
Systematic Review: Contribution of the Gut Microbiome to the Volatile Metabolic Fingerprint of Colorectal Neoplasia
 , ,
, ,
Abstract
:1. Introduction
1.1. Colorectal Neoplasia
1.2. Volatile Organic Compounds
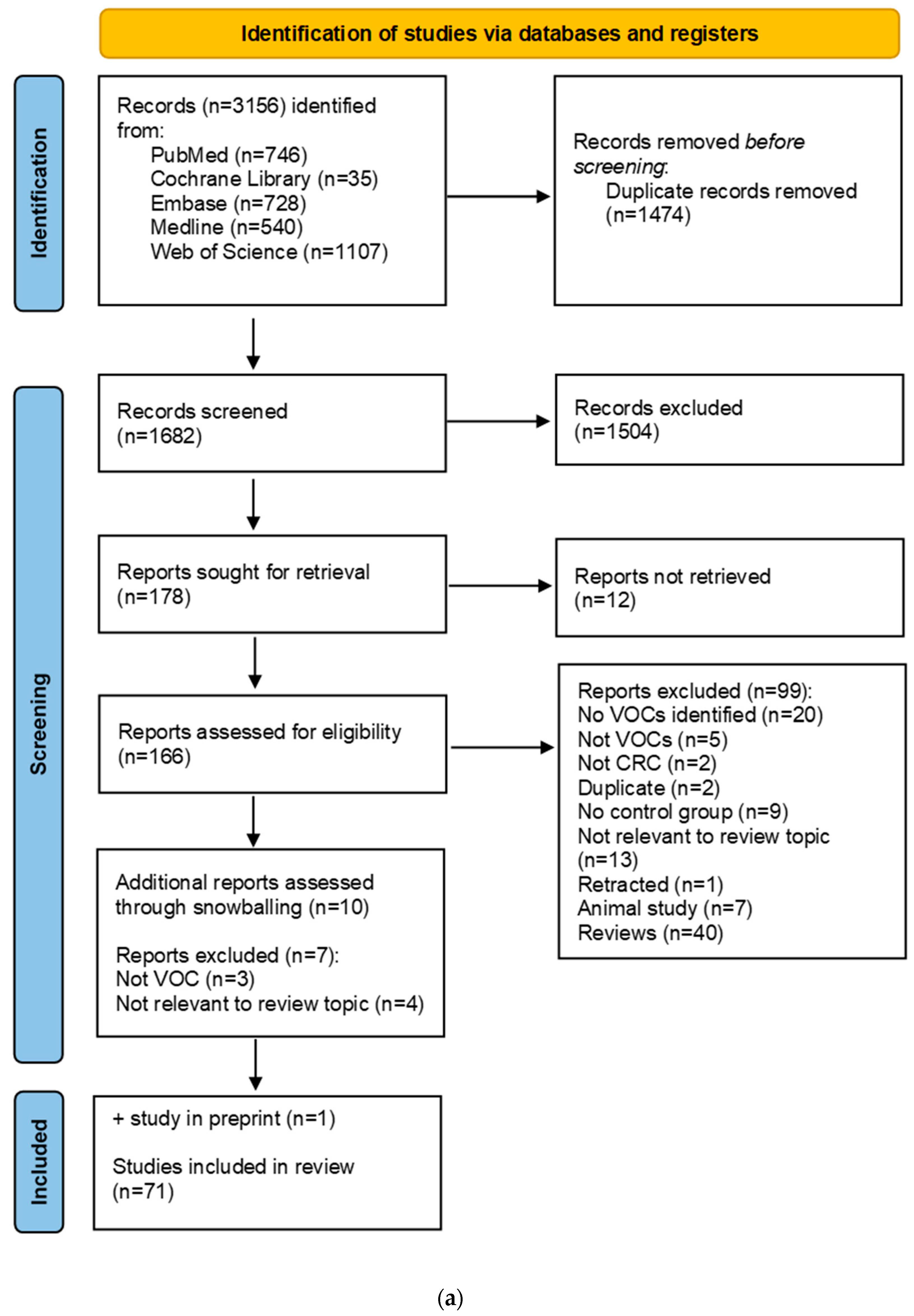
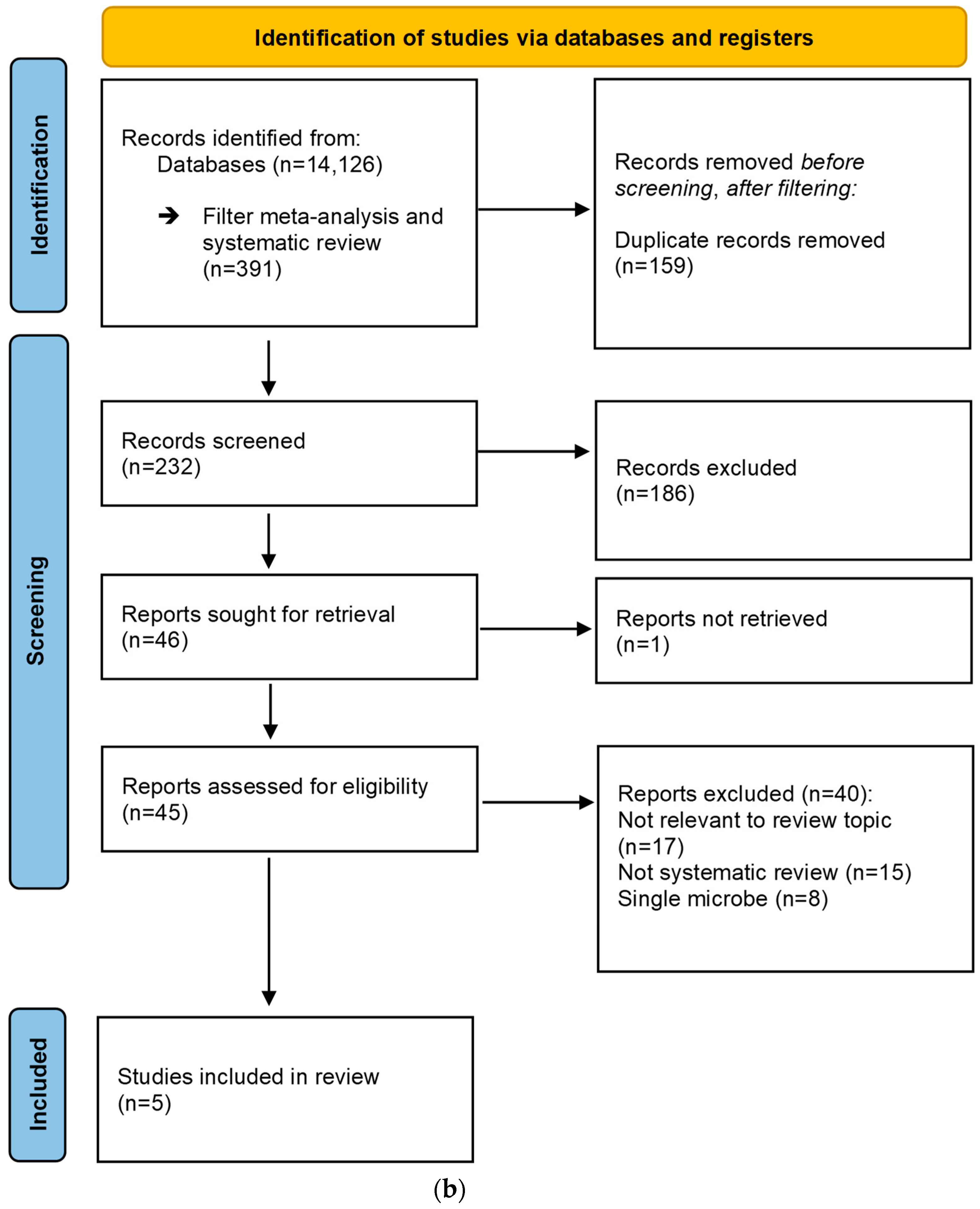
2. Materials and Methods
2.1. Search Strategy
2.2. Selection Criteria
2.3. Data Extraction
2.4. VOC Data Interpretation
3. Results
3.1. Description of VOC Studies
3.2. Fecal VOC Profile
3.3. Dynamics between Fecal VOCs and Gut Microbiota
3.4. Urinary VOC Profile
3.5. Exhaled Breath VOC Profile
3.6. Blood VOC Profile
3.7. Tissue VOC Profile
3.8. Saliva VOC Profile
3.9. Summary of the Volatolome and Gut Microbial Microenvironment in Colorectal Neoplasia
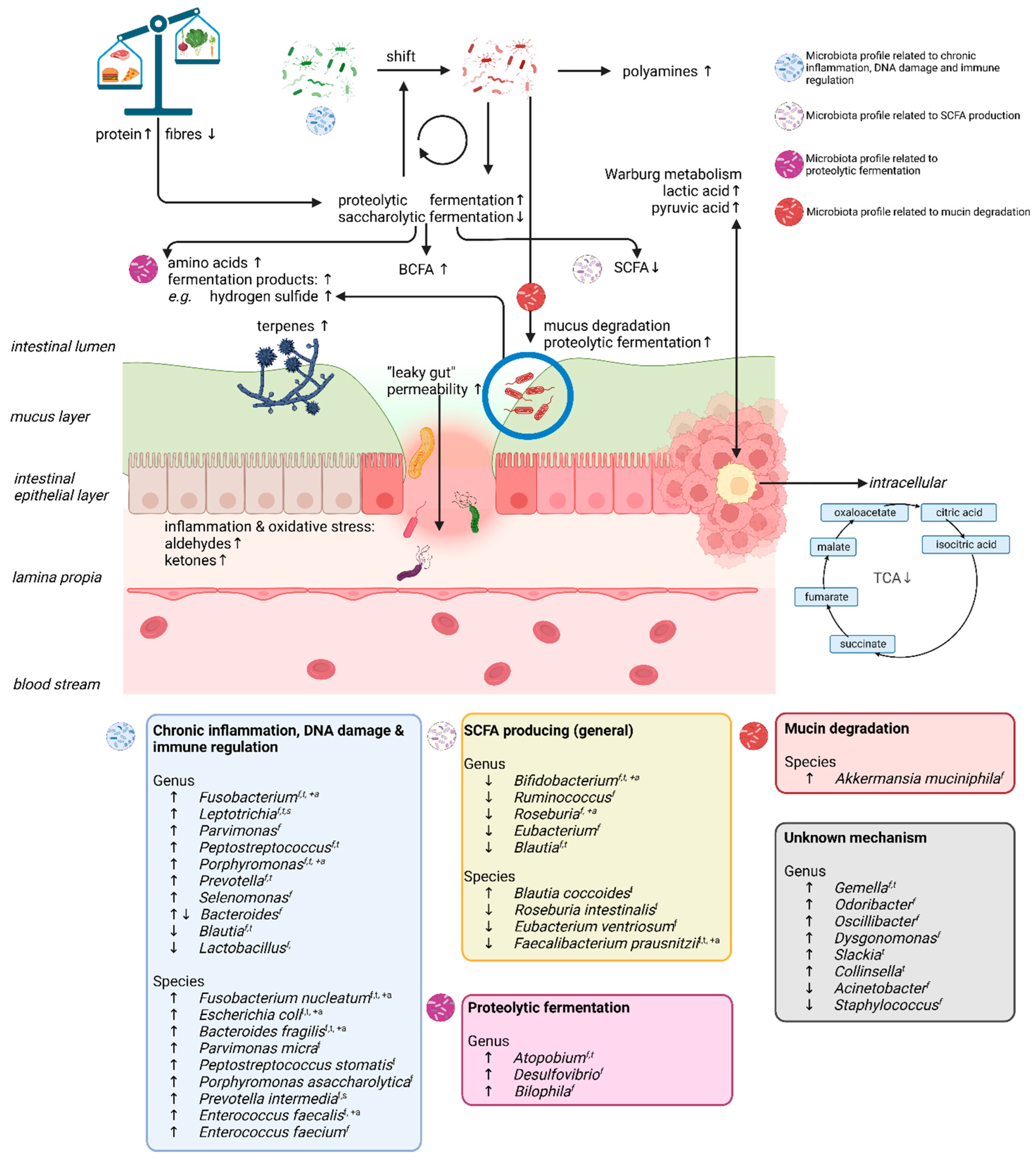
3.10. CRC-Associated Gut Microbiota
4. Discussion
4.1. SCFAs and Proteolytic Fermentation Products
4.2. Amino Acids
4.3. Tricarboxylic Acid Cycle and Warburg Metabolism
4.4. Aldehydes and Ketones
4.5. Terpenes and Furans
4.6. Alcohols and Other Bacterial Fermentation Products
5. Experimental Designs and Future Research
6. Limitations
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Safiri, S.; Sepanlou, S.G.; Ikuta, K.S.; Bisignano, C.; Salimzadeh, H.; Delavari, A.; Ansari, R.; Roshandel, G.; Merat, S.; Fitzmaurice, C.; et al. The global, regional, and national burden of colorectal cancer and its attributable risk factors in 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol. Hepatol. 2019, 4, 913–933. [Google Scholar] [CrossRef] [Green Version]
- Luengo-Fernandez, R.; Leal, J.; Gray, A.; Sullivan, R. Economic burden of cancer across the European Union: A population-based cost analysis. Lancet Oncol. 2013, 14, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, K.E.; Murphy, N.; Key, T.J. Diet and colorectal cancer in UK Biobank: A prospective study. Int. J. Epidemiol. 2020, 49, 246–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Chan, A.T. Environmental factors, gut microbiota, and colorectal cancer prevention. Clin. Gastroenterol. Hepatol. 2020, 17, 275–289. [Google Scholar] [CrossRef]
- Gacesa, R.; Kurilshikov, A.; Vich Vila, A.; Sinha, T.; Klaassen, M.A.Y.; Bolte, L.A.; Andreu-Sánchez, S.; Chen, L.; Collij, V.; Hu, S.; et al. Environmental factors shaping the gut microbiome in a Dutch population. Nature 2022, 604, 732–739. [Google Scholar] [CrossRef]
- Tilg, H.; Adolph, T.E.; Gerner, R.R.; Moschen, A.R. The Intestinal Microbiota in Colorectal Cancer. Cancer Cell 2018, 33, 954–964. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.H.; Yu, J. Gut microbiota in colorectal cancer: Mechanisms of action and clinical applications. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 690–704. [Google Scholar] [CrossRef]
- Nguyen, L.H.; Goel, A.; Chung, D.C. Pathways of Colorectal Carcinogenesis. Gastroenterology 2020, 158, 291–302. [Google Scholar] [CrossRef]
- Winawer, S.J.; Zauber, A.G.; Ho, M.N.; O’brien, M.J.; Gottlieb, L.S.; Sternberg, S.S.; Waye, J.D.; Schapiro, M.; Bond, J.H.; Panish, J.F.; et al. Prevention of Colorectal Cancer by Colonoscopic Polypectomy. N. Engl. J. Med. 1993, 329, 1977–1981. [Google Scholar] [CrossRef]
- Zauber, A.G.; Winawer, S.J.; O’Brien, M.J.; Lansdorp-Vogelaar, I.; Van Ballegooijen, M.; Hankey, B.F.; Shi, W.; Bond, J.H.; Schapiro, M.; Panish, J.F.; et al. Colonoscopic polypectomy and long-term prevention of colorectal-cancer deaths. N. Engl. J. Med. 2012, 366, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Gies, A.; Bhardwaj, M.; Stock, C.; Schrotz-King, P.; Brenner, H. Quantitative fecal immunochemical tests for colorectal cancer screening. Int. J. Cancer 2018, 143, 234–244. [Google Scholar] [CrossRef] [Green Version]
- Boots, A.W.; Bos, L.D.; van der Schee, M.P.; van Schooten, F.J.; Sterk, P.J. Exhaled Molecular Fingerprinting in Diagnosis and Monitoring: Validating Volatile Promises. Trends Mol. Med. 2015, 21, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Pauling, L.; Robinson, A.B.; Teranishi, R.; Cary, P. Quantitative analysis of urine vapor and breath by gas-liquid partition chromatography. Proc. Natl. Acad. Sci. USA 1971, 68, 2374–2376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amann, A.; Costello, B.D.L.; Miekisch, W.; Schubert, J.; Buszewski, B.; Pleil, J.; Ratcliffe, N.; Risby, T. The human volatilome: Volatile organic compounds (VOCs) in exhaled breath, skin emanations, urine, feces and saliva. J. Breath Res. 2014, 8, 034001. [Google Scholar] [CrossRef] [PubMed]
- Best, L.M.; Takwoingi, Y.; Siddique, S.; Selladurai, A.; Gandhi, A.; Low, B.; Yaghoobi, M.; Gurusamy, K.S. Non-invasive diagnostic tests for Helicobacter pylori infection. Cochrane Database Syst. Rev. 2018, 3, CD012080. [Google Scholar] [CrossRef] [Green Version]
- Rubin, R. First Breathalyzer Test to Diagnose COVID-19. JAMA 2022, 327, 1860. [Google Scholar] [CrossRef]
- Filipiak, W.; Mochalski, P.; Filipiak, A.; Ager, C.; Cumeras, R.; Davis, C.E.; Agapiou, A.; Unterkofler, K.; Troppmair, J. A Compendium of Volatile Organic Compounds (VOCs) Released By Human Cell Lines. Curr. Med. Chem. 2016, 23, 2112–2131. [Google Scholar] [CrossRef] [Green Version]
- Visconti, A.; Le Roy, C.I.; Rosa, F.; Rossi, N.; Martin, T.C.; Mohney, R.P.; Li, W.; de Rinaldis, E.; Bell, J.T.; Venter, J.C.; et al. Interplay between the human gut microbiome and host metabolism. Nat. Commun. 2019, 10, 4505. [Google Scholar] [CrossRef]
