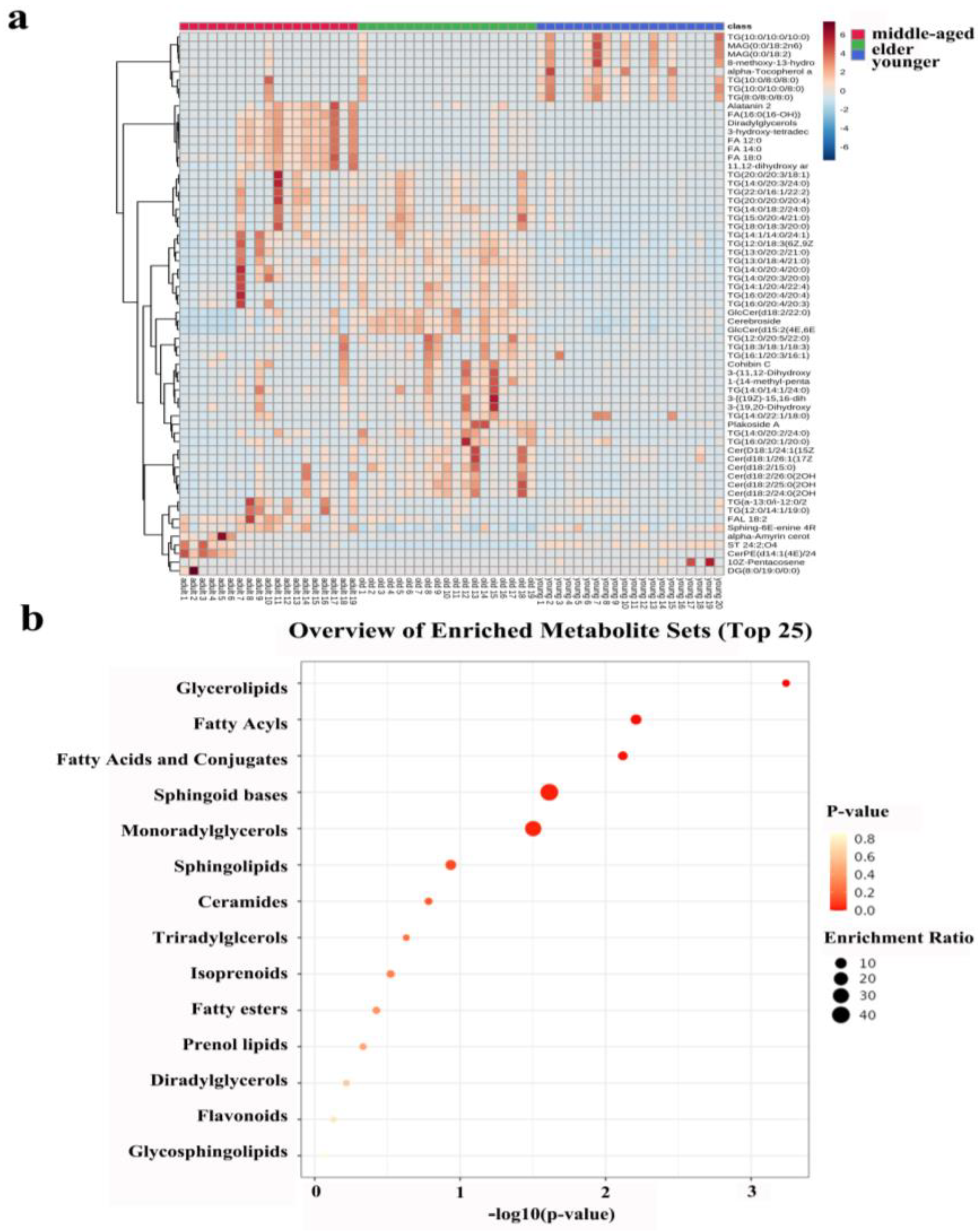
3.2.1. Sixty-Three Individual Lipids Were Responsible for the Differences between Skin Surface Lipids in Skin Samples
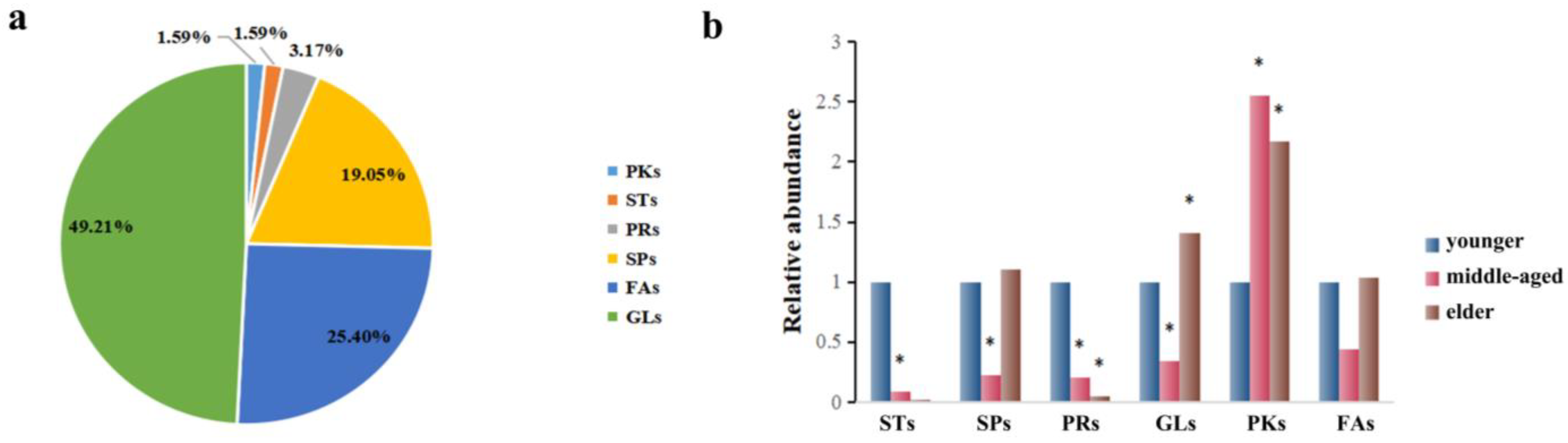
3.2.2. Classification and Relative Abundance Alterations of Specific Differentiated Lipid Metabolites in Skin Samples
3.3. Enrichment Analysis of Skin Lipid Metabolism of Right-Hand SSL in Females of Different Age Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, R.A.; Ghosh, K.; Tonnesen, M.G. Tissue engineering for cutaneous wounds. J. Investig. Dermatol. 2007, 127, 1018–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Q.; Hill, M.C.; Chen, F.; Mo, Q.; Ku, A.T.; Ramos, C.; Sock, E.; Lefebvre, V.; Nguyen, H. SOX11 and SOX4 drive the reactivation of an embryonic gene program during murine wound repair. Nat. Commun. 2019, 10, 4042. [Google Scholar] [CrossRef] [Green Version]
- Breiden, B.; Sandhoff, K. The role of sphingolipid metabolism in cutaneous permeability barrier formation. Biochim. Et Biophys. Acta 2014, 1841, 441–452. [Google Scholar] [CrossRef]
- Morelli, P.; Gaspari, M.; Gabriele, C.; Dastoli, S.; Bennardo, L.; Pavel, A.B.; Patruno, C.; Del Duca, E.; Nisticò, S.P. Proteomic analysis from skin swabs reveals a new set of proteins identifying skin impairment in atopic dermatitis. Exp. Dermatol. 2021, 30, 811–819. [Google Scholar] [CrossRef]
- Hänel, K.H.; Cornelissen, C.; Lüscher, B.; Baron, J.M. Cytokines and the skin barrier. Int. J. Mol. Sci. 2013, 14, 6720–6745. [Google Scholar] [CrossRef] [Green Version]
- Feingold, K.R. Lamellar bodies: The key to cutaneous barrier function. J. Investig. Dermatol. 2012, 132, 1951–1953. [Google Scholar] [CrossRef] [Green Version]
- Hadgraft, J.; Lane, M.E. Transepidermal water loss and skin site: A hypothesis. Int. J. Pharm. 2009, 373, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Biniek, K.; Kaczvinsky, J.; Matts, P.; Dauskardt, R.H. Understanding age-induced alterations to the biomechanical barrier function of human stratum corneum. J. Dermatol. Sci. 2015, 80, 94–101. [Google Scholar] [CrossRef]
- Rittié, L.; Fisher, G.J. Natural and sun-induced aging of human skin. Cold Spring Harb. Perspect. Med. 2015, 5, a015370. [Google Scholar] [CrossRef]
