

Chemodiversity and Anti-Leukemia Effect of Metabolites from Penicillium setosum CMLD 18
 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Experimental Design
Equipment
3. Procedure
3.1. Biological Assays
3.1.1. Stock Solutions
3.1.2. Cell Cultures
3.1.3. Peripheral Blood Mononuclear Isolation
3.1.4. Resazurin Assay
3.1.5. Annexin V/7-AAD Flow Cytometry Assay
3.1.6. Intracellular Protein Labeling
3.1.7. Cell Cycle Analysis
3.1.8. Cell Differentiation by Immunophenotyping
3.1.9. Statistical Analyses
3.2. Isolation of Penicillium setosum from Swinglea glutinosa
3.3. Fungi Cultivation and Initial Characterization
3.3.1. DNA Extraction, PCR, and Sequencing
3.3.2. Phylogenetic Analysis and Identification
3.3.3. Penicillium setosum Cultivation in Hominy (M1) and Rice (M2)
3.3.4. Penicillium setosum Cultivation in CYB (M3) and YES (M4)
3.3.5. Penicillium setosum Cultivation in Yes + Agar (M6)
3.4. Compounds Isolation
4. Results
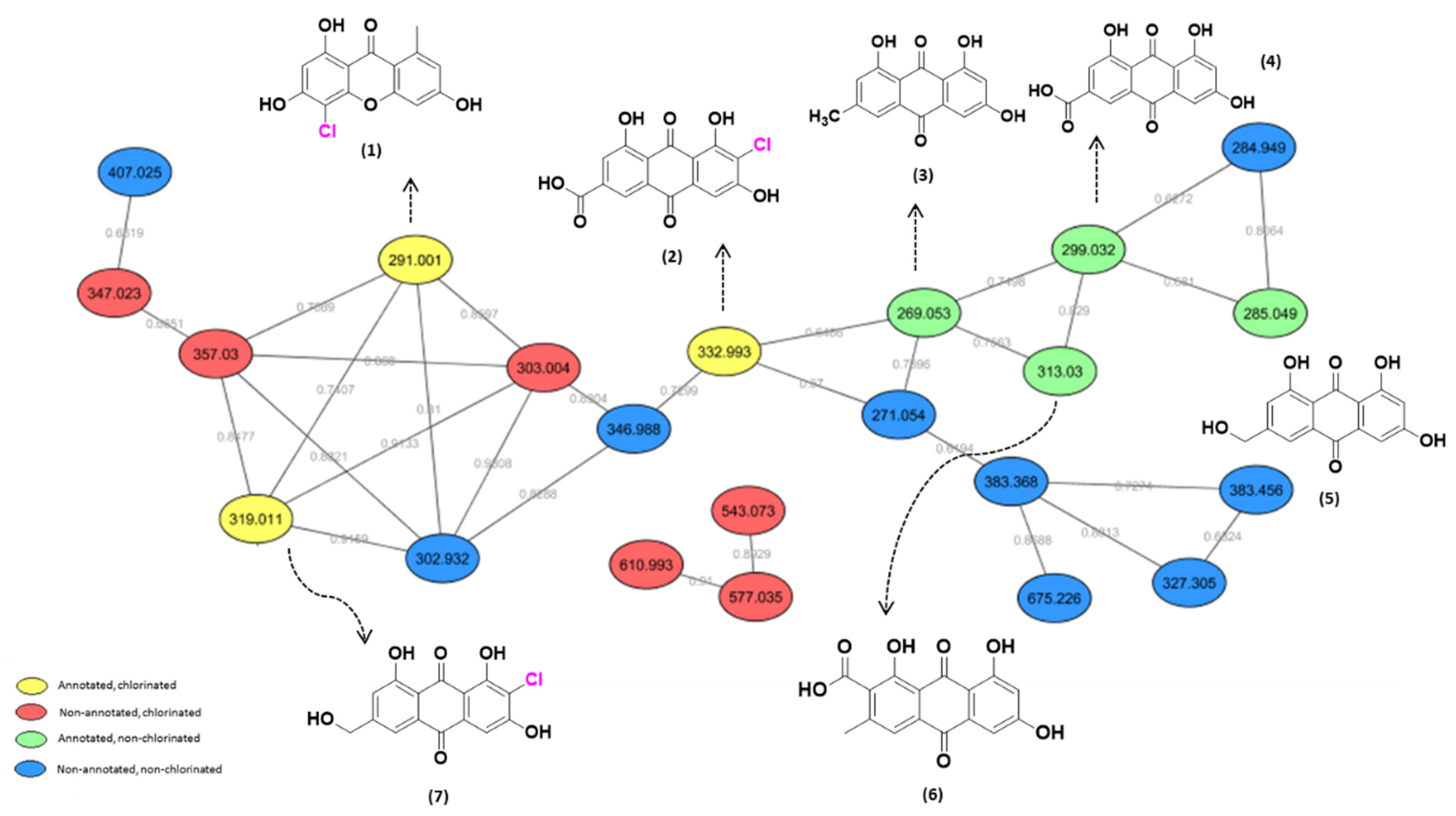
4.1. Exploring the Chemodiversity of P. setosum
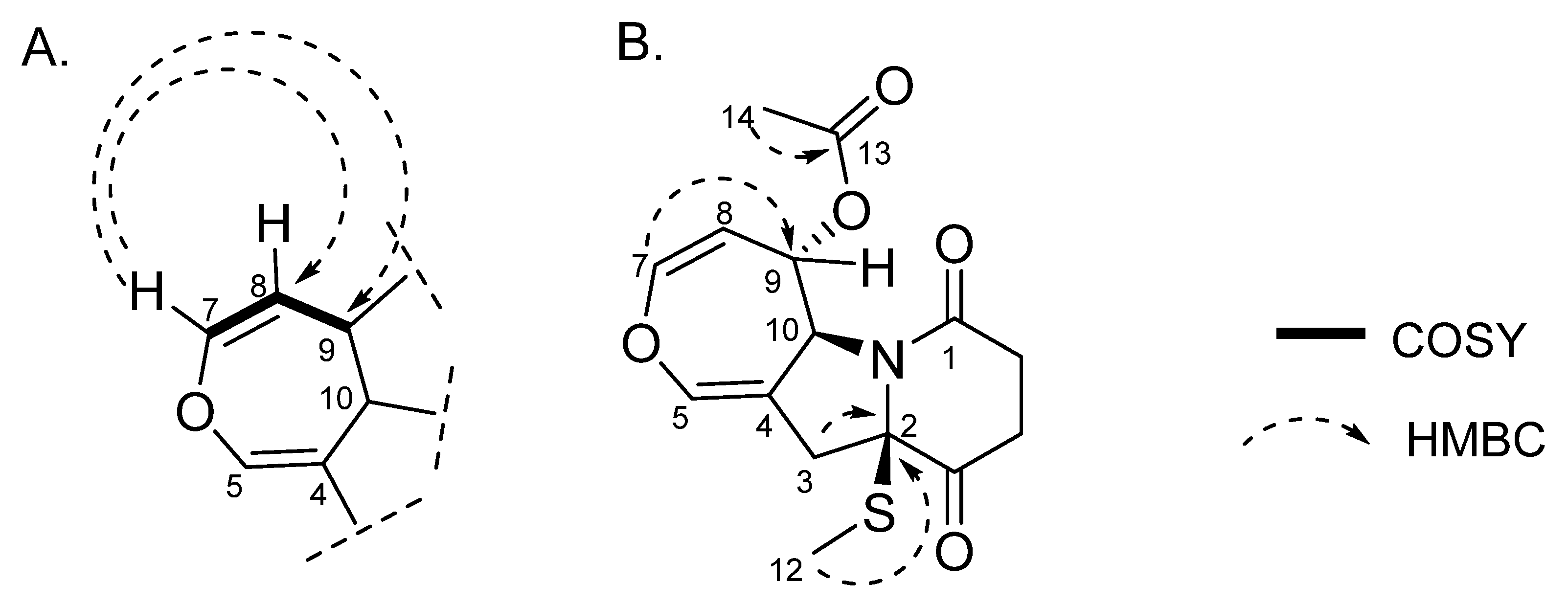
4.2. The Isolation and Characterization of Metabolites from P. setosum
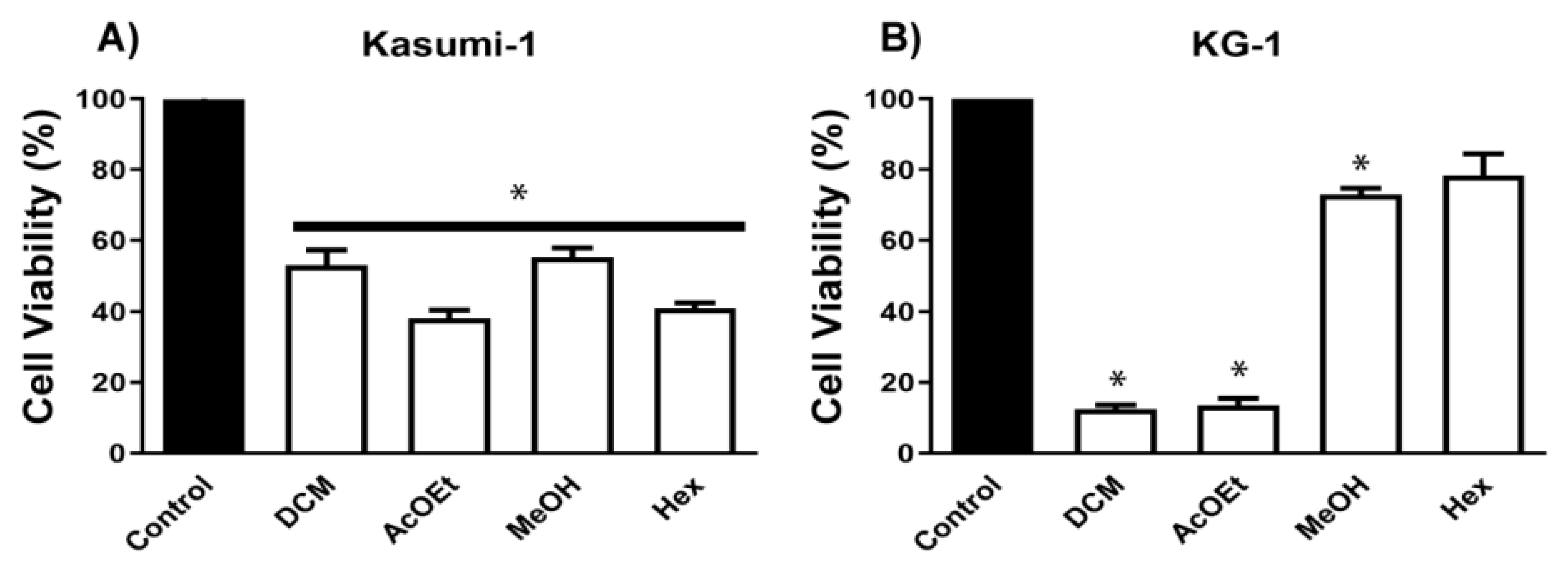
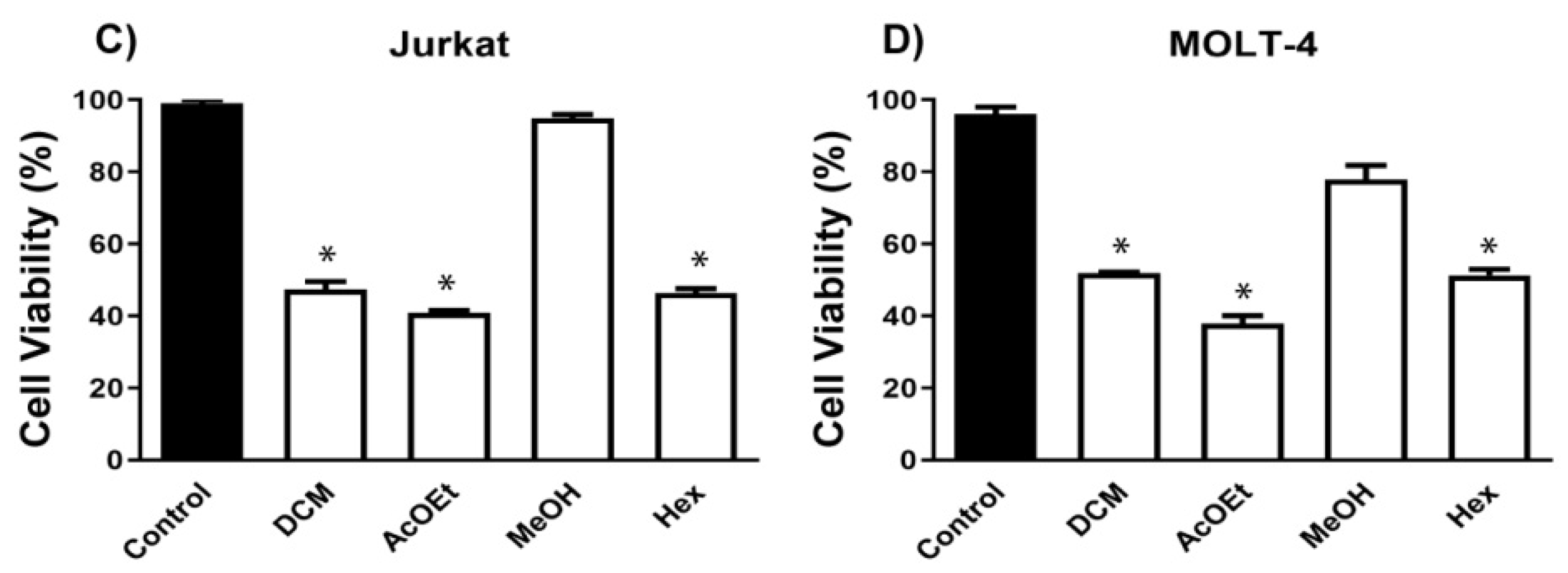
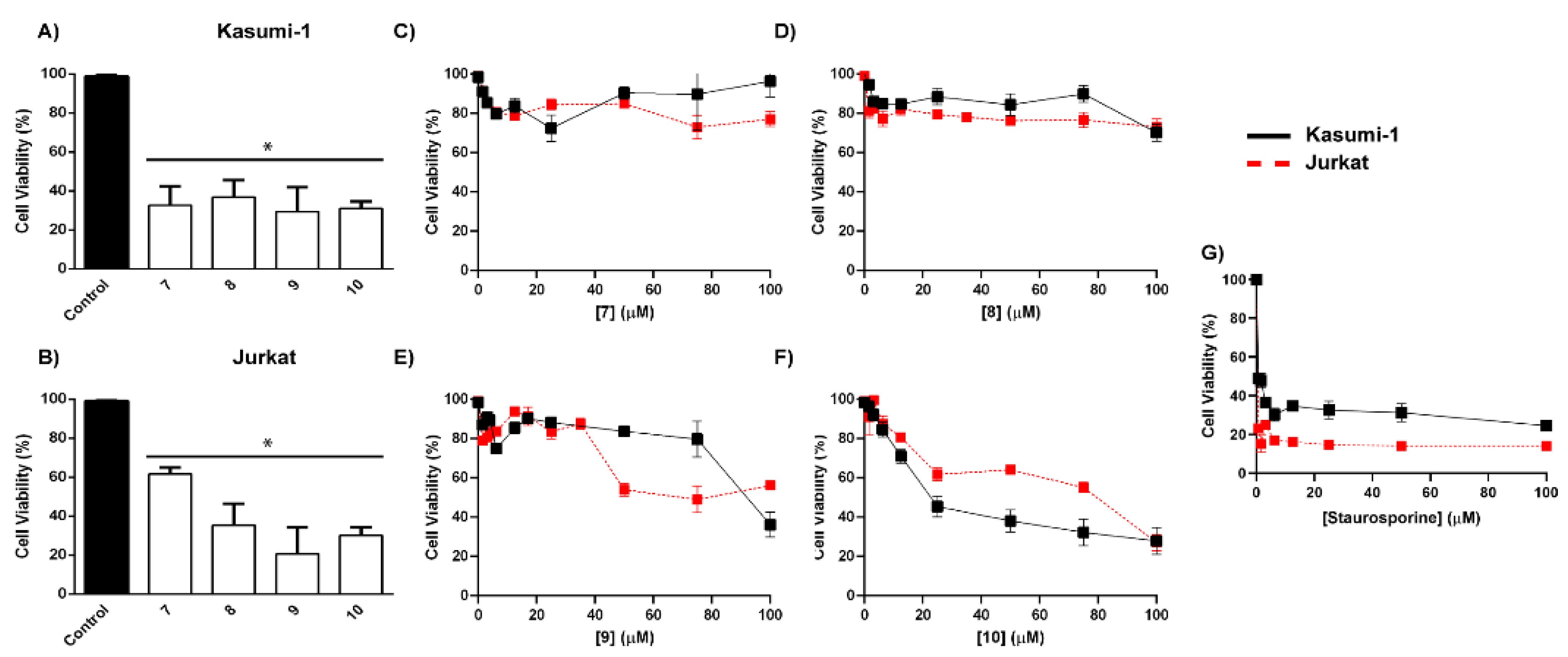
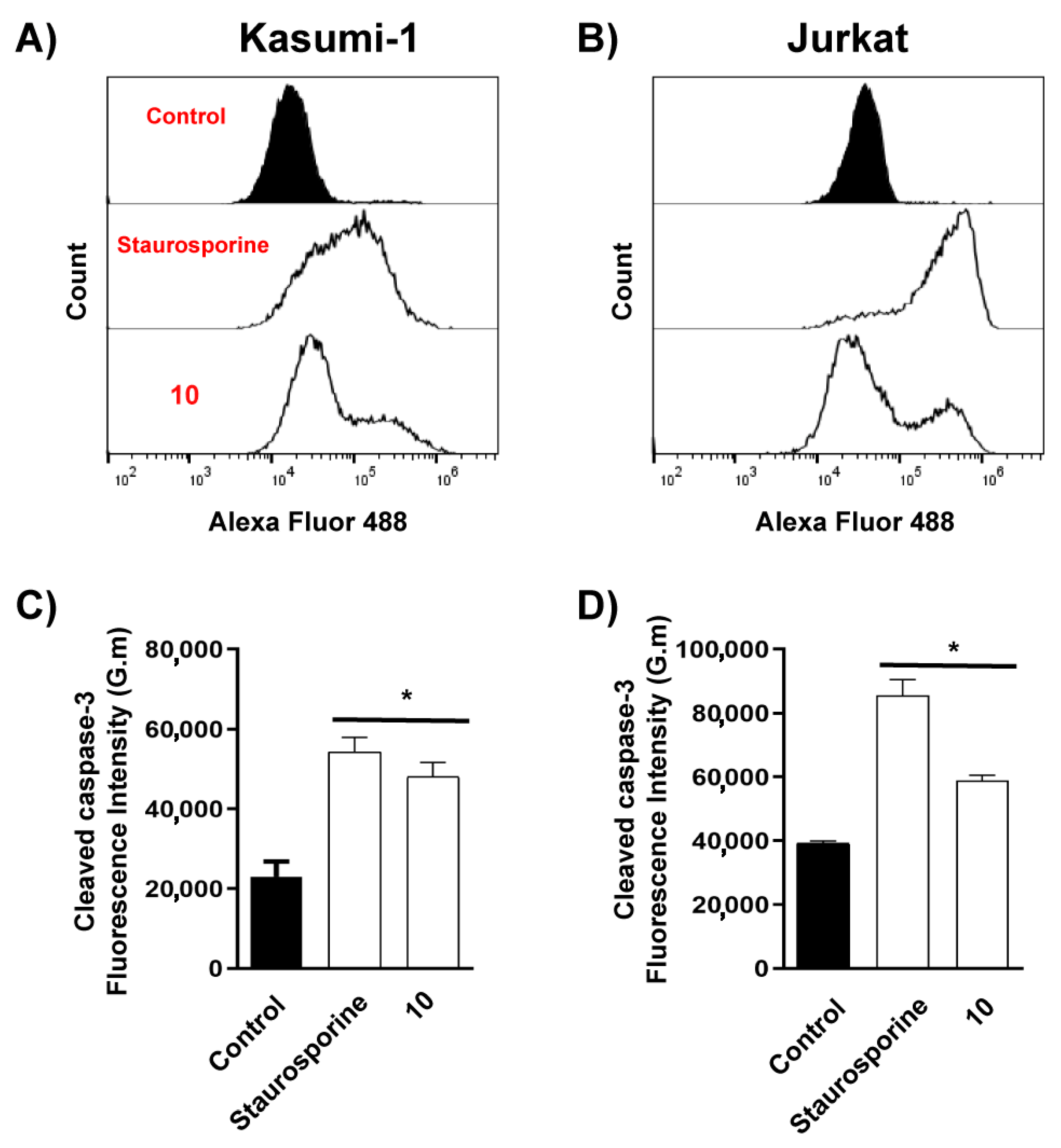
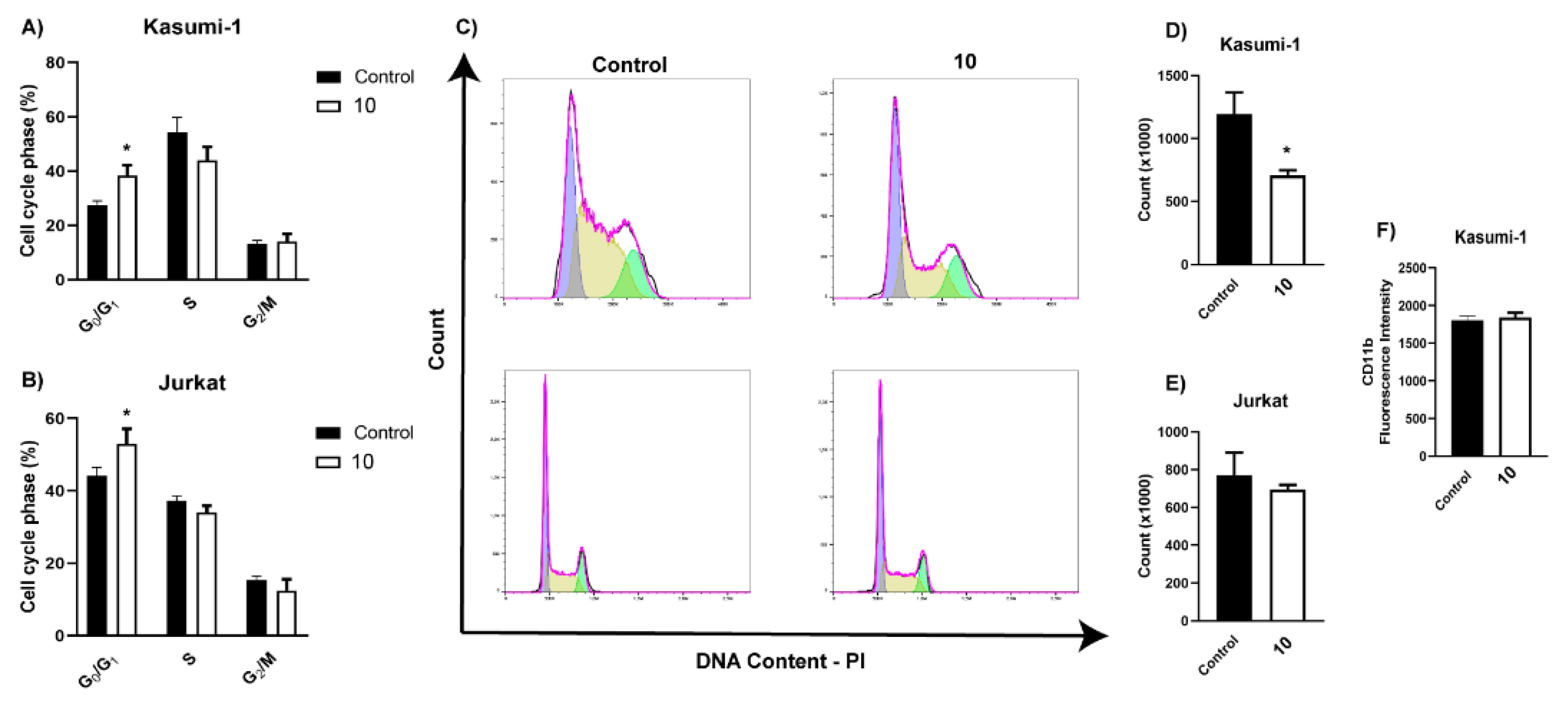
4.3. Biological Activity
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Egbuta, M.A.; Mwanza, M.; Babalola, O.O. A review of the ubiquity of ascomycetes filamentous fungi in relation to their economic and medical importance. Adv. Microbiol. 2016, 06, 1140–1158. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho, A.C.; Ogawa, C.Y.; De Camillis Rodrigues, L.; de Medeiros, L.S.; Veiga, T.A.M. Penicillium genus as a source for anti-leukemia compounds: An overview from 1984 to 2020. Leuk. Lymphoma 2021, 62, 2079–2093. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wei, Q.; Yuan, X.; Xu, K. Newly reported alkaloids produced by marine-derived Penicillium species (covering 2014–2018). Bioorg. Chem. 2020, 99, 103840. [Google Scholar] [CrossRef] [PubMed]
- Koul, M.; Singh, S. Penicillium spp.: Prolific producer for harnessing cytotoxic secondary metabolites. Anticancer Drugs 2017, 28, 11–30. [Google Scholar] [CrossRef] [PubMed]
- George, T.K.; Houbraken, J.; Mathew, L.; Jisha, M.S. Penicillium setosum, a new species from Withania somnifera (L.) Dunal. Mycology 2019, 10, 49–60. [Google Scholar] [CrossRef] [Green Version]
- George, T.K.; Devadasan, D.; Jisha, M.S. Chemotaxonomic profiling of Penicillium setosum using high-resolution mass spectrometry (LC-Q-ToF-MS). Heliyon 2019, 5, e02484. [Google Scholar] [CrossRef] [Green Version]
- Crüsemann, M. Coupling mass spectral and genomic information to improve bacterial natural product discovery workflows. Mar. Drugs 2021, 19, 142. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://gnps.ucsd.edu/ProteoSAFe/static/gnps-splash.jsp (accessed on 16 April 2022).
- Petrini, O.; Sieber, T.N.; Toti, L.; Viret, O. Ecology, metabolite production, and substrate utilization in endophytic fungi. Nat. Toxins 1992, 1, 185–196. [Google Scholar] [CrossRef]
