
Mycotoxin Contamination Status of Cereals in China and Potential Microbial Decontamination Methods


Abstract
:1. Introduction
2. Cereal Contamination by Mycotoxins in China and the Existing Decontamination Methods
2.1. Impact of Climate Change on the Mycotoxin Contamination Rate of Cereals in China
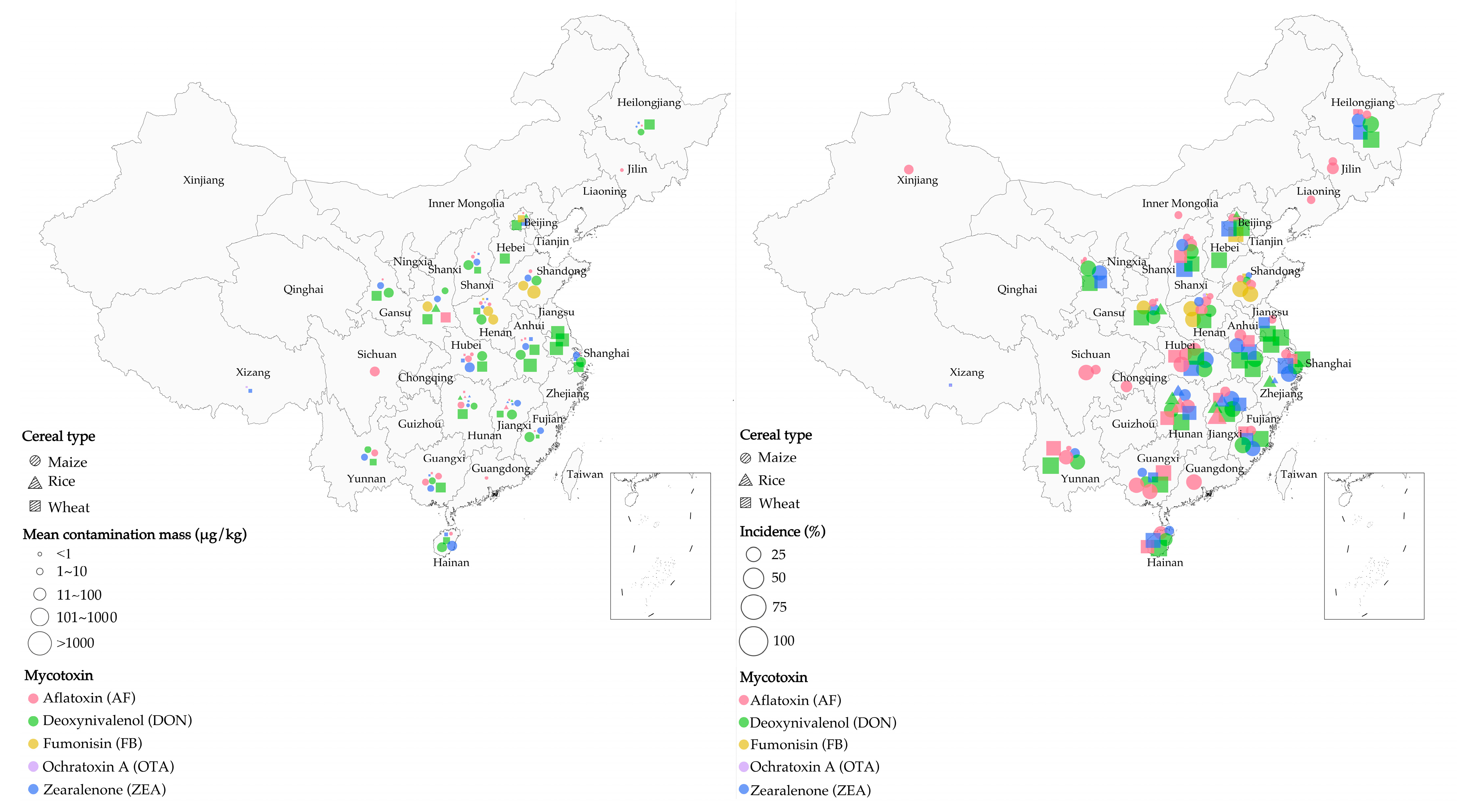
2.2. Correlation between Cereal Types and Mycotoxin Contaminants in China
2.3. Existing Mycotoxin Detoxification Methods
3. Microbial Methods of Inhibiting Mycotoxin Growth and Detoxifying Mycotoxins
3.1. Decontamination Methods Based on Microorganisms
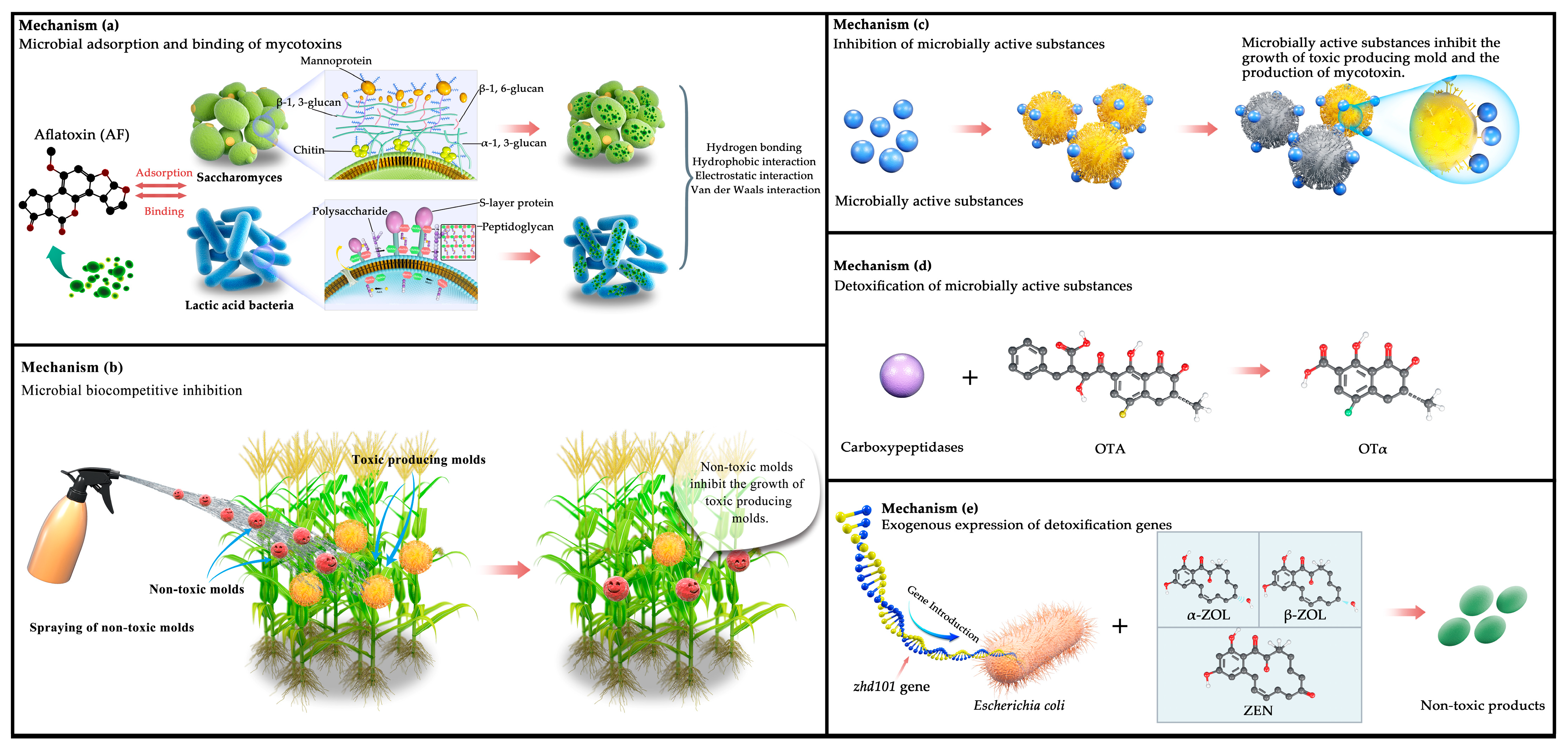
3.1.1. Adsorption and Binding Using Microorganisms
3.1.2. Biocompetitive Inhibition Using Microorganisms
3.2. Microbial Active Substance Decontamination Methods
3.2.1. Inhibition Using Microbial Active Substances
3.2.2. Detoxification Using Microbial Active Substances
3.3. Other Microbial Inhibition Methods
3.4. Mechanism of Mycotoxin Reduction Using Microbiological Methods
4. Conclusions and Prospects
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef]
- Chhaya, R.S.; O’Brien, J.; Cummins, E. Feed to fork risk assessment of mycotoxins under climate change influences—Recent developments. Trends Food Sci. Technol. 2022, 126, 126–141. [Google Scholar] [CrossRef]
- Krska, R.; Schubert-Ullrich, P.; Molinelli, A.; Sulyok, M.; MacDonald, S.; Crews, C. Mycotoxin analysis: An update. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2008, 25, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Castegnaro, M.; Wild, C.P. IARC activities in mycotoxin research. Nat. Toxins 1995, 3, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-Q.; Yoshizawa, T.; Kawamura, O.; Luo, X.-Y.; Li, Y.-W. Aflatoxins and Fumonisins in Corn from the High-Incidence Area for Human Hepatocellular Carcinoma in Guangxi, China. J. Agric. Food Chem. 2001, 49, 4122–4126. [Google Scholar] [CrossRef] [PubMed]
- Wogan, G.N. Aflatoxin as a human carcinogen. Hepatology 1999, 30, 573–575. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Abbitt, B.; Cotter, S.; Murphy, M.; Reagor, J.; Robinson, R.; West, J.; Whitford, H. Bovine abortion and death associated with consumption of aflatoxin-contaminated peanuts. J. Am. Vet. Med. Assoc. 1986, 188, 1187–1188. [Google Scholar]
- Lombard, M.J. Mycotoxin Exposure and Infant and Young Child Growth in Africa: What Do We Know? Ann. Nutr. Metab. 2014, 64 (Suppl. 2), 42–52. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petkova-Bocharova, T.; Chernozemsky, I.N.; Castegnaro, M. Ochratoxin A in human blood in relation to Balkan endemic nephropathy and urinary system tumours in Bulgaria. Food Addit. Contam. 1988, 5, 299–301. [Google Scholar] [CrossRef]
- Bbosa, G.; Kitya, D.; Lubega, A.; Ogwal-Okeng, J.; Anokbonggo, W.; Kyegombe, D. Review of the Biological and Health Effects of Aflatoxins on Body Organs and Body Systems. In Aflatoxins: Recent Advances and Future Prospects; Razzaghi-Abyaneh, M., Ed.; InTech: Rijeka, Croatia, 2013; Volume 3, pp. 239–265. [Google Scholar]
- Fouad, A.M.; Ruan, D.; El-Senousey, H.K.; Chen, W.; Jiang, S.; Zheng, C. Harmful Effects and Control Strategies of Aflatoxin B1 Produced by Aspergillus flavus and Aspergillus parasiticus Strains on Poultry: Review. Toxins 2019, 11, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coulombe, R.A., Jr.; Sharma, R.P. Effect of repeated dietary exposure of aflatoxin B1 on brain biogenic amines and metabolites in the rat. Toxicol. Appl. Pharmacol. 1985, 80, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Ogunade, I.M.; Vyas, D.; Adesogan, A.T. Aflatoxin in Dairy Cows: Toxicity, Occurrence in Feedstuffs and Milk and Dietary Mitigation Strategies. Toxins 2021, 13, 283. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, A.; Hernandez, M.; Verde, M.; Sanz, M. Effect of aflatoxin on performance, hematology, and clinical immunology in lambs. Can. J. Vet. Res. 2000, 64, 53–58. [Google Scholar] [PubMed]
- Bui-Klimke, T.R.; Wu, F. Ochratoxin A and human health risk: A review of the evidence. Crit. Rev. Food Sci. Nutr. 2015, 55, 1860–1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denli, M.; Perez, J.F. Ochratoxins in Feed, a Risk for Animal and Human Health: Control Strategies. Toxins 2010, 2, 1065–1077. [Google Scholar] [CrossRef] [PubMed]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Malir, J.; Toman, J. Ochratoxin A: 50 Years of Research. Toxins 2016, 8, 191. [Google Scholar] [CrossRef] [Green Version]
- Kamle, M.; Mahato, D.K.; Devi, S.; Lee, K.E.; Kang, S.G.; Kumar, P. Fumonisins: Impact on Agriculture, Food, and Human Health and their Management Strategies. Toxins 2019, 11, 328. [Google Scholar] [CrossRef] [Green Version]
- Bucci, T.J.; Howard, P.C. Effect of Fumonisin Mycotoxins in Animals. J. Toxicol. Toxin Rev. 1996, 15, 293–302. [Google Scholar] [CrossRef]
- Carlson, D.B.; Williams, D.E.; Spitsbergen, J.M.; Ross, P.F.; Bacon, C.W.; Meredith, F.I.; Riley, R.T. Fumonisin B1 Promotes Aflatoxin B1 and N-Methyl-N′-nitro-nitrosoguanidine-Initiated Liver Tumors in Rainbow Trout. Toxicol. Appl. Pharmacol. 2001, 172, 29–36. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, L.; Wu, D. Effects of zearalenone on proliferation and apoptosis in MCF-7 cells. Zhonghua Yu Fang Yi Xue Za Zhi 2005, 39, 328–331. [Google Scholar] [PubMed]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, J.P.F.; Placinta, C.M.; Macdonald, A.M.C. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed Sci. Technol. 1999, 80, 183–205. [Google Scholar] [CrossRef] [Green Version]
- Dalefield, R. Chapter 21—Mycotoxins and Mushrooms. In Veterinary Toxicology for Australia and New Zealand; Dalefield, R., Ed.; Elsevier: Oxford, UK, 2017; pp. 373–419. [Google Scholar]
- Ahamad, D.B. Toxic Effects of Citrinin in Animals and Poultry. Shanlax Int. J. Vet. Sci. 2019, 5, 12–31. [Google Scholar]
- Sobrova, P.; Adam, V.; Vasatkova, A.; Beklova, M.; Zeman, L.; Kizek, R. Deoxynivalenol and its toxicity. Interdiscip. Toxicol. 2010, 3, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Turna, N.S.; Wu, F. Risk assessment of dietary deoxynivalenol exposure in wheat products worldwide: Are new codex DON guidelines adequately protective? Trends Food Sci. Technol. 2019, 89, 11–25. [Google Scholar] [CrossRef]
- Adhikari, M.; Negi, B.; Kaushik, N.; Adhikari, A.; Al-Khedhairy, A.A.; Kaushik, N.K.; Choi, E.H. T-2 mycotoxin: Toxicological effects and decontamination strategies. Oncotarget 2017, 8, 33933–33952. [Google Scholar] [CrossRef] [Green Version]
- Fleurat-Lessard, F. Integrated management of the risks of stored grain spoilage by seedborne fungi and contamination by storage mould mycotoxins—An update. J. Stored Prod. Res. 2017, 71, 22–40. [Google Scholar] [CrossRef]
- Juan, C.; Covarelli, L.; Beccari, G.; Colasante, V.; Mañes, J. Simultaneous analysis of twenty-six mycotoxins in durum wheat grain from Italy. Food Control 2016, 62, 322–329. [Google Scholar] [CrossRef]
- Tralamazza, S.M.; Bemvenuti, R.H.; Zorzete, P.; de Souza Garcia, F.; Corrêa, B. Fungal diversity and natural occurrence of deoxynivalenol and zearalenone in freshly harvested wheat grains from Brazil. Food Chem. 2016, 196, 445–450. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, S.; Hu, B.; Zhou, Y.; Liang, Z.; Jia, X.; Huang, M.; Wei, J.; Shi, Z. A comprehensive framework for assessing the impact of potential agricultural pollution on grain security and human health in economically developed areas. Environ. Pollut. 2020, 263, 114653. [Google Scholar] [CrossRef]
