
Expression of STING in Women with Morbid Obesity and Nonalcoholic Fatty Liver Disease

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Sample Size
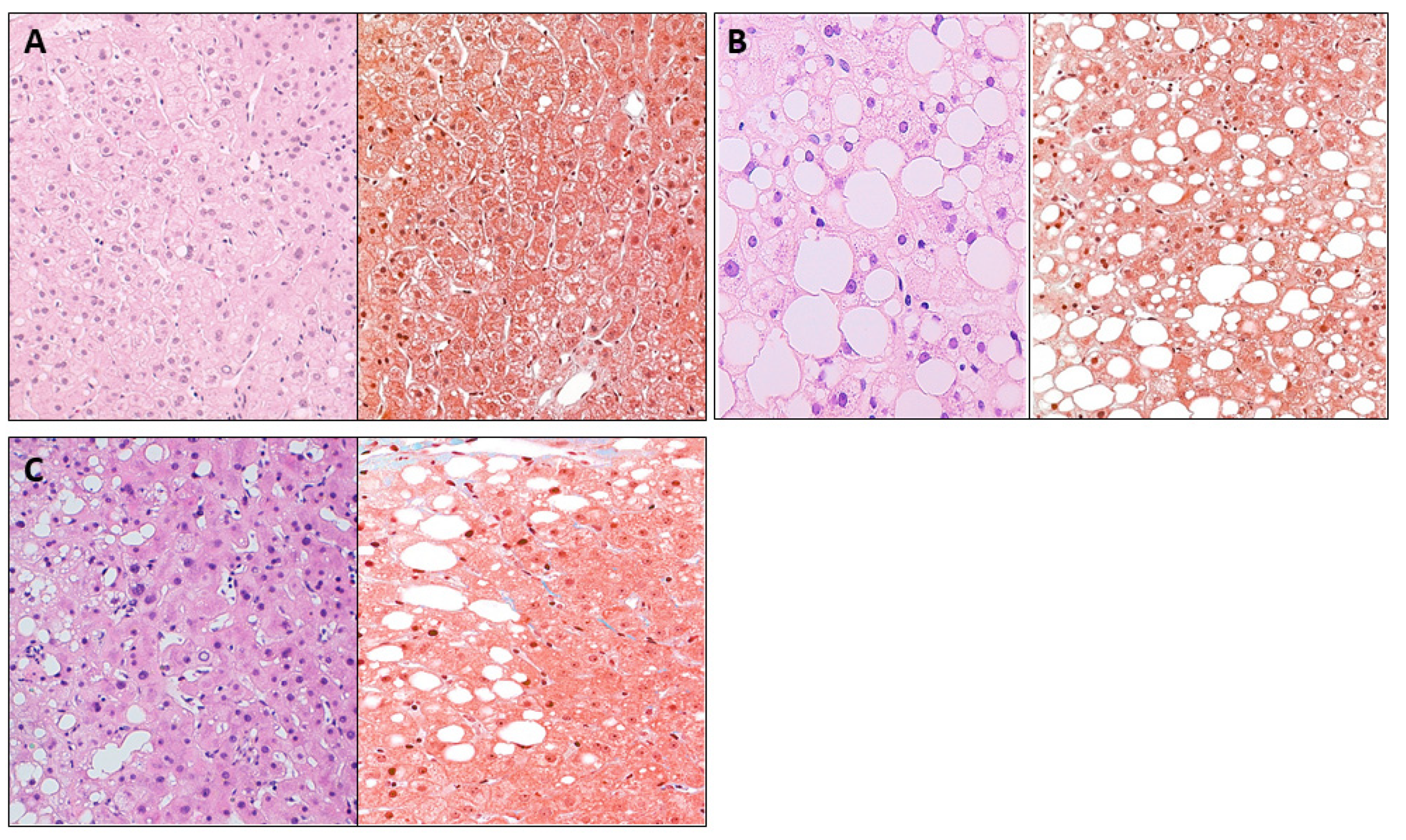
2.3. Liver Pathology
2.4. Biochemical Analyses
2.5. Gene Expression in Liver
2.6. Immunohistochemistry Analysis
2.7. Statistical Analysis
3. Results
3.1. Baseline Features of Subjects
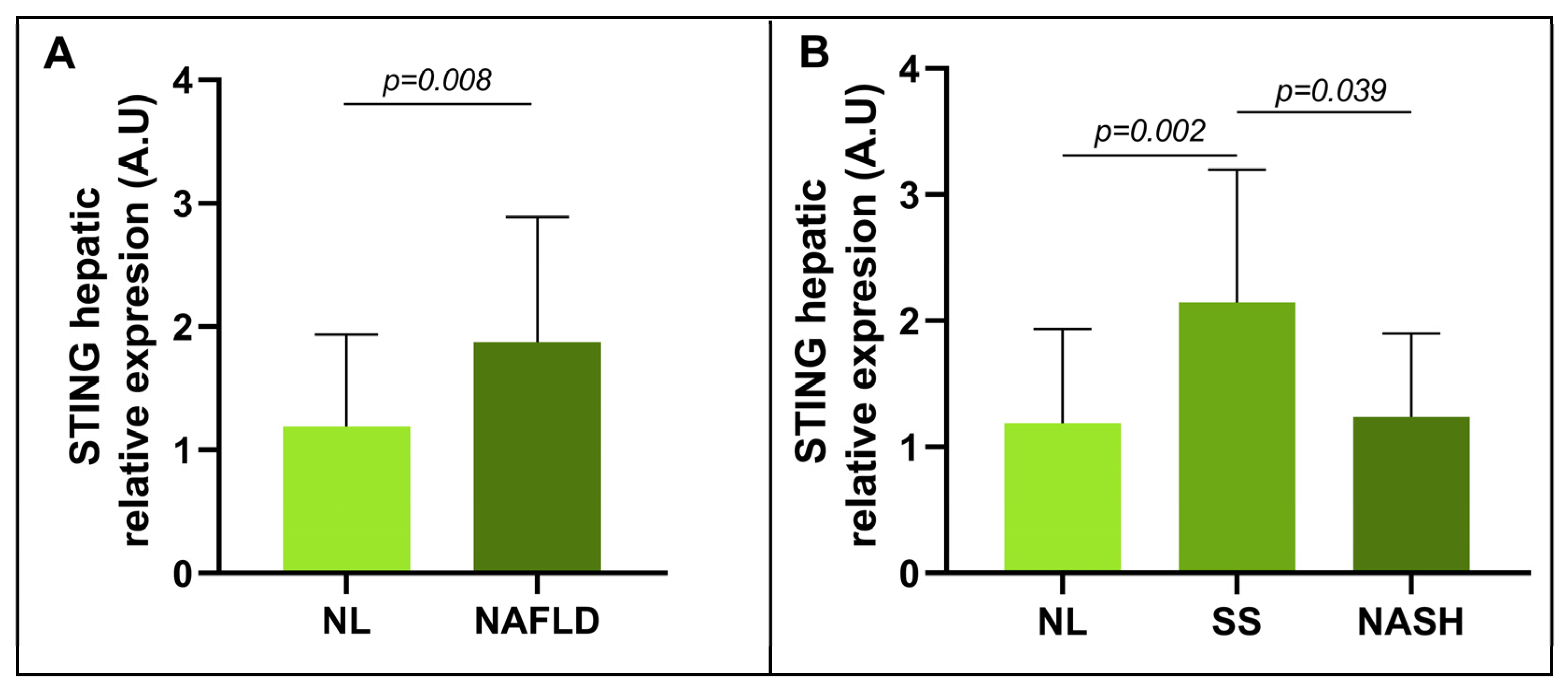
3.2. Evaluation of the Relative mRNA Abundance of STING in Relation to Hepatic Histology
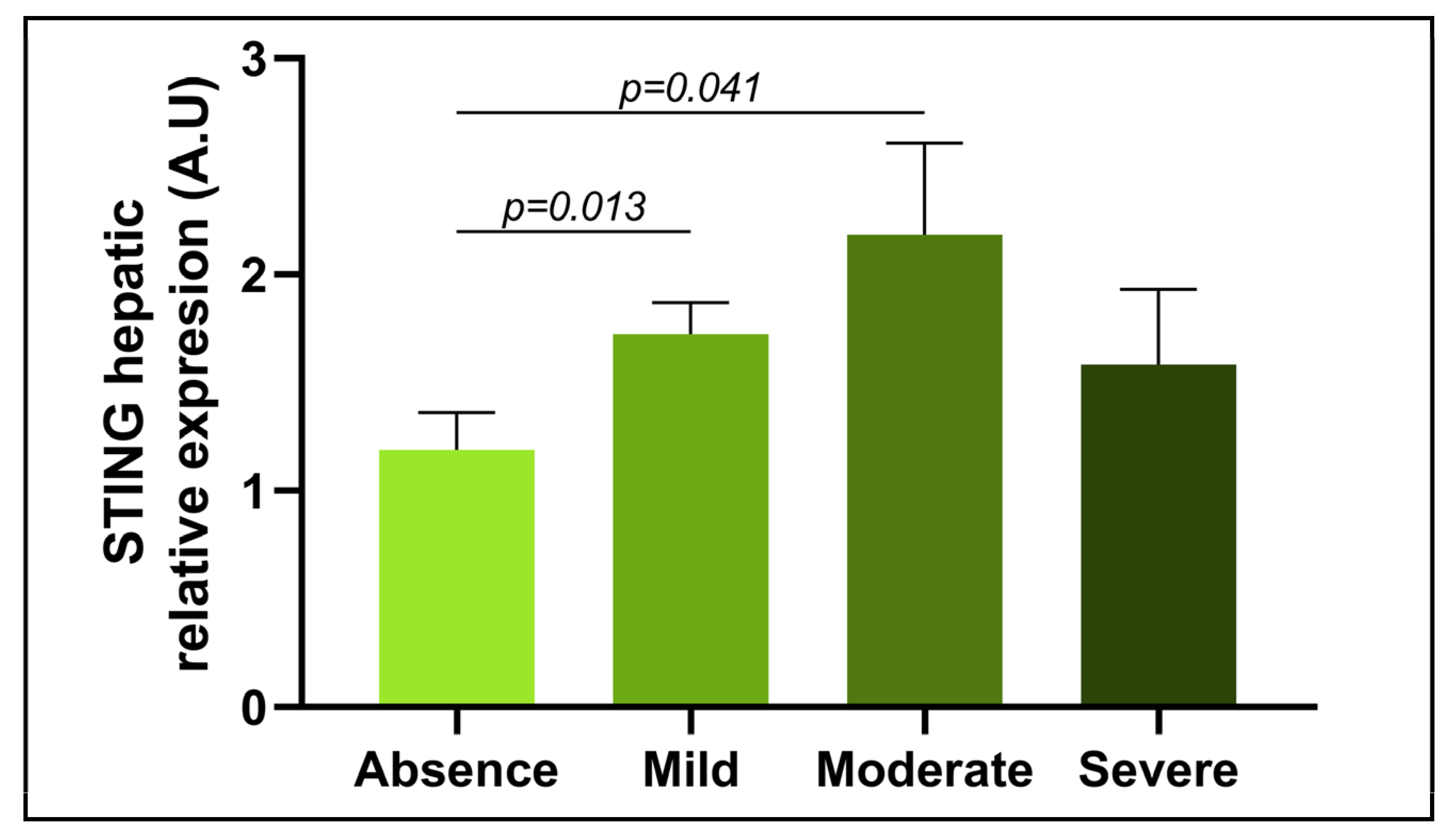
3.3. Evaluation of the Relative mRNA Abundance of STING in Relation to the Severity of Steatosis
3.4. Evaluation of the Relative mRNA Abundance of STING in Relation to NASH-Related Parameters
3.5. Correlation of Relative mRNA Abundance of Hepatic STING with Clinical and Biochemical NAFLD-Related Features
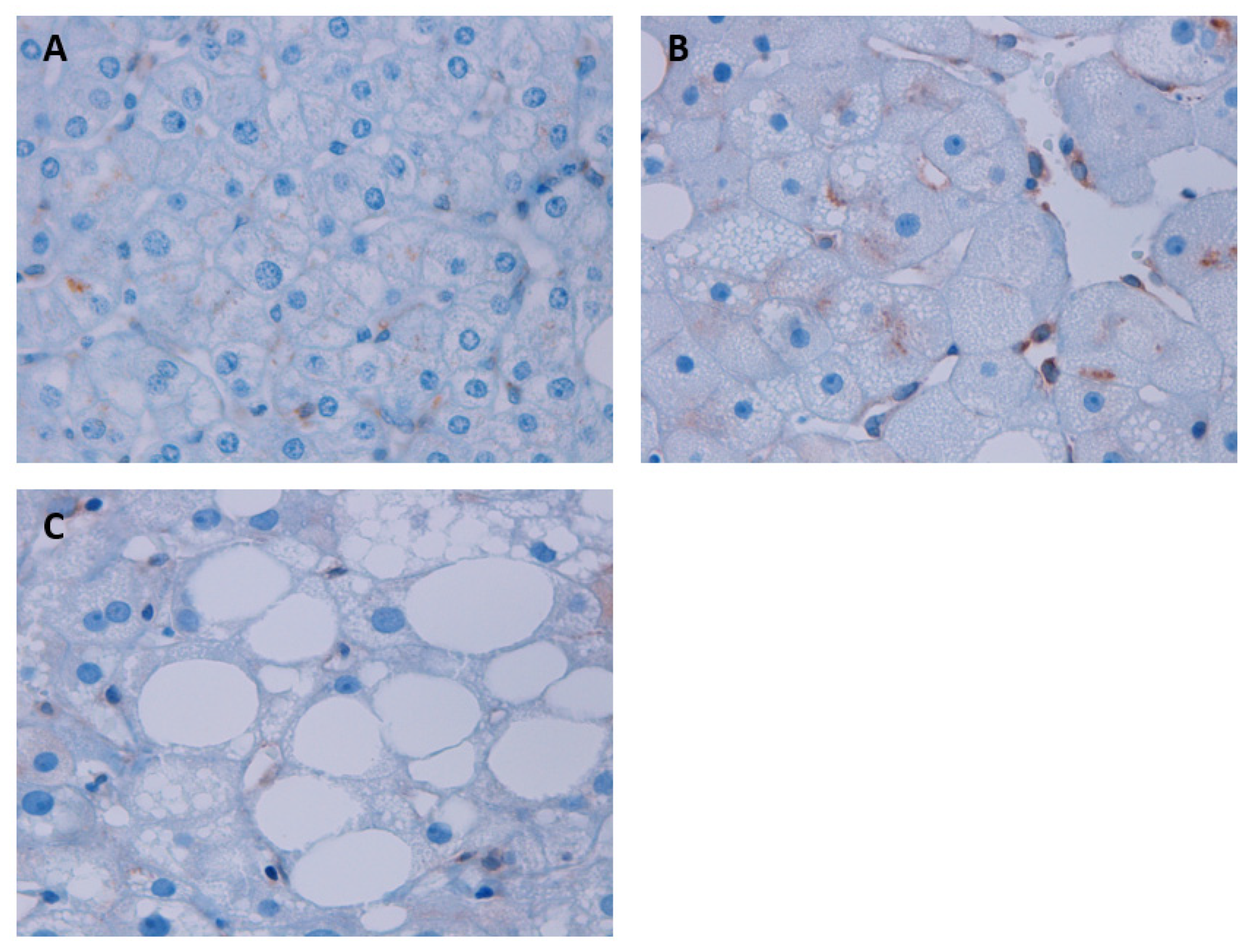
3.6. Assessment of STING Protein Expression According to Liver Histology through the IHC Analysis in Liver Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Celebi, G.; Cicek, A.F.; Gurel, H.; Genc, H.; Kirik, A.; Ercin, C.N.; Dogru, T. Microvesicular Steatosis: A Missed Item in the Management of Nonalcoholic Fatty Liver Disease? Acta Gastro-Enterol. Belg. 2020, 83, 565–570. [Google Scholar]
- Mirmiran, P.; Amirhamidi, Z.; Ejtahed, H.-S.; Bahadoran, Z.; Azizi, F. Relationship between Diet and Non-Alcoholic Fatty Liver Disease: A Review Article. Iran. J. Public Health 2017, 46, 1007–1017. [Google Scholar] [PubMed]
- Byrne, C.D.; Targher, G. NAFLD: A Multisystem Disease. J. Hepatol. 