- Chalyk, N.E.; Bandaletova, T.Y.; Kyle, N.H.; Petyaev, I.M. Morphological Characteristics of Residual Skin Surface Components Collected from the Surface of Facial Skin in Women of Different Age. Ann. Dermatol. 2017, 29, 454–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bize, C.; Le Gélébart, E.; Moga, A.; Payré, B.; Garcia, C. Barrier disruption, dehydration and inflammation: Investigation of the vicious circle underlying dry skin. Int. J. Cosmet. Sci. 2021, 43, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Langan, S.M.; Irvine, A.D.; Weidinger, S. Atopic dermatitis. Lancet 2020, 396, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Han, X. Lipidomics for studying metabolism. Nat. Rev. Endocrinol. 2016, 12, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ning, S.; Ghandi, M.; Kryukov, G.V.; Gopal, S.; Deik, A.; Souza, A.; Pierce, K.; Keskula, P.; Hernandez, D.; et al. The landscape of cancer cell line metabolism. Nat. Med. 2019, 25, 850–860. [Google Scholar] [CrossRef]
- Chatelaine, H.; Dey, P.; Mo, X.; Mah, E.; Bruno, R.S.; Kopec, R.E. Vitamin A and D Absorption in Adults with Metabolic Syndrome versus Healthy Controls: A Pilot Study Utilizing Targeted and Untargeted LC-MS Lipidomics. Mol. Nutr. Food Res. 2021, 65, e2000413. [Google Scholar] [CrossRef]
- Sobolev, V.V.; Mezentsev, A.V.; Ziganshin, R.H.; Soboleva, A.G.; Denieva, M.; Korsunskaya, I.M.; Svitich, O.A. LC-MS/MS analysis of lesional and normally looking psoriatic skin reveals significant changes in protein metabolism and RNA processing. PLoS ONE 2021, 16, e0240956. [Google Scholar] [CrossRef]
- Jeong, H.; Kim, S.; Seo, Y.; Koh, J.; Baek, J. Investigation of symptoms of hand skin changes with aging in Korean women and development of a new standard grading scale for hand aging. Ski. Res. Technol. Off. J. Int. Soc. Bioeng. Ski. 2020, 26, 788–793. [Google Scholar] [CrossRef]
- Carmeli, E.; Patish, H.; Coleman, R. The aging hand. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2003, 58, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Shetage, S.S.; Traynor, M.J.; Brown, M.B.; Chilcott, R.P. Sebomic identification of sex- and ethnicity-specific variations in residual skin surface components (RSSC) for bio-monitoring or forensic applications. Lipids Health Dis. 2018, 17, 194. [Google Scholar] [CrossRef] [Green Version]
