
The Different Metabolic Responses of Resistant and Susceptible Wheats to Fusarium graminearum Inoculation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Fungus Material Culture and Plant Materials
2.3. Metabolite Extraction for Wheat
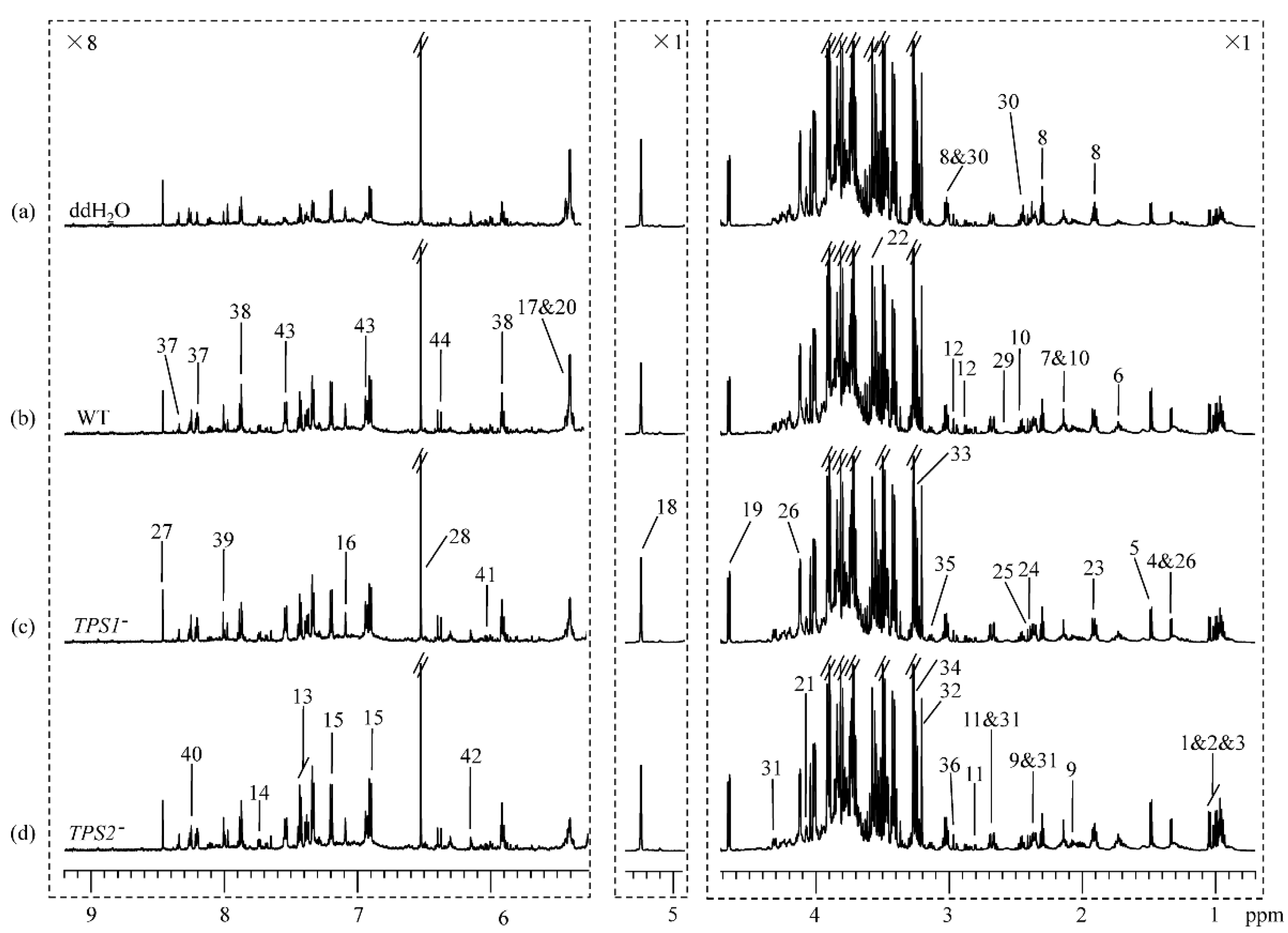
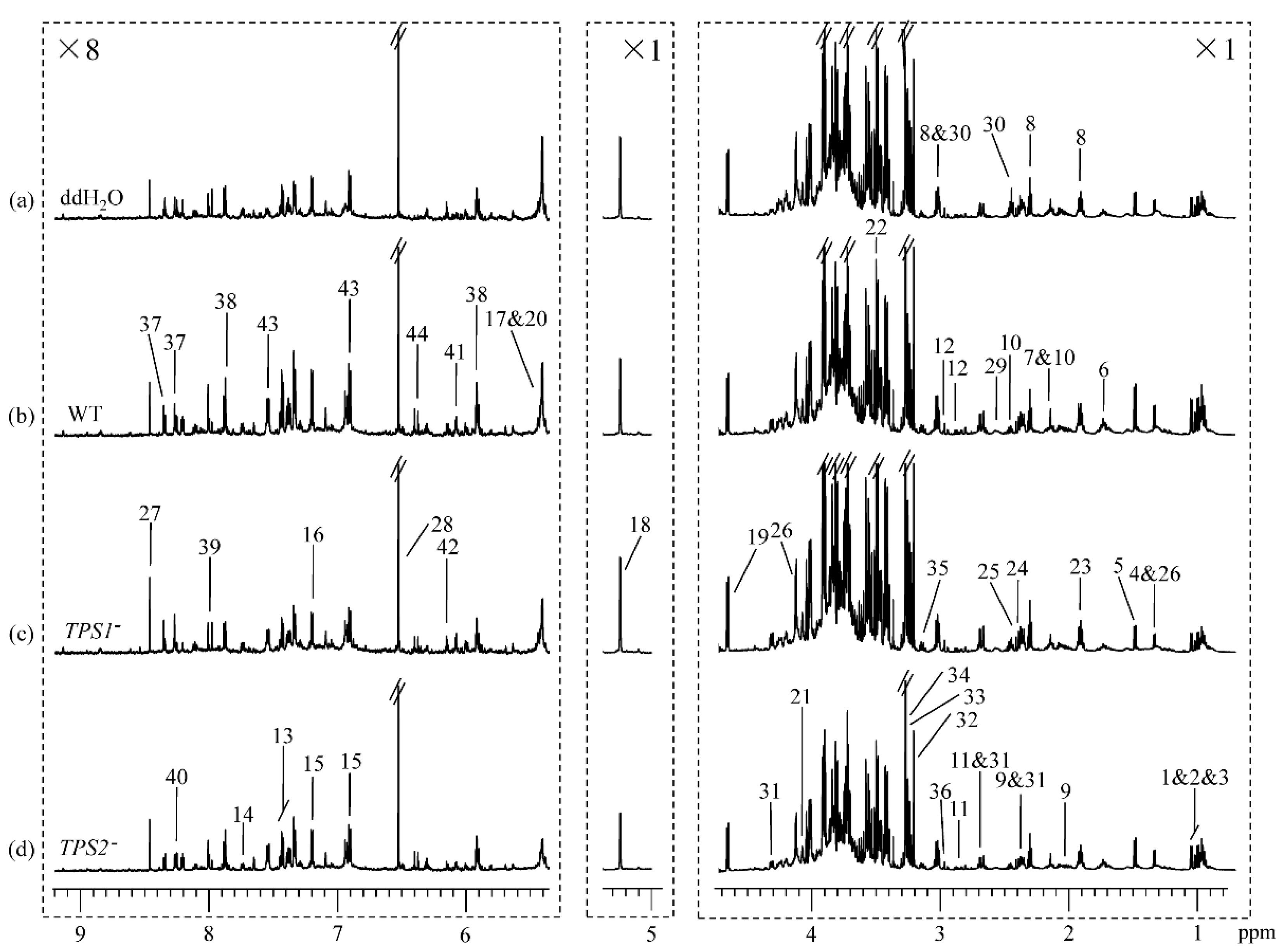
2.4. NMR Measurements
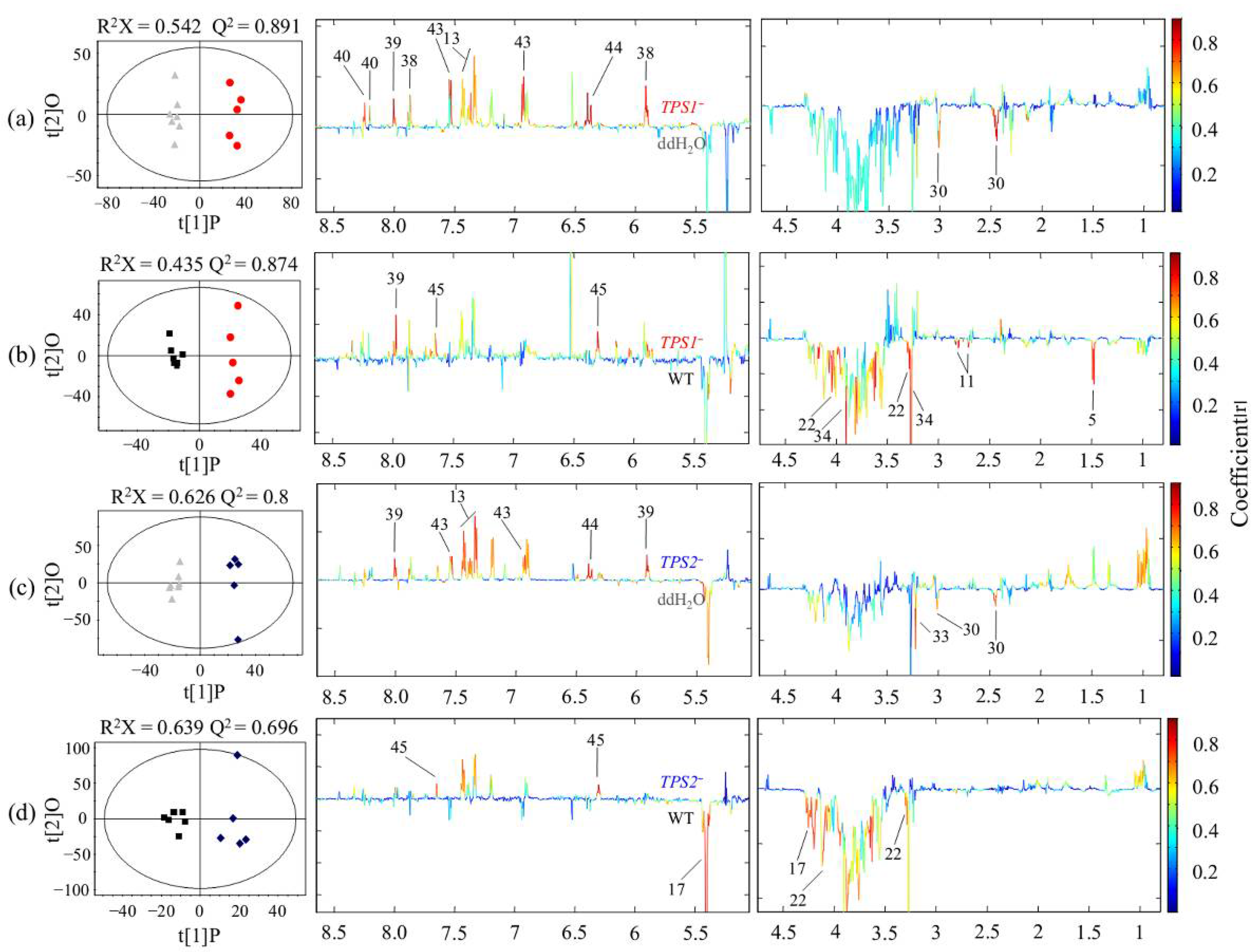
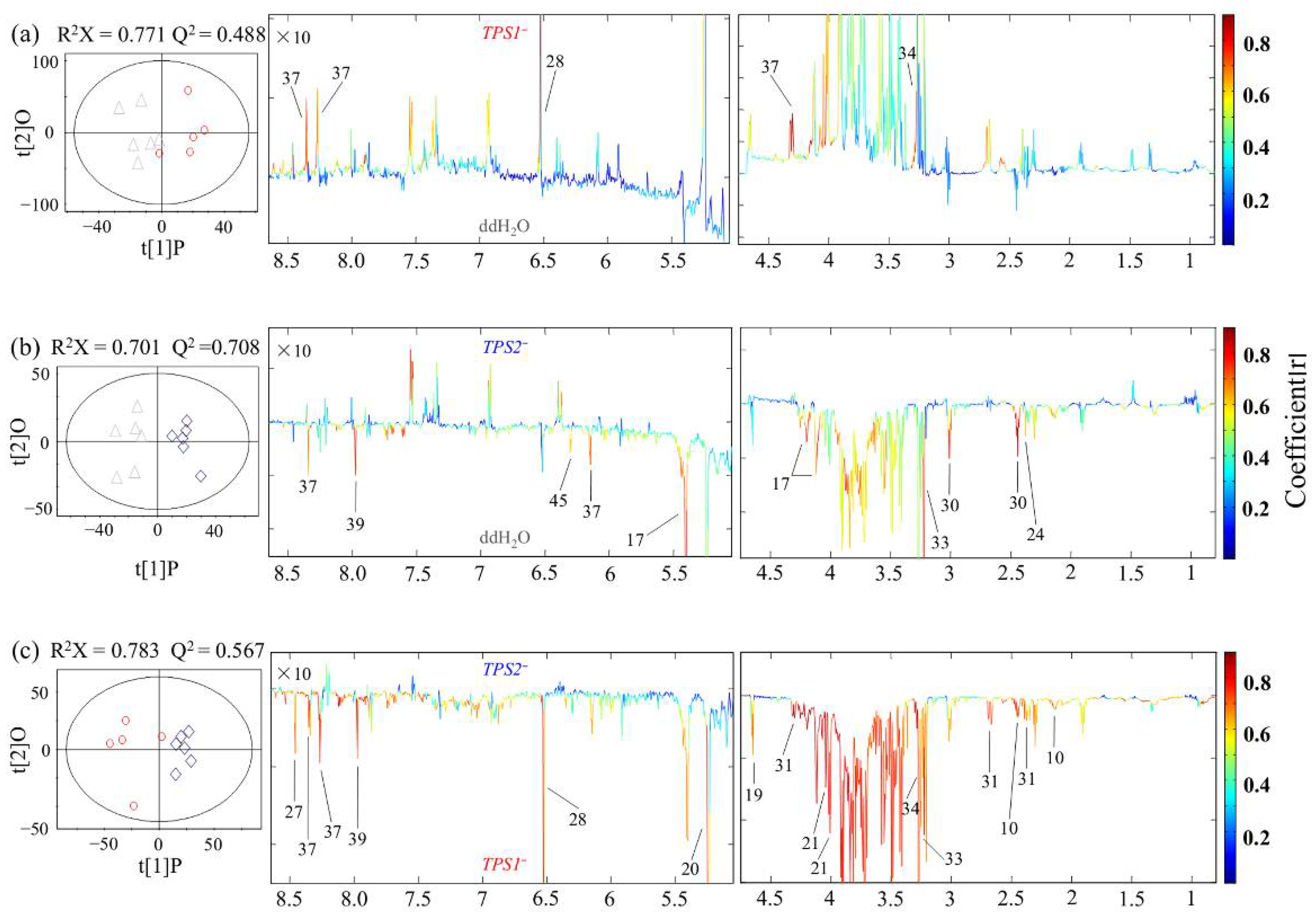
2.5. Spectral Processing and Multivariate Data Analysis
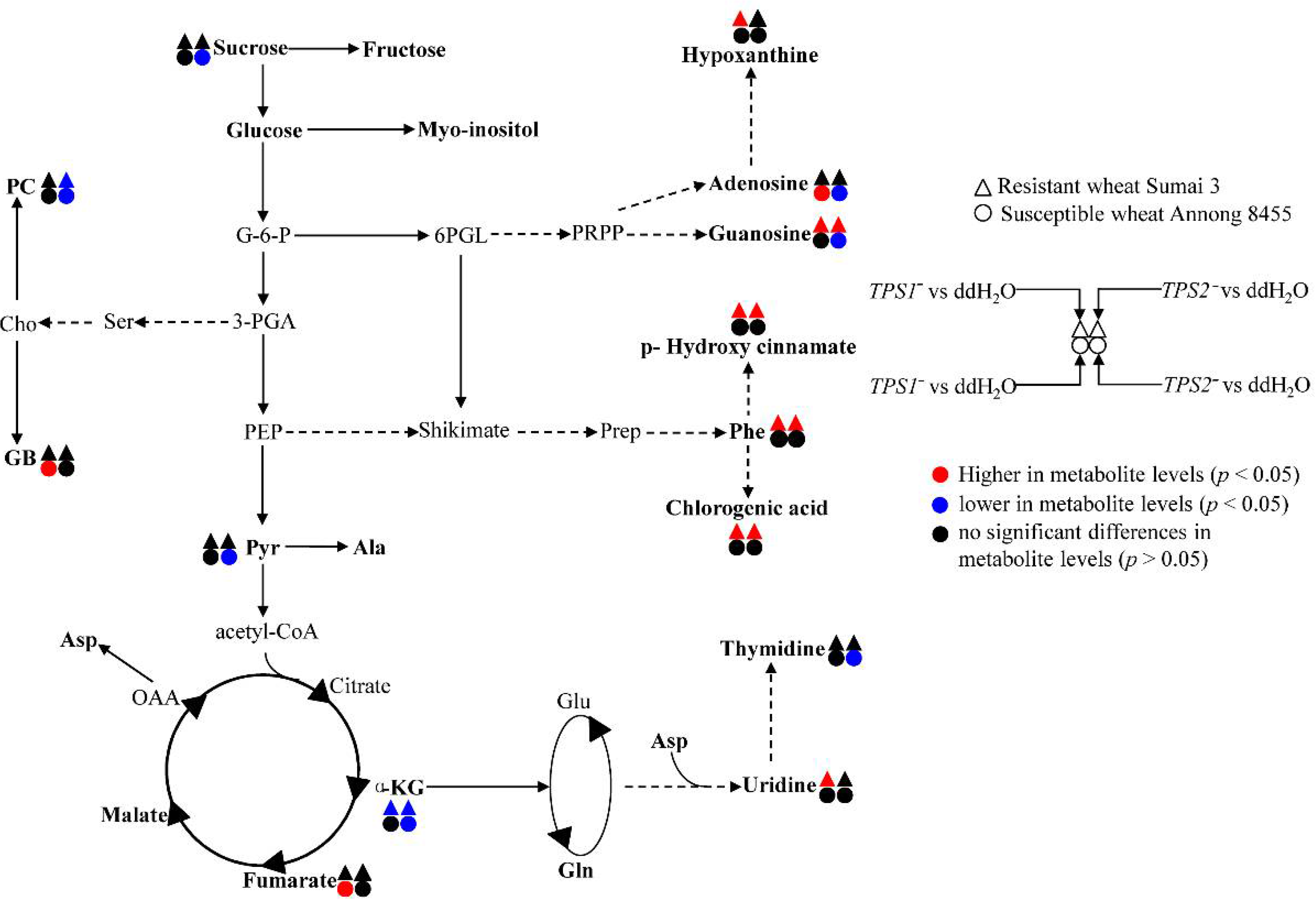
3. Results
3.1. Metabolic Profiles for FHB-Susceptible and -Resistant Wheats
3.2. Different Metabolic Responses of FHB-Resistant and -Susceptible Wheats to Three Fusarium Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blanco, A.; Bellomo, M.; Cenci, A.; De Giovanni, C.; D’ovidio, R.; Iacono, E.; Laddomada, B.; Pagnotta, M.; Porceddu, E.; Sciancalepore, A. A genetic linkage map of durum wheat. Theor. Appl. Genet. 1998, 97, 721–728. [Google Scholar] [CrossRef]
- Otto, C.; Kianian, S.; Elias, E.; Stack, R.; Joppa, L. Genetic dissection of a major Fusarium head blight QTL in tetraploid wheat. Plant Mol. Biol. 2002, 48, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Duffeck, M.R.; Bandara, A.Y.; Weerasooriya, D.K.; Collins, A.A.; Jensen, P.J.; Kuldau, G.A.; Del Ponte, E.M.; Esker, P.D. Fusarium head blight of small grains in pennsylvania: Unravelling species diversity, toxin types, growth, and triazole sensitivity. Phytopathology 2022, 112, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Podgorska-Kryszczuk, I.; Solarska, E.; Kordowska-Wiater, M. Reduction of the fusarium mycotoxins: Deoxynivalenol, nivalenol and zearalenone by selected non-conventional yeast strains in wheat grains and bread. Molecules 2022, 27, 1578. [Google Scholar] [CrossRef]
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef]
- Xu, X.-M.; Parry, D.; Nicholson, P.; Thomsett, M.; Simpson, D.; Edwards, S.; Cooke, B.; Doohan, F.; Brennan, J.; Moretti, A. Predominance and association of pathogenic fungi causing Fusarium ear blightin wheat in four European countries. Eur. J. Plant Pathol. 2005, 112, 143–154. [Google Scholar] [CrossRef]
- Rudd, J.; Horsley, R.; McKendry, A.; Elias, E. Host plant resistance genes for Fusarium head blight. Crop Sci. 2001, 41, 620–627. [Google Scholar] [CrossRef] [Green Version]
- Avonce, N.; MendozaVargas, A.; Morett, E.; Iturriaga, G. Insights on the evolution of trehalose biosynthesis. BMC Evol. Biol. 2006, 6, 109. [Google Scholar] [CrossRef] [Green Version]