2015, 62, S47–S64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurice, J.; Manousou, P. Non-Alcoholic Fatty Liver Disease. Clin. Med. 2018, 18, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Caballeria, L.; Augustin, S.; Broquetas, T.; Morillas, R.M.; Vergara, M.; Virolés, S.; Hernández, M.R.; Serra, I.; Goday, A.; Vila, L.; et al. Recomendaciones para la detección, diagnóstico y seguimiento de los pacientes con enfermedad por hígado graso no alcohólico en atención primaria y hospitalaria. Med. Clínica 2019, 153, 169–177. [Google Scholar] [CrossRef]
- Brunt, E.M. Pathology of Nonalcoholic Fatty Liver Disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 195–203. [Google Scholar] [CrossRef]
- Pappachan, J.M.; Babu, S.; Krishnan, B.; Ravindran, N.C. Non-Alcoholic Fatty Liver Disease: A Clinical Update. J. Clin. Transl. Hepatol. 2017, 5, 384–393. [Google Scholar] [CrossRef] [Green Version]
- Masarone, M.; Rosato, V.; Dallio, M.; Gravina, A.G.; Aglitti, A.; Loguercio, C.; Federico, A.; Persico, M. Role of Oxidative Stress in Pathophysiology of Nonalcoholic Fatty Liver Disease. Oxid. Med. Cell. Longev. 2018, 2018, 9547613. [Google Scholar] [CrossRef]
- Bessone, F.; Razori, M.V.; Roma, M.G. Molecular Pathways of Nonalcoholic Fatty Liver Disease Development and Progression. Cell. Mol. Life Sci. 2019, 76, 99–128. [Google Scholar] [CrossRef]
- Caldwell, S.; Ikura, Y.; Dias, D.; Isomoto, K.; Yabu, A.; Moskaluk, C.; Pramoonjago, P.; Simmons, W.; Scruggs, H.; Rosenbaum, N.; et al. Hepatocellular Ballooning in NASH. J. Hepatol. 2010, 53, 719–723. [Google Scholar] [CrossRef] [Green Version]
- Bolondi, L.; Gramantieri, L. From Liver Cirrhosis to HCC. Intern. Emerg. Med. 2011, 6, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.P. Liver Cirrhosis, Transplantation and Organ Shortage. Dtsch. Ärztebl. Int. 2013, 11, 83–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnabl, B.; Brenner, D.A. Interactions between the Intestinal Microbiome and Liver Diseases. Gastroenterology 2014, 146, 1513–1524. [Google Scholar] [CrossRef] [Green Version]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The Gut-Liver Axis in Liver Disease: Pathophysiological Basis for Therapy. J. Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascale, A.; Marchesi, N.; Marelli, C.; Coppola, A.; Luzi, L.; Govoni, S.; Giustina, A.; Gazzaruso, C. Microbiota and Metabolic Diseases. Endocrine 2018, 61, 357–371. [Google Scholar] [CrossRef]
- Kanmani, P.; Suganya, K.; Kim, H. The Gut Microbiota: How Does It Influence the Development and Progression of Liver Diseases. Biomedicines 2020, 8, 501. [Google Scholar] [CrossRef]
- Imajo, K.; Yoneda, M.; Ogawa, Y.; Wada, K.; Nakajima, A. Microbiota and Nonalcoholic Steatohepatitis. Semin. Immunopathol. 2014, 36, 115–132. [Google Scholar] [CrossRef]
- Milosevic, I.; Vujovic, A.; Barac, A.; Djelic, M.; Korac, M.; Radovanovic Spurnic, A.; Gmizic, I.; Stevanovic, O.; Djordjevic, V.; Lekic, N.; et al. Gut-Liver Axis, Gut Microbiota, and Its Modulation in the Management of Liver Diseases: A Review of the Literature. Int. J. Mol. Sci. 2019, 20, 395. [Google Scholar] [CrossRef] [Green Version]
- Kirpich, I.A.; Marsano, L.S.; McClain, C.J. Gut-Liver Axis, Nutrition, and Non-Alcoholic Fatty Liver Disease. Clin. Biochem. 2015, 48, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Yang, M. Current Options and Future Directions for NAFLD and NASH Treatment. Int. J. Mol. Sci. 2021, 22, 7571. [Google Scholar] [CrossRef]
- Patel, S.; Jin, L. TMEM173 Variants and Potential Importance to Human Biology and Disease. Genes Immun. 2019, 20, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Qiao, J.T.; Cui, C.; Qing, L.; Wang, L.S.; He, T.Y.; Yan, F.; Liu, F.Q.; Shen, Y.H.; Hou, X.G.; Chen, L. Activation of the STING-IRF3 Pathway Promotes Hepatocyte Inflammation, Apoptosis and Induces Metabolic Disorders in Nonalcoholic Fatty Liver Disease. Metabolism 2018, 81, 13–24. [Google Scholar] [CrossRef]