- Xian, M.; Wawrzyniak, P.; Rückert, B.; Duan, S.; Meng, Y.; Sokolowska, M.; Globinska, A.; Zhang, L.; Akdis, M.; Akdis, C.A. Anionic surfactants and commercial detergents decrease tight junction barrier integrity in human keratinocytes. J. Allergy Clin. Immunol. 2016, 138, 890–893.e899. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Vierkötter, A.; Schikowski, T.; Hüls, A.; Ding, A.; Matsui, M.S.; Deng, B.; Ma, C.; Ren, A.; Zhang, J.; et al. Epidemiological evidence that indoor air pollution from cooking with solid fuels accelerates skin aging in Chinese women. J. Dermatol. Sci. 2015, 79, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Guo, S.; He, Y.; Zuo, Q.; Liu, D.; Xiao, M.; Fan, J.; Li, X. COVID-19 Is Distinct From SARS-CoV-2-Negative Community-Acquired Pneumonia. Front. Cell. Infect. Microbiol. 2020, 10, 322. [Google Scholar] [CrossRef]
- Alexander, L.; Wood, C.M.; Gaskin, P.L.R.; Sawiak, S.J.; Fryer, T.D.; Hong, Y.T.; McIver, L.; Clarke, H.F.; Roberts, A.C. Over-activation of primate subgenual cingulate cortex enhances the cardiovascular, behavioral and neural responses to threat. Nat. Commun. 2020, 11, 5386. [Google Scholar] [CrossRef]
- Huang, J.; Heng, S.; Zhang, W.; Liu, Y.; Xia, T.; Ji, C.; Zhang, L.J. Dermal extracellular matrix molecules in skin development, homeostasis, wound regeneration and diseases. Semin. Cell Dev. Biol. 2022, 128, 137–144. [Google Scholar] [CrossRef]
- Starr, N.J.; Johnson, D.J.; Wibawa, J.; Marlow, I.; Bell, M.; Barrett, D.A.; Scurr, D.J. Age-Related Changes to Human Stratum Corneum Lipids Detected Using Time-of-Flight Secondary Ion Mass Spectrometry Following in Vivo Sampling. Anal. Chem. 2016, 88, 4400–4408. [Google Scholar] [CrossRef] [PubMed]
- Zorina, A.; Zorin, V.; Kudlay, D.; Kopnin, P. Molecular Mechanisms of Changes in Homeostasis of the Dermal Extracellular Matrix: Both Involutional and Mediated by Ultraviolet Radiation. Int. J. Mol. Sci. 2022, 23, 6655. [Google Scholar] [CrossRef] [PubMed]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [Green Version]
- Low, E.; Alimohammadiha, G.; Smith, L.A.; Costello, L.F.; Przyborski, S.A.; von Zglinicki, T.; Miwa, S. How good is the evidence that cellular senescence causes skin ageing? Ageing Res. Rev. 2021, 71, 101456. [Google Scholar] [CrossRef]
- Jia, Y.; Gan, Y.; He, C.; Chen, Z.; Zhou, C. The mechanism of skin lipids influencing skin status. J. Dermatol. Sci. 2018, 89, 112–119. [Google Scholar] [CrossRef]
- de Szalay, S.; Wertz, P.W. Protective Barriers Provided by the Epidermis. Int. J. Mol. Sci. 2023, 24, 3145. [Google Scholar] [CrossRef]
- Feingold, K.R. Thematic review series: Skin lipids. The role of epidermal lipids in cutaneous permeability barrier homeostasis. J. Lipid Res. 2007, 48, 2531–2546. [Google Scholar] [CrossRef] [Green Version]