- Stielow, J.B.; Lévesque, C.A.; Seifert, K.A.; Meyer, W.; Iriny, L.; Smits, D.; Renfurm, R.; Verkley, G.J.M.; Groenewald, M.; Chaduli, D.; et al. One fungus, which genes? Development and assessment of universal primers for potential secondary fungal DNA barcodes. Persoonia 2015, 35, 242–263. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. Chembiochem 2002, 3, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.A.; Towers, G.H.N. Anthraquinones and Phenanthroperylenequinones from Nephroma laevigatum. J. Nat. Prod. 1995, 58, 520–526. [Google Scholar] [CrossRef]
- Figueroa, M.; Jarmusch, A.K.; Raja, H.A.; El-Elimat, T.; Kavanaugh, J.S.; Horswill, A.R.; Cooks, R.G.; Cech, N.B.; Oberlies, N.H. Polyhydroxyanthraquinones as quorum sensing inhibitors from the guttates of Penicillium restrictum and their analysis by desorption electrospray ionization mass spectrometry. J. Nat. Prod. 2014, 77, 1351–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, P.A.; Towers, G.N. Biosynthetic studies on chlorinated anthraquinones in the lichen Nephroma laevigatum. Phytochemistry 1996, 42, 1325–1329. [Google Scholar] [CrossRef]
- Louis, I.; Hungerford, N.L.; Humphries, E.J.; McLeod, M.D. Enantioselective total synthesis of (−)-dactylolide. Org. Lett. 2006, 8, 1117–1120. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Cao, F.; Yu, H.; Zhu, H.-J. Secondary Metabolites from the Marine Fungus Aspergillus sydowii. Chem. Nat. Compd. 2017, 53, 1204–1207. [Google Scholar] [CrossRef]
- Kozlovsky, A.G.; Vinokurova, N.G.; Adanin, V.M.; Burkhardt, G.; Dahse, H.M.; Gräfe, U. New diketopiperazine alkaloids from Penicillium fellutanum. J. Nat. Prod. 2000, 63, 698–700. [Google Scholar] [CrossRef]