- Smolinska, A.; Tedjo, D.I.; Blanchet, L.; Bodelier, A.; Pierik, M.J.; Masclee, A.A.M.; Dallinga, J.; Savelkoul, P.H.M.; Jonkers, D.M.A.E.; Penders, J.; et al. Volatile metabolites in breath strongly correlate with gut microbiome in CD patients. Anal. Chim. Acta 2018, 1025, 1–11. [Google Scholar] [CrossRef]
- Zhou, W.; Tao, J.; Li, J.; Tao, S. Volatile organic compounds analysis as a potential novel screening tool for colorectal cancer: A systematic review and meta-analysis. Medicine 2020, 99, e20937. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Wu, S.; Hua, Q.; Bao, C.; Liu, H. Volatile Organic Compounds in Human Exhaled Breath to Diagnose Gastrointestinal Cancer: A Meta-Analysis. Front. Oncol. 2021, 11, 606915. [Google Scholar] [CrossRef] [PubMed]
- Di Lena, M.; Porcelli, F.; Altomare, D.F. Volatile organic compounds as new biomarkers for colorectal cancer: A review. Color Dis. 2016, 18, 654–663. [Google Scholar] [CrossRef]
- Monedeiro, F.; Monedeiro-milanowski, M.; Ligor, T.; Buszewski, B. A review of gc-based analysis of non-invasive biomarkers of colorectal cancer and related pathways. J. Clin. Med. 2020, 9, 3191. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; An, Y.; Qin, X.; Wu, X.; Wang, X.; Hou, H. Gut Microbiota-Derived Metabolites in Colorectal Cancer: The Bad and the Challenges. Front. Oncol. 2021, 11, 4287. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 10, 1–11. [Google Scholar] [CrossRef]
- Altomare, D.F.; Picciariello, A.; Rotelli, M.T.; De Fazio, M.; Aresta, A.; Zambonin, C.G.; Vincenti, L.; Trerotoli, P.; De Vietro, N. Chemical signature of colorectal cancer: Case-control study for profiling the breath print. BJS Open 2020, 4, 1189–1199. [Google Scholar] [CrossRef]
- Altomare, D.F.; Di Lena, M.; Porcelli, F.; Trizio, L.; Travaglio, E.; Tutino, M.; Dragonieri, S.; Memeo, V.; De Gennaro, G. Exhaled volatile organic compounds identify patients with colorectal cancer. Br. J. Surg. 2013, 100, 144–150. [Google Scholar] [CrossRef]
- Bel’skaya, L.V.; Sarf, E.A.; Shalygin, S.P.; Postnova, T.V.; Kosenok, V.K. Identification of salivary volatile organic compounds as potential markers of stomach and colorectal cancer: A pilot study. J. Oral Biosci. 2020, 62, 212–221. [Google Scholar] [CrossRef]
- Bond, A.; Greenwood, R.; Lewis, S.; Corfe, B.; Sarkar, S.; O’Toole, P.; Rooney, P.; Burkitt, M.; Hold, G.; Probert, C. Volatile organic compounds emitted from faeces as a biomarker for colorectal cancer. Aliment. Pharmacol. Ther. 2019, 49, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.G.; Rao, S.; Weir, T.L.; O’Malia, J.; Bazan, M.; Brown, R.J.; Ryan, E.P. Metabolomics and metabolic pathway networks from human colorectal cancers, adjacent mucosa, and stool. Cancer Metab. 2016, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.M.; Yu, Y.N.; Wang, J.L.; Lin, Y.W.; Kong, X.; Yang, C.Q.; Yang, L.; Liu, Z.J.; Yuan, Y.Z.; Liu, F.; et al. Decreased dietary fiber intake and structural alteration of gut microbiota in patients with advanced colorectal adenoma. Am. J. Clin. Nutr. 2013, 97, 1044–1052. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.L.; Fan, J.; Yan, L.S.; Guo, H.Q.; Xiong, J.J.; Ren, Y.; Hu, J.D. Urine metabolite profiling of human colorectal cancer by capillary electrophoresis mass spectrometry based on MRB. Gastroenterol. Res. Pract. 2012, 2012, 125890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Xie, G.; Chen, T.; Qiu, Y.; Zou, X.; Zheng, M.; Tan, B.; Feng, B.; Dong, T.; He, P.; et al. Distinct urinary metabolic profile of human colorectal cancer. J. Proteome Res. 2012, 11, 1354–1363. [Google Scholar] [CrossRef]
- Coker, O.O.; Liu, C.; Wu, W.K.K.; Wong, S.H.; Jia, W.; Sung, J.J.Y.; Yu, J. Altered gut metabolites and microbiota interactions are implicated in colorectal carcinogenesis and can be non-invasive diagnostic biomarkers. Microbiome 2022, 10, 35. [Google Scholar] [CrossRef]
- Cross, A.J.; Moore, S.C.; Boca, S.; Huang, W.-Y.; Xiong, X.; Stolzenberg-Solomon, R.; Sinha, R.; Sampson, J.N. A prospective study of serum metabolites and colorectal cancer risk. Cancer 2014, 120, 3049–3057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crotti, S.; Agnoletto, E.; Cancemi, G.; Di Marco, V.; Traldi, P.; Pucciarelli, S.; Nitti, D.; Agostini, M. Altered plasma levels of decanoic acid in colorectal cancer as a new diagnostic biomarker. Anal. Bioanal. Chem. 2016, 408, 6321–6328. [Google Scholar] [CrossRef]
- De Vietro, N.; Aresta, A.M.; Picciariello, A.; Rotelli, M.T.; Zambonin, C. Determination of VOCs in surgical resected tissues from colorectal cancer patients by solid phase microextraction coupled to gas chromatography–mass spectrometry. Appl. Sci. 2021, 11, 6910. [Google Scholar] [CrossRef]
- Altomare, D.F.; Di Lena, M.; Porcelli, F.; Travaglio, E.; Longobardi, F.; Tutino, M.; Depalma, N.; Tedesco, G.; Sardaro, A.; Memeo, R.; et al. Effects of curative colorectal cancer surgery on exhaled volatile organic compounds and potential implications in clinical follow-up. Ann. Surg. 2015, 262, 862–867. [Google Scholar] [CrossRef]