- Paterson, R.R.M.; Lima, N. Thermophilic Fungi to Dominate Aflatoxigenic/Mycotoxigenic Fungi on Food under Global Warming. Int. J. Environ. Res. Public Health 2017, 14, 199. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, S.J.; Campbell, B.M.; Ingram, J.S.I. Climate Change and Food Systems. Annu. Rev. Environ. Resour. 2012, 37, 195–222. [Google Scholar] [CrossRef] [Green Version]
- Kagot, V.; Okoth, S.; De Boevre, M.; De Saeger, S. Biocontrol of Aspergillus and Fusarium Mycotoxins in Africa: Benefits and Limitations. Toxins 2019, 11, 109. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Hassan, Y.I.; Lepp, D.; Shao, S.; Zhou, T. Strategies and Methodologies for Developing Microbial Detoxification Systems to Mitigate Mycotoxins. Toxins 2017, 9, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannaa, M.; Kim, K.D. Microbe-Mediated Control of Mycotoxigenic Grain Fungi in Stored Rice with Focus on Aflatoxin Biodegradation and Biosynthesis Inhibition. Mycobiology 2016, 44, 67–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonetti, E.; Roberts, I.N.; Montecchia, M.S.; Gutierrez-Boem, F.H.; Gomez, F.M.; Ruiz, J.A. A novel Burkholderia ambifaria strain able to degrade the mycotoxin fusaric acid and to inhibit Fusarium spp. growth. Microbiol. Res. 2018, 206, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Ahad, R.; Zhou, T.; Lepp, D.; Pauls, K.P. Microbial detoxification of eleven food and feed contaminating trichothecene mycotoxins. BMC Biotechnol. 2017, 17, 30. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Halder, J.; White, R.P.; Hughes, D.J.; Ye, Z.; Wang, C.; Xu, R.; Gan, B.; Fitt, B.D.L. Climate change increases risk of fusarium ear blight on wheat in central China. Ann. Appl. Biol. 2014, 164, 384–395. [Google Scholar] [CrossRef] [Green Version]
- Abate, D.; Mitiku, F.; Negash, R. Commercialization level and determinants of market participation of smallholder wheat farmers in northern Ethiopia. Afr. J. Sci. Technol. Innov. Dev. 2022, 14, 428–439. [Google Scholar] [CrossRef]
- Fei, L.; Meijun, Z.; Jiaqi, S.; Zehui, C.; Xiaoli, W.; Jiuchun, Y. Maize, wheat and rice production potential changes in China under the background of climate change. Agric. Sys. 2020, 182, 102853. [Google Scholar] [CrossRef]
- WHO; FAO. Report of the Expert Meeting on Ciguatera Poisoning: Rome, 19–23 November 2018; Food Safaty and Quality Series No. 9. WHO: Rome, Italy, 2020. [Google Scholar]
- Liu, C.; Van der Fels-Klerx, H.J. Quantitative Modeling of Climate Change Impacts on Mycotoxins in Cereals: A Review. Toxins 2021, 13, 276. [Google Scholar] [CrossRef]
- Selvaraj, J.N.; Wang, Y.; Zhou, L.; Zhao, Y.; Xing, F.; Dai, X.; Liu, Y. Recent mycotoxin survey data and advanced mycotoxin detection techniques reported from China: A review. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Wang, S.; Hu, X.; Su, J.; Zhang, Y.; Xie, Y.; Zhang, H.; Tang, L.; Wang, J.S. Co-contamination of aflatoxin B1 and fumonisin B1 in food and human dietary exposure in three areas of China. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2011, 28, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.; Xu, J.; Liu, X.; Yin, X.; Shi, J. Natural occurrence of deoxynivalenol and zearalenone in wheat from Jiangsu province, China. Food Chem. 2014, 157, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Khaneghah, A.M.; Martins, L.M.; von Hertwig, A.M.; Bertoldo, R.; Sant’Ana, A.S. Deoxynivalenol and its masked forms: Characteristics, incidence, control and fate during wheat and wheat based products processing—A review. Trends Food Sci. Technol. 2018, 71, 13–24. [Google Scholar] [CrossRef]
- Deng, Y.Y.; Li, W.; Zhang, P.; Sun, H.Y.; Zhang, X.X.; Zhang, A.X.; Chen, H.G. Fusarium pseudograminearum as an emerging pathogen of crown rot of wheat in eastern China. Plant Pathol. 2020, 69, 240–248. [Google Scholar] [CrossRef]
- Cui, L.; Selvaraj, J.N.; Xing, F.; Zhao, Y.; Zhou, L.; Liu, Y. A minor survey of deoxynivalenol in Fusarium infected wheat from Yangtze–Huaihe river basin region in China. Food Control 2013, 30, 469–473. [Google Scholar] [CrossRef]
- Han, Z.; Nie, D.; Ediage, E.N.; Yang, X.; Wang, J.; Chen, B.; Li, S.; On, S.L.; De Saeger, S.; Wu, A. Cumulative health risk assessment of co-occurring mycotoxins of deoxynivalenol and its acetyl derivatives in wheat and maize: Case study, Shanghai, China. Food Chem. Toxicol. 2014, 74, 334–342. [Google Scholar] [CrossRef]
- Lahouar, A.; Marin, S.; Crespo-Sempere, A.; Saïd, S.; Sanchis, V. Influence of temperature, water activity and incubation time on fungal growth and production of ochratoxin A and zearalenone by toxigenic Aspergillus tubingensis and Fusarium incarnatum isolates in sorghum seeds. Int. J. Food Microbiol. 2017, 242, 53–60. [Google Scholar] [CrossRef]
- Khan, R.; Ghazali, F.M.; Mahyudin, N.A.; Samsudin, N.I. Biocontrol of Aflatoxins Using Non-Aflatoxigenic Aspergillus flavus: A Literature Review. J. Fungi 2021, 7, 381. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Wang, S.; Hu, X.; Su, J.; Huang, T.; Yu, J.; Tang, L.; Gao, W.; Wang, J.-S. Fumonisin B1 contamination of home-grown corn in high-risk areas for esophageal and liver cancer in China. Food Addit. Contam. 2007, 24, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Wang, S.; Yu, M.; Sun, Y.; Xu, J.; Shi, J. Natural occurrence of deoxynivalenol and deoxynivalenol-3-glucoside in various wheat cultivars grown in Jiangsu province, China. World Mycotoxin J. 2017, 10, 285–293. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, Y.; Wang, L.; Chang, F.; Yang, L. Occurrence of deoxynivalenol in wheat, Hebei Province, China. Food Chem. 2016, 197, 1271–1274. [Google Scholar] [CrossRef]
- Ji, F.; He, D.; Olaniran, A.O.; Mokoena, M.P.; Xu, J.; Shi, J. Occurrence, toxicity, production and detection of Fusarium mycotoxin: A review. Food Prod. Process. Nutr. 2019, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.D.; Su, P.; Shan, H. Mycotoxin Contamination of Maize in China. Compr. Rev. Food Sci. Food Saf. 2017, 16, 835–849. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Liu, X.M. Contamination of aflatoxins in different kinds of foods in China. Biomed. Environ. Sci. 2007, 20, 483–487. [Google Scholar]
- Zhu, F.; Wang, L.; Lin, Y. Investigation of mycotoxin contamination on common sheep feed in Shandong province. Chin. J. Anim. Sci. 2014, 10, 72–76. [Google Scholar]
- Lai, X.; Liu, R.; Ruan, C.; Zhang, H.; Liu, C. Occurrence of aflatoxins and ochratoxin A in rice samples from six provinces in China. Food Control 2015, 50, 401–404. [Google Scholar] [CrossRef]
- Shi, H.; Li, S.; Bai, Y.; Prates, L.L.; Lei, Y.; Yu, P. Mycotoxin contamination of food and feed in China: Occurrence, detection techniques, toxicological effects and advances in mitigation technologies. Food Control 2018, 91, 202–215. [Google Scholar] [CrossRef]
- Zhao, K.; Shao, B.; Yang, D.; Li, F.; Zhu, J. Natural Occurrence of Alternaria Toxins in Wheat-Based Products and Their Dietary Exposure in China. PLoS ONE 2015, 10, e0132019. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.; Jiang, K.; Fan, Z.; Diana Di Mavungu, J.; Dong, M.; Guo, W.; Fan, K.; Campbell, K.; Zhao, Z.; Wu, Y. Multi-walled carbon nanotubes-based magnetic solid-phase extraction for the determination of zearalenone and its derivatives in maize by ultra-high performance liquid chromatography-tandem mass spectrometry. Food Control 2017, 79, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Xing, F.; Liu, X.; Wang, L.; Selvaraj, J.N.; Jin, N.; Wang, Y.; Zhao, Y.; Liu, Y. Distribution and variation of fungi and major mycotoxins in pre- and post-nature drying maize in North China Plain. Food Control 2017, 80, 244–251. [Google Scholar] [CrossRef]
