

Mechanisms of Maternal Diet-Induced Obesity Affecting the Offspring Brain and Development of Affective Disorders

Abstract
:1. Introduction
2. Clinical Studies

{kind=link}
{kind=link}
| Maternal Risk Factor | Child Outcome (Age at Follow-Up) | Study Design and Sample Size | Covariates Investigated | Author, Year |
|---|---|---|---|---|
| ↑ Pre-pregnancy BMI and/or gestational weight gain | ↑ teacher-rated ADHD symptomology (7–8 years or 10–12 years) | Cohort study, N = 12,556 | Maternal age, social adversity, maternal smoking, gestation length, birth weight, infant sex, maternal education, family structure | Rodriguez et al., 2008 [35] |
| ↑ teacher-rated ADHD symptomology (5 years) | Cohort study, N = 1714 | Rodriguez, 2010 [36] | ||
| Pre-pregnancy obesity | ↑ Affective behavioural problems as determined by primary caregiver (5, 8, 10, 14, 17 years) | Longitudinal cohort study, N = 1754 | Maternal age, maternal education, household income, maternal smoking, alcohol consumption, family structure, maternal life stress, gestation length, gestational diabetes, duration of breastfeeding, birth weight | Robinson et al., 2012 [37] |
| ↑ Pre-pregnancy BMI | ↑ Internalising and externalizing problems in childhood and adolescence (5, 8, 10, 14, 17 years) | Longitudinal cohort study, N = 2868 | Maternal psychopathology, paternal psychopathology, maternal age, alcohol intake, smoking, socioeconomic status, education, marital status, maternal life stress, maternal diabetes | Van Lieshout et al., 2013 [38] |
| Pre-pregnancy obesity | ↓Child psychosocial development at age 6↑ likelihood of prior ADD/ADHD diagnosis, speech/language therapy, psychological or special needs services (6 years) | Cohort study, N = 1542 | Maternal age, ethnicity, marital status, education, parity, household income, maternal smoking, infant sex, infant current weight, child enrichment activities (e.g., reading to child >3 times per week), breastfeeding duration, postpartum depression, birth weight | Jo et al., 2015 [39] |
| ↑ Pre-pregnancy BMI, pre-pregnancy and gestational diabetes | ↑ risk of neurodevelopmental, ADD/ADHD, psychotic, mood, stress-related disorders in offspring (up to 11 years, median 5.5 years) | Population study, N = 649,043 | Infant birth year, infant sex, perinatal problems, gestation length, parity, mode of delivery, maternal age, family structure, ethnicity, maternal smoking, maternal inflammatory comorbidities | Kong et al., 2018 [40] |
| Infant birth year, infant sex, perinatal problems, gestation length, parity, mode of delivery, maternal age, family structure, ethnicity, maternal smoking, maternal inflammatory comorbidities, maternal psychopathology | Kong et al., 2020 [42] |
3. Maternal Factors Influencing Offspring Neurodevelopment and Behaviour
3.1. Maternal Care Behaviour
3.2. The Maternal Microbiome
3.3. Maternal Inflammation
3.4. Breast Milk Composition
3.5. Neurotransmitter Systems
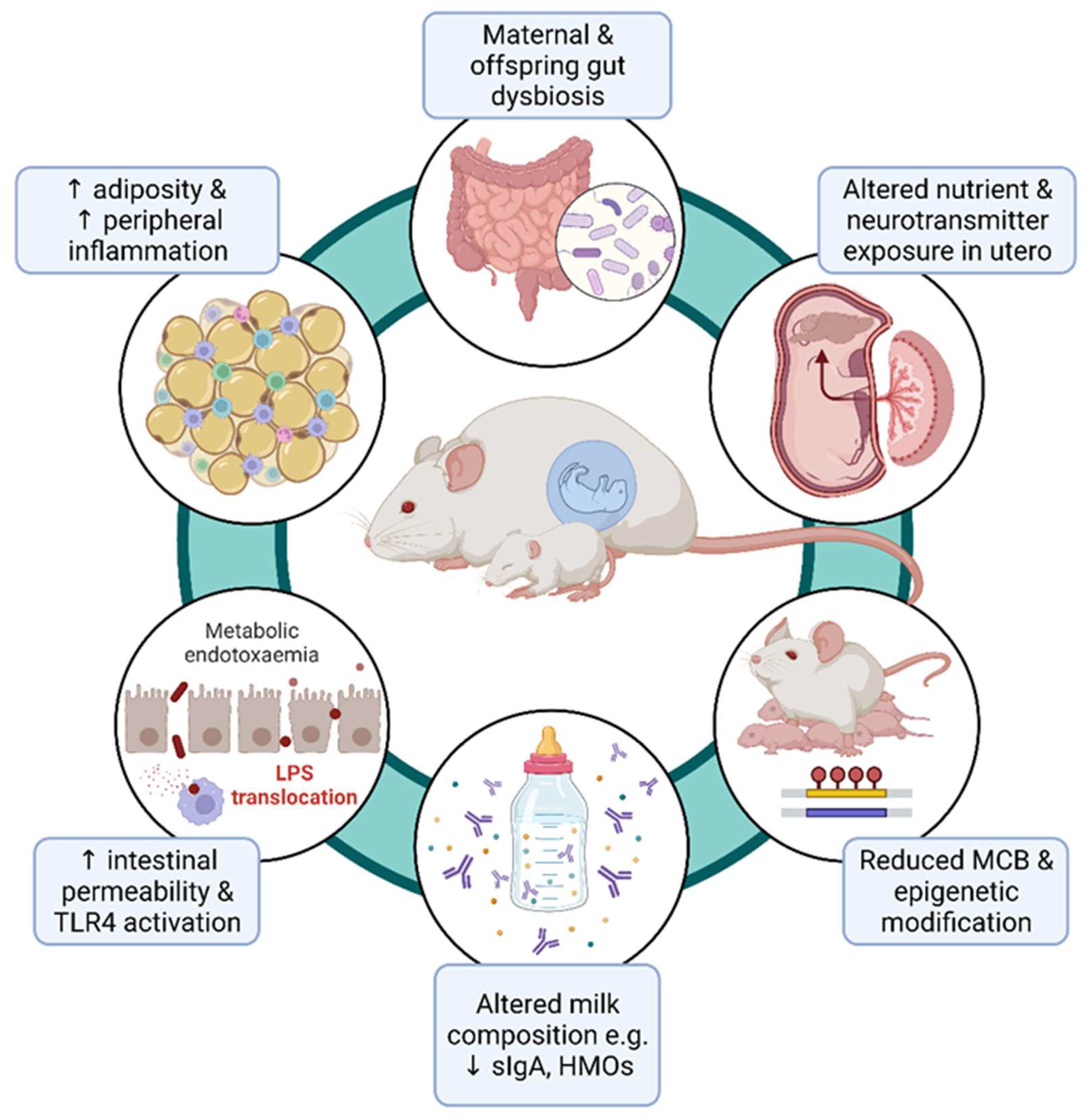
4. Perturbed Metabolism in Offspring Exposed to Maternal Obesity
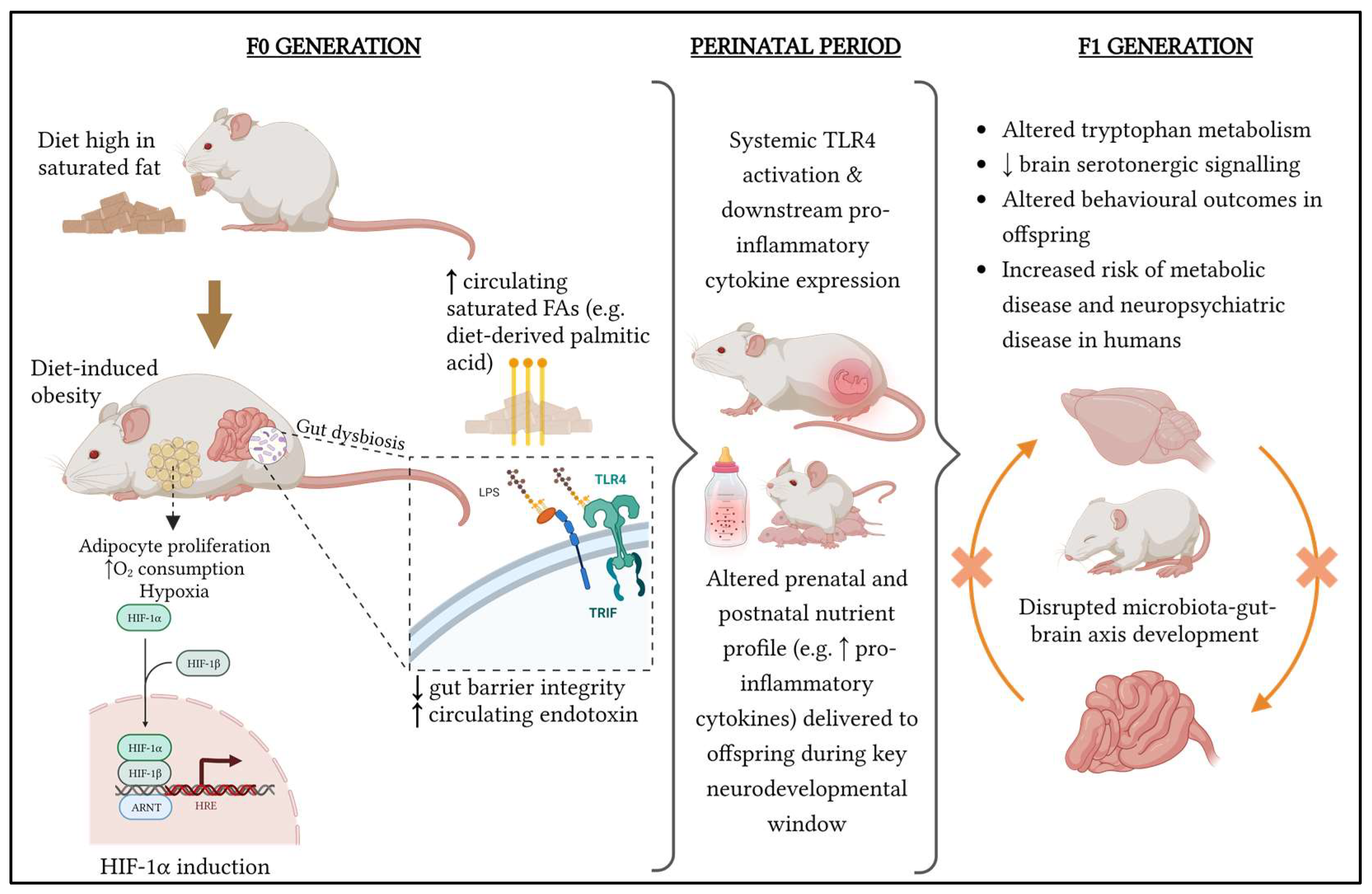
5. Overview of Maternal Influences on the Offspring and Looking beyond the First Filial Generation
6. Translational Perspectives and Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Afshin, A.; Reitsma, M.B.; Murray, C.J.L. Health Effects of Overweight and Obesity in 195 Countries. N. Engl. J. Med. 2017, 377, 1496–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NMPA Project Team. National Maternity and Perinatal Audit: Clinical Report 2017; RCOG: London, UK, 2017. [Google Scholar]
- Devlieger, R.; Benhalima, K.; Damm, P.; Van Assche, A.; Mathieu, C.; Mahmood, T.; Dunne, F.; Bogaerts, A. Maternal Obesity in Europe: Where Do We Stand and How to Move Forward? A Scientific Paper Commissioned by the European Board and College of Obstetrics and Gynaecology (EBCOG). Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 201, 203–208. [Google Scholar] [CrossRef]
- Gaillard, R. Maternal Obesity during Pregnancy and Cardiovascular Development and Disease in the Offspring. Eur. J. Epidemiol. 2015, 30, 1141–1152. [Google Scholar] [CrossRef] [Green Version]
- Campbell, E.E.; Dworatzek, P.D.N.; Penava, D.; de Vrijer, B.; Gilliland, J.; Matthews, J.I.; Seabrook, J.A. Factors That Influence Excessive Gestational Weight Gain: Moving beyond Assessment and Counselling. J. Matern. Fetal. Neonatal Med. 2016, 29, 3527–3531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fung, T.T.; Rimm, E.B.; Spiegelman, D.; Rifai, N.; Tofler, G.H.; Willett, W.C.; Hu, F.B. Association between Dietary Patterns and Plasma Biomarkers of Obesity and Cardiovascular Disease Risk. Am. J. Clin. Nutr. 2001, 73, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Leddy, M.A.; Power, M.L.; Schulkin, J. The Impact of Maternal Obesity on Maternal and Fetal Health. Rev. Obstet. Gynecol. 2008, 1, 170–178. [Google Scholar]
- Mulligan, C.M.; Friedman, J.E. Maternal Modifiers of the Infant Gut Microbiota: Metabolic Consequences. J. Endocrinol. 2017, 235, R1–R12. [Google Scholar] [CrossRef] [Green Version]
- Edlow, A.G. Maternal Obesity and Neurodevelopmental and Psychiatric Disorders in Offspring. Prenat. Diagn. 2017, 37, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Bilbo, S.D.; Tsang, V. Enduring Consequences of Maternal Obesity for Brain Inflammation and Behavior of Offspring. FASEB J. 2010, 24, 2104–2115. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, E.L.; Grayson, B.; Takahashi, D.; Robertson, N.; Maier, A.; Bethea, C.L.; Smith, M.S.; Coleman, K.; Grove, K.L. Chronic Consumption of a High-Fat Diet during Pregnancy Causes Perturbations in the Serotonergic System and Increased Anxiety-like Behavior in Nonhuman Primate Offspring. J. Neurosci. 2010, 30, 3826–3830. [Google Scholar] [CrossRef] [Green Version]
- Radford-Smith, D.E.; Probert, F.; Burnet, P.W.J.; Anthony, D.C. Modifying the Maternal Microbiota Alters the Gut-Brain Metabolome and Prevents Emotional Dysfunction in the Adult Offspring of Obese Dams. Proc. Natl. Acad. Sci. USA 2022, 119, e2108581119. [Google Scholar] [CrossRef] [PubMed]
- Rincel, M.; Lépinay, A.L.; Delage, P.; Fioramonti, J.; Théodorou, V.S.; Layé, S.; Darnaudéry, M. Maternal High-Fat Diet Prevents Developmental Programming by Early-Life Stress. Transl. Psychiatry 2016, 6, e966. [Google Scholar] [CrossRef] [PubMed]
- Baptissart, M.; Lamb, H.E.; To, K.; Bradish, C.; Tehrani, J.; Reif, D.; Cowley, M. Neonatal Mice Exposed to a High-Fat Diet in Utero Influence the Behaviour of Their Nursing Dam. Proc. Biol. Sci. 2018, 285, 20181237. [Google Scholar] [CrossRef] [Green Version]
- Abuaish, S.; Spinieli, R.L.; McGowan, P.O. Perinatal High Fat Diet Induces Early Activation of Endocrine Stress Responsivity and Anxiety-like Behavior in Neonates. Psychoneuroendocrinology 2018, 98, 11–21. [Google Scholar] [CrossRef]
- Buffington, S.A.; Di Prisco, G.V.; Auchtung, T.A.; Ajami, N.J.; Petrosino, J.F.; Costa-Mattioli, M. Microbial Reconstitution Reverses Maternal Diet-Induced Social and Synaptic Deficits in Offspring. Cell 2016, 165, 1762–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, T.R.; Kim, D.W.; Glendining, K.A.; Jasoni, C.L. Maternal Obesity and IL-6 Lead to Aberrant Developmental Gene Expression and Deregulated Neurite Growth in the Fetal Arcuate Nucleus. Endocrinology 2014, 155, 2566–2577. [Google Scholar] [CrossRef]
- Cohen, S.; Ke, X.; Liu, Q.; Fu, Q.; Majnik, A.; Lane, R. Adverse Early Life Environment Increases Hippocampal Microglia Abundance in Conjunction with Decreased Neural Stem Cells in Juvenile Mice. Int. J. Dev. Neurosci. 2016, 55, 56–65. [Google Scholar] [CrossRef]
- Thompson, J.R.; Valleau, J.C.; Barling, A.N.; Franco, J.G.; DeCapo, M.; Bagley, J.L.; Sullivan, E.L. Exposure to a High-Fat Diet during Early Development Programs Behavior and Impairs the Central Serotonergic System in Juvenile Non-Human Primates. Front. Endocrinol. 2017, 8, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safi-Stibler, S.; Thévenot, E.A.; Jouneau, L.; Jouin, M.; Seyer, A.; Jammes, H.; Rousseau-Ralliard, D.; Baly, C.; Gabory, A. Differential Effects of Post-Weaning Diet and Maternal Obesity on Mouse Liver and Brain Metabolomes. Nutrients 2020, 12, 1572. [Google Scholar] [CrossRef]
- Winther, G.; Eskelund, A.; Bay-Richter, C.; Elfving, B.; Müller, H.K.; Lund, S.; Wegener, G. Grandmaternal High-Fat Diet Primed Anxiety-like Behaviour in the Second-Generation Female Offspring. Behav. Brain Res. 2019, 359, 47–55. [Google Scholar] [CrossRef]
- McDonald, S.D.; Han, Z.; Mulla, S.; Beyene, J.; Knowledge Synthesis Group. Overweight and Obesity in Mothers and Risk of Preterm Birth and Low Birth Weight Infants: Systematic Review and Meta-Analyses. BMJ 2010, 341, c3428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, M.C.; Gortmaker, S.L.; Sobol, A.M. Very Low Birth Weight Children: Behavior Problems and School Difficulty in a National Sample. J. Pediatr. 1990, 117, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Kelly, Y.J.; Nazroo, J.Y.; McMunn, A.; Boreham, R.; Marmot, M. Birthweight and Behavioural Problems in Children: A Modifiable Effect? Int. J. Epidemiol. 2001, 30, 88–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breslau, N.; Klein, N.; Allen, L. Very Low Birthweight: Behavioral Sequelae at Nine Years of Age. J. Am. Acad. Child Adolesc. Psychiatry 1988, 27, 605–612. [Google Scholar] [CrossRef]
- Ross, G.; Lipper, E.G.; Auld, P.A. Social Competence and Behavior Problems in Premature Children at School Age. Pediatrics 1990, 86, 391–397. [Google Scholar] [CrossRef]
- Klebanov, P.K.; Brooks-Gunn, J.; McCormick, M.C. Classroom Behavior of Very Low Birth Weight Elementary School Children. Pediatrics 1994, 94, 700–708. [Google Scholar] [CrossRef]
- Pharoah, P.O.; Stevenson, C.J.; Cooke, R.W.; Stevenson, R.C. Prevalence of Behaviour Disorders in Low Birthweight Infants. Arch. Dis. Child. 1994, 70, 271–274. [Google Scholar] [CrossRef] [Green Version]
- Chapieski, M.L.; Evankovich, K.D. Behavioral Effects of Prematurity. Semin. Perinatol. 1997, 21, 221–239. [Google Scholar] [CrossRef]
- Sykes, D.H.; Hoy, E.A.; Bill, J.M.; McClure, B.G.; Halliday, H.L.; Reid, M.M. Behavioural Adjustment in School of Very Low Birthweight Children. J. Child Psychol. Psychiatry 1997, 38, 315–325. [Google Scholar] [CrossRef]