- De Vietro, N.; Aresta, A.; Rotelli, M.T.; Zambonin, C.; Lippolis, C.; Picciariello, A.; Altomare, D.F. Relationship between cancer tissue derived and exhaled volatile organic compound from colorectal cancer patients. Preliminary results. J. Pharm. Biomed. Anal. 2020, 180, 113055. [Google Scholar] [CrossRef] [PubMed]
- Denkert, C.; Budczies, J.; Weichert, W.; Wohlgemuth, G.; Scholz, M.; Kind, T.; Niesporek, S.; Noske, A.; Buckendahl, A.; Dietel, M.; et al. Metabolite profiling of human colon carcinoma—Deregulation of TCA cycle and amino acid turnover. Mol. Cancer 2008, 7, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Giovanni, N.; Meuwis, M.A.; Louis, E.; Focant, J.F. Specificity of metabolic colorectal cancer biomarkers in serum through effect size. Metabolomics 2020, 16, 88. [Google Scholar] [CrossRef] [PubMed]
- Farshidfar, F.; Weljie, A.M.; Kopciuk, K.A.; Hilsden, R.; McGregor, S.E.; Buie, W.D.; MacLean, A.; Vogel, H.J.; Bathe, O.F. A validated metabolomic signature for colorectal cancer: Exploration of the clinical value of metabolomics. Br. J. Cancer 2016, 115, 848–857. [Google Scholar] [CrossRef] [Green Version]
- Genua, F.; Mirković, B.; Mullee, A.; Levy, M.; Gallagher, W.M.; Vodicka, P.; Hughes, D.J. Association of circulating short chain fatty acid levels with colorectal adenomas and colorectal cancer. Clin. Nutr. ESPEN 2021, 46, 297–304. [Google Scholar] [CrossRef]
- Hassan, H.A.; Ammar, N.M.; Serag, A.; Shaker, O.G.; El Gendy, A.N.; Abdel-Hamid, A.H.Z. Metabolomics driven analysis of obesity-linked colorectal cancer patients via GC-MS and chemometrics: A pilot study. Microchem. J. 2020, 155, 104742. [Google Scholar] [CrossRef]
- Hou, X.; Hu, J.; Zhao, X.; Wei, Q.; Zhao, R.; Li, M.; Li, Q. Taurine Attenuates the Hypotaurine-Induced Progression of CRC via ERK/RSK Signaling. Front. Cell Dev. Biol. 2021, 9, 631163. [Google Scholar] [CrossRef]
- Ikeda, A.; Nishiumi, S.; Shinohara, M.; Yoshie, T.; Hatano, N.; Okuno, T.; Bamba, T.; Fukusaki, E.; Takenawa, T.; Azuma, T.; et al. Serum metabolomics as a novel diagnostic approach for gastrointestinal cancer. Biomed. Chromatogr. 2012, 26, 548–558. [Google Scholar] [CrossRef]
- Ishibe, A.; Ota, M.; Takeshita, A.; Tsuboi, H.; Kizuka, S.; Oka, H.; Suwa, Y.; Suzuki, S.; Nakagawa, K.; Suwa, H.; et al. Detection of gas components as a novel diagnostic method for colorectal cancer. Ann. Gastroenterol. Surg. 2018, 2, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Kibi, M.; Nishiumi, S.; Kobayashi, T.; Kodama, Y.; Yoshida, M. GC/MS and LC/MS-based tissue metabolomic analysis detected increased levels of antioxidant metabolites in colorectal cancer. Kobe J. Med. Sci. 2019, 65, E19–E27. [Google Scholar]
- Amal, H.; Leja, M.; Funka, K.; Lasina, I.; Skapars, R.; Sivins, A.; Ancans, G.; Kikuste, I.; Vanags, A.; Tolmanis, I.; et al. Breath testing as potential colorectal cancer screening tool. Int. J. Cancer 2016, 138, 229–236. [Google Scholar] [CrossRef]
- Kim, D.J.; Yang, J.; Seo, H.; Lee, W.H.; Lee, D.H.; Kym, S.; Park, Y.S.; Kim, J.G.; Jang, I.-J.; Kim, Y.-K.; et al. Colorectal cancer diagnostic model utilizing metagenomic and metabolomic data of stool microbial extracellular vesicles. Sci. Rep. 2020, 10, 2860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Yin, X.; Prodhan, M.A.I.; Zhang, X.; Zhong, Z.; Kato, I. Global plasma profiling for colorectal cancer-associated volatile organic compounds: A proof-of-principle study. J. Chromatogr. Sci. 2019, 57, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, R.V.; Suryawanshi, Y.R.; Essani, K. Nitrogen isotopes provide clues to amino acid metabolism in human colorectal cancer cells. Sci. Rep. 2017, 7, 2562. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Li, Y.; Wang, G.; Guo, N.; Liu, D.; Li, D.; Guo, L.; Zheng, X.; Yu, K.; Yu, K.; et al. Release of volatile organic compounds (VOCs) from colorectal cancer cell line LS174T. Anal. Biochem. 2019, 581, 113340. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Liu, W.; Peng, J.; Huang, L.; Zhang, P.; Zhao, X.; Cheng, Y.; Qin, H. A pilot study of gas chromatograph/mass spectrometry-based serum metabolic profiling of colorectal cancer after operation. Mol. Biol. Rep. 2010, 37, 1403–1411. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, P.; Wang, F.; Liu, W.; Yang, J.; Qin, H. An integrated proteomics and metabolomics approach for defining oncofetal biomarkers in the colorectal cancer. Ann. Surg. 2012, 255, 720–730. [Google Scholar] [CrossRef]
- Mal, M.; Koh, P.K.; Cheah, P.Y.; Chan, E.C.Y. Metabotyping of human colorectal cancer using two-dimensional gas chromatography mass spectrometry. Anal. Bioanal. Chem. 2012, 403, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Mal, M.; Koh, P.K.; Cheah, P.Y.; Chan, E.C.Y. Development and validation of a gas chromatography/mass spectrometry method for the metabolic profiling of human colon tissue. Rapid Commun. Mass Spectrom. 2009, 23, 487–494. [Google Scholar] [CrossRef]
- Markar, S.R.; Chin, S.; Romano, A.; Wiggins, T.; Antonowicz, S.; Paraskeva, P.; Ziprin, P.; Darzi, A.; Hanna, G.B. Breath Volatile Organic Compound Profiling of Colorectal Cancer Using Selected Ion Flow-tube Mass Spectrometry. Ann. Surg. 2019, 269, 903–910. [Google Scholar] [CrossRef]
- Miller-Atkins, G.; Acevedo-Moreno, L.-A.; Grove, D.; Dweik, R.A.; Tonelli, A.R.; Brown, J.M.; Allende, D.S.; Aucejo, F.; Rotroff, D.M. Breath Metabolomics Provides an Accurate and Noninvasive Approach for Screening Cirrhosis, Primary, and Secondary Liver Tumors. Hepatol. Commun. 2020, 4, 1041–1055. [Google Scholar] [CrossRef] [PubMed]