- Gancedo, C.; Flores, C.L. The importance of a functional trehalose biosynthetic pathway for the life of yeasts and fungi. FEMS Yeast Res. 2004, 4, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Purvis, J.E.; Yomano, L.; Ingram, L. Enhanced trehalose production improves growth of Escherichia coli under osmotic stress. Appl. Environ. Microb. 2005, 71, 3761–3769. [Google Scholar] [CrossRef] [Green Version]
- Crowe, J.H.; Crowe, L.M.; Chapman, D. Preservation of membranes in anhydrobiotic organisms: The role of trehalose. Science 1984, 223, 701–703. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.A.; Lindquist, S. Multiple effects of trehalose on protein folding in vitro and in vivo. Mol. Cell 1998, 1, 639–648. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, Y.; Dai, B.; Wang, B.; Zhang, H.; Zhu, Z.; Xu, Y.; Cao, Y.; Jiang, Y.; Zhang, G. Trehalose is an important mediator of Cap1p oxidative stress response in Candida albicans. Biol. Pharm. Bull. 2008, 31, 421–425. [Google Scholar] [CrossRef] [Green Version]
- Zakharova, K.; Tesei, D.; Marzban, G.; Dijksterhuis, J.; Wyatt, T.; Sterflinger, K. Microcolonial fungi on rocks: A life in constant drought? Mycopathologia 2013, 175, 537–547. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.W.; Sun, M.; Gao, Y.M.; Luo, Y. Exogenous trehalose differently improves photosynthetic carbon assimilation capacities in maize and wheat under heat stress. J. Plant Interact. 2022, 17, 361–370. [Google Scholar] [CrossRef]
- Elbein, A.D.; Pan, Y.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17–27. [Google Scholar] [CrossRef]
- Bell, W.; Klaassen, P.; Ohnacker, M.; Boller, T.; Herweijer, M.; Schoppink, P.; Vanderzee, P.; Wiemken, A. Characterization of the 56-kDa subunit of yeast trehalose-6-phosphate synthase and cloning of its gene reveal its identity with the product of CIF1, a regulator of carbon catabolite inactivation. FEBS J. 1992, 209, 951–959. [Google Scholar] [CrossRef]
- Vuorio, O.E.; Kalkkinen, N.; Londesborough, J. Cloning of two related genes encoding the 56-kDa and 123-kDa subunits of trehalose synthase from the yeast Saccharomyces cerevisiae. FEBS J. 1993, 216, 849–861. [Google Scholar] [CrossRef]
- Al-Bader, N.; Vanier, G.; Liu, H.; Gravelat, F.N.; Urb, M.; Hoareau, C.M.-Q.; Campoli, P.; Chabot, J.; Filler, S.G.; Sheppard, D.C. Role of trehalose biosynthesis in Aspergillus fumigatus development, stress response, and virulence. Infect. Immun. 2010, 78, 3007–3018. [Google Scholar] [CrossRef] [Green Version]