- Burdette, D.L.; Vance, R.E. STING and the Innate Immune Response to Nucleic Acids in the Cytosol. Nat. Immunol. 2013, 14, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Nakhaei, P.; Hiscott, J.; Lin, R. STING-Ing the Antiviral Pathway. J. Mol. Cell Biol. 2010, 2, 110–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vila, I.K.; Chamma, H.; Steer, A.; Saccas, M.; Taffoni, C.; Turtoi, E.; Reinert, L.S.; Hussain, S.; Marines, J.; Jin, L.; et al. STING Orchestrates the Crosstalk between Polyunsaturated Fatty Acid Metabolism and Inflammatory Responses. Cell Metab. 2022, 34, 125–139.e8. [Google Scholar] [CrossRef] [PubMed]
- Couillin, I.; Riteau, N. STING Signaling and Sterile Inflammation. Front. Immunol. 2021, 12, 753789. [Google Scholar] [CrossRef]
- Chen, C.; Yang, R.-X.; Xu, H.-G. STING and Liver Disease. J. Gastroenterol. 2021, 56, 704–712. [Google Scholar] [CrossRef]
- Cheng, Z.; Dai, T.; He, X.; Zhang, Z.; Xie, F.; Wang, S.; Zhang, L.; Zhou, F. The Interactions between CGAS-STING Pathway and Pathogens. Signal Transduct. Target. Ther. 2020, 5, 91. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Liu, F. The CGAS-CGAMP-STING Pathway: A Molecular Link Between Immunity and Metabolism. Diabetes 2019, 68, 1099–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Møhlenberg, M.; Terczynska-Dyla, E.; Thomsen, K.L.; George, J.; Eslam, M.; Grønbæk, H.; Hartmann, R. The Role of IFN in the Development of NAFLD and NASH. Cytokine 2019, 124, 154519. [Google Scholar] [CrossRef]
- Chen, R.; Du, J.; Zhu, H.; Ling, Q. The Role of CGAS-STING Signalling in Liver Diseases. JHEP Rep. 2021, 3, 100324. [Google Scholar] [CrossRef]