- Arasaradnam, R.P.; McFarlane, M.J.; Ryan-Fisher, C.; Westenbrink, E.; Hodges, P.; Thomas, M.G.; Chambers, S.; O’Connell, N.; Bailey, C.; Harmston, C.; et al. Detection of colorectal cancer (CRC) by urinary volatile organic compound analysis. PLoS ONE 2014, 9, e108750. [Google Scholar] [CrossRef]
- Niccolai, E.; Baldi, S.; Ricci, F.; Russo, E.; Nannini, G.; Menicatti, M.; Poli, G.; Taddei, A.; Bartolucci, G.; Calabrò, A.S.; et al. Evaluation and comparison of short chain fatty acids composition in gut diseases. World J. Gastroenterol. 2019, 25, 5543–5558. [Google Scholar] [CrossRef] [PubMed]
- Ning, W.; Li, H.; Meng, F.; Cheng, J.; Song, X.; Zhang, G.; Wang, W.; Wu, S.; Fang, J.; Ma, K.; et al. Identification of differential metabolic characteristics between tumor and normal tissue from colorectal cancer patients by gas chromatography–mass spectrometry. Biomed. Chromatogr. 2017, 31, e3999. [Google Scholar] [CrossRef]
- Ning, W.; Qiao, N.; Zhang, X.; Pei, D.; Wang, W. Metabolic profiling analysis for clinical urine of colorectal cancer. Asia Pac. J. Clin. Oncol. 2021, 17, 403–413. [Google Scholar] [CrossRef]
- Nishiumi, S.; Kobayashi, T.; Ikeda, A.; Yoshie, T.; Kibi, M.; Izumi, Y.; Okuno, T.; Hayashi, N.; Kawano, S.; Takenawa, T.; et al. A novel serum metabolomics-based diagnostic approach for colorectal cancer. PLoS ONE 2012, 7, e40459. [Google Scholar] [CrossRef] [PubMed]
- Nishiumi, S.; Kobayashi, T.; Kawana, S.; Unno, Y.; Sakai, T.; Okamoto, K.; Yamada, Y.; Sudo, K.; Yamaji, T.; Saito, Y.; et al. Investigations in the possibility of early detection of colorectal cancer by gas chromatography/triple-quadrupole mass spectrometry. Oncotarget 2017, 8, 17115–17126. [Google Scholar] [CrossRef] [Green Version]
- Nugent, J.L.; McCoy, A.N.; Addamo, C.J.; Jia, W.; Sandler, R.S.; Keku, T.O. Altered tissue metabolites correlate with microbial dysbiosis in colorectal adenomas. J. Proteome. Res. 2014, 13, 1921–1929. [Google Scholar] [CrossRef]
- Ong, E.S.; Zou, L.; Li, S.; Cheah, P.Y.; Eu, K.W.; Ong, C.N. Metabolic profiling in colorectal cancer reveals signature metabolic shifts during tumorigenesis. Mol. Cell. Proteom. 2010. [Google Scholar] [CrossRef]
- Peng, G.; Hakim, M.; Broza, Y.Y.; Billan, S.; Abdah-Bortnyak, R.; Kuten, A.; Tisch, U.; Haick, H. Detection of lung, breast, colorectal, and prostate cancers from exhaled breath using a single array of nanosensors. Br. J. Cancer 2010, 103, 542–551. [Google Scholar] [CrossRef]
- Phua, L.C.; Chue, X.P.; Koh, P.K.; Cheah, P.Y.; Ho, H.K.; Chan, E.C.Y. Non-invasive fecal metabonomic detection of colorectal cancer. Cancer Biol. Ther. 2014, 15, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Politi, L.; Monasta, L.; Rigressi, M.; Princivalle, A.; Gonfiotti, A.; Camiciottoli, G.; Perbellini, L. Discriminant profiles of volatile compounds in the alveolar air of patients with squamous cell lung cancer, lung adenocarcinoma or colon cancer. Molecules 2021, 26, 550. [Google Scholar] [CrossRef] [PubMed]
- Arshad, A.Z.; Munajat, Y.; Ghoshal, S.K.; Jamal, R.; Johdi, N.A.; Ibrahim, R.K.R.; Zulkhairi, H. Volatolomics combined terahertz time domain spectral analyses of colon cancer in vitro. J. Teknol. 2019, 81, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Porto-Figueira, P.; Pereira, J.A.M.; Câmara, J.S. Exploring the potential of needle trap microextraction combined with chromatographic and statistical data to discriminate different types of cancer based on urinary volatomic biosignature. Anal. Chim. Acta 2018, 1023, 53–63. [Google Scholar] [CrossRef]
- Qiu, Y.; Cai, G.; Su, M.; Chen, T.; Liu, Y.; Xu, Y.; Ni, Y.; Zhao, A.; Cai, S.; Xu, L.X.; et al. Urinary metabonomic study on colorectal cancer. J. Proteome. Res. 2010, 9, 1627–1634. [Google Scholar] [CrossRef]
- Qiu, Y.; Cai, G.; Su, M.; Chen, T.; Zheng, X.; Xu, Y.; Ni, Y.; Zhao, A.; Xu, L.X.; Cai, S.; et al. Serum metabolite profiling of human colorectal cancer using GC-TOFMS and UPLC-QTOFMS. J. Proteome. Res. 2009, 8, 4844–4850. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Cai, G.; Zhou, B.; Li, D.; Zhao, A.; Xie, G.; Li, H.; Cai, S.; Xie, D.; Huang, C.; et al. A distinct metabolic signature of human colorectal cancer with prognostic potential. Clin. Cancer Res. 2014, 20, 2136–2146. [Google Scholar] [CrossRef] [Green Version]
- Rozhentsov, A.A.; Koptina, A.V.; Mitrakov, N.N.; Sharipova, T.; Tsapaev, I.; Ryzhkov, V.L.; Lychagin, K.A.; Furina, R.R.; Mitrakova, N.N. A new method to diagnose cancer based on image analysis of mass chromatograms of volatile organic compounds in urine. Sovrem. Tehnol. V Med. 2014, 6, 151–157. [Google Scholar]