- Stjernqvist, K.; Svenningsen, N.W. Ten-Year Follow-up of Children Born before 29 Gestational Weeks: Health, Cognitive Development, Behaviour and School Achievement. Acta Paediatr. 1999, 88, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Zubrick, S.R.; Kurinczuk, J.J.; McDermott, B.M.; McKelvey, R.S.; Silburn, S.R.; Davies, L.C. Fetal Growth and Subsequent Mental Health Problems in Children Aged 4 to 13 Years. Dev. Med. Child Neurol. 2000, 42, 14–20. [Google Scholar] [CrossRef]
- Hille, E.T.; den Ouden, A.L.; Saigal, S.; Wolke, D.; Lambert, M.; Whitaker, A.; Pinto-Martin, J.A.; Hoult, L.; Meyer, R.; Feldman, J.F.; et al. Behavioural Problems in Children Who Weigh 1000 g or Less at Birth in Four Countries. Lancet 2001, 357, 1641–1643. [Google Scholar] [CrossRef]
- Mick, E.; Biederman, J.; Prince, J.; Fischer, M.J.; Faraone, S.V. Impact of Low Birth Weight on Attention-Deficit Hyperactivity Disorder. J. Dev. Behav. Pediatr. 2002, 23, 16–22. [Google Scholar] [CrossRef]
- Rodriguez, A.; Miettunen, J.; Henriksen, T.B.; Olsen, J.; Obel, C.; Taanila, A.; Ebeling, H.; Linnet, K.M.; Moilanen, I.; Järvelin, M.-R. Maternal Adiposity Prior to Pregnancy Is Associated with ADHD Symptoms in Offspring: Evidence from Three Prospective Pregnancy Cohorts. Int. J. Obes. 2008, 32, 550–557. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A. Maternal Pre-Pregnancy Obesity and Risk for Inattention and Negative Emotionality in Children. J. Child Psychol. Psychiatry 2010, 51, 134–143. [Google Scholar] [CrossRef]
- Robinson, M.; Zubrick, S.R.; Pennell, C.E.; Van Lieshout, R.J.; Jacoby, P.; Beilin, L.J.; Mori, T.A.; Stanley, F.J.; Newnham, J.P.; Oddy, W.H. Pre-Pregnancy Maternal Overweight and Obesity Increase the Risk for Affective Disorders in Offspring. J. Dev. Orig. Health Dis. 2013, 4, 42–48. [Google Scholar] [CrossRef]
- Van Lieshout, R.J.; Robinson, M.; Boyle, M.H. Maternal Pre-Pregnancy Body Mass Index and Internalizing and Externalizing Problems in Offspring. Can. J. Psychiatry 2013, 58, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Jo, H.; Schieve, L.A.; Sharma, A.J.; Hinkle, S.N.; Li, R.; Lind, J.N. Maternal Prepregnancy Body Mass Index and Child Psychosocial Development at 6 Years of Age. Pediatrics 2015, 135, e1198–e1209. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Norstedt, G.; Schalling, M.; Gissler, M.; Lavebratt, C. The Risk of Offspring Psychiatric Disorders in the Setting of Maternal Obesity and Diabetes. Pediatrics 2018, 142, e20180776. [Google Scholar] [CrossRef] [Green Version]
- Cosgrove, V.E.; Rhee, S.H.; Gelhorn, H.L.; Boeldt, D.; Corley, R.C.; Ehringer, M.A.; Young, S.E.; Hewitt, J.K. Structure and Etiology of Co-Occurring Internalizing and Externalizing Disorders in Adolescents. J. Abnorm. Child Psychol. 2011, 39, 109–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.; Nilsson, I.A.K.; Brismar, K.; Gissler, M.; Lavebratt, C. Associations of Different Types of Maternal Diabetes and Body Mass Index with Offspring Psychiatric Disorders. JAMA Netw. Open 2020, 3, e1920787. [Google Scholar] [CrossRef] [PubMed]
- Harlow, H.F.; Zimmermann, R.R. Affectional Responses in the Infant Monkey; Orphaned Baby Monkeys Develop a Strong and Persistent Attachment to Inanimate Surrogate Mothers. Science 1959, 130, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Harlow, H.F.; Dodsworth, R.O.; Harlow, M.K. Total Social Isolation in Monkeys. Proc. Natl. Acad. Sci. USA 1965, 54, 90–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of Stress throughout the Lifespan on the Brain, Behaviour and Cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef]
- Sacks, R.M.; Takemoto, E.; Andrea, S.; Dieckmann, N.F.; Bauer, K.W.; Boone-Heinonen, J. Childhood Maltreatment and BMI Trajectory: The Mediating Role of Depression. Am. J. Prev. Med. 2017, 53, 625–633. [Google Scholar] [CrossRef]
- Gershon, A.; Hayward, C.; Schraedley-Desmond, P.; Rudolph, K.D.; Booster, G.D.; Gotlib, I.H. Life Stress and First Onset of Psychiatric Disorders in Daughters of Depressed Mothers. J. Psychiatr. Res. 2011, 45, 855–862. [Google Scholar] [CrossRef] [Green Version]
- Orso, R.; Creutzberg, K.C.; Wearick-Silva, L.E.; Wendt Viola, T.; Tractenberg, S.G.; Benetti, F.; Grassi-Oliveira, R. How Early Life Stress Impact Maternal Care: A Systematic Review of Rodent Studies. Front. Behav. Neurosci. 2019, 13, 197. [Google Scholar] [CrossRef]
- Norman, R.E.; Byambaa, M.; De, R.; Butchart, A.; Scott, J.; Vos, T. The Long-Term Health Consequences of Child Physical Abuse, Emotional Abuse, and Neglect: A Systematic Review and Meta-Analysis. PLoS Med. 2012, 9, e1001349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teicher, M.H.; Andersen, S.L.; Polcari, A.; Anderson, C.M.; Navalta, C.P.; Kim, D.M. The Neurobiological Consequences of Early Stress and Childhood Maltreatment. Neurosci. Biobehav. Rev. 2003, 27, 33–44. [Google Scholar] [CrossRef]
- Pechtel, P.; Pizzagalli, D.A. Effects of Early Life Stress on Cognitive and Affective Function: An Integrated Review of Human Literature. Psychopharmacology 2011, 214, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Davey, C.G.; Yücel, M.; Allen, N.B. The Emergence of Depression in Adolescence: Development of the Prefrontal Cortex and the Representation of Reward. Neurosci. Biobehav. Rev. 2008, 32, 1–19. [Google Scholar] [CrossRef]
- Abrahám, H.; Vincze, A.; Jewgenow, I.; Veszprémi, B.; Kravják, A.; Gömöri, E.; Seress, L. Myelination in the Human Hippocampal Formation from Midgestation to Adulthood. Int. J. Dev. Neurosci. 2010, 28, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Diorio, D.; Viau, V.; Meaney, M.J. The Role of the Medial Prefrontal Cortex (Cingulate Gyrus) in the Regulation of Hypothalamic-Pituitary-Adrenal Responses to Stress. J. Neurosci. 1993, 13, 3839–3847. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.D.; Lopez, J.F.; Lyons, D.M.; Burke, S.; Wallace, M.; Schatzberg, A.F. Glucocorticoid and Mineralocorticoid Receptor MRNA Expression in Squirrel Monkey Brain. J. Psychiatr. Res. 2000, 34, 383–392. [Google Scholar] [CrossRef]
- Anacker, C.; Zunszain, P.A.; Carvalho, L.A.; Pariante, C.M. The Glucocorticoid Receptor: Pivot of Depression and of Antidepressant Treatment? Psychoneuroendocrinology 2011, 36, 415–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, S.; Wiener, S.G. Psychoendocrine Aspects of Mother-Infant Relationships in Nonhuman Primates. Psychoneuroendocrinology 1988, 13, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Anisman, H.; Zaharia, M.D.; Meaney, M.J.; Merali, Z. Do Early-Life Events Permanently Alter Behavioral and Hormonal Responses to Stressors? Int. J. Dev. Neurosci. 1998, 16, 149–164. [Google Scholar] [CrossRef]
- Liu, D.; Diorio, J.; Tannenbaum, B.; Caldji, C.; Francis, D.; Freedman, A.; Sharma, S.; Pearson, D.; Plotsky, P.M.; Meaney, M.J. Maternal Care, Hippocampal Glucocorticoid Receptors, and Hypothalamic-Pituitary-Adrenal Responses to Stress. Science 1997, 277, 1659–1662. [Google Scholar] [CrossRef] [Green Version]
- Numakawa, T.; Odaka, H.; Adachi, N. Actions of Brain-Derived Neurotrophic Factor and Glucocorticoid Stress in Neurogenesis. Int. J. Mol. Sci. 2017, 18, 2312. [Google Scholar] [CrossRef] [Green Version]
- Weaver, I.C.G.; Meaney, M.J.; Szyf, M. Maternal Care Effects on the Hippocampal Transcriptome and Anxiety-Mediated Behaviors in the Offspring That Are Reversible in Adulthood. Proc. Natl. Acad. Sci. USA 2006, 103, 3480–3485. [Google Scholar] [CrossRef] [Green Version]
- Champagne, F.A.; Meaney, M.J. Stress during Gestation Alters Postpartum Maternal Care and the Development of the Offspring in a Rodent Model. Biol. Psychiatry 2006, 59, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Maestripieri, D.; McCormack, K.; Lindell, S.G.; Higley, J.D.; Sanchez, M.M. Influence of Parenting Style on the Offspring’s Behaviour and CSF Monoamine Metabolite Levels in Crossfostered and Noncrossfostered Female Rhesus Macaques. Behav. Brain Res. 2006, 175, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Fahlke, C.; Lorenz, J.G.; Long, J.; Champoux, M.; Suomi, S.J.; Higley, J.D. Rearing Experiences and Stress-Induced Plasma Cortisol as Early Risk Factors for Excessive Alcohol Consumption in Nonhuman Primates. Alcohol. Clin. Exp. Res. 2000, 24, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Coplan, J.D.; Andrews, M.W.; Rosenblum, L.A.; Owens, M.J.; Friedman, S.; Gorman, J.M.; Nemeroff, C.B. Persistent Elevations of Cerebrospinal Fluid Concentrations of Corticotropin-Releasing Factor in Adult Nonhuman Primates Exposed to Early-Life Stressors: Implications for the Pathophysiology of Mood and Anxiety Disorders. Proc. Natl. Acad. Sci. USA 1996, 93, 1619–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danesch, U.; Hashimoto, S.; Renkawitz, R.; Schütz, G. Transcriptional Regulation of the Tryptophan Oxygenase Gene in Rat Liver by Glucocorticoids. J. Biol. Chem. 1983, 258, 4750–4753. [Google Scholar] [CrossRef]
- Nakamura, T.; Niimi, S.; Nawa, K.; Noda, C.; Ichihara, A.; Takagi, Y.; Anai, M.; Sakaki, Y. Multihormonal Regulation of Transcription of the Tryptophan 2,3-Dioxygenase Gene in Primary Cultures of Adult Rat Hepatocytes with Special Reference to the Presence of a Transcriptional Protein Mediating the Action of Glucocorticoids. J. Biol. Chem. 1987, 262, 727–733. [Google Scholar] [CrossRef]
- Coplan, J.D.; George, R.; Syed, S.A.; Rozenboym, A.V.; Tang, J.E.; Fulton, S.L.; Perera, T.D. Early Life Stress and the Fate of Kynurenine Pathway Metabolites. Front. Hum. Neurosci. 2021, 15, 636144. [Google Scholar] [CrossRef]
- Hohmann, C.F.; Hamon, R.; Batshaw, M.L.; Coyle, J.T. Transient Postnatal Elevation of Serotonin Levels in Mouse Neocortex. Brain Res. 1988, 471, 163–166. [Google Scholar] [CrossRef]
- Rebello, T.J.; Yu, Q.; Goodfellow, N.M.; Caffrey Cagliostro, M.K.; Teissier, A.; Morelli, E.; Demireva, E.Y.; Chemiakine, A.; Rosoklija, G.B.; Dwork, A.J.; et al. Postnatal Day 2 to 11 Constitutes a 5-HT-Sensitive Period Impacting Adult MPFC Function. J. Neurosci. 2014, 34, 12379–12393. [Google Scholar] [CrossRef] [Green Version]
- Courtiol, E.; Wilson, D.A.; Shah, R.; Sullivan, R.M.; Teixeira, C.M. Maternal Regulation of Pups’ Cortical Activity: Role of Serotonergic Signaling. eNeuro 2018, 5, ENEURO.0093-18.2018. [Google Scholar] [CrossRef] [Green Version]