- Luo, X.; Li, H.; Ma, L.; Zhou, J.; Guo, X.; Woo, S.-L.; Pei, Y.; Knight, L.R.; Deveau, M.; Chen, Y.; et al. Expression of STING Is Increased in Liver Tissues From Patients With NAFLD and Promotes Macrophage-Mediated Hepatic Inflammation and Fibrosis in Mice. Gastroenterology 2018, 155, 1971–1984.e4. [Google Scholar] [CrossRef] [Green Version]
- Basaranoglu, M. Fructose as a Key Player in the Development of Fatty Liver Disease. World J. Gastroenterol. 2013, 19, 1166. [Google Scholar] [CrossRef] [PubMed]
- Akhmetova, K.; Balasov, M.; Chesnokov, I. Drosophila STING Protein Has a Role in Lipid Metabolism. eLife 2021, 10, e67358. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, N.; Li, Z.; Xu, G.; Zhan, X.; Tang, J.; Xiao, X.; Bai, Z. The Cytosolic DNA-Sensing CGAS-STING Pathway in Liver Diseases. Front. Cell Dev. Biol. 2021, 9, 717610. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Tian, Y.; Xia, Q.; Ke, B. The CGAS-STING Pathway: Novel Perspectives in Liver Diseases. Front. Immunol. 2021, 12, 682736. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Rao, X.; Wang, X.; Luo, Z.; Wang, J.; Sheng, S.; Liu, Y.; Zhang, N.; Jin, S.; Chen, H.; et al. CGAS-STING Signaling Pathway and Liver Disease: From Basic Research to Clinical Practice. Front. Pharmacol. 2021, 12, 719644. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, Q.; Yan, C.; Niu, C.; Zhou, J.; Liu, J.; Song, Y.; Zhou, F.; Fan, Y.; Ren, J.; et al. The Absence of STING Ameliorates Non-Alcoholic Fatty Liver Disease and Reforms Gut Bacterial Community. Front. Immunol. 2022, 13, 931176. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.