- Shu, X.; Xiang, Y.-B.; Rothman, N.; Yu, D.; Li, H.-L.; Yang, G.; Cai, H.; Ma, X.; Lan, Q.; Gao, Y.-T.; et al. Prospective study of blood metabolites associated with colorectal cancer risk. Int. J. Cancer 2018, 143, 527–534. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.L.; Passos, M.; Cmara, J.S. Investigation of urinary volatile organic metabolites as potential cancer biomarkers by solid-phase microextraction in combination with gas chromatography-mass spectrometry. Br. J. Cancer 2011, 105, 1894–1904. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.; Qiu, Y.; Zou, X.; Chen, T.; Xie, G.; Cheng, Y.; Dong, T.; Zhao, L.; Feng, B.; Hu, X.; et al. Metabonomics identifies serum metabolite markers of colorectal cancer. J. Proteome Res. 2013, 12, 3000–3009. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, H.; Daulton, E.; Bannaga, A.S.; Arasaradnam, R.P.; Covington, J.A. Non-invasive detection and staging of colorectal cancer using a portable electronic nose. Sensors 2021, 21, 5440. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, K.; Yagi, N.; Mizushima, K.; Higashimura, Y.; Hirai, Y.; Okayama, T.; Yoshida, N.; Katada, K.; Handa, O.; Ishikawa, T.; et al. Serum metabolomics analysis for early detection of colorectal cancer. J. Gastroenterol. 2017, 52, 677–694. [Google Scholar] [CrossRef]
- Baldi, S.; Menicatti, M.; Nannini, G.; Niccolai, E.; Russo, E.; Ricci, F.; Pallecchi, M.; Romano, F.; Pedone, M.; Poli, G.; et al. Free fatty acids signature in human intestinal disorders: Significant association between butyric acid and celiac disease. Nutrients 2021, 13, 742. [Google Scholar] [CrossRef]
- Wang, C.; Ke, C.; Wang, X.; Chi, C.; Guo, L.; Luo, S.; Guo, Z.; Xu, G.; Zhang, F.; Li, E. Noninvasive detection of colorectal cancer by analysis of exhaled breath. Anal. Bioanal. Chem. 2014, 406, 4757–4763. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, P.; Lian, A.; Sun, B.; Wang, X.; Guo, L.; Chi, C.; Liu, S.; Zhao, W.; Luo, S.; et al. Blood volatile compounds as biomarkers for colorectal cancer. Cancer Biol. Ther. 2014, 15, 200–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weir, T.L.; Manter, D.K.; Sheflin, A.M.; Barnett, B.A.; Heuberger, A.L.; Ryan, E.P. Stool Microbiome and Metabolome Differences between Colorectal Cancer Patients and Healthy Adults. PLoS ONE 2013, 8, e70803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Wu, M.; Wu, Q. Identification of potential metabolite markers for colon cancer and rectal cancer using serum metabolomics. J. Clin. Lab. Anal. 2020, 34, e23333. [Google Scholar] [CrossRef] [Green Version]
- Yachida, S.; Mizutani, S.; Shiroma, H.; Shiba, S.; Nakajima, T.; Sakamoto, T.; Watanabe, H.; Masuda, K.; Nishimoto, Y.; Kubo, M.; et al. Metagenomic and metabolomic analyses reveal distinct stage-specific phenotypes of the gut microbiota in colorectal cancer. Nat. Med. 2019, 25, 968–976. [Google Scholar] [CrossRef]
- Yang, Y.; Misra, B.; Liang, L.; Bi, D.; Weng, W.; Wu, W.; Cai, S.; Qin, H.; Goel, A.; Li, X.; et al. Integrated microbiome and metabolome analysis reveals a novel interplay between commensal bacteria and metabolites in colorectal cancer. Theranostics 2019, 9, 4101–4114. [Google Scholar] [CrossRef]
- Yuan, F.; Kim, S.; Yin, X.; Zhang, X.; Kato, I. Integrating two-dimensional gas and liquid chromatography-mass spectrometry for untargeted colorectal cancer metabolomics: A proof-of-principle study. Metabolites 2020, 10, 343. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, F.; Adewiah, S.; Syam, A.F.; Fatchiyah, F. Altered profile of gut microbiota and the level short chain fatty acids in colorectal cancer patients. J. Phys. Conf. Ser. 2019, 1146, 012037. [Google Scholar] [CrossRef]
- Zhu, G.; Wang, Y.; Wang, W.; Shang, F.; Pei, B.; Zhao, Y.; Kong, D.; Fan, Z. Untargeted GC-MS-Based Metabolomics for Early Detection of Colorectal Cancer. Front. Oncol. 2021, 11, 729512. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, D.; Hartmann, M.; Moyer, M.P.; Nolte, J.; Baumbach, J.I. Determination of volatile products of human colon cell line metabolism by GC/MS analysis. Metabolomics 2007, 3, 13–17. [Google Scholar] [CrossRef]
- Barberini, L.; Restivo, A.; Noto, A.; Deidda, S.; Fattuoni, C.; Fanos, V.; Saba, L.; Zorcolo, L.; Mussap, M. A gas chromatography-mass spectrometry (GC-MS) metabolomic approach in human colorectal cancer (CRC): The emerging role of monosaccharides and amino acids. Ann. Transl. Med. 2019, 7, 727. [Google Scholar] [CrossRef]
- Cheng, H.R.; van Vorstenbosch, R.W.; Pachen, D.M.; Meulen, L.W.; Straathof, J.W.A.; Dallinga, J.W.; Jonkers, D.M.; Masclee, A.A.; van Schooten, F.-J.; Mujagic, Z.; et al. Detecting Colorectal Adenomas and Cancer Via Volatile Organic Compounds in Exhaled Breath, a Proof of Principle Study to Improve Screening. Clin. Transl. Gastroenterol. 2022, 13, e00518. [Google Scholar] [CrossRef]
- Barberis, E.; Joseph, S.; Amede, E.; Clavenna, M.G.; La Vecchia, M.; Sculco, M.; Aspesi, A.; Occhipinti, P.; Robotti, E.; Boldorini, R.; et al. A new method for investigating microbiota-produced small molecules in adenomatous polyps. Anal. Chim. Acta 2021, 1179, 338841. [Google Scholar] [CrossRef]
- Batty, C.A.; Cauchi, M.; Lourenço, C.; Hunter, J.O.; Turner, C. Use of the analysis of the volatile faecal metabolome in screening for colorectal cancer. PLoS ONE 2015, 10, e0130301. [Google Scholar] [CrossRef] [Green Version]
- Aprile, F.; Bruno, G.; Palma, R.; Mascellino, M.; Panetta, C.; Scalese, G.; Oliva, A.; Severi, C.; Pontone, S. Microbiota alterations in precancerous colon lesions: A systematic review. Cancers 2021, 13, 3061. [Google Scholar] [CrossRef]
- Liu, H.; Wu, H.; Bilegsaikhan, E.; Lu, E.X.; Shen, X.; Liu, T. Differential expression of intestinal microbiota in colorectal cancer compared with healthy controls: A systematic review and meta-analysis. Int. J. Clin. Exp. Med. 2016, 9, 10923–10930. [Google Scholar]