- Elias, P.M. The skin barrier as an innate immune element. Semin. Immunopathol. 2007, 29, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Wohlrab, J.; Gabel, A.; Wolfram, M.; Grosse, I.; Neubert, R.H.H.; Steinbach, S.C. Age- and Diabetes-Related Changes in the Free Fatty Acid Composition of the Human Stratum Corneum. Ski. Pharmacol. Physiol. 2018, 31, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Mika, A.; Pakiet, A.; Szczygielski, O.; Woźniak, K.; Osipowicz, K.; Kowalewski, C.; Krześniak, N.; Noszczyk, B.H.; Wertheim-Tysarowska, K. Fatty acid profiles in various lipid fractions in the female epidermis. Does the body site and age matter? Acta Biochim. Pol. 2022, 69, 657–671. [Google Scholar] [CrossRef]
- Sanford, J.A.; O’Neill, A.M.; Zouboulis, C.C.; Gallo, R.L. Short-Chain Fatty Acids from Cutibacterium acnes Activate Both a Canonical and Epigenetic Inflammatory Response in Human Sebocytes. J. Immunol. 2019, 202, 1767–1776. [Google Scholar] [CrossRef] [Green Version]
- Pilkington, S.M.; Watson, R.E.; Nicolaou, A.; Rhodes, L.E. Omega-3 polyunsaturated fatty acids: Photoprotective macronutrients. Exp. Dermatol. 2011, 20, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Chiang, H.M.; Chan, S.Y.; Chu, Y.; Wen, K.C. Fisetin Ameliorated Photodamage by Suppressing the Mitogen-Activated Protein Kinase/Matrix Metalloproteinase Pathway and Nuclear Factor-κB Pathways. J. Agric. Food Chem. 2015, 63, 4551–4560. [Google Scholar] [CrossRef]
- Nováčková, A.; Sagrafena, I.; Pullmannová, P.; Paraskevopoulos, G.; Dwivedi, A.; Mazumder, A.; Růžičková, K.; Slepička, P.; Zbytovská, J.; Vávrová, K. Acidic pH Is Required for the Multilamellar Assembly of Skin Barrier Lipids In Vitro. J. Investig. Dermatol. 2021, 141, 1915–1921.e1914. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.L.; Timmermann, V.; Illing, T.; Elsner, P. Contact Dermatitis in the Elderly: Predisposing Factors, Diagnosis, and Management. Drugs Aging 2019, 36, 411–417. [Google Scholar] [CrossRef]
- Lodén, M.; Bárány, E. Skin-identical lipids versus petrolatum in the treatment of tape-stripped and detergent-perturbed human skin. Acta Derm. Venereol. 2000, 80, 412–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.J.; Kim, Y.K.; Kim, S.; Kim, J.E.; Tian, Y.D.; Doh, E.J.; Lee, D.H.; Chung, J.H. Adipochemokines induced by ultraviolet irradiation contribute to impaired fat metabolism in subcutaneous fat cells. Br. J. Dermatol. 2018, 178, 492–501. [Google Scholar] [CrossRef]