- Bhansali, P.; Dunning, J.; Singer, S.E.; David, L.; Schmauss, C. Early Life Stress Alters Adult Serotonin 2C Receptor Pre-MRNA Editing and Expression of the Alpha Subunit of the Heterotrimeric G-Protein G q. J. Neurosci. 2007, 27, 1467–1473. [Google Scholar] [CrossRef] [Green Version]
- Bremner, J.D.; Randall, P.; Vermetten, E.; Staib, L.; Bronen, R.A.; Mazure, C.; Capelli, S.; McCarthy, G.; Innis, R.B.; Charney, D.S. Magnetic Resonance Imaging-Based Measurement of Hippocampal Volume in Posttraumatic Stress Disorder Related to Childhood Physical and Sexual Abuse—A Preliminary Report. Biol. Psychiatry 1997, 41, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Driessen, M.; Herrmann, J.; Stahl, K.; Zwaan, M.; Meier, S.; Hill, A.; Osterheider, M.; Petersen, D. Magnetic Resonance Imaging Volumes of the Hippocampus and the Amygdala in Women with Borderline Personality Disorder and Early Traumatization. Arch. Gen. Psychiatry 2000, 57, 1115–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buss, C.; Lord, C.; Wadiwalla, M.; Hellhammer, D.H.; Lupien, S.J.; Meaney, M.J.; Pruessner, J.C. Maternal Care Modulates the Relationship between Prenatal Risk and Hippocampal Volume in Women but Not in Men. J. Neurosci. 2007, 27, 2592–2595. [Google Scholar] [CrossRef] [Green Version]
- Hart, H.; Rubia, K. Neuroimaging of Child Abuse: A Critical Review. Front. Hum. Neurosci. 2012, 6, 52. [Google Scholar] [CrossRef] [Green Version]
- Tyborowska, A.; Volman, I.; Niermann, H.C.M.; Pouwels, J.L.; Smeekens, S.; Cillessen, A.H.N.; Toni, I.; Roelofs, K. Early-Life and Pubertal Stress Differentially Modulate Grey Matter Development in Human Adolescents. Sci. Rep. 2018, 8, 9201. [Google Scholar] [CrossRef] [PubMed]
- Heyn, S.A.; Keding, T.J.; Ross, M.C.; Cisler, J.M.; Mumford, J.A.; Herringa, R.J. Abnormal Prefrontal Development in Pediatric Posttraumatic Stress Disorder: A Longitudinal Structural and Functional Magnetic Resonance Imaging Study. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2019, 4, 171–179. [Google Scholar] [CrossRef]
- Underwood, M.D.; Bakalian, M.J.; Escobar, T.; Kassir, S.; Mann, J.J.; Arango, V. Early-Life Adversity, but Not Suicide, Is Associated with Less Prefrontal Cortex Gray Matter in Adulthood. Int. J. Neuropsychopharmacol. 2019, 22, 349–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrion, V.G.; Weems, C.F.; Eliez, S.; Patwardhan, A.; Brown, W.; Ray, R.D.; Reiss, A.L. Attenuation of Frontal Asymmetry in Pediatric Posttraumatic Stress Disorder. Biol. Psychiatry 2001, 50, 943–951. [Google Scholar] [CrossRef]
- Geuze, E.; Vermetten, E.; Bremner, J.D. MR-Based in Vivo Hippocampal Volumetrics: 2. Findings in Neuropsychiatric Disorders. Mol. Psychiatry 2005, 10, 160–184. [Google Scholar] [CrossRef] [Green Version]
- De Bellis, M.D.; Keshavan, M.S.; Clark, D.B.; Casey, B.J.; Giedd, J.N.; Boring, A.M.; Frustaci, K.; Ryan, N.D. A.E. Bennett Research Award. Developmental Traumatology. Part II: Brain Development. Biol. Psychiatry 1999, 45, 1271–1284. [Google Scholar] [CrossRef] [PubMed]
- Bernard-Bonnin, A.-C. Maternal Depression and Child Development. Paediatr. Child Health 2004, 9, 575–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupien, S.J.; King, S.; Meaney, M.J.; McEwen, B.S. Child’s Stress Hormone Levels Correlate with Mother’s Socioeconomic Status and Depressive State. Biol. Psychiatry 2000, 48, 976–980. [Google Scholar] [CrossRef] [PubMed]
- Halligan, S.L.; Herbert, J.; Goodyer, I.M.; Murray, L. Exposure to Postnatal Depression Predicts Elevated Cortisol in Adolescent Offspring. Biol. Psychiatry 2004, 55, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Halligan, S.L.; Herbert, J.; Goodyer, I.; Murray, L. Disturbances in Morning Cortisol Secretion in Association with Maternal Postnatal Depression Predict Subsequent Depressive Symptomatology in Adolescents. Biol. Psychiatry 2007, 62, 40–46. [Google Scholar] [CrossRef]
- Cogill, S.R.; Caplan, H.L.; Alexandra, H.; Robson, K.M.; Kumar, R. Impact of Maternal Postnatal Depression on Cognitive Development of Young Children. Br. Med. J. Clin. Res. Ed. 1986, 292, 1165–1167. [Google Scholar] [CrossRef] [Green Version]
- Caplan, H.L.; Cogill, S.R.; Alexandra, H.; Robson, K.M.; Katz, R.; Kumar, R. Maternal Depression and the Emotional Development of the Child. Br. J. Psychiatry 1989, 154, 818–822. [Google Scholar] [CrossRef]
- Madigan, S.; Oatley, H.; Racine, N.; Fearon, R.M.P.; Schumacher, L.; Akbari, E.; Cooke, J.E.; Tarabulsy, G.M. A Meta-Analysis of Maternal Prenatal Depression and Anxiety on Child Socioemotional Development. J. Am. Acad. Child Adolesc. Psychiatry 2018, 57, 645–657.e8. [Google Scholar] [CrossRef]
- Canetti, L.; Bachar, E.; Galili-Weisstub, E.; De-Nour, A.K.; Shalev, A.Y. Parental Bonding and Mental Health in Adolescence. Adolescence 1997, 32, 381–394. [Google Scholar]
- Meaney, M.J. Maternal Care, Gene Expression, and the Transmission of Individual Differences in Stress Reactivity across Generations. Annu. Rev. Neurosci. 2001, 24, 1161–1192. [Google Scholar] [CrossRef]
- Jensen Peña, C.; Champagne, F.A. Implications of Temporal Variation in Maternal Care for the Prediction of Neurobiological and Behavioral Outcomes in Offspring. Behav. Neurosci. 2013, 127, 33–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, I.C.G.; Cervoni, N.; Champagne, F.A.; D’Alessio, A.C.; Sharma, S.; Seckl, J.R.; Dymov, S.; Szyf, M.; Meaney, M.J. Epigenetic Programming by Maternal Behavior. Nat. Neurosci. 2004, 7, 847–854. [Google Scholar] [CrossRef]
- Champagne, F.A. Epigenetic Mechanisms and the Transgenerational Effects of Maternal Care. Front. Neuroendocrinol. 2008, 29, 386–397. [Google Scholar] [CrossRef] [Green Version]
- Dowdney, L.; Skuse, D.; Rutter, M.; Quinton, D.; Mrazek, D. The Nature and Qualities of Parenting Provided by Women Raised in Institutions. J. Child Psychol. Psychiatry 1985, 26, 599–625. [Google Scholar] [CrossRef] [PubMed]
- Bertino, M. Effects of High Fat, Protein Supplemented Diets on Maternal Behavior in Rats. Physiol. Behav. 1982, 29, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Franks, B.; Curley, J.P.; Champagne, F.A. Measuring Variations in Maternal Behavior: Relevance for Studies of Mood and Anxiety. In Mood and Anxiety Related Phenotypes in Mice: Characterization Using Behavioral Tests; Gould, T.D., Ed.; Neuromethods; Humana Press: Totowa, NJ, USA, 2011; Volume II, pp. 209–224. [Google Scholar] [CrossRef] [Green Version]
- Purcell, R.H.; Sun, B.; Pass, L.L.; Power, M.L.; Moran, T.H.; Tamashiro, K.L.K. Maternal Stress and High-Fat Diet Effect on Maternal Behavior, Milk Composition, and Pup Ingestive Behavior. Physiol. Behav. 2011, 104, 474–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Xu, S.; Lin, L.; Gu, X.; Fu, C.; Fang, Y.; Li, X.; Wang, X. High-Fat Diet Mediates Anxiolytic-like Behaviors in a Time-Dependent Manner Through the Regulation of SIRT1 in the Brain. Neuroscience 2018, 372, 237–245. [Google Scholar] [CrossRef]
- John, R.M. Prenatal Adversity Modulates the Quality of Maternal Care Via the Exposed Offspring. Bioessays 2019, 41, e1900025. [Google Scholar] [CrossRef]
- Champagne, F.A.; Francis, D.D.; Mar, A.; Meaney, M.J. Variations in Maternal Care in the Rat as a Mediating Influence for the Effects of Environment on Development. Physiol. Behav. 2003, 79, 359–371. [Google Scholar] [CrossRef]
- Latham, N.; Mason, G. From House Mouse to Mouse House: The Behavioural Biology of Free-Living Mus Musculus and Its Implications in the Laboratory. Appl. Anim. Behav. Sci. 2004, 86, 261–289. [Google Scholar] [CrossRef] [Green Version]
- Silva, D.F.O.; Cobucci, R.N.; Gonçalves, A.K.; Lima, S.C.V.C. Systematic Review of the Association between Dietary Patterns and Perinatal Anxiety and Depression. BMC Pregnancy Childbirth 2019, 19, 212. [Google Scholar] [CrossRef] [Green Version]
- Okubo, H.; Miyake, Y.; Sasaki, S.; Tanaka, K.; Murakami, K.; Hirota, Y.; Osaka Maternal and Child Health Study Group. Dietary Patterns during Pregnancy and the Risk of Postpartum Depression in Japan: The Osaka Maternal and Child Health Study. Br. J. Nutr. 2011, 105, 1251–1257. [Google Scholar] [CrossRef] [Green Version]
- Northstone, K.; Joinson, C.; Emmett, P. Dietary Patterns and Depressive Symptoms in a UK Cohort of Men and Women: A Longitudinal Study. Public Health Nutr. 2018, 21, 831–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, M.C.; Isolauri, E.; Laitinen, K.; Salminen, S. Distinct Composition of Gut Microbiota during Pregnancy in Overweight and Normal-Weight Women. Am. J. Clin. Nutr. 2008, 88, 894–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santacruz, A.; Collado, M.C.; García-Valdés, L.; Segura, M.T.; Martín-Lagos, J.A.; Anjos, T.; Martí-Romero, M.; Lopez, R.M.; Florido, J.; Campoy, C.; et al. Gut Microbiota Composition Is Associated with Body Weight, Weight Gain and Biochemical Parameters in Pregnant Women. Br. J. Nutr. 2010, 104, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.M.; Antony, K.M.; Ma, J.; Prince, A.L.; Showalter, L.; Moller, M.; Aagaard, K.M. The Early Infant Gut Microbiome Varies in Association with a Maternal High-Fat Diet. Genome Med. 2016, 8, 77. [Google Scholar] [CrossRef] [Green Version]
- Paul, H.A.; Bomhof, M.R.; Vogel, H.J.; Reimer, R.A. Diet-Induced Changes in Maternal Gut Microbiota and Metabolomic Profiles Influence Programming of Offspring Obesity Risk in Rats. Sci. Rep. 2016, 6, 20683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tochitani, S.; Ikeno, T.; Ito, T.; Sakurai, A.; Yamauchi, T.; Matsuzaki, H. Administration of Non-Absorbable Antibiotics to Pregnant Mice to Perturb the Maternal Gut Microbiota Is Associated with Alterations in Offspring Behavior. PLoS ONE 2016, 11, e0138293. [Google Scholar] [CrossRef]
- de Goffau, M.C.; Lager, S.; Sovio, U.; Gaccioli, F.; Cook, E.; Peacock, S.J.; Parkhill, J.; Charnock-Jones, D.S.; Smith, G.C.S. Human Placenta Has No Microbiome but Can Contain Potential Pathogens. Nature 2019, 572, 329–334. [Google Scholar] [CrossRef]
- Macpherson, A.J.; de Agüero, M.G.; Ganal-Vonarburg, S.C. How Nutrition and the Maternal Microbiota Shape the Neonatal Immune System. Nat. Rev. Immunol. 2017, 17, 508–517. [Google Scholar] [CrossRef]