- Temba, M.C.; Njobeh, P.B.; Kayitesi, E. Storage stability of maize-groundnut composite flours and an assessment of aflatoxin B1 and ochratoxin A contamination in flours and porridges. Food Control 2017, 71, 178–186. [Google Scholar] [CrossRef]
- Albuzaudi, M.; Eerikäinen, T.; Turunen, O.; Ghelawi, M.; El Haj Assad, M.; Tawalbeh, M.; Bedade, D.; Shamekh, S. Effect of gamma irradiation and heat treatment on the artificial contamination of maize grains by Aspergillus flavus Link NRRL 5906. J. Stored Prod. Res. 2017, 71, 57–63. [Google Scholar] [CrossRef]
- Basappa, S.; Shantha, T. Methods for detoxification of aflatoxins in foods and feeds-a critical appraisal. J. Food Sci. Technol. 1996, 33, 95–107. [Google Scholar]
- Bata, Á.; Lásztity, R. Detoxification of mycotoxin-contaminated food and feed by microorganisms. Trends Food Sci. Technol. 1999, 10, 223–228. [Google Scholar] [CrossRef]
- Womack, E.D.; Brown, A.E.; Sparks, D.L. A recent review of non-biological remediation of aflatoxin-contaminated crops. J. Sci. Food Agric. 2014, 94, 1706–1714. [Google Scholar] [CrossRef]
- Shanakhat, H.; Sorrentino, A.; Raiola, A.; Romano, A.; Masi, P.; Cavella, S. Current methods for mycotoxins analysis and innovative strategies for their reduction in cereals: An overview. J. Sci. Food Agric. 2018, 98, 4003–4013. [Google Scholar] [CrossRef] [PubMed]
- Cazzaniga, D.; Basílico, J.C.; González, R.J.; Torres, R.L.; De Greef, D.M. Mycotoxins inactivation by extrusion cooking of corn flour. Lett. Appl. Microbiol. 2001, 33, 144–147. [Google Scholar] [CrossRef]
- Samarajeewa, U.; Sen, A.C.; Cohen, M.D.; Wei, C.I. Detoxification of Aflatoxins in Foods and Feeds by Physical and Chemical Methods 1. J. Food Prot. 1990, 53, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Mir, S.A.; Dar, B.N.; Shah, M.A.; Sofi, S.A.; Hamdani, A.M.; Oliveira, C.A.F.; Hashemi Moosavi, M.; Mousavi Khaneghah, A.; Sant’Ana, A.S. Application of new technologies in decontamination of mycotoxins in cereal grains: Challenges, and perspectives. Food Chem. Toxicol. 2021, 148, 111976. [Google Scholar] [CrossRef]
- Gavahian, M.; Cullen, P.J. Cold Plasma as an Emerging Technique for Mycotoxin-Free Food: Efficacy, Mechanisms, and Trends. Food Rev. Int. 2020, 36, 193–214. [Google Scholar] [CrossRef]
- Conte, G.; Fontanelli, M.; Galli, F.; Cotrozzi, L.; Pagni, L.; Pellegrini, E. Mycotoxins in Feed and Food and the Role of Ozone in Their Detoxification and Degradation: An Update. Toxins 2020, 12, 486. [Google Scholar] [CrossRef] [PubMed]
- Freitas-Silva, O.; Venâncio, A. Ozone applications to prevent and degrade mycotoxins: A review. Drug Metab. Rev. 2010, 42, 612–620. [Google Scholar] [CrossRef]
- Sun, X.D.; Su, P.; Shan, H. Mycotoxin Contamination of Rice in China. J. Food Sci. 2017, 82, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Dorner, J.W.; Cole, R.J.; Blankenship, P.D. Use of a Biocompetitive Agent to Control Preharvest Aflatoxin in Drought Stressed Peanuts. J. Food Prot. 1992, 55, 888–892. [Google Scholar] [CrossRef]
- Stanley, V.G.; Ojo, R.; Woldesenbet, S.; Hutchinson, D.H.; Kubena, L.F. The use of Saccharomyces cerevisiae to suppress the effects of aflatoxicosis in broiler chicks. Poult. Sci. 1993, 72, 1867–1872. [Google Scholar] [CrossRef]
- Dvegowda, G.; Raju, M.V.L.N.; Swamy, H.D.N. Mycotoxins: Novel solutions for their counteraction. Feedstuffs 1998, 70, 12–17. [Google Scholar]
- Armando, M.; Dogi, C.; Pizzolitto, R.; Escobar, F.; Peirano, M.; Salvano, M.; Sabini, L.; Combina, M.; Dalcero, A.; Cavaglieri, L. Saccharomyces cerevisiae strains from animal environment with in vitro aflatoxin B1 binding ability and anti-pathogenic bacterial influence. World Mycotoxin J. 2011, 4, 59–68. [Google Scholar] [CrossRef]
- Raju, M.V.L.N.; Devegowda, G. Esterified-Glucomannan in Broiler Chicken Diets-Contaminated with Aflatoxin, Ochratoxin and T-2 Toxin: Evaluation of its Binding Ability (in vitro) and Efficacy as Immunomodulator. Asian-Australas. J. Anim. Sci. 2002, 15, 1051–1056. [Google Scholar] [CrossRef]
- Yiannikouris, A.; André, G.; Poughon, L.; François, J.; Dussap, C.-G.; Jeminet, G.; Bertin, G.; Jouany, J.-P. Chemical and Conformational Study of the Interactions Involved in Mycotoxin Complexation with β-d-Glucans. Biomacromolecules 2006, 7, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Freimund, S.; Sauter, M.; Rys, P. Efficient Adsorption of the Mycotoxins Zearalenone and T-2 Toxin on a Modified Yeast Glucan. J. Environ. Sci. Health Part B 2003, 38, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Mani-López, E.; Arrioja-Bretón, D.; López-Malo, A. The impacts of antimicrobial and antifungal activity of cell-free supernatants from lactic acid bacteria in vitro and foods. Compr. Rev. Food Sci. Food Saf. 2022, 21, 604–641. [Google Scholar] [CrossRef] [PubMed]
- Haskard Carolyn, A.; El-Nezami Hani, S.; Kankaanpää Pasi, E.; Salminen, S.; Ahokas Jorma, T. Surface Binding of Aflatoxin B1 by Lactic Acid Bacteria. Appl. Environ. Microbiol. 2001, 67, 3086–3091. [Google Scholar] [CrossRef] [Green Version]
- El-Nezami, H.; Polychronaki, N.; Salminen, S.; Mykkänen, H. Binding Rather Than Metabolism May Explain the Interaction of Two Food-Grade Lactobacillus Strains with Zearalenone and Its Derivative ά-Zearalenol. Appl. Environ. Microbiol. 2002, 68, 3545–3549. [Google Scholar] [CrossRef] [Green Version]
- Niderkorn, V.; Boudra, H.; Morgavi, D.P. Binding of Fusarium mycotoxins by fermentative bacteria in vitro. J. Appl. Microbiol. 2006, 101, 849–856. [Google Scholar] [CrossRef]
- Lahtinen, S.J.; Haskard, C.A.; Ouwehand, A.C.; Salminen, S.J.; Ahokas, J.T. Binding of aflatoxin B1 to cell wall components of Lactobacillus rhamnosus strain GG. Food Addit. Contam. 2004, 21, 158–164. [Google Scholar] [CrossRef]
- Hernandez-Mendoza, A.; Guzman-de-Peña, D.; Garcia, H.S. Key role of teichoic acids on aflatoxin B1 binding by probiotic bacteria. J. Appl. Microbiol. 2009, 107, 395–403. [Google Scholar] [CrossRef]
- Bueno, D.J.; Casale, C.H.; Pizzolitto, R.P.; Salvano, M.A.; Oliver, G. Physical Adsorption of Aflatoxin B1 by Lactic Acid Bacteria and Saccharomyces cerevisiae: A Theoretical Model. J. Food Prot. 2007, 70, 2148–2154. [Google Scholar] [CrossRef]
- Oluwafemi, F.; Silva, F.A.d. Removal of aflatoxins by viable and heat-killed Lactobacillus species isolated from fermented maize. J. Appl. Biosci. 2009, 16, 871–876. [Google Scholar]
- Cleveland, T.E.; Dowd, P.F.; Desjardins, A.E.; Bhatnagar, D.; Cotty, P.J. United States Department of Agriculture—Agricultural Research Service research on pre-harvest prevention of mycotoxins and mycotoxigenic fungi in US crops. Pest Manag. Sci. 2003, 59, 629–642. [Google Scholar]
- Cotty, P. Influence of field application of an atoxigenic strain of Aspergillus flavus on the populations of A. flavus infecting cotton bolls and on the aflatoxin content of cottonseed. Phytopathology 1994, 84, 1270–1277. [Google Scholar] [CrossRef]
- Abbas, H.K.; Zablotowicz, R.M.; Bruns, H.A.; Abel, C.A. Biocontrol of aflatoxin in corn by inoculation with non-aflatoxigenic Aspergillus flavus isolates. Biocontrol Sci. Technol. 2006, 16, 437–449. [Google Scholar] [CrossRef]