- Puttikamonkul, S.; Willger, S.D.; Grahl, N.; Perfect, J.R.; Movahed, N.; Bothner, B.; Park, S.; Paderu, P.; Perlin, D.S.; Cramer, R.A., Jr. Trehalose 6-phosphate phosphatase is required for cell wall integrity and fungal virulence but not trehalose biosynthesis in the human fungal pathogen Aspergillus fumigatus. Mol. Microbiol. 2010, 77, 891–911. [Google Scholar] [CrossRef] [Green Version]
- Song, X.-S.; Li, H.-P.; Zhang, J.-B.; Song, B.; Huang, T.; Du, X.-M.; Gong, A.-D.; Liu, Y.-K.; Feng, Y.-N.; Agboola, R.S. Trehalose 6-phosphate phosphatase is required for development, virulence and mycotoxin biosynthesis apart from trehalose biosynthesis in Fusarium graminearum. Fungal Genet. Biol. 2014, 63, 24–41. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chen, F.; Zhang, J.; Liu, L.; Lei, H.; Li, H.; Wang, Y.; Liao, Y.-C.; Tang, H. Metabolic changes of Fusarium graminearum induced by TPS gene deletion. J. Proteome Res. 2019, 18, 3317–3327. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Liu, C.; Zhang, J.; Lei, H.; Li, H.-P.; Liao, Y.