- Nagarajan, R.; Huckstep, L.L.; Lively, D.H.; DeLong, D.C.; Marsh, M.M.; Neuss, N. Aranotin and related metabolites from Arachniotus aureus. I. Determination of structure. J. Am. Chem. Soc. 1968, 90, 2980–2982. [Google Scholar] [CrossRef]
- Antonsson, A.; Persson, J.L. Induction of apoptosis by staurosporine involves the inhibition of expression of the major cell cycle proteins at the G(2)/m checkpoint accompanied by alterations in Erk and Akt kinase activities. Anticancer Res. 2009, 29, 2893–2898. [Google Scholar]
- El-Elimat, T.; Figueroa, M.; Ehrmann, B.M.; Cech, N.B.; Pearce, C.J.; Oberlies, N.H. High-resolution MS, MS/MS, and UV database of fungal secondary metabolites as a dereplication protocol for bioactive natural products. J. Nat. Prod. 2013, 76, 1709–1716. [Google Scholar] [CrossRef] [Green Version]
- Freire, V.F.; Gubiani, J.R.; Spencer, T.M.; Hajdu, E.; Ferreira, A.G.; Ferreira, D.A.S.; de Castro Levatti, E.V.; Burdette, J.E.; Camargo, C.H.; Tempone, A.G.; et al. Feature-Based Molecular Networking Discovery of Bromopyrrole Alkaloids from the Marine Sponge Agelas dispar. J. Nat. Prod. 2022, 85, 1340–1350. [Google Scholar] [CrossRef] [PubMed]
- Avalon, N.E.; Murray, A.E.; Baker, B.J. Integrated Metabolomic-Genomic Workflows Accelerate Microbial Natural Product Discovery. Anal. Chem. 2022, 94, 11959–11966. [Google Scholar] [CrossRef] [PubMed]
- E Silva, J.P.R.; Pereira, L.C.O.; Abreu, L.S.; Lins, F.S.V.; de Souza, T.A.; do Espírito-Santo, R.F.; Barros, R.P.C.; Villarreal, C.F.; de Melo, J.I.M.; Scotti, M.T.; et al. Targeted Isolation of Anti-inflammatory Lignans from Justicia aequilabris by Molecular Networking Approach. J. Nat. Prod. 2022, 85, 2184–2191. [Google Scholar] [CrossRef] [PubMed]