- Jašarević, E.; Howerton, C.L.; Howard, C.D.; Bale, T.L. Alterations in the Vaginal Microbiome by Maternal Stress Are Associated with Metabolic Reprogramming of the Offspring Gut and Brain. Endocrinology 2015, 156, 3265–3276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuong, H.E.; Pronovost, G.N.; Williams, D.W.; Coley, E.J.L.; Siegler, E.L.; Qiu, A.; Kazantsev, M.; Wilson, C.J.; Rendon, T.; Hsiao, E.Y. The Maternal Microbiome Modulates Fetal Neurodevelopment in Mice. Nature 2020, 586, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Yu, X.-N.; Kubo, C.; Koga, Y. Postnatal Microbial Colonization Programs the Hypothalamic-Pituitary-Adrenal System for Stress Response in Mice. J. Physiol. 2004, 558 Pt 1, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Soderborg, T.K.; Clark, S.E.; Mulligan, C.E.; Janssen, R.C.; Babcock, L.; Ir, D.; Young, B.; Krebs, N.; Lemas, D.J.; Johnson, L.K.; et al. The Gut Microbiota in Infants of Obese Mothers Increases Inflammation and Susceptibility to NAFLD. Nat. Commun. 2018, 9, 4462. [Google Scholar] [CrossRef] [Green Version]
- Aye, I.L.M.H.; Rosario, F.J.; Powell, T.L.; Jansson, T. Adiponectin Supplementation in Pregnant Mice Prevents the Adverse Effects of Maternal Obesity on Placental Function and Fetal Growth. Proc. Natl. Acad. Sci. USA 2015, 112, 12858–12863. [Google Scholar] [CrossRef] [Green Version]
- Cheung, S.G.; Goldenthal, A.R.; Uhlemann, A.-C.; Mann, J.J.; Miller, J.M.; Sublette, M.E. Systematic Review of Gut Microbiota and Major Depression. Front. Psychiatry 2019, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus Reuteri in Human Health and Diseases. Front. Microbiol. 2018, 9, 757. [Google Scholar] [CrossRef] [Green Version]
- Bruce-Keller, A.J.; Fernandez-Kim, S.-O.; Townsend, R.L.; Kruger, C.; Carmouche, R.; Newman, S.; Salbaum, J.M.; Berthoud, H.-R. Maternal Obese-Type Gut Microbiota Differentially Impact Cognition, Anxiety and Compulsive Behavior in Male and Female Offspring in Mice. PLoS ONE 2017, 12, e0175577. [Google Scholar] [CrossRef] [Green Version]
- Bruce-Keller, A.J.; Salbaum, J.M.; Luo, M.; Blanchard, E.; Taylor, C.M.; Welsh, D.A.; Berthoud, H.-R. Obese-Type Gut Microbiota Induce Neurobehavioral Changes in the Absence of Obesity. Biol. Psychiatry 2015, 77, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Round, J.L.; Mazmanian, S.K. The Gut Microbiota Shapes Intestinal Immune Responses during Health and Disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef]
- Lee, S.; Kim, H.; You, G.; Kim, Y.-M.; Lee, S.; Le, V.-H.; Kwon, O.; Im, S.-H.; Kim, Y.-M.; Kim, K.S.; et al. Bone Marrow CX3CR1+ Mononuclear Cells Relay a Systemic Microbiota Signal to Control Hematopoietic Progenitors in Mice. Blood 2019, 134, 1312–1322. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Chen, G.-L.; Hannemann, N.; Ipseiz, N.; Krönke, G.; Bäuerle, T.; Munos, L.; Wirtz, S.; Schett, G.; Bozec, A. Microbiota from Obese Mice Regulate Hematopoietic Stem Cell Differentiation by Altering the Bone Niche. Cell Metab. 2015, 22, 886–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manicone, A.M.; Gong, K.; Johnston, L.K.; Giannandrea, M. Diet-Induced Obesity Alters Myeloid Cell Populations in Naïve and Injured Lung. Respir. Res. 2016, 17, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strekalova, T.; Evans, M.; Costa-Nunes, J.; Bachurin, S.; Yeritsyan, N.; Couch, Y.; Steinbusch, H.M.W.; Eleonore Köhler, S.; Lesch, K.-P.; Anthony, D.C. Tlr4 Upregulation in the Brain Accompanies Depression- and Anxiety-like Behaviors Induced by a High-Cholesterol Diet. Brain Behav. Immun. 2015, 48, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.L.; Buonomo, E.; Carey, M.; Cowardin, C.; Naylor, C.; Noor, Z.; Wills-Karp, M.; Petri, W.A. Bone Marrow Dendritic Cells from Mice with an Altered Microbiota Provide Interleukin 17A-Dependent Protection against Entamoeba Histolytica Colitis. mBio 2014, 5, e01817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Mantrana, I.; Selma-Royo, M.; González, S.; Parra-Llorca, A.; Martínez-Costa, C.; Collado, M.C. Distinct Maternal Microbiota Clusters Are Associated with Diet during Pregnancy: Impact on Neonatal Microbiota and Infant Growth during the First 18 Months of Life. Gut Microbes 2020, 11, 962–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schachter, J.; Martel, J.; Lin, C.-S.; Chang, C.-J.; Wu, T.-R.; Lu, C.-C.; Ko, Y.-F.; Lai, H.-C.; Ojcius, D.M.; Young, J.D. Effects of Obesity on Depression: A Role for Inflammation and the Gut Microbiota. Brain Behav. Immun. 2018, 69, 1–8. [Google Scholar] [CrossRef]
- Schneeberger, M.; Everard, A.; Gómez-Valadés, A.G.; Matamoros, S.; Ramírez, S.; Delzenne, N.M.; Gomis, R.; Claret, M.; Cani, P.D. Akkermansia Muciniphila Inversely Correlates with the Onset of Inflammation, Altered Adipose Tissue Metabolism and Metabolic Disorders during Obesity in Mice. Sci. Rep. 2015, 5, 16643. [Google Scholar] [CrossRef] [Green Version]
- Gohir, W.; Kennedy, K.M.; Wallace, J.G.; Saoi, M.; Bellissimo, C.J.; Britz-McKibbin, P.; Petrik, J.J.; Surette, M.G.; Sloboda, D.M. High-Fat Diet Intake Modulates Maternal Intestinal Adaptations to Pregnancy and Results in Placental Hypoxia, as Well as Altered Fetal Gut Barrier Proteins and Immune Markers. J. Physiol. 2019, 597, 3029–3051. [Google Scholar] [CrossRef]
- Mlakar, J.; Korva, M.; Tul, N.; Popović, M.; Poljšak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Resman Rus, K.; Vesnaver Vipotnik, T.; Fabjan Vodušek, V.; et al. Zika Virus Associated with Microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [CrossRef]
- Stanelle-Bertram, S.; Walendy-Gnirß, K.; Speiseder, T.; Thiele, S.; Asante, I.A.; Dreier, C.; Kouassi, N.M.; Preuß, A.; Pilnitz-Stolze, G.; Müller, U.; et al. Male Offspring Born to Mildly ZIKV-Infected Mice Are at Risk of Developing Neurocognitive Disorders in Adulthood. Nat. Microbiol. 2018, 3, 1161–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estes, M.L.; McAllister, A.K. Maternal Immune Activation: Implications for Neuropsychiatric Disorders. Science 2016, 353, 772–777. [Google Scholar] [CrossRef] [Green Version]
- Winter, C.; Djodari-Irani, A.; Sohr, R.; Morgenstern, R.; Feldon, J.; Juckel, G.; Meyer, U. Prenatal Immune Activation Leads to Multiple Changes in Basal Neurotransmitter Levels in the Adult Brain: Implications for Brain Disorders of Neurodevelopmental Origin Such as Schizophrenia. Int. J. Neuropsychopharmacol. 2009, 12, 513–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronovsky, M.; Berger, S.; Molz, B.; Berger, A.; Pollak, D.D. Animal Models of Maternal Immune Activation in Depression Research. Curr. Neuropharmacol. 2016, 14, 688–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simanek, A.M.; Meier, H.C.S. Association Between Prenatal Exposure to Maternal Infection and Offspring Mood Disorders: A Review of the Literature. Curr. Probl. Pediatr. Adolesc. Health Care 2015, 45, 325–364. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yan, J.-Y.; Lo, Y.-K.; Carvey, P.M.; Ling, Z. Dopaminergic and Serotoninergic Deficiencies in Young Adult Rats Prenatally Exposed to the Bacterial Lipopolysaccharide. Brain Res. 2009, 1265, 196–204. [Google Scholar] [CrossRef]
- Babri, S.; Doosti, M.-H.; Salari, A.-A. Strain-Dependent Effects of Prenatal Maternal Immune Activation on Anxiety- and Depression-like Behaviors in Offspring. Brain Behav. Immun. 2014, 37, 164–176. [Google Scholar] [CrossRef]
- Khan, D.; Fernando, P.; Cicvaric, A.; Berger, A.; Pollak, A.; Monje, F.J.; Pollak, D.D. Long-Term Effects of Maternal Immune Activation on Depression-like Behavior in the Mouse. Transl. Psychiatry 2014, 4, e363. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-L.; Wang, S. Prenatal Lipopolysaccharide Exposure Increases Depression-like Behaviors and Reduces Hippocampal Neurogenesis in Adult Rats. Behav. Brain Res. 2014, 259, 24–34. [Google Scholar] [CrossRef]
- Depino, A.M. Early Prenatal Exposure to LPS Results in Anxiety- and Depression-Related Behaviors in Adulthood. Neuroscience 2015, 299, 56–65. [Google Scholar] [CrossRef]
- Estes, M.L.; McAllister, A.K. Immune Mediators in the Brain and Peripheral Tissues in Autism Spectrum Disorder. Nat. Rev. Neurosci. 2015, 16, 469–486. [Google Scholar] [CrossRef] [Green Version]
- Das, U.N. Is Obesity an Inflammatory Condition? Nutrition 2001, 17, 953–966. [Google Scholar] [CrossRef]
- Sureshchandra, S.; Marshall, N.E.; Messaoudi, I. Impact of Pregravid Obesity on Maternal and Fetal Immunity: Fertile Grounds for Reprogramming. J. Leukoc. Biol. 2019, 106, 1035–1050. [Google Scholar] [CrossRef] [PubMed]
- Pendeloski, K.P.T.; Ono, E.; Torloni, M.R.; Mattar, R.; Daher, S. Maternal Obesity and Inflammatory Mediators: A Controversial Association. Am. J. Reprod. Immunol. 2017, 77, e12674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challier, J.C.; Basu, S.; Bintein, T.; Minium, J.; Hotmire, K.; Catalano, P.M.; Hauguel-de Mouzon, S. Obesity in Pregnancy Stimulates Macrophage Accumulation and Inflammation in the Placenta. Placenta 2008, 29, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Saben, J.; Zhong, Y.; Gomez-Acevedo, H.; Thakali, K.M.; Borengasser, S.J.; Andres, A.; Shankar, K. Early Growth Response Protein-1 Mediates Lipotoxicity-Associated Placental Inflammation: Role in Maternal Obesity. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E1–E14. [Google Scholar] [CrossRef] [PubMed]
- Frias, A.E.; Morgan, T.K.; Evans, A.E.; Rasanen, J.; Oh, K.Y.; Thornburg, K.L.; Grove, K.L. Maternal High-Fat Diet Disturbs Uteroplacental Hemodynamics and Increases the Frequency of Stillbirth in a Nonhuman Primate Model of Excess Nutrition. Endocrinology 2011, 152, 2456–2464. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, E.Y.; Patterson, P.H. Activation of the Maternal Immune System Induces Endocrine Changes in the Placenta via IL-6. Brain Behav. Immun. 2011, 25, 604–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, F.M.; Freeman, D.J.; Ramsay, J.E.; Greer, I.A.; Caslake, M.; Ferrell, W.R. Longitudinal Assessment of Maternal Endothelial Function and Markers of Inflammation and Placental Function throughout Pregnancy in Lean and Obese Mothers. J. Clin. Endocrinol. Metab. 2007, 92, 969–975. [Google Scholar] [CrossRef] [Green Version]
- Kretschmer, T.; Schulze-Edinghausen, M.; Turnwald, E.-M.; Janoschek, R.; Bae-Gartz, I.; Zentis, P.; Handwerk, M.; Wohlfarth, M.; Schauss, A.; Hucklenbruch-Rother, E.; et al. Effect of Maternal Obesity in Mice on IL-6 Levels and Placental Endothelial Cell Homeostasis. Nutrients 2020, 12, 296. [Google Scholar] [CrossRef] [Green Version]