- Li, W.H.; Pappas, A.; Zhang, L.; Ruvolo, E.; Cavender, D. IL-11, IL-1α, IL-6, and TNF-α are induced by solar radiation in vitro and may be involved in facial subcutaneous fat loss in vivo. J. Dermatol. Sci. 2013, 71, 58–66. [Google Scholar] [CrossRef]
- Kim, E.J.; Kim, Y.K.; Kim, J.E.; Kim, S.; Kim, M.K.; Park, C.H.; Chung, J.H. UV modulation of subcutaneous fat metabolism. J. Investig. Dermatol. 2011, 131, 1720–1726. [Google Scholar] [CrossRef] [Green Version]
- Kuehne, A.; Hildebrand, J.; Soehle, J.; Wenck, H.; Terstegen, L.; Gallinat, S.; Knott, A.; Winnefeld, M.; Zamboni, N. An integrative metabolomics and transcriptomics study to identify metabolic alterations in aged skin of humans in vivo. BMC Genom. 2017, 18, 169. [Google Scholar] [CrossRef] [Green Version]
- Martin, R.P.; Varela, P.; Gomes, C.P.; Marins, M.M.; Filippelli-Silva, R.; Yarak, S.; Soares, J.L.M.; Sanudo, A.; Idkowiak-Baldys, J.; Chen, S.; et al. Transcriptomic and histological analysis of exposed facial skin areas wrinkled or not and unexposed skin. Mol. Biol. Rep. 2022, 49, 1669–1678. [Google Scholar] [CrossRef]
- Dalmau, N.; Andrieu-Abadie, N.; Tauler, R.; Bedia, C. Phenotypic and lipidomic characterization of primary human epidermal keratinocytes exposed to simulated solar UV radiation. J. Dermatol. Sci. 2018, 92, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Egawa, M.; Tagami, H. Comparison of the depth profiles of water and water-binding substances in the stratum corneum determined in vivo by Raman spectroscopy between the cheek and volar forearm skin: Effects of age, seasonal changes and artificial forced hydration. Br. J. Dermatol. 2008, 158, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, N.; Yan, Y.Q.; Liu, Y.; Xiong, K.; Liu, Y.; Xia, Q.M.; Zhang, H.; Liu, Z.D. Recent advances in the anti-aging effects of phytoestrogens on collagen, water content, and oxidative stress. Phytother. Res. PTR 2020, 34, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Bedord, C.J.; Young, J.M. A comparison of comedonal and skin surface lipids from hairless dogs showing clinical signs of acne. J. Investig. Dermatol. 1981, 77, 341–344. [Google Scholar] [CrossRef] [Green Version]