- Ramatapa, T.; Msobo, A.; Maphari, P.W.; Ncube, E.N.; Nogemane, N.; Mhlongo, M.I. Identification of Plant-Derived Bioactive Compounds Using Affinity Mass Spectrometry and Molecular Networking. Metabolites 2022, 12, 863. [Google Scholar] [CrossRef]
- Luo, M.; Chang, S.; Li, Y.; Xi, X.; Chen, M.; He, N.; Wang, M.; Zhao, W.; Xie, Y. Molecular Networking-Based Screening Led to the Discovery of a Cyclic Heptadepsipeptide from an Endolichenic Xylaria sp. J. Nat. Prod. 2022, 85, 972–979. [Google Scholar] [CrossRef]
- Paguigan, N.D.; El-Elimat, T.; Kao, D.; Raja, H.A.; Pearce, C.J.; Oberlies, N.H. Enhanced dereplication of fungal cultures via use of mass defect filtering. J. Antibiot. 2017, 70, 553–561. [Google Scholar] [CrossRef] [Green Version]
- Calheiros de Carvalho, A.; De Camillis Rodrigues, L.; Ribeiro, A.I.; Fernandes da Silva, M.F.d.G.; Soman de Medeiros, L.; Moura Veiga, T.A. Integrated Analytical Tools for Accessing Acridones and Unrelated Phenylacrylamides from Swinglea glutinosa. Molecules 2019, 25, 153. [Google Scholar] [CrossRef] [Green Version]
- Marcucci, M.C.; Oliveira, C.R.; Spindola, D.; Antunes, A.A.; Santana, L.Y.K.; Cavalaro, V.; Costa, I.B.; de Carvalho, A.C.; Veiga, T.A.M.; Medeiros, L.S.; et al. Molecular Dereplication and In Vitro and In Silico Pharmacological Evaluation of Coriandrum sativum against Neuroblastoma Cells. Molecules 2022, 27, 5389. [Google Scholar] [CrossRef]