- Probst, C.; Bandyopadhyay, R.; Price, L.E.; Cotty, P.J. Identification of Atoxigenic Aspergillus flavus Isolates to Reduce Aflatoxin Contamination of Maize in Kenya. Plant Dis. 2010, 95, 212–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorner, J.W.; Cole, R.J.; Connick, W.J.; Daigle, D.J.; McGuire, M.R.; Shasha, B.S. Evaluation of biological control formulations to reduce aflatoxin contamination in peanuts. Biol. Control 2003, 26, 318–324. [Google Scholar] [CrossRef]
- Khan, N.I.; Schisler, D.A.; Boehm, M.J.; Lipps, P.E.; Slininger, P.J. Field testing of antagonists of Fusarium head blight incited by Gibberella zeae. Biol. Control 2004, 29, 245–255. [Google Scholar] [CrossRef]
- Leite de Souza, E. Insights into the current evidence on the effects of essential oils toward beneficial microorganisms in foods with a special emphasis to lactic acid bacteria—A review. Trends Food Sci. Technol. 2021, 114, 333–341. [Google Scholar] [CrossRef]
- Farbo, M.G.; Urgeghe, P.P.; Fiori, S.; Marcello, A.; Oggiano, S.; Balmas, V.; Hassan, Z.U.; Jaoua, S.; Migheli, Q. Effect of yeast volatile organic compounds on ochratoxin A-producing Aspergillus carbonarius and A. ochraceus. Int. J. Food Microbiol. 2018, 284, 1–10. [Google Scholar] [CrossRef]
- Guimarães, A.; Santiago, A.; Teixeira, J.A.; Venâncio, A.; Abrunhosa, L. Anti-aflatoxigenic effect of organic acids produced by Lactobacillus plantarum. Int. J. Food Microbiol. 2018, 264, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Lavermicocca, P.; Valerio, F.; Evidente, A.; Lazzaroni, S.; Corsetti, A.; Gobbetti, M. Purification and Characterization of Novel Antifungal Compounds from the Sourdough Lactobacillus plantarum Strain 21B. Appl. Environ. Microbiol. 2000, 66, 4084–4090. [Google Scholar] [CrossRef] [Green Version]
- Muhialdin, B.J.; Hassan, Z.; Bakar, F.A.; Saari, N. Identification of antifungal peptides produced by Lactobacillus plantarum IS10 grown in the MRS broth. Food Control 2016, 59, 27–30. [Google Scholar] [CrossRef]
- Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; Said, L.B.; Gaudreau, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a new generation of antimicrobials: Toxicity aspects and regulations. FEMS Microbiol. Rev. 2021, 45, fuaa039. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Lu, L.; Pang, J.; Hu, Y.; Guo, Q.; Li, Z.; Wu, S.; Liu, H.; Wang, C. Biocontrol activity of volatile organic compounds from Streptomyces alboflavus TD-1 against Aspergillus flavus growth and aflatoxin production. J. Microbiol. 2019, 57, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Sangmanee, P.; Hongpattarakere, T. Inhibitory of multiple antifungal components produced by Lactobacillus plantarum K35 on growth, aflatoxin production and ultrastructure alterations of Aspergillus flavus and Aspergillus parasiticus. Food Control 2014, 40, 224–233. [Google Scholar] [CrossRef]
- Munimbazi, C.; Bullerman, L.B. Inhibition of aflatoxin production of Aspergillus parasiticus NRRL 2999 by Bacillus pumilus. Mycopathologia 1997, 140, 163–169. [Google Scholar] [CrossRef]
- Munimbazi, C.; Bullerman, L.B. Isolation and partial characterization of antifungal metabolites of Bacillus pumilus. J. Appl. Microbiol. 1998, 84, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Sakuda, S.; Suzuki, A.; Isogai, A. Aflastatin A, a novel inhibitor of aflatoxin production by aflatoxigenic fungi. J. Antibiot. 1997, 50, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, A.-D.; Dong, F.-Y.; Hu, M.-J.; Kong, X.-W.; Wei, F.-F.; Gong, S.-J.; Zhang, Y.-M.; Zhang, J.-B.; Wu, A.-B.; Liao, Y.-C. Antifungal activity of volatile emitted from Enterobacter asburiae Vt-7 against Aspergillus flavus and aflatoxins in peanuts during storage. Food Control 2019, 106, 106718. [Google Scholar] [CrossRef]
- De Simone, N.; Capozzi, V.; de Chiara, M.L.; Amodio, M.L.; Brahimi, S.; Colelli, G.; Drider, D.; Spano, G.; Russo, P. Screening of Lactic Acid Bacteria for the Bio-Control of Botrytis cinerea and the Potential of Lactiplantibacillus plantarum for Eco-Friendly Preservation of Fresh-Cut Kiwifruit. Microorganisms 2021, 9, 773. [Google Scholar] [CrossRef]
- Guo, Y.; Qin, X.; Tang, Y.; Ma, Q.; Zhang, J.; Zhao, L. CotA laccase, a novel aflatoxin oxidase from Bacillus licheniformis, transforms aflatoxin B1 to aflatoxin Q1 and epi-aflatoxin Q1. Food Chem. 2020, 325, 126877. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, J.; Jin, Y.; Wu, C.; Shen, D.; Zhang, S.; Zhou, R. Aflatoxin B1 degradation by salt tolerant Tetragenococcus halophilus CGMCC 3792. Food Chem. Toxicol. 2018, 121, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, J.; Jin, Y.; Wu, C.; Shen, D.; Zhang, S.; Zhou, R. Mechanism and kinetics of degrading aflatoxin B1 by salt tolerant Candida versatilis CGMCC 3790. J. Hazard. Mater. 2018, 359, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Stander, M.A.; Bornscheuer, U.T.; Henke, E.; Steyn, P.S. Screening of Commercial Hydrolases for the Degradation of Ochratoxin A. J. Agric. Food Chem. 2000, 48, 5736–5739. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Yu, M.; Dong, F.; Shi, J.; Xu, J. Esterase activity inspired selection and characterization of zearalenone degrading bacteria Bacillus pumilus ES-21. Food Control 2017, 77, 57–64. [Google Scholar] [CrossRef]
- Wang, J.; Ogata, M.; Hirai, H.; Kawagishi, H. Detoxification of aflatoxin B1 by manganese peroxidase from the white-rot fungus Phanerochaete sordida YK-624. FEMS Microbiol. Lett. 2011, 314, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, J.; Liu, Z.; Shi, Y.; Liu, J.; Xu, X.; Hao, S.; Mu, P.; Deng, F.; Deng, Y. Aflatoxin B1 Degradation and Detoxification by Escherichia coli CG1061 Isolated From Chicken Cecum. Front. Pharmacol. 2019, 9, 130. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, Y.; Gong, A.; Liu, N.; Chen, S.; Zhao, X.; Li, X.; Chen, L.; Zhou, C.; Wang, J. Biodegradation of mycotoxin fumonisin B1 by a novel bacterial consortium SAAS79. Appl. Microbiol. Biotechnol. 2019, 103, 7129–7140. [Google Scholar] [CrossRef]
- Zhou, W.-W.; Tang, X.; Zheng, X.; Zhang, Y.; Liu, S. Screening and Application of Bacillus Strains to Degrade Aflatoxin B1. Patent No. CN108102963A, 1 June 2018. [Google Scholar]
- Takahashi-Ando, N.; Kimura, M.; Kakeya, H.; Osada, H.; Yamaguchi, I. A novel lactonohydrolase responsible for the detoxification of zearalenone: Enzyme purification and gene cloning. Biochem. J. 2002, 365, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Yeong-Hsiang, C.; Ching-Feng, W.; Bao-Ji, C.; Ming-Huang, C. Toxicity of different Fusarium mycotoxins on growth performance, immune responses and efficacy of a mycotoxin degrading enzyme in pigs. Anim. Res. 2006, 55, 579–590. [Google Scholar]
- Utermark, J.; Karlovsky, P. Role of Zearalenone Lactonase in Protection of Gliocladium roseum from Fungitoxic Effects of the Mycotoxin Zearalenone. Appl. Environ. Microbiol. 2007, 73, 637–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altalhi, A.D.; El-Deeb, B. Localization of zearalenone detoxification gene(s) in pZEA-1 plasmid of Pseudomonas putida ZEA-1 and expressed in Escherichia coli. J. Hazard. Mater. 2009, 161, 1166–1172. [Google Scholar] [CrossRef] [PubMed]
- Teniola, O.D.; Addo, P.A.; Brost, I.M.; Färber, P.; Jany, K.D.; Alberts, J.F.; van Zyl, W.H.; Steyn, P.S.; Holzapfel, W.H. Degradation of aflatoxin B1 by cell-free extracts of Rhodococcus erythropolis and Mycobacterium fluoranthenivorans sp. nov. DSM44556T. Int. J. Food Microbiol. 2005, 105, 111–117. [Google Scholar] [CrossRef]