-C.; Tang, H. Combined metabonomic and quantitative RT-PCR analyses revealed metabolic reprogramming associated with Fusarium graminearum resistance in transgenic Arabidopsis thaliana. Front. Plant Sci. 2017, 8, 2177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.X.; Ding, F.; Hao, F.H.; Yu, M.; Lei, H.H.; Wu, X.Y.; Zhao, Z.X.; Guo, H.X.; Yin, J.; Wang, Y.L.; et al. Reprogramming of seed metabolism facilitates pre-harvest sprouting resistance of wheat. Sci. Rep. 2016, 6, 20593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Hao, F.; Hu, J.; Zhang, W.; Wan, L.; Zhu, L.; Tang, H.; He, G. Revealing different systems responses to brown planthopper infestation for pest susceptible and resistant rice plants with the combined metabonomic and gene-expression analysis. J. Proteome Res. 2010, 9, 6774–6785. [Google Scholar] [CrossRef]
- Marti, G.; Erb, M.; Boccard, J.; Glauser, G.; Doyen, G.R.; Villard, N.; Robert, C.A.M.; Turlings, T.C.; Rudaz, S.; Wolfender, J.L. Metabolomics reveals herbivore-induced metabolites of resistance and susceptibility in maize leaves and roots. Plant Cell Environ. 2013, 36, 621–639. [Google Scholar] [CrossRef]