- Zaretsky, M.V.; Alexander, J.M.; Byrd, W.; Bawdon, R.E. Transfer of Inflammatory Cytokines across the Placenta. Obstet. Gynecol. 2004, 103, 546–550. [Google Scholar] [CrossRef] [PubMed]
- Hrolfsdottir, L.; Schalkwijk, C.G.; Birgisdottir, B.E.; Gunnarsdottir, I.; Maslova, E.; Granström, C.; Strøm, M.; Olsen, S.F.; Halldorsson, T.I. Maternal Diet, Gestational Weight Gain, and Inflammatory Markers during Pregnancy. Obesity 2016, 24, 2133–2139. [Google Scholar] [CrossRef]
- Bay-Richter, C.; Janelidze, S.; Hallberg, L.; Brundin, L. Changes in Behaviour and Cytokine Expression upon a Peripheral Immune Challenge. Behav. Brain Res. 2011, 222, 193–199. [Google Scholar] [CrossRef]
- Lasselin, J.; Elsenbruch, S.; Lekander, M.; Axelsson, J.; Karshikoff, B.; Grigoleit, J.-S.; Engler, H.; Schedlowski, M.; Benson, S. Mood Disturbance during Experimental Endotoxemia: Predictors of State Anxiety as a Psychological Component of Sickness Behavior. Brain Behav. Immun. 2016, 57, 30–37. [Google Scholar] [CrossRef]
- Dalman, C.; Allebeck, P.; Gunnell, D.; Harrison, G.; Kristensson, K.; Lewis, G.; Lofving, S.; Rasmussen, F.; Wicks, S.; Karlsson, H. Infections in the CNS during Childhood and the Risk of Subsequent Psychotic Illness: A Cohort Study of More than One Million Swedish Subjects. Am. J. Psychiatry 2008, 165, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Abrahao, A.L.; Focaccia, R.; Gattaz, W.F. Childhood Meningitis Increases the Risk for Adult Schizophrenia. World J. Biol. Psychiatry 2005, 6 (Suppl. S2), 44–48. [Google Scholar] [CrossRef] [PubMed]
- Blomström, Å.; Karlsson, H.; Svensson, A.; Frisell, T.; Lee, B.K.; Dal, H.; Magnusson, C.; Dalman, C. Hospital Admission with Infection during Childhood and Risk for Psychotic Illness--a Population-Based Cohort Study. Schizophr. Bull. 2014, 40, 1518–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronaldson, A.; Arias de la Torre, J.; Sima, R.; Ashworth, M.; Armstrong, D.; Bakolis, I.; Hotopf, M.; Dregan, A. Prospective Associations between Depression and Risk of Hospitalisation for Infection: Findings from the UK Biobank. Brain Behav. Immun. 2022, 102, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Köhler-Forsberg, O.; Petersen, L.; Gasse, C.; Mortensen, P.B.; Dalsgaard, S.; Yolken, R.H.; Mors, O.; Benros, M.E. A Nationwide Study in Denmark of the Association Between Treated Infections and the Subsequent Risk of Treated Mental Disorders in Children and Adolescents. JAMA Psychiatry 2019, 76, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Yates, A.G.; Weglinski, C.M.; Ying, Y.; Dunstan, I.K.; Strekalova, T.; Anthony, D.C. Nafamostat Reduces Systemic Inflammation in TLR7-Mediated Virus-like Illness. J. Neuroinflamm. 2022, 19, 8. [Google Scholar] [CrossRef]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From Inflammation to Sickness and Depression: When the Immune System Subjugates the Brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammen, C. Stress and Depression. Annu. Rev. Clin. Psychol. 2005, 1, 293–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisman, H.; Merali, Z. Cytokines, Stress and Depressive Illness: Brain-Immune Interactions. Ann. Med. 2003, 35, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Kiecolt-Glaser, J.K.; Preacher, K.J.; MacCallum, R.C.; Atkinson, C.; Malarkey, W.B.; Glaser, R. Chronic Stress and Age-Related Increases in the Proinflammatory Cytokine IL-6. Proc. Natl. Acad. Sci. USA 2003, 100, 9090–9095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wohleb, E.S.; Delpech, J.-C. Dynamic Cross-Talk between Microglia and Peripheral Monocytes Underlies Stress-Induced Neuroinflammation and Behavioral Consequences. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 79 Pt A, 40–48. [Google Scholar] [CrossRef]
- Munshi, S.; Loh, M.K.; Ferrara, N.; DeJoseph, M.R.; Ritger, A.; Padival, M.; Record, M.J.; Urban, J.H.; Rosenkranz, J.A. Repeated Stress Induces a Pro-Inflammatory State, Increases Amygdala Neuronal and Microglial Activation, and Causes Anxiety in Adult Male Rats. Brain Behav. Immun. 2020, 84, 180–199. [Google Scholar] [CrossRef]
- Kowalska, K.; Pasinska, P.; Klimiec-Moskal, E.; Pera, J.; Slowik, A.; Klimkowicz-Mrowiec, A.; Dziedzic, T. C-Reactive Protein and Post-Stroke Depressive Symptoms. Sci. Rep. 2020, 10, 1431. [Google Scholar] [CrossRef] [Green Version]
- Roman, M.; Irwin, M.R. Novel Neuroimmunologic Therapeutics in Depression: A Clinical Perspective on What We Know so Far. Brain Behav. Immun. 2020, 83, 7–21. [Google Scholar] [CrossRef]
- Liu, J.J.; Wei, Y.B.; Strawbridge, R.; Bao, Y.; Chang, S.; Shi, L.; Que, J.; Gadad, B.S.; Trivedi, M.H.; Kelsoe, J.R.; et al. Peripheral Cytokine Levels and Response to Antidepressant Treatment in Depression: A Systematic Review and Meta-Analysis. Mol. Psychiatry 2020, 25, 339–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohmann, C.F.; Odebode, G.; Naidu, L.; Koban, M. Early Life Stress Alters Adult Inflammatory Responses in a Mouse Model for Depression. Ann. Psychiatry Ment. Health 2017, 5, 1095. [Google Scholar] [PubMed]
- Bolton, J.L.; Bilbo, S.D. Developmental Programming of Brain and Behavior by Perinatal Diet: Focus on Inflammatory Mechanisms. Dialogues Clin. Neurosci. 2014, 16, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.; Petri, W.A. Microglia: Immune Regulators of Neurodevelopment. Front. Immunol. 2018, 9, 2576. [Google Scholar] [CrossRef] [Green Version]
- Schafer, D.P.; Stevens, B. Microglia Function in Central Nervous System Development and Plasticity. Cold Spring Harb. Perspect. Biol. 2015, 7, a020545. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Barres, B.A. Microglia and Macrophages in Brain Homeostasis and Disease. Nat. Rev. Immunol. 2018, 18, 225–242. [Google Scholar] [CrossRef]
- Thion, M.S.; Ginhoux, F.; Garel, S. Microglia and Early Brain Development: An Intimate Journey. Science 2018, 362, 185–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thion, M.S.; Low, D.; Silvin, A.; Chen, J.; Grisel, P.; Schulte-Schrepping, J.; Blecher, R.; Ulas, T.; Squarzoni, P.; Hoeffel, G.; et al. Microbiome Influences Prenatal and Adult Microglia in a Sex-Specific Manner. Cell 2018, 172, 500–516.e16. [Google Scholar] [CrossRef] [Green Version]
- Mousa, A.; SeigerA, N.; Kjaeldgaard, A.; Bakhiet, M. Human First Trimester Forebrain Cells Express Genes for Inflammatory and Anti-Inflammatory Cytokines. Cytokine 1999, 11, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Mousa, A.; Bakhiet, M. Role of Cytokine Signaling during Nervous System Development. Int. J. Mol. Sci. 2013, 14, 13931–13957. [Google Scholar] [CrossRef] [Green Version]
- Bauer, S.; Kerr, B.J.; Patterson, P.H. The Neuropoietic Cytokine Family in Development, Plasticity, Disease and Injury. Nat. Rev. Neurosci. 2007, 8, 221–232. [Google Scholar] [CrossRef]
- Samuelsson, A.-M.; Jennische, E.; Hansson, H.-A.; Holmäng, A. Prenatal Exposure to Interleukin-6 Results in Inflammatory Neurodegeneration in Hippocampus with NMDA/GABA(A) Dysregulation and Impaired Spatial Learning. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1345–R1356. [Google Scholar] [CrossRef] [Green Version]
- Graham, A.M.; Rasmussen, J.M.; Rudolph, M.D.; Heim, C.M.; Gilmore, J.H.; Styner, M.; Potkin, S.G.; Entringer, S.; Wadhwa, P.D.; Fair, D.A.; et al. Maternal Systemic Interleukin-6 During Pregnancy Is Associated with Newborn Amygdala Phenotypes and Subsequent Behavior at 2 Years of Age. Biol. Psychiatry 2018, 83, 109–119. [Google Scholar] [CrossRef]
- Curry, A.E.; Vogel, I.; Skogstrand, K.; Drews, C.; Schendel, D.E.; Flanders, W.D.; Hougaard, D.M.; Thorsen, P. Maternal Plasma Cytokines in Early- and Mid-Gestation of Normal Human Pregnancy and Their Association with Maternal Factors. J. Reprod. Immunol. 2008, 77, 152–160. [Google Scholar] [CrossRef]
- Catalano, P.M.; Presley, L.; Minium, J.; Hauguel-de Mouzon, S. Fetuses of Obese Mothers Develop Insulin Resistance in Utero. Diabetes Care 2009, 32, 1076–1080. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Haghiac, M.; Surace, P.; Challier, J.-C.; Guerre-Millo, M.; Singh, K.; Waters, T.; Minium, J.; Presley, L.; Catalano, P.M.; et al. Pregravid Obesity Associates with Increased Maternal Endotoxemia and Metabolic Inflammation. Obesity 2011, 19, 476–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, K.A.; Riley, S.C.; Reynolds, R.M.; Barr, S.; Evans, M.; Statham, A.; Hor, K.; Jabbour, H.N.; Norman, J.E.; Denison, F.C. Placental Structure and Inflammation in Pregnancies Associated with Obesity. Placenta 2011, 32, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Friis, C.M.; Paasche Roland, M.C.; Godang, K.; Ueland, T.; Tanbo, T.; Bollerslev, J.; Henriksen, T. Adiposity-Related Inflammation: Effects of Pregnancy. Obesity 2013, 21, E124–E130. [Google Scholar] [CrossRef] [PubMed]
- Christian, L.M.; Porter, K. Longitudinal Changes in Serum Proinflammatory Markers across Pregnancy and Postpartum: Effects of Maternal Body Mass Index. Cytokine 2014, 70, 134–140. [Google Scholar] [CrossRef] [Green Version]
- Sen, S.; Iyer, C.; Meydani, S.N. Obesity during Pregnancy Alters Maternal Oxidant Balance and Micronutrient Status. J. Perinatol. 2014, 34, 105–111. [Google Scholar] [CrossRef]
- Holtan, S.G.; Chen, Y.; Kaimal, R.; Creedon, D.J.; Enninga, E.A.L.; Nevala, W.K.; Markovic, S.N. Growth Modeling of the Maternal Cytokine Milieu throughout Normal Pregnancy: Macrophage-Derived Chemokine Decreases as Inflammation/Counterregulation Increases. J. Immunol. Res. 2015, 2015, 952571. [Google Scholar] [CrossRef] [Green Version]
- Ratnayake, U.; Quinn, T.; Walker, D.W.; Dickinson, H. Cytokines and the Neurodevelopmental Basis of Mental Illness. Front. Neurosci. 2013, 7, 180. [Google Scholar] [CrossRef] [Green Version]
- Torres-Platas, S.G.; Cruceanu, C.; Chen, G.G.; Turecki, G.; Mechawar, N. Evidence for Increased Microglial Priming and Macrophage Recruitment in the Dorsal Anterior Cingulate White Matter of Depressed Suicides. Brain Behav. Immun. 2014, 42, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Kalkman, H.O.; Feuerbach, D. Antidepressant Therapies Inhibit Inflammation and Microglial M1-Polarization. Pharmacol. Ther. 2016, 163, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Catale, C.; Gironda, S.; Lo Iacono, L.; Carola, V. Microglial Function in the Effects of Early-Life Stress on Brain and Behavioral Development. J. Clin. Med. 2020, 9, 468. [Google Scholar] [CrossRef] [Green Version]