- Tinyiro, S.E.; Wokadala, C.; Xu, D.; Yao, W. Adsorption and degradation of zearalenone by bacillus strains. Folia Microbiol. 2011, 56, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Eisa Ahmed, M.F.; Sangare, L.; Zhao, Y.; Selvaraj, J.N.; Xing, F.; Wang, Y.; Yang, H.; Liu, Y. Novel Aflatoxin-Degrading Enzyme from Bacillus shackletonii L7. Toxins 2017, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Farzaneh, M.; Shi, Z.-Q.; Ghassempour, A.; Sedaghat, N.; Ahmadzadeh, M.; Mirabolfathy, M.; Javan-Nikkhah, M. Aflatoxin B1 degradation by Bacillus subtilis UTBSP1 isolated from pistachio nuts of Iran. Food Control 2012, 23, 100–106. [Google Scholar] [CrossRef]
- Zhou, W.-W.; Wei, X.; Wang, S.; Qian, C. High-Efficiency Fermentation Method of Aflatoxin-Degrading Bacteria. Patent No. CN111154679A, 15 May 2020. [Google Scholar]
- Wang, Y.; Zhang, H.; Yan, H.; Yin, C.; Liu, Y.; Xu, Q.; Liu, X.; Zhang, Z. Effective Biodegradation of Aflatoxin B1 Using the Bacillus licheniformis (BL010) Strain. Toxins 2018, 10, 497. [Google Scholar] [CrossRef] [Green Version]
- Shu, X.; Wang, Y.; Zhou, Q.; Li, M.; Hu, H.; Ma, Y.; Chen, X.; Ni, J.; Zhao, W.; Huang, S.; et al. Biological Degradation of Aflatoxin B1 by Cell-Free Extracts of Bacillus velezensis DY3108 with Broad PH Stability and Excellent Thermostability. Toxins 2018, 10, 330. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Sun, J.; Cui, Y.; Wang, X.; Sang, Y. Biological degradation of aflatoxin M1 by Bacillus pumilus E-1-1-1. MicrobiologyOpen 2019, 8, e00663. [Google Scholar] [CrossRef] [Green Version]
- Shao, S.; Cai, J.; Du, X.; Wang, C.; Lin, J.; Dai, J. Biotransformation and detoxification of aflatoxin B1 by extracellular extract of Cladosporium uredinicola. Food Sci. Biotechnol. 2016, 25, 1789–1794. [Google Scholar] [CrossRef]
- Sangare, L.; Zhao, Y.; Folly, Y.M.; Chang, J.; Li, J.; Selvaraj, J.N.; Xing, F.; Zhou, L.; Wang, Y.; Liu, Y. Aflatoxin B1 Degradation by a Pseudomonas Strain. Toxins 2014, 6, 3028–3040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, J.; Mehta, A. Protein-mediated degradation of aflatoxin B1 by Pseudomonas putida. Braz. J. Microbiol. 2019, 50, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-H.; Li, W.-Y.; Hsu, I.N.; Liao, Y.-Y.; Yang, C.-Y.; Taylor, M.C.; Liu, Y.-F.; Huang, W.-H.; Chang, H.-H.; Huang, H.-L.; et al. Recombinant Aflatoxin-Degrading F420H2-Dependent Reductase from Mycobacterium smegmatis Protects Mammalian Cells from Aflatoxin Toxicity. Toxins 2019, 11, 259. [Google Scholar] [CrossRef] [Green Version]
- Elaasser, M.; El Kassas, R. Detoxification of aflatoxin B1 by certain bacterial species isolated from Egyptian soil. World Mycotoxin J. 2011, 4, 169–176. [Google Scholar] [CrossRef]
- Sokoutifar, R.; Razavilar, V.; Anvar, A.A.; Shoeiby, S. Degraded aflatoxin M1 in artificially contaminated fermented milk using Lactobacillus acidophilus and Lactobacillus plantarum affected by some bio-physical factors. J. Food Saf. 2018, 38, e12544. [Google Scholar] [CrossRef]
- Smiley, R.D.; Draughon, F.A. Preliminary evidence that degradation of aflatoxin B1 by Flavobacterium aurantiacum is enzymatic. J. Food Prot. 2000, 63, 415–418. [Google Scholar] [CrossRef]
- Guan, S.; Ji, C.; Zhou, T.; Li, J.; Ma, Q.; Niu, T. Aflatoxin B1 Degradation by Stenotrophomonas Maltophilia and Other Microbes Selected Using Coumarin Medium. Int. J. Mol. Sci. 2008, 9, 1489–1503. [Google Scholar] [CrossRef] [Green Version]
- Yi, P.-J.; Pai, C.-K.; Liu, J.-R. Isolation and characterization of a Bacillus licheniformis strain capable of degrading zearalenone. World J. Microbiol. Biotechnol. 2011, 27, 1035–1043. [Google Scholar] [CrossRef]
- He, X.; Li, S.; Li, Y.; Gu, W.; Sun, Y.; Sun, X. Evaluation of reduced toxicity of zearalenone as measured by the Hep G2 cell assay on degradation enzymes. Food Control 2015, 57, 161–168. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Venâncio, A. Isolation and purification of an enzyme hydrolyzing ochratoxin A from Aspergillus niger. Biotechnol. Lett. 2007, 29, 1909–1914. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yang, H.; Apaliya, M.T.; Zhao, L.; Gu, X.; Zheng, X.; Hu, W.; Zhang, H. The mechanisms involved in ochratoxin A elimination by Yarrowia lipolytica Y-2. Ann. Appl. Biol. 2018, 173, 164–174. [Google Scholar] [CrossRef]
- Liuzzi, V.C.; Fanelli, F.; Tristezza, M.; Haidukowski, M.; Picardi, E.; Manzari, C.; Lionetti, C.; Grieco, F.; Logrieco, A.F.; Thon, M.R.; et al. Transcriptional Analysis of Acinetobacter sp. neg1 Capable of Degrading Ochratoxin A. Front. Microbiol. 2017, 7, 2162. [Google Scholar]
- Chang, X.; Wu, Z.; Wu, S.; Dai, Y.; Sun, C. Degradation of ochratoxin A by Bacillus amyloliquefaciens ASAG1. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 564–571. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, X.Y.; Xu, S.H.; Yuan, G.Q.; Shi, X.J.; Liang, Z.H. Degradation of ochratoxin A by supernatant and ochratoxinase of Aspergillus niger W-35 isolated from cereals. World Mycotoxin J. 2020, 13, 287–298. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Y.; Yin, T.; Wang, J.; Zhang, X. Heterologous Expression and Characterization of A Novel Ochratoxin A Degrading Enzyme, N-acyl-L-amino Acid Amidohydrolase, from Alcaligenes faecalis. Toxins 2019, 11, 518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinl, S.; Hartinger, D.; Thamhesl, M.; Vekiru, E.; Krska, R.; Schatzmayr, G.; Moll, W.D.; Grabherr, R. Degradation of fumonisin B1 by the consecutive action of two bacterial enzymes. J. Biotechnol. 2010, 145, 120–129. [Google Scholar] [CrossRef]
- Heinl, S.; Hartinger, D.; Thamhesl, M.; Schatzmayr, G.; Moll, W.D.; Grabherr, R. An aminotransferase from bacterium ATCC 55552 deaminates hydrolyzed fumonisin B1. Biodegradation 2011, 22, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.-K.; Matsushima, K.; Takahashi, T.; Yu, J.; Abe, K.; Bhatnagar, D.; Yuan, G.-F.; Koyama, Y.; Cleveland, T.E. Understanding nonaflatoxigenicity of Aspergillus sojae: A windfall of aflatoxin biosynthesis research. Appl. Microbiol. Biotechnol. 2007, 76, 977–984. [Google Scholar] [CrossRef]
- Pfliegler, W.P.; Pusztahelyi, T.; Pócsi, I. Mycotoxins—Prevention and decontamination by yeasts. J. Basic Microbiol. 2015, 55, 805–818. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Dou, K.; Lu, Z.; Wang, X.; Li, Y.; Chen, J. Enhanced biocontrol activity of cellulase from Trichoderma harzianum against Fusarium graminearum through activation of defense-related genes in maize. Physiol. Mol. Plant Pathol. 2018, 103, 130–136. [Google Scholar] [CrossRef]
- Ece, S.; Lambertz, C.; Fischer, R.; Commandeur, U. Heterologous expression of a Streptomyces cyaneus laccase for biomass modification applications. AMB Express 2017, 7, 86. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Wu, H.; Tang, Y.; Qiu, L. Cloning, expression of a peroxiredoxin gene from Acinetobacter sp. SM04 and characterization of its recombinant protein for zearalenone detoxification. Microbiol. Res. 2012, 167, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Chang, J.; Wang, P.; Liu, C.; Yin, Q.; Zhu, Q.; Lu, F.; Gao, T. Effect of the combined compound probiotics with mycotoxin-degradation enzyme on detoxifying aflatoxin B1 and zearalenone. J. Toxicol. Sci. 2018, 43, 377–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, R.Y.; Chang, J.; Yin, Q.Q.; Wang, P.; Yang, Y.R.; Wang, X.; Wang, G.Q.; Zheng, Q.H. Effect of the combined probiotics with aflatoxin B₁-degrading enzyme on aflatoxin detoxification, broiler production performance and hepatic enzyme gene expression. Food Chem. Toxicol. 2013, 59, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.S.; Yu, D.; Liu, N.; Wu, A. Degrading Ochratoxin A and Zearalenone Mycotoxins Using a Multifunctional Recombinant Enzyme. Toxins 2019, 11, 301. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Qin, X.; Hao, Z.; Luo, H.; Yao, B.; Su, X. Degradation of Four Major Mycotoxins by Eight Manganese Peroxidases in Presence of a Dicarboxylic Acid. Toxins 2019, 11, 566. [Google Scholar] [CrossRef] [Green Version]