- Lasch, J.; Schönfelder, U.; Walke, M.; Zellmer, S.; Beckert, D. Oxidative damage of human skin lipids. Dependence of lipid peroxidation on sterol concentration. Biochim. Et Biophys. Acta 1997, 1349, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Camera, E.; Ludovici, M.; Tortorella, S.; Sinagra, J.L.; Capitanio, B.; Goracci, L.; Picardo, M. Use of lipidomics to investigate sebum dysfunction in juvenile acne. J. Lipid Res. 2016, 57, 1051–1058. [Google Scholar] [CrossRef] [Green Version]
- Misawa, E.; Tanaka, M.; Saito, M.; Nabeshima, K.; Yao, R.; Yamauchi, K.; Abe, F.; Yamamoto, Y.; Furukawa, F. Protective effects of Aloe sterols against UVB-induced photoaging in hairless mice. Photodermatol. Photoimmunol. Photomed. 2017, 33, 101–111. [Google Scholar] [CrossRef]
- Nojiri, K.; Fudetani, S.; Arai, A.; Kitamura, T.; Sassa, T.; Kihara, A. Impaired Skin Barrier Function Due to Reduced ω-O-Acylceramide Levels in a Mouse Model of Sjögren-Larsson Syndrome. Mol. Cell. Biol. 2021, 41, e0035221. [Google Scholar] [CrossRef] [PubMed]
- Rinnov, M.R.; Halling, A.S.; Gerner, T.; Ravn, N.H.; Knudgaard, M.H.; Trautner, S.; Goorden, S.M.I.; Ghauharali-van der Vlugt, K.J.M.; Stet, F.S.; Skov, L.; et al. Skin biomarkers predict development of atopic dermatitis in infancy. Allergy 2022, 78, 791–802. [Google Scholar] [CrossRef]
- Toncic, R.J.; Jakasa, I.; Hadzavdic, S.L.; Goorden, S.M.; Vlugt, K.J.G.; Stet, F.S.; Balic, A.; Petkovic, M.; Pavicic, B.; Zuzul, K.; et al. Altered Levels of Sphingosine, Sphinganine and Their Ceramides in Atopic Dermatitis Are Related to Skin Barrier Function, Disease Severity and Local Cytokine Milieu. Int. J. Mol. Sci. 2020, 21, 1958. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (min) | A Phase (%) | B Phase (%) |
|---|---|---|
| 0.0 | 80 | 20 |
| 1.0 | 80 | 20 |
| 5.0 | 40 | 60 |
| 11.0 | 20 | 80 |
| 18.0 | 10 | 90 |
| 19.0 | 0 | 100 |
| 21.0 | 0 | 100 |
| 21.1 | 80 | 20 |
| 22.0 | 80 | 20 |
| Retention Time (min) | m/z | Formula | Description | Anova (p) | Highest Mean |
|---|---|---|---|---|---|
| 0.8523 | 262.2384 | C14H28O3 | 3-hydroxy-tetradecanoic acid | 0.000 | middle-aged |
| 0.8523 | 218.2124 | C12H24O2 | FA(12:0) | 0.000 | middle-aged |
| 1.8561 | 302.3059 | C18H36O2 | FA(18:0) | 0.000 | middle-aged |
| 0.9692 | 290.2702 | C16H32O3 | FA(16:0(16-OH)) | 0.000 | middle-aged |
| 0.9692 | 246.2436 | C14H28O2 | FA(14:0) | 0.000 | middle-aged |
| 4.3784 | 282.2800 | C18H32O | FAL(18:2) | 0.000 | middle-aged |
| 13.2548 | 575.5057 | C37H66O4 | 3-(11,12-Dihydroxy-15,19-dotriacontadienyl)-5-methyl-2(5H)-furanone, 9ci | 0.000 | elder |
| 13.7980 | 577.5213 | C37H68O4 | 3-[(19Z)-15,16-dihydroxydotriacont-19-en-1-yl]-5-methyl-5H-furan-2-one | 0.000 | elder |