- Al-Haddad, B.J.S.; Oler, E.; Armistead, B.; Elsayed, N.A.; Weinberger, D.R.; Bernier, R.; Burd, I.; Kapur, R.; Jacobsson, B.; Wang, C.; et al. The Fetal Origins of Mental Illness. Am. J. Obstet. Gynecol. 2019, 221, 549–562. [Google Scholar] [CrossRef]
- Ballard, O.; Morrow, A.L. Human Milk Composition: Nutrients and Bioactive Factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, X.; Mi, L.; Li, C.; Zhang, Y.; Bi, R.; Pang, J.; Li, Y. Comparative Proteomic Analysis of Proteins in Breast Milk during Different Lactation Periods. Nutrients 2022, 14, 3648. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, M.G.; Bottanelli, C.; Cattaneo, A.; Pariante, C.M.; Borsini, A. Nutritional and Immunological Factors in Breast Milk: A Role in the Intergenerational Transmission from Maternal Psychopathology to Child Development. Brain Behav. Immun. 2020, 85, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.M. The Origin of Human Milk Bacteria: Is There a Bacterial Entero-Mammary Pathway during Late Pregnancy and Lactation? Adv. Nutr. 2014, 5, 779–784. [Google Scholar] [CrossRef] [Green Version]
- Moossavi, S.; Sepehri, S.; Robertson, B.; Bode, L.; Goruk, S.; Field, C.J.; Lix, L.M.; de Souza, R.J.; Becker, A.B.; Mandhane, P.J.; et al. Composition and Variation of the Human Milk Microbiota Are Influenced by Maternal and Early-Life Factors. Cell Host Microbe 2019, 25, 324–335.e4. [Google Scholar] [CrossRef] [Green Version]
- Cabrera-Rubio, R.; Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E.; Mira, A. The Human Milk Microbiome Changes over Lactation and Is Shaped by Maternal Weight and Mode of Delivery. Am. J. Clin. Nutr. 2012, 96, 544–551. [Google Scholar] [CrossRef] [Green Version]
- Borre, Y.E.; O’Keeffe, G.W.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota and Neurodevelopmental Windows: Implications for Brain Disorders. Trends Mol. Med. 2014, 20, 509–518. [Google Scholar] [CrossRef] [PubMed]
- De Leoz, M.L.A.; Kalanetra, K.M.; Bokulich, N.A.; Strum, J.S.; Underwood, M.A.; German, J.B.; Mills, D.A.; Lebrilla, C.B. Human Milk Glycomics and Gut Microbial Genomics in Infant Feces Show a Correlation between Human Milk Oligosaccharides and Gut Microbiota: A Proof-of-Concept Study. J. Proteome Res. 2015, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Morrow, A.L.; Ruiz-Palacios, G.M.; Jiang, X.; Newburg, D.S. Human-Milk Glycans That Inhibit Pathogen Binding Protect Breast-Feeding Infants against Infectious Diarrhea. J. Nutr. 2005, 135, 1304–1307. [Google Scholar] [CrossRef] [Green Version]
- Korpela, K.; Salonen, A.; Hickman, B.; Kunz, C.; Sprenger, N.; Kukkonen, K.; Savilahti, E.; Kuitunen, M.; de Vos, W.M. Fucosylated Oligosaccharides in Mother’s Milk Alleviate the Effects of Caesarean Birth on Infant Gut Microbiota. Sci. Rep. 2018, 8, 13757. [Google Scholar] [CrossRef] [Green Version]
- Soyyılmaz, B.; Mikš, M.H.; Röhrig, C.H.; Matwiejuk, M.; Meszaros-Matwiejuk, A.; Vigsnæs, L.K. The Mean of Milk: A Review of Human Milk Oligosaccharide Concentrations throughout Lactation. Nutrients 2021, 13, 2737. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.E.; Heisel, T.; Allert, M.; Fürst, A.; Yerabandi, N.; Knights, D.; Jacobs, K.M.; Lock, E.F.; Bode, L.; Fields, D.A.; et al. Human Milk Variation Is Shaped by Maternal Genetics and Impacts the Infant Gut Microbiome. bioRxiv 2023. [Google Scholar] [CrossRef]
- Binns, C.; Lee, M.; Low, W.Y. The Long-Term Public Health Benefits of Breastfeeding. Asia Pac. J. Public Health 2016, 28, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Ellsworth, L.; Perng, W.; Harman, E.; Das, A.; Pennathur, S.; Gregg, B. Impact of Maternal Overweight and Obesity on Milk Composition and Infant Growth. Matern. Child Nutr. 2020, 16, e12979. [Google Scholar] [CrossRef]
- Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E. Maternal Weight and Excessive Weight Gain during Pregnancy Modify the Immunomodulatory Potential of Breast Milk. Pediatr. Res. 2012, 72, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Sela, D.A.; Chapman, J.; Adeuya, A.; Kim, J.H.; Chen, F.; Whitehead, T.R.; Lapidus, A.; Rokhsar, D.S.; Lebrilla, C.B.; German, J.B.; et al. The Genome Sequence of Bifidobacterium Longum Subsp. Infantis Reveals Adaptations for Milk Utilization within the Infant Microbiome. Proc. Natl. Acad. Sci. USA 2008, 105, 18964–18969. [Google Scholar] [CrossRef] [Green Version]
- Andreas, N.J.; Hyde, M.J.; Gale, C.; Parkinson, J.R.C.; Jeffries, S.; Holmes, E.; Modi, N. Effect of Maternal Body Mass Index on Hormones in Breast Milk: A Systematic Review. PLoS ONE 2014, 9, e115043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuyama, H.; Hiramatsu, Y. Additive Effects of Maternal High Fat Diet during Lactation on Mouse Offspring. PLoS ONE 2014, 9, e92805. [Google Scholar] [CrossRef] [PubMed]
- Erliana, U.D.; Fly, A.D. The Function and Alteration of Immunological Properties in Human Milk of Obese Mothers. Nutrients 2019, 11, 1284. [Google Scholar] [CrossRef] [Green Version]
- Pettersen-Dahl, A.; Murzakanova, G.; Sandvik, L.; Laine, K. Maternal Body Mass Index as a Predictor for Delivery Method. Acta Obstet. Gynecol. Scand. 2018, 97, 212–218. [Google Scholar] [CrossRef] [Green Version]
- Bever Babendure, J.; Reifsnider, E.; Mendias, E.; Moramarco, M.W.; Davila, Y.R. Reduced Breastfeeding Rates among Obese Mothers: A Review of Contributing Factors, Clinical Considerations and Future Directions. Int. Breastfeed. J. 2015, 10, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballesta-Castillejos, A.; Gomez-Salgado, J.; Rodriguez-Almagro, J.; Ortiz-Esquinas, I.; Hernandez-Martinez, A. Relationship between Maternal Body Mass Index with the Onset of Breastfeeding and Its Associated Problems: An Online Survey. Int. Breastfeed. J. 2020, 15, 55. [Google Scholar] [CrossRef]
- Banić, M.; Butorac, K.; Čuljak, N.; Leboš Pavunc, A.; Novak, J.; Bellich, B.; Kazazić, S.; Kazazić, S.; Cescutti, P.; Šušković, J.; et al. The Human Milk Microbiota Produces Potential Therapeutic Biomolecules and Shapes the Intestinal Microbiota of Infants. Int. J. Mol. Sci. 2022, 23, 14382. [Google Scholar] [CrossRef]
- Pannaraj, P.S.; Li, F.; Cerini, C.; Bender, J.M.; Yang, S.; Rollie, A.; Adisetiyo, H.; Zabih, S.; Lincez, P.J.; Bittinger, K.; et al. Association Between Breast Milk Bacterial Communities and Establishment and Development of the Infant Gut Microbiome. JAMA Pediatr. 2017, 171, 647–654. [Google Scholar] [CrossRef] [Green Version]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel Disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Thompson, A.L. Intergenerational Impact of Maternal Obesity and Postnatal Feeding Practices on Pediatric Obesity. Nutr. Rev. 2013, 71, S55–S61. [Google Scholar] [CrossRef] [Green Version]
- Bar, S.; Milanaik, R.; Adesman, A. Long-Term Neurodevelopmental Benefits of Breastfeeding. Curr. Opin. Pediatr. 2016, 28, 559–566. [Google Scholar] [CrossRef]
- Horta, B.L.; de Sousa, B.A.; de Mola, C.L. Breastfeeding and Neurodevelopmental Outcomes. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Deng, Q.; Wang, J.; Wang, H.; Li, Q.; Zhu, B.; Ji, C.; Xu, X.; Johnston, L. The Impact of Breast Milk Feeding on Early Brain Development in Preterm Infants in China: An Observational Study. PLoS ONE 2022, 17, e0272125. [Google Scholar] [CrossRef] [PubMed]
- Bann, D.; Johnson, W.; Li, L.; Kuh, D.; Hardy, R. Socioeconomic Inequalities in Body Mass Index across Adulthood: Coordinated Analyses of Individual Participant Data from Three British Birth Cohort Studies Initiated in 1946, 1958 and 1970. PLoS Med. 2017, 14, e1002214. [Google Scholar] [CrossRef] [Green Version]
- English, S.; Steele, A.; Williams, A.; Blacklay, J.; Sorinola, O.; Wernisch, L.; Grammatopoulos, D.K. Modelling of Psychosocial and Lifestyle Predictors of Peripartum Depressive Symptoms Associated with Distinct Risk Trajectories: A Prospective Cohort Study. Sci. Rep. 2018, 8, 12799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abel, K.M.; Hope, H.; Swift, E.; Parisi, R.; Ashcroft, D.M.; Kosidou, K.; Osam, C.S.; Dalman, C.; Pierce, M. Prevalence of Maternal Mental Illness among Children and Adolescents in the UK between 2005 and 2017: A National Retrospective Cohort Analysis. Lancet Public Health 2019, 4, e291–e300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, M.S.; Fombonne, E.; Matush, L.; Bogdanovich, N.; Dahhou, M.; Platt, R.W. Long-Term Behavioural Consequences of Infant Feeding: The Limits of Observational Studies. Paediatr. Perinat. Epidemiol. 2011, 25, 500–506. [Google Scholar] [CrossRef]
- Taveras, E.M.; Capra, A.M.; Braveman, P.A.; Jensvold, N.G.; Escobar, G.J.; Lieu, T.A. Clinician Support and Psychosocial Risk Factors Associated with Breastfeeding Discontinuation. Pediatrics 2003, 112 Pt 1, 108–115. [Google Scholar] [CrossRef]
- Garratt, E.A.; Chandola, T.; Purdam, K.; Wood, A.M. Income and Social Rank Influence UK Children’s Behavioral Problems: A Longitudinal Analysis. Child Dev. 2017, 88, 1302–1320. [Google Scholar] [CrossRef] [Green Version]
- Hafner, H.; Chang, E.; Carlson, Z.; Zhu, A.; Varghese, M.; Clemente, J.; Abrishami, S.; Bagchi, D.P.; MacDougald, O.A.; Singer, K.; et al. Lactational High-Fat Diet Exposure Programs Metabolic Inflammation and Bone Marrow Adiposity in Male Offspring. Nutrients 2019, 11, 1393. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhang, Y.; Miller, D.; Rehman, N.O.; Cheng, X.; Yeo, J.-Y.; Joe, B.; Hill, J.W. Microbial Reconstitution Reverses Early Female Puberty Induced by Maternal High-Fat Diet During Lactation. Endocrinology 2020, 161, bqz041. [Google Scholar] [CrossRef]
- Wan, Y.; Wang, F.; Yuan, J.; Li, J.; Jiang, D.; Zhang, J.; Li, H.; Wang, R.; Tang, J.; Huang, T.; et al. Effects of Dietary Fat on Gut Microbiota and Faecal Metabolites, and Their Relationship with Cardiometabolic Risk Factors: A 6-Month Randomised Controlled-Feeding Trial. Gut 2019, 68, 1417–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, S.H.; Belenguer, A.; Holtrop, G.; Johnstone, A.M.; Flint, H.J.; Lobley, G.E. Reduced Dietary Intake of Carbohydrates by Obese Subjects Results in Decreased Concentrations of Butyrate and Butyrate-Producing Bacteria in Feces. Appl. Environ. Microbiol. 2007, 73, 1073–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The Neuroactive Potential of the Human Gut Microbiota in Quality of Life and Depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, J.; Yang, S.; Utturkar, S.; Crodian, J.; Cummings, S.; Thimmapuram, J.; San Miguel, P.; Kuang, S.; Gribskov, M.; et al. Effect of High-Fat Diet on Secreted Milk Transcriptome in Midlactation Mice. Physiol. Genom. 2017, 49, 747–762. [Google Scholar] [CrossRef]