- Armando, M.R.; Pizzolitto, R.P.; Dogi, C.A.; Cristofolini, A.; Merkis, C.; Poloni, V.; Dalcero, A.M.; Cavaglieri, L.R. Adsorption of ochratoxin A and zearalenone by potential probiotic Saccharomyces cerevisiae strains and its relation with cell wall thickness: Ochratoxin A and zearalenone adsorption by yeasts. J. Appl. Microbiol. 2012, 113, 256–264. [Google Scholar] [CrossRef]
- Guo, C.; Yuan, Y.; Yue, T.; Hatab, S.; Wang, Z. Binding mechanism of patulin to heat-treated yeast cell. Lett. Appl. Microbiol. 2012, 55, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wang, J.; Liu, B.; Wang, Z.; Yuan, Y.; Yue, T. Effect of Yeast Cell Morphology, Cell Wall Physical Structure and Chemical Composition on Patulin Adsorption. PLoS ONE 2015, 10, e0136045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Liu, X.; Yuan, L.; Li, J. Complicated interactions between bio-adsorbents and mycotoxins during mycotoxin adsorption: Current research and future prospects. Trends Food Sci. Technol. 2020, 96, 127–134. [Google Scholar] [CrossRef]
- Haskard, C.; Binnion, C.; Ahokas, J. Factors affecting the sequestration of aflatoxin by Lactobacillusrhamnosus strain GG. Chem. Biol. Interact. 2000, 128, 39–49. [Google Scholar] [CrossRef]
- Peltonen, K.; El-Nezami, H.; Haskard, C.; Ahokas, J.; Salminen, S. Aflatoxin B1 Binding by Dairy Strains of Lactic Acid Bacteria and Bifidobacteria. J. Dairy Sci. 2001, 84, 2152–2156. [Google Scholar] [CrossRef]
- Takahashi-Ando, N.; Ohsato, S.; Shibata, T.; Hamamoto, H.; Yamaguchi, I.; Kimura, M. Metabolism of Zearalenone by Genetically Modified Organisms Expressing the Detoxification Gene from Clonostachys rosea. Appl. Environ. Microbiol. 2004, 70, 3239–3245. [Google Scholar] [PubMed] [Green Version]
- Xu, W.; Han, X.; Li, F.; Zhang, L. Natural Occurrence of Alternaria Toxins in the 2015 Wheat from Anhui Province, China. Toxins 2016, 8, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, M.; Feng, Y.; He, X.; Zhang, D.; Wang, W.; Liu, D. Mycotoxins in livestock feed in China—Current status and future challenges. Toxicon 2022, 214, 112–120. [Google Scholar] [CrossRef]
- Abdi, M.; Asadi, A.; Maleki, F.; Kouhsari, E.; Fattahi, A.; Ohadi, E.; Lotfali, E.; Ahmadi, A.; Ghafouri, Z. Microbiological Detoxification of Mycotoxins: Focus on Mechanisms and Advances. Infect. Disord. Drug Targets 2021, 21, 339–357. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Wang, L.; Sun, J.; Wang, L.; Guo, H.; Ye, Y.; Sun, X. Microbial detoxification of mycotoxins in food and feed. Crit. Rev. Food Sci. Nutr. 2022, 62, 4951–4969. [Google Scholar] [CrossRef]
- Siahmoshteh, F.; Hamidi-Esfahani, Z.; Spadaro, D.; Shams-Ghahfarokhi, M.; Razzaghi-Abyaneh, M. Unraveling the mode of antifungal action of Bacillus subtilis and Bacillus amyloliquefaciens as potential biocontrol agents against aflatoxigenic Aspergillus parasiticus. Food Control 2018, 89, 300–307. [Google Scholar] [CrossRef]
- Chen, Y.; Kong, Q.; Liang, Y. Three newly identified peptides from Bacillus megaterium strongly inhibit the growth and aflatoxin B1 production of Aspergillus flavus. Food Control 2019, 95, 41–49. [Google Scholar] [CrossRef]
- Scillato, M.; Spitale, A.; Mongelli, G.; Privitera, G.F.; Mangano, K.; Cianci, A.; Stefani, S.; Santagati, M. Antimicrobial properties of Lactobacillus cell-free supernatants against multidrug-resistant urogenital pathogens. MicrobiologyOpen 2021, 10, e1173. [Google Scholar] [CrossRef]
- Yang, E.J.; Chang, H.C. Purification of a new antifungal compound produced by Lactobacillus plantarum AF1 isolated from kimchi. Int. J. Food Microbiol. 2010, 139, 56–63. [Google Scholar] [CrossRef]
- Abdel-Kareem, M.M.; Rasmey, A.M.; Zohri, A.A. The action mechanism and biocontrol potentiality of novel isolates of Saccharomyces cerevisiae against the aflatoxigenic Aspergillus flavus. Lett. Appl. Microbiol. 2019, 68, 104–111. [Google Scholar] [CrossRef]
- Hua, S.S.T.; Beck, J.J.; Sarreal, S.B.L.; Gee, W. The major volatile compound 2-phenylethanol from the biocontrol yeast, Pichia anomala, inhibits growth and expression of aflatoxin biosynthetic genes of Aspergillus flavus. Mycotoxin Res. 2014, 30, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.M.d.; Prietto, L.; Ribeiro, A.C.; Souza, T.D.d.; Badiale-Furlong, E. Assessment of the antifungal activity of Spirulina platensis phenolic extract against Aspergillus flavus. Ciênc. Agrotecnol. 2011, 35, 1050–1058. [Google Scholar]
- Tayel, A.A.; El-Tras, W.F.; Moussa, S.H.; El-Agamy, M.A. Antifungal action of Pichia anomala against aflatoxigenic Aspergillus flavus and its application as a feed supplement. J. Sci. Food Agric. 2013, 93, 3259–3263. [Google Scholar] [CrossRef]
- Yoshinari, T.; Akiyama, T.; Nakamura, K.; Kondo, T.; Takahashi, Y.; Muraoka, Y.; Nonomura, Y.; Nagasawa, H.; Sakuda, S. Dioctatin A is a strong inhibitor of aflatoxin production by Aspergillus parasiticus. Microbiology 2007, 153, 2774–2780. [Google Scholar] [CrossRef] [Green Version]
- Shakeel, Q.; Lyu, A.; Zhang, J.; Wu, M.; Li, G.; Hsiang, T.; Yang, L. Biocontrol of Aspergillus flavus on Peanut Kernels Using Streptomyces yanglinensis 3-10. Front. Microbiol. 2018, 9, 1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, J.J.; Huang, W.Q.; Li, Z.W.; Lu, D.L.; Zhang, Y.; Luo, X.C. Biocontrol activity of recombinant aspartic protease from Trichoderma harzianum against pathogenic fungi. Enzyme Microb. Technol. 2018, 112, 35–42. [Google Scholar] [CrossRef]
- Akocak, P.B.; Churey, J.J.; Worobo, R.W. Antagonistic effect of chitinolytic Pseudomonas and Bacillus on growth of fungal hyphae and spores of aflatoxigenic Aspergillus flavus. Food Biosci. 2015, 10, 48–58. [Google Scholar] [CrossRef]
- Jaibangyang, S.; Nasanit, R.; Limtong, S. Biological control of aflatoxin-producing Aspergillus flavus by volatile organic compound-producing antagonistic yeasts. BioControl 2020, 65, 377–386. [Google Scholar] [CrossRef]
- Corsetti, A.; Gobbetti, M.; Rossi, J.; Damiani, P. Antimould activity of sourdough lactic acid bacteria: Identification of a mixture of organic acids produced by Lactobacillus sanfrancisco CB1. Appl. Microbiol. Biotechnol. 1998, 50, 253–256. [Google Scholar] [CrossRef]
- Klich, M.A.; Lax, A.R.; Bland, J.M. Inhibition of some mycotoxigenic fungi by iturin A, a peptidolipid produced by Bacillus subtilis. Mycopathologia 1991, 116, 77–80. [Google Scholar] [CrossRef]
- Medina, Á.; Jiménez, M.; Mateo, R.; Magan, N. Efficacy of natamycin for control of growth and ochratoxin A production by Aspergillus carbonarius strains under different environmental conditions. J. Appl. Microbiol. 2007, 103, 2234–2239. [Google Scholar] [CrossRef] [PubMed]
- Masoud, W.; Poll, L.; Jakobsen, M. Influence of volatile compounds produced by yeasts predominant during processing of Coffea arabica in East Africa on growth and ochratoxin A (OTA) production by Aspergillus ochraceus. Yeast 2005, 22, 1133–1142. [Google Scholar] [CrossRef] [Green Version]