-C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and Validation of a Histological Scoring System for Nonalcoholic Fatty Liver Disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Brunt, E.M.; Janney, C.G.; Di Bisceglie, A.M.; Neuschwander-Tetri, B.A.; Bacon, B.R. Nonalcoholic Steatohepatitis: A Proposal for Grading and Staging The Histological Lesions. Am. J. Gastroenterol. 1999, 94, 2467–2474. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Rao, H.; Zhao, J.; Wee, A.; Li, X.; Fei, R.; Huang, R.; Wu, C.; Liu, F.; Wei, L. STING Expression in Monocyte-Derived Macrophages Is Associated with the Progression of Liver Inflammation and Fibrosis in Patients with Nonalcoholic Fatty Liver Disease. Lab. Investig. 2020, 100, 542–552. [Google Scholar] [CrossRef]
- Vila, I.K.; Guha, S.; Kalucka, J.; Olagnier, D.; Laguette, N. Alternative Pathways Driven by STING: From Innate Immunity to Lipid Metabolism. Cytokine Growth Factor Rev. 2022, 68, 54–68. [Google Scholar] [CrossRef] [PubMed]
- Neuman, M.G.; Malnick, S.; Chertin, L. Gamma Glutamyl Transferase—An Underestimated Marker for Cardiovascular Disease and the Metabolic Syndrome. J. Pharm. Pharm. Sci. 2020, 23, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien-Min, K.; Cheng-Chuan, L. Clinical Criteria Correlated with the Incidence of Patients with Non-Alcoholic Fatty Liver Disease. Ann. Clin. Lab. Sci. 2017, 47, 191–200. [Google Scholar] [PubMed]
- Zhou, Y.-J.; Zou, H.; Zheng, J.-N.; Zou, T.-T.; Vitale, A.; Miele, L.; Van Poucke, S.; Liu, W.-Y.; Shen, S.; Zhang, D.-C.; et al. Serum Alkaline Phosphatase, a Risk Factor for Non-Alcoholic Fatty Liver, but Only for Women in Their 30s and 40s: Evidence from a Large Cohort Study. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 269–276. [Google Scholar] [CrossRef]