- Shour, S.; Iranshahy, M.; Pham, N.; Quinn, R.J.; Iranshahi, M. Dereplication of cytotoxic compounds from different parts of Sophora pachycarpa using an integrated method of HPLC, LC-MS and 1H-NMR techniques. Nat. Prod. Res. 2017, 31, 1270–1276. [Google Scholar] [CrossRef]
- Shishido, T.K.; Popin, R.V.; Jokela, J.; Wahlsten, M.; Fiore, M.F.; Fewer, D.P.; Herfindal, L.; Sivonen, K. Dereplication of Natural Products with Antimicrobial and Anticancer Activity from Brazilian Cyanobacteria. Toxins 2019, 12, 12. [Google Scholar] [CrossRef] [Green Version]
- Kushwaha, M.; Qayum, A.; Sharma, N.; Abrol, V.; Choudhary, P.; Murtaza, M.; Singh, S.K.; Vishwakarma, R.A.; Goutam, U.; Jain, S.K.; et al. LC–PDA–MS/MS-Based Dereplication Guided Isolation of a New Optical Isomer of 19, 20-Epoxycytochalasin-N and Its Cytotoxic Activity. ACS Omega 2022, 7, 29135–29141. [Google Scholar] [CrossRef] [PubMed]
- Geiger, M.; Desanglois, G.; Hogeveen, K.; Fessard, V.; Leprêtre, T.; Mondeguer, F.; Guitton, Y.; Hervé, F.; Séchet, V.; Grovel, O.; et al. Cytotoxicity, fractionation and dereplication of extracts of the dinoflagellate Vulcanodinium rugosum, a producer of pinnatoxin G. Mar. Drugs 2013, 11, 3350–3371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, K.; Kato, T.; Mori, H.; Kitamura, J.; Nozawa, Y. A comparative study on cytotoxicities and biochemical properties of anthraquinone mycotoxins emodin and skyrin from Penicillium islandicum Sopp. Toxicol. Lett. 1984, 20, 155–160. [Google Scholar] [CrossRef]
- Bai, J.; Guo, F.; Wang, R.; Chen, G.; Li, Z.; Shao, M.; Xue, C.; Hua, H. Asperterzine, a symmetric aromatized derivative of epipolythiodioxopiperazine, from the endophytic fungus Aspergillus terreus PR-P-2. Chin. Chem. Lett. 2018, 29, 535–537. [Google Scholar] [CrossRef]