- Huybrechts, I.; Zouiouich, S.; Loobuyck, A.; Vandenbulcke, Z.; Vogtmann, E.; Pisanu, S.; Iguacel, I.; Scalbert, A.; Indave, I.; Smelov, V.; et al. The human microbiome in relation to cancer risk: A systematic review of epidemiologic studies. Cancer Epidemiol. Biomark. Prev. 2020, 29, 1856–1868. [Google Scholar] [CrossRef] [PubMed]
- Amitay, E.L.; Krilaviciute, A.; Brenner, H. Systematic review: Gut microbiota in fecal samples and detection of colorectal neoplasms. Gut Microbes 2018, 9, 293–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges-canha, M.; Portela-cidade, J.P.; Dinis-ribeiro, M.; Leite-moreira, A.F.; Pimentel-nunes, P. Role of colonic microbiota in colorectal carcinogenesis: A systematic review. Rev. Española De Enferm. Dig. 2015, 107, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.C.S.; Huang, J.; Huang, J.L.W.; Pang, T.W.Y.; Choi, P.; Wang, J.; Chiang, J.I.; Jiang, J.Y. Global Prevalence of Colorectal Neoplasia: A Systematic Review and Meta-Analysis. Clin. Gastroenterol. Hepatol. 2020, 18, 553–561.e10. [Google Scholar] [CrossRef]
- Dai, Z.; Zhang, J.; Wu, Q.; Chen, J.; Liu, J.; Wang, L.; Chen, C.; Xu, J.; Zhang, H.; Shi, C.; et al. The role of microbiota in the development of colorectal cancer. Int. J. Cancer 2019, 145, 2032–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pleguezuelos-Manzano, C.; Puschhof, J.; Rosendahl Huber, A.; van Hoeck, A.; Wood, H.M.; Nomburg, J.; Gurjao, C.; Manders, F.; Dalmasso, G.; Stege, P.B.; et al. Mutational signature in colorectal cancer caused by genotoxic pks + E. coli. Nature 2020, 580, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Ríos-covián, D.; Ruas-madiedo, P.; Margolles, A.; Gueimonde, M.; Reyes-gavilán, C.G.D.L.; Salazar, N. Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [Green Version]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Jardon, K.M.; Canfora, E.E.; Goossens, G.H.; Blaak, E.E. Dietary macronutrients and the gut microbiome: A precision nutrition approach to improve cardiometabolic health. Gut 2022, 71, 1214–1226. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Dąbek-Drobny, A.; Kaczmarczyk, O.; Woźniakiewicz, M.; Paśko, P.; Dobrowolska-Iwanek, J.; Woźniakiewicz, A.; Piątek-Guziewicz, A.; Zagrodzki, P.; Zwolińska-Wcisło, M. Association between Fecal Short-Chain Fatty Acid Levels, Diet, and Body Mass Index in Patients with Inflammatory Bowel Disease. Biology 2022, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- de Jesús Rodríguez-Romero, J.; Durán-Castañeda, A.C.; Cárdenas-Castro, A.P.; Sánchez-Burgos, J.A.; Zamora-Gasga, V.M.; Sáyago-Ayerdi, G.S. What we know about protein gut metabolites: Implications and insights for human health and diseases. Food Chem. X 2022, 13, 100195. [Google Scholar] [CrossRef] [PubMed]
- Neis, E.P.J.G.; Dejong, C.H.C.; Rensen, S.S. The role of microbial amino acid metabolism in host metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef] [PubMed]
- Bansal, T.; Alaniz, R.C.; Wood, T.K.; Jayaraman, A. The bacterial signal indole increases epithelial-cell tight-junction resistance and attenuates indicators of inflammation. Proc. Natl. Acad. Sci. USA 2010, 107, 228–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieu, E.L.; Nguyen, T.; Rhyne, S.; Kim, J. Amino acids in cancer. Exp. Mol. Med. 2020, 52, 15–30. [Google Scholar] [CrossRef]
- Oliphant, K.; Allen-Vercoe, E. Macronutrient metabolism by the human gut microbiome: Major fermentation by-products and their impact on host health. Microbiome 2019, 7, 91. [Google Scholar] [CrossRef]
- Paone, P.; Cani, P.D. Mucus barrier, mucins and gut microbiota: The expected slimy partners? Gut 2020, 69, 2232–2243. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, F.; Liu, H.; Xie, Y.; Zhao, D.; Li, C. Transcriptomics and metabolomics reveal the adaption of Akkermansia muciniphila to high mucin by regulating energy homeostasis. Sci. Rep. 2021, 11, 9073. [Google Scholar] [CrossRef]
- Rebersek, M. Gut microbiome and its role in colorectal cancer. BMC Cancer 2021, 21, 1325. [Google Scholar] [CrossRef]
- Mohseni, A.H.; Taghinezhad-S, S.; Fu, X. Gut microbiota-derived metabolites and colorectal cancer: New insights and updates. Microb. Pathog. 2020, 149, 104569. [Google Scholar] [CrossRef]
- Tsuruya, A.; Kuwahara, A.; Saito, Y.; Yamaguchi, H.; Tenma, N.; Inai, M.; Takahashi, S.; Tsutsumi, E.; Suwa, Y.; Totsuka, Y.; et al. Major Anaerobic Bacteria Responsible for the Production of Carcinogenic Acetaldehyde from Ethanol in the Colon and Rectum. Alcohol Alcohol. 2016, 51, 395–401. [Google Scholar] [CrossRef] [Green Version]
- Ratcliffe, N.; Wieczorek, T.; Drabińska, N.; Drabińska, N.; Gould, O.; Osborne, A.; De Lacy Costello, B. A mechanistic study and review of volatile products from peroxidation of unsaturated fatty acids: An aid to understanding the origins of volatile organic compounds from the human body. J. Breath Res. 2020, 14, 034001. [Google Scholar] [CrossRef]
- den Besten, G.; Lange, K.; Havinga, R.; van Dijk, T.H.; Gerding, A.; van Eunen, K.; Müller, M.; Groen, A.K.; Hooiveld, G.J.; Bakker, B.M.; et al. Gut-derived short-chain fatty acids are vividly assimilated into host carbohydrates and lipids. Am. J. Physiol.-Gastrointest. Liver Physiol. 2013, 305, 900–910. [Google Scholar] [CrossRef]