| 6.6815 | 355.2852 | C21H38O4 | MAG(0:0/18:2) | 0.000 | younger |
| 7.3001 | 355.2852 | C21H38O4 | MAG(0:0/18:2n6) | 0.000 | younger |
| 5.6706 | 368.4260 | C25H50 | 10Z-Pentacosene | 0.002 | younger |
| 13.2617 | 577.5207 | C37H68O4 | Cohibin C | 0.002 | elder |
| 5.8905 | 327.2537 | C19H34O4 | FA(19:2;O2) | 0.006 | middle-aged |
| 13.7980 | 551.5058 | C35H66O4 | 3-(19,20-Dihydroxytriacontyl)-5-methyl-2(5H)-furanone | 0.006 | elder |
| 1.2578 | 274.2747 | C16H32O2 | Diradylglycerols | 0.046 | middle-aged |
| 1.2509 | 362.3267 | C20H40O4 | 11,12-dihydroxy arachidic acid | 0.048 | middle-aged |
| 13.8186 | 876.8028 | C55H102O6 | TG(14:1/14:0/24:1) | 0.000 | middle-aged |
| 13.2479 | 874.7869 | C55H100O6 | TG(12:0/18:3(6Z,9Z,12Z)/22:0) | 0.000 | elder |
| 14.4376 | 904.8344 | C57H106O6 | TG(13:0/20:2/21:0) | 0.000 | elder |
| 12.7256 | 872.7716 | C55H98O6 | TG(13:0/18:4/21:0) | 0.000 | elder |
| 13.2548 | 900.8024 | C57H102O6 | TG(14:0/20:4/20:0) | 0.000 | elder |
| 12.7325 | 881.7627 | C57H100O6 | TG(12:0/20:5/22:0) | 0.000 | elder |
| 13.8049 | 603.5368 | C39H70O4 | DG(O-36:4) | 0.000 | elder |
| 13.8255 | 902.8183 | C57H104O6 | TG(14:0/20:3/20:0) | 0.000 | middle-aged |
| 7.9599 | 572.4903 | C33H62O6 | TG(10:0/10:0/10:0) | 0.000 | younger |
| 14.4376 | 878.8193 | C55H104O6 | TG(14:0/14:1/24:0) | 0.000 | elder |
| 7.3070 | 544.4583 | C31H58O6 | TG(10:0/10:0/8:0) | 0.000 | younger |
| 14.4582 | 956.8639 | C61H110O6 | TG(20:0/20:3/18:1) | 0.000 | middle-aged |
| 15.7642 | 960.8970 | C61H114O6 | TG(14:0/20:2/24:0) | 0.000 | elder |
| 6.1036 | 488.3956 | C27H50O6 | TG(8:0/8:0/8:0) | 0.001 | younger |
| 15.0217 | 958.8799 | C61H112O6 | TG(14:0/20:3/24:0) | 0.001 | middle-aged |
| 6.6746 | 516.4270 | C29H54O6 | TG(10:0/8:0/8:0) | 0.001 | younger |
| 15.7504 | 934.8814 | C59H112O6 | TG(16:0/20:1/20:0) | 0.001 | elder |
| 12.2370 | 901.7284 | C59H96O6 | TG(14:1/20:4/22:4) | 0.001 | elder |
| 12.7325 | 903.7431 | C59H98O6 | TG(16:0/20:4/20:4) | 0.001 | elder |
| 13.2617 | 905.7588 | C59H100O6 | TG(16:0/20:4/20:3) | 0.001 | elder |
| 15.0767 | 932.8662 | C59H110O6 | TG(14:0/18:2/24:0) | 0.002 | elder |
| 11.8041 | 894.7567 | C57H96O6 | TG(18:3/18:1/18:3) | 0.003 | elder |
| 15.0905 | 906.8505 | C57H108O6 | TG(14:0/22:1/18:0) | 0.004 | elder |
| 13.9492 | 928.8351 | C59H106O6 | TG(15:0/20:4/21:0) | 0.004 | elder |
| 14.4720 | 930.8490 | C59H108O6 | TG(18:0/18:3/20:0) | 0.006 | elder |