- Ul Hassan, Z.; Al Thani, R.; Alnaimi, H.; Migheli, Q.; Jaoua, S. Investigation and Application of Bacillus licheniformis Volatile Compounds for the Biological Control of Toxigenic Aspergillus and Penicillium spp. ACS Omega 2019, 4, 17186–17193. [Google Scholar] [CrossRef] [PubMed]
- Scaglioni, P.T.; de Oliveira Garcia, S.; Badiale-Furlong, E. Inhibition of in vitro trichothecenes production by microalgae phenolic extracts. Food Res. Int. 2019, 124, 175–180. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, Q.; Wang, K.; Brian, K.; Liu, C.; Gu, Y. Study of the antifungal activity of Bacillus vallismortis ZZ185 in vitro and identification of its antifungal components. Bioresour. Technol. 2010, 101, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Shatha, A.; Shafiq, S. Antagonistic activity of probiotic and sea weed extract against vegetative growth for some fungi and Zearalenone production. World J. Pharm. Res. 2015, 4, 1577–1585. [Google Scholar]
- Mahdi, L.H.; Shafiq, S.A.; Ajaa, H. Effects of crude and purified bacteriocin of Pediococcus pentosaceus on the growth and zearalenone production by Fusarium graminearum. Int. J. Curr Eng. Technol. 2013, 4, 2277–4106. [Google Scholar]
- Lee, T.; Park, D.; Kim, K.; Lim, S.M.; Yu, N.H.; Kim, S.; Kim, H.-Y.; Jung, K.S.; Jang, J.Y.; Park, J.-C.; et al. Characterization of Bacillus amyloliquefaciens DA12 Showing Potent Antifungal Activity against Mycotoxigenic Fusarium Species. Plant Pathol. J. 2017, 33, 499–507. [Google Scholar] [CrossRef] [Green Version]
- Gong, A.-D.; Li, H.-P.; Yuan, Q.-S.; Song, X.-S.; Yao, W.; He, W.-J.; Zhang, J.-B.; Liao, Y.-C. Antagonistic Mechanism of Iturin A and Plipastatin A from Bacillus amyloliquefaciens S76-3 from Wheat Spikes against Fusarium graminearum. PLoS ONE 2015, 10, e0116871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juodeikiene, G.; Bartkiene, E.; Cernauskas, D.; Cizeikiene, D.; Zadeike, D.; Lele, V.; Bartkevics, V. Antifungal activity of lactic acid bacteria and their application for Fusarium mycotoxin reduction in malting wheat grains. LWT 2018, 89, 307–314. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Mycotoxin (Toxic Dose) | Food Commodity | Toxicology | |
|---|---|---|---|
| Human | Animals | ||
| Aflatoxins (AFs) (>2 ppm or 300 μg/kg) | Maize, sorghum, pearl millet, rice, wheat, oilseeds, spices, and tree nuts | 1. Ingestion of medium or high doses of AFs can cause acute liver damage, potentially leading to death. Chronic AF poisoning can lead to cirrhosis and liver cancer [11]. | 1. Increased mortality, reduced hatchability, growth rate, meat and egg yield, and quality in poultry [12]. |
| 2. Neurotransmitter deficiency impairing the central nervous system [13]. | 2. In dairy cows and beef cattle, acute AF poisoning can decrease milk production and cause weight loss and death [14]. | ||
| 3. Immunosuppression [15]. | |||
| Ochratoxin A (OTA) (>100 ng/g) | Rice, barley, maize, wheat, flour, and bran | 1. Balkan endemic nephropathy (BEN) and chronic interstitial nephropathy (CIN) [16]. | 1. Excessive intake of OTA can cause kidney disease in pigs. 2. In poultry, weight loss, decreased egg production and eggshell quality, and renal toxicity [17]. |
| 2. Genotoxic and carcinogenic [18]. | |||
| Fumonisin (>200 μg/g) | Rice, wheat, peanut, barley, maize, rye, oat, and millet | 1. Esophageal cancer, oral cancer, pharyngeal cancer, and fetal neural tube defects [19]. | 1. Pulmonary edema in pigs and leukoencephalomalacia in horses. 2. Toxic to the liver and kidneys of poultry and livestock [20]. |
| 2. Promotes AF-initiated liver tumors [21]. | |||
| Zearalenone (ZEN) (>0.5 μg/kg) | Maize, rice, wheat, and barley | 1. Increased risk of breast cancer [22]. | Miscarriage and infertility in livestock [23]. |
| 2. Excessive ZEN intake during pregnancy can reduce embryo survival [24]. | |||
| 3. Genetic, immune, blood, and liver toxicity [23]. | |||
| Citrinin (>20–40 mg/kg) | Wheat, rye, barley, and beans | 1. Chronic poisoning can cause kidney failure and weight loss. | 1. Slow growth and watery feces in poultry. |
| 2. Embryotoxic, immunotoxic, and teratogenic effects [25]. | 2. Chickens, rabbits, and mice experience varying degrees of liver and kidney damage [26]. | ||
| Deoxynivalenol (DON) (>19.3 ng/g) | Wheat, barley, oats, rye, and maize | Excessive intake of DON can cause nausea, vomiting, diarrhea, abdominal pain, headache, and fever [27]. | 1. Excessive intake of DON can cause acute gastrointestinal diseases and immune dysfunction. |
| 2. Weight loss and anorexia [28]. | |||
| T-2 toxin (>0.05–10 mg/kg) | Wheat, maize, barley, rice, soybeans, and oats | 1. Skin blistering, pain, burning, itching, and inflammation. | 1. Cows were diagnosed with gastroenteritis lesions, intestinal bleeding, decreased milk production, and no estrus cycle. |
| 2. Dyspnea and coughing after inhalation, as well as vomiting, diarrhea, and anorexia. | 2. Poultry were diagnosed with impaired immune systems, damaged hematopoietic systems, and altered feather patterns. | ||
| 3. Genetically toxic and cytotoxic with adverse effects on the immune system [29]. | 3. Pigs were diagnosed with gastric bleeding, intestinal necrosis, and a refusal to feed [29]. | ||
| Detoxification Approach | Specific Method | Pros | Cons | References |
|---|---|---|---|---|
| Physical | Sorting | Reduces aflatoxin contamination level by up to 70–80%. | Inefficient and laborious. | [72] |
| Extrusion | Reduces aflatoxin contamination level by 50–80%. | Causes a loss of nutritional ingredients in the cereals. | ||
| Reduces the toxicity of the initial compounds. | ||||
| Heat treatment | Kills molds attached to the surface of the cereal. | Damages cereals’ nutritional values and sensory quality. | [73] | |
| Destroys some natural toxins by 10–25%. | High energy consumption. Restricted by the heat resistance and moisture of cereals. | |||
| Ultraviolet (UV) radiation | Reduces aflatoxin concentration by up to 40–45%. | Limitations, including low penetration and narrow wavelength range, mean the industry has not yet recognized the patented process of UV inactivation of aflatoxin. | [74] | |
| Main degradation compound of AFB1 seems to retain residual toxicity when exposed to UV light, requiring further degrading into non-toxic forms. | ||||
| Irradiation | High efficiency in eliminating microorganisms and other potential pathogens infecting cereals (over 68.9% for AFB1, over 51% for OTA). | High-energy consumption. Destroys water-soluble vitamins and proteins in cereals. | [75] | |
| Cold plasma | Does not cause heat damage to processed food or affect the protein content. | Plasma treatment of irregularly shaped or bulk food materials can be challenging. | [76] | |
| The potential cytotoxic effect remains unclear. | ||||
| Amplification and continuous processing are challenges of current plasma equipment design. | ||||
| Pulsed light | Potential substitute for traditional technology which does not reduce food quality. | Seldom able to penetrate the cereal, so it is difficult to deal with mycotoxins deep in the cereal. Reduces the germination rate of the seeds. | [75] | |
| Cost-effective non-thermal technology leaves no residue on food materials. | ||||
| Chemical | Fungicides and pesticides | Minimize fungal infections or insect damage to crops, reducing mycotoxin contamination by up to 55–75%. | Can produce residues hazardous to food safety and the environment. | [77] |