- Sattar, N.; Forrest, E.; Preiss, D. Non-Alcoholic Fatty Liver Disease. BMJ 2014, 349, g4596. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, Y.; An, W.; Song, J.; Zhang, Y.; Zhao, X. STING-Mediated Inflammation in Kupffer Cells Contributes to Progression of Nonalcoholic Steatohepatitis. J. Clin. Investig. 2018, 129, 546–555. [Google Scholar] [CrossRef]
- Gottlieb, A.; Canbay, A. Why Bile Acids Are So Important in Non-Alcoholic Fatty Liver Disease (NAFLD) Progression. Cells 2019, 8, 1358. [Google Scholar] [CrossRef]
- Fontes-Cal, T.C.M.; Mattos, R.T.; Medeiros, N.I.; Pinto, B.F.; Belchior-Bezerra, M.; Roque-Souza, B.; Dutra, W.O.; Ferrari, T.C.A.; Vidigal, P.V.T.; Faria, L.C.; et al. Crosstalk Between Plasma Cytokines, Inflammation, and Liver Damage as a New Strategy to Monitoring NAFLD Progression. Front. Immunol. 2021, 12, 708959. [Google Scholar] [CrossRef]
- Khanna, D.; Khanna, S.; Khanna, P.; Kahar, P.; Patel, B.M. Obesity: A Chronic Low-Grade Inflammation and Its Markers. Cureus 2022, 14, e22711. [Google Scholar] [CrossRef]
- Mraz, M.; Lacinova, Z.; Drapalova, J.; Haluzikova, D.; Horinek, A.; Matoulek, M.; Trachta, P.; Kavalkova, P.; Svacina, S.; Haluzik, M. The Effect of Very-Low-Calorie Diet on MRNA Expression of Inflammation-Related Genes in Subcutaneous Adipose Tissue and Peripheral Monocytes of Obese Patients with Type 2 Diabetes Mellitus. J. Clin. Endocrinol. Metab. 2011, 96, E606–E613. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | NL (n = 27) | SS (n = 26) | NASH (n = 16) |
|---|---|---|---|
| Weight (kg) | 117 (107–131) | 114 (108.98–128.6) | 110.5 (104.33–120.75) |
| BMI (kg/m2) | 43.50 (40.89–46.88) | 44.35 (40.87–46.8) | 44.19 (40.69–45.81) |
| SBP (mmHg) | 120 (100–132.5) | 117.5 (108.5–127) | 115 (102–127) |
| DBP (mmHg) | 63 (57.5–73) | 62 (59.5–73.75) | 64 (55–70) |
| Glucose (mg/dL) | 85.04 (76.03–93.14) | 93.14 (87.2–107.02) * | 91.52 (82.33–101.97) |