- Liao, Y.; Du, X.; Li, J.; Lönnerdal, B. Human Milk Exosomes and Their MicroRNAs Survive Digestion in Vitro and Are Taken up by Human Intestinal Cells. Mol. Nutr. Food Res. 2017, 61, 1700082. [Google Scholar] [CrossRef]
- Innis, S.M. Dietary Omega 3 Fatty Acids and the Developing Brain. Brain Res. 2008, 1237, 35–43. [Google Scholar] [CrossRef]
- Colombo, J.; Carlson, S.E.; Cheatham, C.L.; Fitzgerald-Gustafson, K.M.; Kepler, A.; Doty, T. Long-Chain Polyunsaturated Fatty Acid Supplementation in Infancy Reduces Heart Rate and Positively Affects Distribution of Attention. Pediatr. Res. 2011, 70, 406–410. [Google Scholar] [CrossRef]
- Ansorge, M.S.; Zhou, M.; Lira, A.; Hen, R.; Gingrich, J.A. Early-Life Blockade of the 5-HT Transporter Alters Emotional Behavior in Adult Mice. Science 2004, 306, 879–881. [Google Scholar] [CrossRef] [Green Version]
- Bonnin, A.; Torii, M.; Wang, L.; Rakic, P.; Levitt, P. Serotonin Modulates the Response of Embryonic Thalamocortical Axons to Netrin-1. Nat. Neurosci. 2007, 10, 588–597. [Google Scholar] [CrossRef]
- Clarke, G.; Grenham, S.; Scully, P.; Fitzgerald, P.; Moloney, R.D.; Shanahan, F.; Dinan, T.G.; Cryan, J.F. The Microbiome-Gut-Brain Axis during Early Life Regulates the Hippocampal Serotonergic System in a Sex-Dependent Manner. Mol. Psychiatry 2013, 18, 666–673. [Google Scholar] [CrossRef] [Green Version]
- Pfaff, A.W.; Mousli, M.; Sénégas, A.; Marcellin, L.; Takikawa, O.; Klein, J.-P.; Candolfi, E. Impact of Foetus and Mother on IFN-Gamma-Induced Indoleamine 2,3-Dioxygenase and Inducible Nitric Oxide Synthase Expression in Murine Placenta Following Toxoplasma Gondii Infection. Int. J. Parasitol. 2008, 38, 249–258. [Google Scholar] [CrossRef]
- Kannan, S.; Saadani-Makki, F.; Balakrishnan, B.; Dai, H.; Chakraborty, P.K.; Janisse, J.; Muzik, O.; Romero, R.; Chugani, D.C. Decreased Cortical Serotonin in Neonatal Rabbits Exposed to Endotoxin in Utero. J. Cereb. Blood Flow Metab. 2011, 31, 738–749. [Google Scholar] [CrossRef] [Green Version]
- Beutler, B. Tlr4: Central Component of the Sole Mammalian LPS Sensor. Curr. Opin. Immunol. 2000, 12, 20–26. [Google Scholar] [CrossRef]
- Koga, K.; Izumi, G.; Mor, G.; Fujii, T.; Osuga, Y. Toll-like Receptors at the Maternal-Fetal Interface in Normal Pregnancy and Pregnancy Complications. Am. J. Reprod. Immunol. 2014, 72, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.A.; Hutchinson, M.R.; Rice, K.C.; Chin, P.-Y.; Moldenhauer, L.M.; Stark, M.J.; Olson, D.M.; Keelan, J.A. Targeting Toll-like Receptor-4 to Tackle Preterm Birth and Fetal Inflammatory Injury. Clin. Transl. Immunol. 2020, 9, e1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Li, M.; Haghiac, M.; Catalano, P.M.; O’Tierney-Ginn, P.; Hauguel-de Mouzon, S. Causal Relationship between Obesity-Related Traits and TLR4-Driven Responses at the Maternal-Fetal Interface. Diabetologia 2016, 59, 2459–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnin, A.; Goeden, N.; Chen, K.; Wilson, M.L.; King, J.; Shih, J.C.; Blakely, R.D.; Deneris, E.S.; Levitt, P. A Transient Placental Source of Serotonin for the Fetal Forebrain. Nature 2011, 472, 347–350. [Google Scholar] [CrossRef] [Green Version]
- Goeden, N.; Velasquez, J.; Arnold, K.A.; Chan, Y.; Lund, B.T.; Anderson, G.M.; Bonnin, A. Maternal Inflammation Disrupts Fetal Neurodevelopment via Increased Placental Output of Serotonin to the Fetal Brain. J. Neurosci. 2016, 36, 6041–6049. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.A.; Fairburn, C.G.; Cowen, P.J. Relapse of Depression after Rapid Depletion of Tryptophan. Lancet 1997, 349, 915–919. [Google Scholar] [CrossRef]
- Del Colle, A.; Israelyan, N.; Gross Margolis, K. Novel Aspects of Enteric Serotonergic Signaling in Health and Brain-Gut Disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G130–G143. [Google Scholar] [CrossRef] [Green Version]
- Gross, C.; Hen, R. The Developmental Origins of Anxiety. Nat. Rev. Neurosci. 2004, 5, 545–552. [Google Scholar] [CrossRef]
- Chaput, Y.; de Montigny, C.; Blier, P. Effects of a Selective 5-HT Reuptake Blocker, Citalopram, on the Sensitivity of 5-HT Autoreceptors: Electrophysiological Studies in the Rat Brain. Naunyn Schmiedebergs Arch. Pharmacol. 1986, 333, 342–348. [Google Scholar] [CrossRef]
- Kubo, A.; Ferrara, A.; Laurent, C.A.; Windham, G.C.; Greenspan, L.C.; Deardorff, J.; Hiatt, R.A.; Quesenberry, C.P.; Kushi, L.H. Associations Between Maternal Pregravid Obesity and Gestational Diabetes and the Timing of Pubarche in Daughters. Am. J. Epidemiol. 2016, 184, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, T.J.; Fonseca, M.A.; Campbell, K.E.; Moyce, B.L.; Cole, L.K.; Hatch, G.M.; Doucette, C.A.; Klein, J.; Aliani, M.; Dolinsky, V.W. Maternal Obesity Characterized by Gestational Diabetes Increases the Susceptibility of Rat Offspring to Hepatic Steatosis via a Disrupted Liver Metabolome. J. Physiol. 2015, 593, 3181–3197. [Google Scholar] [CrossRef] [Green Version]
- Paul, H.A.; Collins, K.H.; Nicolucci, A.C.; Urbanski, S.J.; Hart, D.A.; Vogel, H.J.; Reimer, R.A. Maternal Prebiotic Supplementation Reduces Fatty Liver Development in Offspring through Altered Microbial and Metabolomic Profiles in Rats. FASEB J. 2019, 33, 5153–5167. [Google Scholar] [CrossRef] [PubMed]
- Savva, C.; Helguero, L.A.; González-Granillo, M.; Couto, D.; Melo, T.; Li, X.; Angelin, B.; Domingues, M.R.; Kutter, C.; Korach-André, M. Obese Mother Offspring Have Hepatic Lipidic Modulation That Contributes to Sex-Dependent Metabolic Adaptation Later in Life. Commun. Biol. 2021, 4, 14. [Google Scholar] [CrossRef]
- Bucher, M.; Montaniel, K.R.C.; Myatt, L.; Weintraub, S.; Tavori, H.; Maloyan, A. Dyslipidemia, Insulin Resistance, and Impairment of Placental Metabolism in the Offspring of Obese Mothers. J. Dev. Orig. Health Dis. 2021, 12, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Han, T.-L.; Zhao, Y.; Zhou, X.; Mao, X.; Qi, H.; Baker, P.N.; Zhang, H. A Mouse Model of Pre-Pregnancy Maternal Obesity Combined with Offspring Exposure to a High-Fat Diet Resulted in Cognitive Impairment in Male Offspring. Exp. Cell Res. 2018, 368, 159–166. [Google Scholar] [CrossRef]
- Marosi, K.; Kim, S.W.; Moehl, K.; Scheibye-Knudsen, M.; Cheng, A.; Cutler, R.; Camandola, S.; Mattson, M.P. 3-Hydroxybutyrate Regulates Energy Metabolism and Induces BDNF Expression in Cerebral Cortical Neurons. J. Neurochem. 2016, 139, 769–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, H. Fatty Acid Oxidation. In Encyclopedia of Biological Chemistry, 2nd ed.; Lennarz, W.J., Lane, M.D., Eds.; Academic Press: Waltham, MA, USA, 2013; pp. 281–284. [Google Scholar] [CrossRef]
- De Vadder, F.; Kovatcheva-Datchary, P.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-Produced Succinate Improves Glucose Homeostasis via Intestinal Gluconeogenesis. Cell Metab. 2016, 24, 151–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, K.; Krishnan, A.; Cervenka, E.; Hu, G.; Guadagno, E.; Trakadis, Y. Biomarkers for Major Depressive and Bipolar Disorders Using Metabolomics: A Systematic Review. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2019, 180, 122–137. [Google Scholar] [CrossRef] [PubMed]
- Bakian, A.V.; Huber, R.S.; Scholl, L.; Renshaw, P.F.; Kondo, D. Dietary Creatine Intake and Depression Risk among U.S. Adults. Transl. Psychiatry 2020, 10, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, D.G.; Sung, Y.-H.; Hellem, T.L.; Fiedler, K.K.; Shi, X.; Jeong, E.-K.; Renshaw, P.F. Open-Label Adjunctive Creatine for Female Adolescents with SSRI-Resistant Major Depressive Disorder: A 31-Phosphorus Magnetic Resonance Spectroscopy Study. J. Affect. Disord. 2011, 135, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Harper, D.G.; Jensen, J.E.; Ravichandran, C.; Perlis, R.H.; Fava, M.; Renshaw, P.F.; Iosifescu, D.V. Tissue Type-Specific Bioenergetic Abnormalities in Adults with Major Depression. Neuropsychopharmacology 2017, 42, 876–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Della, F.P.; Abelaira, H.M.; Réus, G.Z.; Ribeiro, K.F.; Antunes, A.R.; Scaini, G.; Jeremias, I.C.; dos Santos, L.M.M.; Jeremias, G.C.; Streck, E.L.; et al. Tianeptine Treatment Induces Antidepressive-like Effects and Alters BDNF and Energy Metabolism in the Brain of Rats. Behav. Brain Res. 2012, 233, 526–535. [Google Scholar] [CrossRef]
- Song, X.; Chen, X.; Yuksel, C.; Yuan, J.; Pizzagalli, D.A.; Forester, B.; Öngür, D.; Du, F. Bioenergetics and Abnormal Functional Connectivity in Psychotic Disorders. Mol. Psychiatry 2021, 26, 2483–2492. [Google Scholar] [CrossRef]
- Sullivan, E.L.; Nousen, E.K.; Chamlou, K.A. Maternal High Fat Diet Consumption during the Perinatal Period Programs Offspring Behavior. Physiol. Behav. 2014, 123, 236–242. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Choi, Y.J.; Joung, S.M.; Lee, B.H.; Jung, Y.-S.; Lee, J.Y. Hypoxic Stress Up-Regulates the Expression of Toll-like Receptor 4 in Macrophages via Hypoxia-Inducible Factor. Immunology 2010, 129, 516–524. [Google Scholar] [CrossRef]
- Korbecki, J.; Bajdak-Rusinek, K. The Effect of Palmitic Acid on Inflammatory Response in Macrophages: An Overview of Molecular Mechanisms. Inflamm. Res. 2019, 68, 915–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilg, H.; Zmora, N.; Adolph, T.E.; Elinav, E. The Intestinal Microbiota Fuelling Metabolic Inflammation. Nat. Rev. Immunol. 2020, 20, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Ballantyne, C.M. Metabolic Inflammation and Insulin Resistance in Obesity. Circ. Res. 2020, 126, 1549–1564. [Google Scholar] [CrossRef] [PubMed]
- Paul, K.H.; Graham, M.L.; Olson, C.M. The Web of Risk Factors for Excessive Gestational Weight Gain in Low Income Women. Matern. Child Health J. 2013, 17, 344–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NICE. Weight Management before, during and after Pregnancy; NICE: London, UK, 2010; p. 56. Available online: https://www.nice.org.uk/guidance/ph27/resources/weight-management-before-during-and-after-pregnancy-pdf-1996242046405 (accessed on 14 March 2023).