- Koerner, S.K.; Hanai, J.-I.; Bai, S.; Jernigan, F.E.; Oki, M.; Komaba, C.; Shuto, E.; Sukhatme, V.P.; Sun, L. Design and synthesis of emodin derivatives as novel inhibitors of ATP-citrate lyase. Eur. J. Med. Chem. 2017, 126, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Cui, Z.; Huang, H.; Song, X.; Sun, A.; Dang, Y.; Lu, L.; Ju, J. Amino Acid Conjugated Anthraquinones from the Marine-Derived Fungus Penicillium sp. SCSIO sof101. J. Nat. Prod. 2017, 80, 1668–1673. [Google Scholar] [CrossRef]
- George, T.K.; Joy, A.; Divya, K.; Jisha, M.S. In vitro and in silico docking studies of antibacterial compounds derived from endophytic Penicillium setosum. Microb. Pathog. 2019, 131, 87–97. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Structure | m/z | Found in the Culture Media |
|---|---|---|---|
| 4-chloro-norliquexanthone (1) |  | 291.001 | M6 |
| 2-chloroemodic acid (2) * |  | 332.993 | M1, M2, M4 |
| emodin (3) |  | 269.053 | M1, M5 |
| emodic acid (4) |  | 299.032 | M1, M4, M5, M6, |
| citreorosein (5) |  | 285.049 | M2, M5, M6 |
| endocrocin (6) |  | 313.03 | M6 |
| 2-chloro-1,3,8-trihydroxy-6-(hydroxymethyl)anthracene-9,10-dione (7) * |  | 319.011 | M1, M2, M4, M5, M6 |
| 7-chloroemodin (8) * |  | 303.008 | M2, M6 |
| bisdethiobis(methylthio)acetylaranotine (9) * |  | 557.1028 | M2 |
| fellutanine C (10) * |  | 507.2750 | M1, M2 |
| canrenone (11) |  | 339.214 | M6, M2, M5 |
| tricin (12) |  | 329.083 | M2 |
| 5,6,2′-trimethoxyflavone (13) |  | 311.084 | M2 |