- Koo, S.; Thomas, H.R.; Daniels, S.D.; Lynch, R.C.; Fortier, S.M.; Shea, M.M.; Rearden, P.; Comolli, J.C.; Baden, L.R.; Marty, F.M. A breath fungal secondary metabolite signature to diagnose invasive aspergillosis. Clin. Infect. Dis. 2014, 59, 1733–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heddergott, C.; Latgé, J.P.; Calvo, A.M. The volatome of Aspergillus fumigatus. Eukaryot. Cell 2014, 13, 1014–1025. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Gu, Y.; Liu, T.; Wang, C.; Zhong, W.; Wang, B.; Cao, H. Gut mycobiome: A promising target for colorectal cancer. Biochim. Biophys. Acta Rev. Cancer 2021, 1875, 188489. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, N.; Kounatidis, D.; Christodoulatos, G.S.; Panagopoulos, F.; Karampela, I.; Dalamaga, M. Mycobiome and cancer: What is the evidence? Cancers 2021, 13, 3149. [Google Scholar] [CrossRef] [PubMed]
- Nakatsu, G.; Zhou, H.; Wu, W.K.K.; Wong, S.H.; Coker, O.O.; Dai, Z.; Li, X.; Szeto, C.H.; Sugimura, N.; Lam, T.Y.T.; et al. Alterations in Enteric Virome Are Associated With Colorectal Cancer and Survival Outcomes. Gastroenterology 2018, 155, 529–541.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anandakumar, A.; Pellino, G.; Tekkis, P.; Kontovounisios, C. Fungal microbiome in colorectal cancer: A systematic review. Updates Surg. 2019, 71, 625–630. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, M.; Greten, F.R. The inflammatory pathogenesis of colorectal cancer. Nat. Rev. Immunol. 2021, 21, 653–667. [Google Scholar] [CrossRef] [PubMed]
- Van Der Lugt, T.; Venema, K.; Van Leeuwen, S.; Vrolijk, M.F.; Opperhuizen, A.; Bast, A. Gastrointestinal digestion of dietary advanced glycation endproducts using an: In vitro model of the gastrointestinal tract (TIM-1). Food Funct. 2020, 11, 6297–6307. [Google Scholar] [CrossRef] [PubMed]
- Blanchet, L.; Smolinska, A.; Baranska, A.; Tigchelaar, E.; Swertz, M.; Zhernakova, A.; Dallinga, J.W.; Wijmenga, C.; Van Schooten, F.J. Factors that influence the volatile organic compound content in human breath. J. Breath Res. 2017, 11, 016013. [Google Scholar] [CrossRef] [PubMed]
- Stegmüller, S.; Beißmann, N.; Kremer, J.I.; Mehl, D.; Baumann, C.; Richling, E. and 2-Methylfuran Metabolites in Human Urine Samples after Coffee Consumption. Molecules 2020, 25, 5104. [Google Scholar] [CrossRef] [PubMed]
- Hanna, G.B.; Boshier, P.R.; Markar, S.R.; Romano, A. Accuracy and Methodologic Challenges of Volatile Organic Compound-Based Exhaled Breath Tests for Cancer Diagnosis: A Systematic Review and Meta-analysis. JAMA Oncol. 2019, 5, e182815. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Chemical Class | VOC | Feces | Urine | Breath | Blood | Tissue | Saliva |
|---|---|---|---|---|---|---|---|
| SCFA | acetic acid | C, n = 5 | I, n = 2 | I, n = 2 | |||
| propanoic acid | C, n = 4 | n = 1 | I, n = 2 | ||||
| butyric acid | C, n = 7 | n = 1 | n = 1 | n = 1 | |||
| valeric acid | C, n = 2 | n = 1 | C, n = 2 | I, n = 2 | |||
| hexanoic acid | n = 1 | n = 1 | |||||
| heptanoic acid | I, n = 2 | ||||||
| octanoic acid | n = 1 | I, n = 3 | n = 1 | ||||
| nonanoic acid | C, n = 2 | ||||||
| total SCFA | n = 1 | ||||||
| BCFA | isobutyric acid | I, n = 4 | n = 1 | I, n = 2 | |||
| isovaleric acid | C, n = 3 | I, n = 2 | I, n = 2 | ||||
| ethyl 3-methylbutuanoate | n = 1 | ||||||
| hexanoic acid 2-methyl | n = 1 | ||||||
| 4-hydroxybutyrate | n = 1 | ||||||
| ethyl-acetate | n = 1 | n = 1 | |||||
| 2-methyl butyric acid | C, n = 2 | ||||||
| 3-hydroxyisovaleric acid | n = 1 | ||||||
| 2-aminoisobutyric acid | n = 1 | ||||||
| 2-aminobutyric acid | I, n = 2 | I, n = 2 | |||||
| 2-hydroxybutanoic acid | C, n = 7 | ||||||
| 2-hydroxy-pentanoic acid | |||||||
| 3-hydroxy butanoic acid | C, n = 6 | n = 1 | |||||
| 3-hydroxyisobutyric acid | n = 1 | ||||||
| 2-hydroxyisovaleric acid | n = 1 | ||||||
| 3-hydroxypropionic acid | n = 1 | ||||||
| Warburg metabolism | lactic acid | n = 1 | n = 1 | n = 1 | C, n = 7 | C, n = 5 | |
| pyruvate | n = 1 | C, n = 5 | n = 1 | ||||
| propyl-pyruvate | n = 1 | ||||||
| Amino acids | alanine | C, n = 4 | n = 1 | C, n = 4 | C, n = 3 | ||
| aspartic acid | C, n = 3 | n = 1 | I, n = 2 | C, n = 4 | |||
| valine | C, n = 4 | C, n = 2 | C, n = 8 | C, n = 5 | |||
| serine | C, n = 4 | n = 1 | n = 1 | C, n = 3 | |||
| proline | C, n = 3 | C, n = 5 | C, n = 7 | ||||
| glycine | C, n = 4 | n = 1 | C, n = 4 | C, n = 6 | |||
| phenylalanine | C, n = 2 | C, n = 5 | C, n = 6 | ||||
| leucine | C, n = 5 | n = 1 | C, n = 5 | C, n = 4 | |||
| threonine | n = 1 | I, n = 2 | C, n = 3 | ||||
| lysine | C, n = 2 | C, n = 2 | C, n = 3 | ||||
| isoleucine | C, n = 2 | C, n = 2 | C, n = 5 | C, n = 5 | |||
| citrulline | C, n = 1 | C, n = 3 | |||||
| tryptophan | C, n = 2 | C, n = 6 | n = 1 | ||||
| histidine | I, n = 3 | n = 1 | |||||
| tyrosine | n = 1 | C, n = 7 | C, n = 2 | ||||
| arginine | n = 1 | C, n = 1 | n = 1 | ||||
| ornithine | C, n = 5 | n = 1 | |||||
| cysteine | n = 1 | C, n = 5 | C, n = 2 | ||||
| beta-alanine | n = 1 | I, n = 3 | C, n = 5 | ||||
| aspargine | I, n = 3 | I, n = 2 | |||||
| glutamine | I, n = 2 | C, n = 2 | |||||
| Aldehydes | acetaldehyde | n = 1 | n = 1 | n = 1 | n = 1 | ||
| heptanal | n = 1 | n = 1 | n = 1 | ||||
| hexanal | I, n = 3 | n = 1 | |||||