| 12.7187 | 782.7254 | C48H92O6 | TG(a-13:0/i-12:0/20:0) | 0.009 | middle-aged |
| 15.6336 | 986.9108 | C63H116O6 | TG(22:0/16:1/22:2) | 0.009 | middle-aged |
| 15.0630 | 984.8953 | C63H114O6 | TG(20:0/20:0/20:4) | 0.017 | middle-aged |
| 12.3264 | 780.7093 | C48H90O6 | TG(12:0/14:1/19:0) | 0.020 | middle-aged |
| 12.2714 | 870.7570 | C55H96O6 | TG(16:1/20:3/16:1) | 0.023 | elder |
| 6.6609 | 516.4614 | C30H58O5 | DG(8:0/19:0/0:0) | 0.039 | middle-aged |
| 9.6592 | 832.2423 | C39H43O20 | Alatanin 2 | 0.023 | middle-aged |
| 8.0079 | 473.4000 | C31H52O3 | alpha-Tocopherol acetate | 0.000 | younger |
| 13.1654 | 822.8074 | C56H100O2 | alpha-Amyrin cerotate | 0.018 | middle-aged |
| 8.5101 | 838.7146 | C50H95NO8 | Cerebroside | 0.000 | elder |
| 6.6540 | 740.6045 | C43H81NO8 | GlcCer(d15:2(4E,6E)/22:0) | 0.000 | elder |
| 5.4162 | 413.2673 | C18H37NO6S | Sphing-6E-enine 4R-sufate | 0.000 | middle-aged |
| 7.3688 | 522.4893 | C33H63NO3 | Cer(d18:2/15:0) | 0.000 | elder |
| 9.7967 | 678.6408 | C43H83NO4 | Cer(d18:2/25:0(2OH)) | 0.000 | elder |
| 7.4169 | 782.6516 | C46H87NO8 | GlcCer(d18:2/22:0) | 0.000 | elder |
| 11.9829 | 948.7871 | C57H105NO9 | Plakoside A | 0.000 | elder |
| 9.7760 | 648.6302 | C42H81NO3 | Cer(D18:1/24:1(15Z)) | 0.001 | elder |
| 9.4457 | 664.6251 | C42H81NO4 | Cer(d18:2/24:0(2OH)) | 0.001 | elder |
| 8.0972 | 750.6111 | C40H81N2O7P | CerPE(d14:1(4E)/24:0(2OH)) | 0.002 | middle-aged |
| 10.4633 | 676.6615 | C44H85NO3 | Cer(d18:1/26:1(17Z)) | 0.003 | elder |
| 10.1333 | 692.6557 | C44H85NO4 | Cer(d18:2/26:0(2OH)) | 0.010 | elder |
| 21.6564 | 391.2851 | C24H38O4 | ST(24:2;O4) | 0.000 | younger |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.; Wang, J.; Mao, Z. Lipid Differences and Related Metabolism Present on the Hand Skin Surface of Different-Aged Asiatic Females—An Untargeted Metabolomics Study. Metabolites 2023, 13, 553. https://doi.org/10.3390/metabo13040553
Chen T, Wang J, Mao Z. Lipid Differences and Related Metabolism Present on the Hand Skin Surface of Different-Aged Asiatic Females—An Untargeted Metabolomics Study. Metabolites. 2023; 13(4):553. https://doi.org/10.3390/metabo13040553
Chicago/Turabian StyleChen, Tian, Juan Wang, and Zhenxing Mao. 2023. "Lipid Differences and Related Metabolism Present on the Hand Skin Surface of Different-Aged Asiatic Females—An Untargeted Metabolomics Study" Metabolites 13, no. 4: 553. https://doi.org/10.3390/metabo13040553