| Fungicide concentrations tested in the laboratory exceeded the maximum solubility levels in aqueous media, so effectiveness is unclear. | ||||
| Long-term use will produce drug resistance in molds if residue remains in the cereals, thus affecting eating quality. | ||||
| Ozone | Can inhibit the growth, spore formation, and germination of fungi. | Antibacterial activity largely depends on the type of vegetable/fungi, growth stage, concentration, and exposure time. | [78] | |
| Loss of nutrition or sensory quality in food/feed is negligible. | Degradation products formed by residues are not yet fully determined. | |||
| Ammonia | Reducese AFs, FBs, and OTA to undetectable levels and inhibits the growth of toxin-producing molds. | Infrastructure is complex, and the European community does not allow this method to be used for human food. | [77] | |
| Acid treatment | Degradation of AFs (barely detectable). | Causes chemical residues, limiting its applicability in cereal due to safety concerns. | [77] |
| Product Name | Mycotoxin Limit (μg/kg) a | ||||||
|---|---|---|---|---|---|---|---|
| AFB1 | DON | OTA | ZEN | T-2 | FB1 + FB2 | ||
| Food category | Maize, cornmeal, maize products | ≤20 | ≤1000 | ≤5.0 | ≤60 | ||
| Rice, brown rice | ≤10 | ||||||
| Wheat, barley, other hulled cereals | ≤5.0 | ≤60 | |||||
| Beans, bean products | ≤5.0 | ||||||
| Peanut, peanut products | ≤20 | ||||||
| Vegetable oils (except peanut and maize oil) | ≤10 | ||||||
| Peanut oil, maize oil | ≤20 | ||||||
| Feed raw material | Cereals, processed products | ≤30 | ≤500 | ≤100 | ≤1000 | ≤500 | |
| Processed maize products | ≤50 | ≤500 | ≤60,000 | ||||
| Vegetable oils (except peanut and maize oil) | ≤10 | ≤1000 | |||||
| Peanut oil, maize oil | ≤20 | ||||||
| Other vegetative feed raw materials | ≤30 | ||||||
| Feed products | Compound feed for piglets and young birds | ≤10 | ≤1000 | ≤150 | ≤500 | ||
| Supplementary feed for calves and lambs | ≤20 | ≤500 | ≤2000 | ||||
| Other compound feed | ≤20 | ≤3000 | ≤100 | ≤500 | |||
| Microbial Source | Detoxification Active Substances | Target Mycotoxin | Mycotoxin Detoxification (%) | Detoxification Conditions | Detoxification Products | Reference |
|---|---|---|---|---|---|---|
| Bacillus shackletonii L7 | Bacillus aflatoxin-degrading enzyme (BADE) | Aflatoxin B1 | 92.1 | 37 °C | - | [129] |
| Aflatoxin B2 | 84.1 | |||||
| Aflatoxin M1 | 90.4 | |||||
| Bacillus subtilis UTBSP1 | Extracellular enzymes | Aflatoxin B1 | 85.66 | 35–40 °C | - | [130] |
| Bacillus aryabhattai DT | Extracellular enzymes | Aflatoxin B1 | 78 | 37 °C | - | [131] |
| Bacillus licheniformis BL010 | Two degrading enzymes (Chia010 and Lac010) | Aflatoxin B1 | 89.1 | 30 °C | Molecular formula is C12H14O4 | [132] |
| Bacillus velezensis DY3108 | Extracellular proteins or enzymes | Aflatoxin B1 | 91.5 | 80 °C, pH = 8 | - | [133] |
| Bacillus pimilus E-1-1-1 | Extracellular extracts | Aflatoxin M1 | 100 | 37 °C | - | [134] |
| Aflatoxin B1 | 89.55 | |||||
| Candida versatilis CGMCC3790 | Intracellular components | Aflatoxin B1 | 70 | 25 °C, pH = 5.0 | Molecular formulas are C14H10O4, C14H12O3, C13H12O2, C11H10O4 | [116] |
| Escherichia coli CG1061 | Heat-resistant protein | Aflatoxin B1 | 93.7 | 55 °C, pH = 8.5 | Molecular formula is C16H14O5 | [120] |
| Cladosporium uredinicola CCTCC M 2015181 | Extracellular enzymes | Aflatoxin B1 | 84.50 ± 5.70 | 37 °C | - | [135] |
| Pseudomonas aeruginosa N17-1 | Extracellular enzymes | Aflatoxin B1 | 82.8 | 37 °C | - | [136] |
| Aflatoxin B2 | 46.8 | |||||
| Aflatoxin M1 | 31.9 | |||||
| Pseudomonas putida MTCC 2445 | Intracellular enzymes | Aflatoxin B1 | 80 | 50 °C, pH = 7.0 | - | [137] |
| Bacillus licheniformis | CotA laccase | Aflatoxin B1 | 96 | 37 °C, pH = 8.0 | Aflatoxin Q1 and epi-aflatoxin Q1 | [114] |
| Mycobacterium smegmatis | MSMEG 5998 (aflatoxin-degrading F420H2-dependent reductase) | Aflatoxin B1 | 31 | 22 °C | - | [138] |
| Pseudomonas putida 12-3 | Intracellular enzymes | Aflatoxin B1 | 83.3 | 30 °C, pH = 8.0 | - | [139] |
| Escherichia coli 12-5 | Aflatoxin B1 | 63.8 | ||||
| Tetragenococcus halophilus CGMCC 3792 | Intracellular enzymes | Aflatoxin B1 | 82.8 | 55 °C | Molecular formula is C14H20O2 | [115] |
| Fusarium sp. WCQ3361 | Protein | Aflatoxin B1 | 95.38 | 30 °C | - | [133] |
| Lacticaseibacillus acidophilus PTCC 1643 | - | Aflatoxin M1 | 100 | 21 °C | - | [140] |
| Lactiplantibacillus plantarum PTCC 1058 | 37 °C | |||||
| Flavobacterium aurantiacum | Crude protein extracts | Aflatoxin B1 | 74.5 | 30 °C, pH = 7.0 | - | [141] |
| Phanerochaete sordida YK-624 | Manganese peroxidase (MnP) | Aflatoxin B1 | 86 | 30 °C, pH = 4.5 | AFB1-8,9-dihydrodiol | [119] |
| Rhodococcus erythropolis DSM 14303 | Cell-free extract | Aflatoxin B1 | 95 | 30 °C, pH = 7.0 | - | [127] |
| Nocardia corynebacterioides DSM 12676 | 70 | |||||
| Nocardia corynebacterioides DSM 20151 | 100 | |||||
| Mycobacterium fluoranthenivorans sp. nov. DSM 14304 | 100 | |||||
| Stenotrophomonas maltophilia 35-3 | Extracellular enzymes | Aflatoxin B1 | 82.5 | 37 °C, pH = 8 | - | [142] |
| Bacillus licheniformis CK1 | Extracellular xylanase, CMCase and protease | Zearalenone | 97 | 20 °C | - | [143] |
| Clonostachys rosa IFO 7063 | Lactonohydrolase ZHD101 | Zearalenone | - | pH = 7–10 | 1-(3,5-dihydroxy-phenyl)-10-hydroxy-1-undecen-6-one | [123] |
| Bacillus subtilis 168 | Culture extract | Zearalenone | 81 | 30 °C, pH = 8.0 | - | [128] |
| Bacillus natto CICC 24640 | Zearalenone | 100 | ||||
| Aspergillus niger FS10 | Protease | Zearalenone | 89.92 | 37 °C | - | [144] |
| Pseudomonas putida ZEA-1 | Cell-free extract | Zearalenone | 100 | 30–37 °C, pH = 7.0–8.0 | - | [126] |
| Bacillus pumilus ES-21 | Esterase | Zearalenone | 95.7 | 40.1 °C, pH = 7.60 | 1-(3,5-dihydroxyphenyl)-6′-hydroxy-l′-undecen-l0′-one | [118] |
| Aspergillus niger | OTA hydrolytic enzyme | Ochratoxin A | 99 | 37 °C, pH = 7.5 | - | [145] |
| Yarrowia lipolytica Y-2 | Carboxypeptidases | Ochratoxin A | 97.2 | 28 °C | Otα | [146] |
| Acinetobacter sp. neg1, ITEM 17016 | Carboxypeptidases | Ochratoxin A | 70 | 37 °C | Otα | [147] |
| Bacillus amyloliquefaciens ASAG1 | Carboxypeptidase | Ochratoxin A | 100 | 37 °C, pH = 7.0 | - | [148] |
| Aspergillus niger | Crude lipase (Amano A) | Ochratoxin A | 100 | 30 °C, pH = 7.5 | Otα and phenylalanine | [117] |
| Aspergillus niger W-35 | Ochratoxinase | Ochratoxin A | 85.1 | 37 °C | - | [149] |
| Alcaligenes faecalis | N-acyl-L-amina acid amidohydrolase (AfOTase) | Ochratoxin A | - | 50 °C, pH = 6.5 | - | [150] |
| Sphingopyxis sp. MTA144 | Recombinant carboxylesterase | Fumonisin B1 | 100 | 30 °C, pH = 8.0 | - | [151] |
| Bacterial consortium SAAS79 | Intracellular enzymes | Fumonisin B1 | 90 | 28 °C, pH = 7.0 | pHFB1a or pHFB1b | [121] |
| Bacterium ATCC 55552 | Aminotransferase | Fumonisin B1 | 100 | 25 °C | - | [152] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Tang, X.; Cai, Y.; Zhou, W.-W. Mycotoxin Contamination Status of Cereals in China and Potential Microbial Decontamination Methods. Metabolites 2023, 13, 551. https://doi.org/10.3390/metabo13040551
Zhang J, Tang X, Cai Y, Zhou W-W. Mycotoxin Contamination Status of Cereals in China and Potential Microbial Decontamination Methods. Metabolites. 2023; 13(4):551. https://doi.org/10.3390/metabo13040551
Chicago/Turabian StyleZhang, Jing, Xi Tang, Yifan Cai, and Wen-Wen Zhou. 2023. "Mycotoxin Contamination Status of Cereals in China and Potential Microbial Decontamination Methods" Metabolites 13, no. 4: 551. https://doi.org/10.3390/metabo13040551