- Denison, F.; Aedla, N.; Keag, O.; Hor, K.; Reynolds, R.; Milne, A.; Diamond, A.; the Royal College of Obstetricians and Gynaecologists. Care of Women with Obesity in Pregnancy. BJOG Int. J. Obstet. Gynaecol. 2019, 126, e62–e106. [Google Scholar] [CrossRef] [Green Version]
- Heslehurst, N.; Rankin, J.; Wilkinson, J.R.; Summerbell, C.D. A Nationally Representative Study of Maternal Obesity in England, UK: Trends in Incidence and Demographic Inequalities in 619 323 Births, 1989–2007. Int. J. Obes. 2010, 34, 420–428. [Google Scholar] [CrossRef] [Green Version]
- Mojtabai, R. Body Mass Index and Serum Folate in Childbearing Age Women. Eur. J. Epidemiol. 2004, 19, 1029–1036. [Google Scholar] [CrossRef]
- Tinker, S.C.; Hamner, H.C.; Berry, R.J.; Bailey, L.B.; Pfeiffer, C.M. Does Obesity Modify the Association of Supplemental Folic Acid with Folate Status among Nonpregnant Women of Childbearing Age in the United States? Birth Defects Res. A Clin. Mol. Teratol. 2012, 94, 749–755. [Google Scholar] [CrossRef]
- Knight, B.A.; Shields, B.M.; Brook, A.; Hill, A.; Bhat, D.S.; Hattersley, A.T.; Yajnik, C.S. Lower Circulating B12 Is Associated with Higher Obesity and Insulin Resistance during Pregnancy in a Non-Diabetic White British Population. PLoS ONE 2015, 10, e0135268. [Google Scholar] [CrossRef] [Green Version]
- van der Windt, M.; Schoenmakers, S.; van Rijn, B.; Galjaard, S.; Steegers-Theunissen, R.; van Rossem, L. Epidemiology and (Patho)Physiology of Folic Acid Supplement Use in Obese Women before and during Pregnancy. Nutrients 2021, 13, 331. [Google Scholar] [CrossRef]
- Jacka, F.N.; Ystrom, E.; Brantsaeter, A.L.; Karevold, E.; Roth, C.; Haugen, M.; Meltzer, H.M.; Schjolberg, S.; Berk, M. Maternal and Early Postnatal Nutrition and Mental Health of Offspring by Age 5 Years: A Prospective Cohort Study. J. Am. Acad. Child Adolesc. Psychiatry 2013, 52, 1038–1047. [Google Scholar] [CrossRef]
- Dawson, S.L.; O’Hely, M.; Jacka, F.N.; Ponsonby, A.-L.; Symeonides, C.; Loughman, A.; Collier, F.; Moreno-Betancur, M.; Sly, P.; Burgner, D.; et al. Maternal Prenatal Gut Microbiota Composition Predicts Child Behaviour. EBioMedicine 2021, 68, 103400. [Google Scholar] [CrossRef]
- Visconti, A.; Le Roy, C.I.; Rosa, F.; Rossi, N.; Martin, T.C.; Mohney, R.P.; Li, W.; de Rinaldis, E.; Bell, J.T.; Venter, J.C.; et al. Interplay between the Human Gut Microbiome and Host Metabolism. Nat. Commun. 2019, 10, 4505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Diener, C.; Qin, S.; Zhou, Y.; Patwardhan, S.; Tang, L.; Lovejoy, J.C.; Magis, A.T.; Price, N.D.; Hood, L.; Gibbons, S.M. Baseline Gut Metagenomic Functional Gene Signature Associated with Variable Weight Loss Responses Following a Healthy Lifestyle Intervention in Humans. mSystems 2021, 6, e00964-21. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Arraño, V.; Martín-Peláez, S. Effects of Probiotics and Synbiotics on Weight Loss in Subjects with Overweight or Obesity: A Systematic Review. Nutrients 2021, 13, 3627. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Olendzki, B.C.; Wang, J.; Persuitte, G.M.; Li, W.; Fang, H.; Merriam, P.A.; Wedick, N.M.; Ockene, I.S.; Culver, A.L.; et al. Single-Component Versus Multicomponent Dietary Goals for the Metabolic Syndrome. Ann. Intern. Med. 2015, 162, 248–257. [Google Scholar] [CrossRef] [Green Version]


| Species (Strain) | Maternal Diet | Intervention Period, Length | Offspring Behavioural Outcomes | Offspring Brain Outcomes | Author, Year |
|---|---|---|---|---|---|
| Mouse (CD-1) | HFD (60% kcal fat) vs. CD (10% kcal fat) | Prenatal + postnatal, 12 weeks (including 6 weeks pre-gestation) | ↑ anxiety OFT PND21 and PND112; ↑ passive stress coping FST PND21 and PND112; ↓ social interaction PND56 | ↓ brain creatine and brain glutamate PND21; ↑ PFC ZIF-268 mRNA PND21; ↓ PFC SYP mRNA PND112 | Radford-Smith et al., 2022 [12] |
| Rat (Sprague Dawley) | HFD (60% kcal sat. fat) vs. trans fat (60% kcal trans-fat) vs. CD (10% kcal fat) | Prenatal + postnatal, 10 weeks (including 4 weeks pre-gestation) | ↑ anxiety EPM (males only), ↑ memory performance MWM in adulthood (3 months age) | ↑ hippocampal microglial activation (CD11b, TLR4) at PND1, ↑ IL-1B protein at PND20, ↑ hippocampal microglial activation and reactivity in adulthood (3 months age) | Bilbo and Tsang, 2010 [10] |
| Japanese macaques | HFD (32% kcal fat) vs. CD (13% kcal fat) | Up to 4 years (including prenatally) | ↑ anxiety female juvenile offspring (PND130) | ↑ TPH2, 5-HT1AR in dorsal raphe in foetuses; | Sullivan et al., 2010 [11] |
| Rat (Wistar) | HFD (40% kcal fat) vs. CD (12% kcal fat) | Gestation at lactation (until PND21) | ↓ anxiety in OFT, ↑ social interaction in HFD adult offspring experiencing maternal separation in early life compared to CD adult offspring | Prevents neurobiological effects (↓ BDNF, 5-HT1AR, CRH) of maternal separation stress in juvenile offspring (PND11) and anxiety-induced cFOS and corticosterone in adulthood (8 months). | Rincel et al., 2016 [13] |
| Mouse (C57Bl/6) | HFD (45% kcal fat) vs. CD (10% kcal fat) | 6 weeks (including gestation) | Pups exposed to HFD in utero altered maternal care behaviour (reduced time spent on nest) independent of maternal diet (cross-fostering experiment) | - | Baptissart et al., 2018 [14] |
| Rat (Long Evans) | HFD (60% kcal fat) vs. chow diet (13.5% kcal fat) | ~6 weeks (including 3 weeks pre-gestation) | Pup isolation test: HFD pups vocalised less at PND7, more at PND13 | ↑ CRH, ↓ NR3C1 in paraventricular nucleus at PND7, ↑ GAD1, ↓ NR3C1 in ventral hippocampus at PND13 | Abuaish et al., 2018 [15] |
| Mouse (C57Bl/6) | HFD (60% kcal fat) vs. chow diet (13.4% kcal fat) | Prenatal + postnatal, 14 weeks (including 8 weeks pre-gestation) | ↓ social interaction (3-chamber social interaction test), ↑ anxiety (OFT and marble burying) 7–12 week offspring | ↓ hypothalamic oxytocin in HFD offspring; ↓ social interaction-induced dopaminergic activity in ventral tegmental area | Buffington et al., 2016 [16] |
| Mouse (C57Bl/6) | HFD (45% kcal fat) vs. CD (10% kcal fat) | Variable pre-gestation until significant weight gain occurred | - | ↓ neuropeptide Y innervation of paraventricular nucleus in foetal offspring | Sanders et al., 2014 [17] |
| Mouse (C57Bl/6) | Chronic mild stress + Western diet (40% kcal fat) vs. no stress CD (10% kcal fat) | Prenatal + postnatal, 11 weeks (including 5 weeks pre-gestation) | - | ↑ hippocampal microglial activation (↑ CD11b+ cells, ↑ AIF1, TLR9 mRNA and protein); ↓ neuronal cell density (NeuN+ cells) in hippocampus | Cohen et al., 2016 [18] |
| Japanese macaques | HFD (36.6% kcal fat) vs. CD (14.7% kcal fat) | Mothers had been consuming HFD for 1.2–8.5 years at birth of offspring | ↑ anxiety and stereotypic behaviours in offspring (11 months age) | ↓ TPH2 mRNA in dorsal raphe; ↓ serotonin immunoreactivity prefrontal cortex (13 months age) | Thompson et al., 2017 [19] |
| Mouse (C57Bl/6) | HFD (60% kcal fat) vs. CD (10% kcal fat) vs. weight loss (HFD → CD) | Prenatal + postnatal, 5.5 months (including 4 months pre-gestation) | - | No significant effect of maternal diet on offspring hypothalamus or olfactory bulb metabolite levels | Safi-Stibler et al., 2020 [20] |
| Rat (Sprague Dawley) | Grandmaternal HFD (60% kcal fat) vs. CD (10% kcal fat) | Prenatal + postnatal, 14 weeks (including 8 weeks pre-gestation) | ↑ anxiety in female F2 offspring in EPM | ↑ hippocampal CRH-R2 in male HFD F2 offspring; ↑ NR3C2 in female HFD F2 offspring | Winther et al., 2019 [21] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radford-Smith, D.E.; Anthony, D.C. Mechanisms of Maternal Diet-Induced Obesity Affecting the Offspring Brain and Development of Affective Disorders. Metabolites 2023, 13, 455. https://doi.org/10.3390/metabo13030455
Radford-Smith DE, Anthony DC. Mechanisms of Maternal Diet-Induced Obesity Affecting the Offspring Brain and Development of Affective Disorders. Metabolites. 2023; 13(3):455. https://doi.org/10.3390/metabo13030455
Chicago/Turabian StyleRadford-Smith, Daniel E., and Daniel C. Anthony. 2023. "Mechanisms of Maternal Diet-Induced Obesity Affecting the Offspring Brain and Development of Affective Disorders" Metabolites 13, no. 3: 455. https://doi.org/10.3390/metabo13030455