| nonanal | n = 1 | I, n = 3 | n = 1 | ||||
| octanal | n = 1 | n = 1 | |||||
| butanal | n = 1 | ||||||
| undecanal | n = 1 | ||||||
| decanal | n = 1 | I, n = 4 | n = 1 | ||||
| propanal | n = 1 | ||||||
| pentanal | n = 1 | ||||||
| benzaldehyde | I, n = 4 | C, n = 2 | |||||
| (branched) Alkanes/enes | heptane | n = 1 | |||||
| octane | n = 1 | ||||||
| pentane | n = 1 | n = 1 | |||||
| nonane | I, n = 2 | ||||||
| decane | n = 1 | ||||||
| dodecane | I, n = 4 | ||||||
| tetradecane | I, n = 4 | ||||||
| isoprene | n = 1 | C, n = 2 | |||||
| 4-methyl octane | n = 1 | n = 1 | |||||
| 2,2,1-dimethyldecane | n = 1 | ||||||
| 2-methyl-butane | I, n = 2 | ||||||
| 2-methylpentane | I, n = 2 | ||||||
| 3-methyl-pentane | n = 1 | ||||||
| 3-ethyl hexane | n = 1 | ||||||
| 5-butyl-nonane | n = 1 | ||||||
| methylhexane | n = 1 | ||||||
| Sulfides | hydrogen sulfide | C, n = 2 | n = 1 | ||||
| dimethylsulfide | n = 1 | ||||||
| dimethyl disulfide | C, n = 2 | ||||||
| carbondisulphide | n = 1 | ||||||
| thiophene | n = 1 | ||||||
| tetrasulfide, dimethyl | I, n = 2 | ||||||
| dimethyl disulfide | I, n = 2 | ||||||
| dimethyl trisulfide | n = 1 | ||||||
| Furans | 2,4-dimethylfuran | n = 1 | |||||
| 2-acetylfuran | n = 1 | ||||||
| 2-ethyl-5-methylfuran | n = 1 | ||||||
| 2-methyl-5-(methylthio)furan | n = 1 | ||||||
| 2-methyl furan | n = 1 | ||||||
| 2-pentyl furan | n = 1 | n = 1 | |||||
| 3-methylfuran | n = 1 | ||||||
| furan | n = 1 | ||||||
| Ketones | hexan-2-one | n = 1 | |||||
| 2-heptanone | n = 1 | I, n = 2 | |||||
| 2-pentanone | I, n = 2 | n = 1 | |||||
| 3-heptanone | n = 1 | ||||||
| 3-hexanone | n = 1 | ||||||
| 4-heptanone | n = 1 | ||||||
| 2,4-dimethyl-3-pentone | I, n = 2 | ||||||
| 3-methyl-2-butanone | n = 1 | ||||||
| acetone | I, n = 2 | C, n = 4 | n = 1 | ||||
| 4-methyl-2-pentanone | I, n = 2 | ||||||
| 4-nonanone | n = 1 | ||||||
| 2-nonanone | I, n = 2 | ||||||
| Protein fermentation | ammonia | n = 1 | |||||
| tyramine | n = 1 | ||||||
| p-cymene | n = 1 | ||||||
| p-cresol | I, n = 3 | n = 1 | |||||
| triethylamine | n = 1 | ||||||
| trimethylamine | C, n = 2 | n = 1 | |||||
| indole | n = 1 | I, n = 2 | |||||
| phenol | n = 1 | I, n = 2 | n = 1 | n = 1 | |||
| phenyl lactic acid | C, n = 2 | ||||||
| phenylacetic acid | n = 1 | ||||||
| 2,4-di-tert-butylphenol | n = 1 | ||||||
| 3,5-di-t-butylphenol | n = 1 | ||||||
| 4-methyl-phenol | n = 1 | ||||||
| 4-tert-butylphenol | n = 1 | ||||||
| urea | C, n = 4 | ||||||
| uric acid | C, n = 3 | n = 1 | |||||
| taurine | I, n = 2 | ||||||
| cadaverine | n = 1 | ||||||
| indole acetate | n = 1 | ||||||
| 5-hydroxy-indoleacetate | n = 1 | ||||||
| 5-hydroxytryptophan | n = 1 | ||||||
| putrescine | n = 1 | ||||||
| Bacterial fermentation | di-nitrogen oxide | n = 1 | |||||
| propanol | n = 1 | ||||||
| propan-2-ol | n = 1 | n = 1 | |||||
| propan-2-ul pentanoate | n = 1 | ||||||
| propan-2-yl butanoate | n = 1 | ||||||
| propan-2-yl propanoate | n = 1 | ||||||
| ethanolamine | n = 1 | ||||||
| methyl mercaptan | n = 1 | n = 1 | |||||
| ethanol | n = 1 | n = 1 | |||||
| choline | C, n = 2 | ||||||
| norvaline | n = 1 | ||||||
| Terpenes | xylene | n = 1 | n = 1 | ||||
| g-terpinene | n = 1 | ||||||
| beta-pinene | n = 1 | n = 1 | |||||
| TCA metabolism | maltose | n = 1 | n = 1 | I, n = 2 | |||
| fructose | C, n = 2 | n = 1 | |||||
| malate | n = 1 | n = 1 | C, n = 3 | C, n = 3 | |||
| oxalic acid | n = 1 | n = 1 | n = 1 | n = 1 | |||
| succinic acid | C, n = 2 | C, n = 3 | n = 1 | n = 1 | |||
| citrate | C, n = 2 | I, n = 4 | n = 1 | ||||
| glucose | n = 1 | I n = 2 | C, n = 5 | ||||
| sorbose | n = 1 | n = 1 | |||||
| xylose | n = 1 | n = 1 | |||||
| arabitol | n = 1 | n = 1 | |||||
| glucuronate | I, n = 2 | n = 1 | |||||
| xylitol | I, n = 2 | ||||||
| arabinose | I, n = 3 | n = 1 | |||||
| isocitric acid | I, n = 2 | I, n = 2 | |||||
| sucrose | n = 1 | ||||||
| threitol | n = 1 | ||||||
| fumaric acid | n = 1 | n = 1 | I, n = 2 | n = 1 | |||
| glutamic acid | n = 1 | n = 1 | I, n = 7 | C, n = 5 | |||
| glycolic acid | I, n = 2 | ||||||
| D-mannose | C, n = 2 | C, n = 4 | |||||
| ribitol | n = 1 | ||||||
| ribulose | n = 1 | ||||||
| glycerol | I, n = 2 | C, n = 4 | |||||
| inositol | n = 1 | C, n = 3 | |||||
| sorbitol | n = 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Vorstenbosch, R.; Cheng, H.R.; Jonkers, D.; Penders, J.; Schoon, E.; Masclee, A.; van Schooten, F.-J.; Smolinska, A.; Mujagic, Z. Systematic Review: Contribution of the Gut Microbiome to the Volatile Metabolic Fingerprint of Colorectal Neoplasia. Metabolites 2023, 13, 55. https://doi.org/10.3390/metabo13010055
van Vorstenbosch R, Cheng HR, Jonkers D, Penders J, Schoon E, Masclee A, van Schooten F-J, Smolinska A, Mujagic Z. Systematic Review: Contribution of the Gut Microbiome to the Volatile Metabolic Fingerprint of Colorectal Neoplasia. Metabolites. 2023; 13(1):55. https://doi.org/10.3390/metabo13010055
Chicago/Turabian Stylevan Vorstenbosch, Robert, Hao Ran Cheng, Daisy Jonkers, John Penders, Erik Schoon, Ad Masclee, Frederik-Jan van Schooten, Agnieszka Smolinska, and Zlatan Mujagic. 2023. "Systematic Review: Contribution of the Gut Microbiome to the Volatile Metabolic Fingerprint of Colorectal Neoplasia" Metabolites 13, no. 1: 55. https://doi.org/10.3390/metabo13010055