| hydroquinidine (14) |  | 325.196 | M2, M4, M5, |
| 4-methyl-5,6-diihydro-2H-pyran-2-one (15) * |  | 113.0601 | M2 |
| Position | Compound (9) | |
|---|---|---|
| δH (Multiplicity, J (Hz)) | δC | |
| 1 | - | 164.5 |
| 2 | - | 70.6 |
| 3 | 2.98–3.10 (m) | 40.6 |
| 4 | - | 109.6 |
| 5 | 6.57 (d, 8.2) | 137.8 |
| 7 | 6.29 (dd, 8.3 and 2.2) | 139.8 |
| 8 | 4.69 (dd, 8.2 and 1.3) | 105.9 |
| 9 | 5.80 (bd, 8.1) | 71.9 |
| 10 | 5.17 (bd, 7.6) | 60.4 |
| 12 | 2.26 (s) | 14.8 |
| 13 | - | 170.2 |
| 14 | 2.07 (s) | 21.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Carvalho, A.C.; Lima, C.S.; Torquato, H.F.V.; Domiciano, A.T.; Silva, S.d.C.; de Abreu, L.M.; Uemi, M.; Paredes-Gamero, E.J.; Vieira, P.C.; Veiga, T.A.M.; et al. Chemodiversity and Anti-Leukemia Effect of Metabolites from Penicillium setosum CMLD 18. Metabolites 2023, 13, 23. https://doi.org/10.3390/metabo13010023
de Carvalho AC, Lima CS, Torquato HFV, Domiciano AT, Silva SdC, de Abreu LM, Uemi M, Paredes-Gamero EJ, Vieira PC, Veiga TAM, et al. Chemodiversity and Anti-Leukemia Effect of Metabolites from Penicillium setosum CMLD 18. Metabolites. 2023; 13(1):23. https://doi.org/10.3390/metabo13010023
Chicago/Turabian Stylede Carvalho, Ana Calheiros, Cauê Santos Lima, Heron Fernandes Vieira Torquato, André Tarsis Domiciano, Sebastião da Cruz Silva, Lucas Magalhães de Abreu, Miriam Uemi, Edgar Julian Paredes-Gamero, Paulo Cezar Vieira, Thiago André Moura Veiga, and et al. 2023. "Chemodiversity and Anti-Leukemia Effect of Metabolites from Penicillium setosum CMLD 18" Metabolites 13, no. 1: 23. https://doi.org/10.3390/metabo13010023