| Insulin (mUI/L) | 9.57 (5.55–16.82) | 10.17 (7.23–13.93) | 7.19 (5.14–26.02) |
| HbA1c (%) | 5.5 (5.3–5.7) | 5.55 (5.3–5.95) | 5.55 (5.15–6.13) |
| HOMA1 | 2.05 (1.03–3.45) | 2.52 (1.38–3.68) | 1.63 (1.26–4.23) |
| Cholesterol (mg/dL) | 170 (148.25–209.5) | 171.15 (136.25–194.25) | 183.9 (152.75–229.5) |
| HDL-c (mg/dL) | 40.6 (32.05–48.5) | 43.5 (33.75–47) | 37.8 (33.5–48.5) |
| LDL-c (mg/dL) | 107.9 (86–134.2) | 104.1 (77.20–126.25) | 94 (79.3–128.03) |
| TG (mg/dL) | 106.5 (94–136) | 117.5 (82.25–172.5) | 153 (116.5–256.5) * |
| AST (UI/L) | 20 (15.5–36.5) | 23 (17–35) | 27 (17.25–43.5) |
| ALT (UI/L) | 22.5 (16–37.5) | 31 (22–35.25) | 32 (16.25–41) |
| GGT (UI/L) | 18 (15.25–26.25) | 21 (16–32.25) | 25.5 (18–28.75) |
| ALP (UI/L) | 58.5 (49.25–71.25) | 74 (64–86.25) * | 63 (55–74.5) $ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertran, L.; Adalid, L.; Vilaró-Blay, M.; Barrientos-Riosalido, A.; Aguilar, C.; Martínez, S.; Sabench, F.; del Castillo, D.; Porras, J.A.; Alibalic, A.; et al. Expression of STING in Women with Morbid Obesity and Nonalcoholic Fatty Liver Disease. Metabolites 2023, 13, 496. https://doi.org/10.3390/metabo13040496
Bertran L, Adalid L, Vilaró-Blay M, Barrientos-Riosalido A, Aguilar C, Martínez S, Sabench F, del Castillo D, Porras JA, Alibalic A, et al. Expression of STING in Women with Morbid Obesity and Nonalcoholic Fatty Liver Disease. Metabolites. 2023; 13(4):496. https://doi.org/10.3390/metabo13040496
Chicago/Turabian StyleBertran, Laia, Laia Adalid, Mercè Vilaró-Blay, Andrea Barrientos-Riosalido, Carmen Aguilar, Salomé Martínez, Fàtima Sabench, Daniel del Castillo, José Antonio Porras, Ajla Alibalic, and et al. 2023. "Expression of STING in Women with Morbid Obesity and Nonalcoholic Fatty Liver Disease" Metabolites 13, no. 4: 496. https://doi.org/10.3390/metabo13040496