- Freitas, D.D.; Carlos, E.F.; Gil, M.C.S.D.; Vieira, L.G.E.; Alcantara, G.B. NMR-based metabolomic analysis of huanglongbing-asymptomatic and -symptomatic citrus trees. J. Agric. Food Chem. 2015, 63, 7582–7588. [Google Scholar] [CrossRef]
- Cuperlovic-Culf, M.; Wang, L.; Forseille, L.; Boyle, K.; Merkley, N.; Burton, I.; Fobert, P.R. Metabolic biomarker panels of response to fusarium head blight infection in different wheat varieties. PLoS ONE 2016, 11, e0153642. [Google Scholar] [CrossRef]
- Baldaccicresp, F.; Chang, C.; Mickaël Maucourt Deborde, C.; Hopkins, J.; Lecomte, P. (Homo) glutathione deficiency impairs root-knot nematode development in Medicago truncatula. PLoS Pathog. 2012, 8, e1002471. [Google Scholar]
- Kumar, Y.; Zhang, L.; Panigrahi, P.; Dholakia, B.B.; Dewangan, V.; Chavan, S.G.; Kunjir, S.M.; Wu, X.; Li, N.; Rajmohanan, P.R. Fusarium oxysporum mediates systems metabolic reprogramming of chickpea roots as revealed by a combination of proteomics and metabolomics. Plant Biotechnol. J. 2016, 14, 1589–1603. [Google Scholar] [CrossRef] [Green Version]
- Marentes-Culma, R.; Cuellar-Cuestas, C.; Ardila, H.D.; Coy-Barrera, E. 5- n -alkylresorcinol-based metabolic response of rice to the interaction with burkholderia glumae: A chemical characterization of the temporal and spatial variations depending on environmental conditions. J. Plant Interact. 2022, 17, 127–139. [Google Scholar] [CrossRef]
- Liu, C.; Du, B.; Hao, F.; Lei, H.; Wan, Q.; He, G.; Wang, Y.; Tang, H. Dynamic metabolic responses of brown planthoppers towards susceptible and resistant rice plants. Plant Biotechnol. J. 2017, 15, 1346–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Li, T.; Gao, M.; Ye, M.; Lin, M.; Wu, D.; Guo, J.; Guan, W.; Wang, J.; Yang, K.; et al. Transcriptome and metabolome profiling reveal the resistance mechanisms of rice against brown planthopper. Int. J. Mol. Sci. 2022, 23, 4083. [Google Scholar] [CrossRef] [PubMed]
- Browne, R.A.; Brindle, K.M. 1H NMR-based metabolite profiling as a potential selection tool for breeding passive resistance against Fusarium head blight (FHB) in wheat. Mol. Plant Pathol. 2007, 8, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Xu, L.; He, X.; Zhang, L.; Shaban, M.; Zhang, X.; Zhu, L. Suppression of tryptophan synthase activates cotton immunity by triggering cell death via promoting SA synthesis. Plant J. 2019, 98, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.B.; Li, H.P.; Dang, F.J.; Qu, B.; Xu, Y.B.; Zhao, C.S.; Liao, Y.C. Determination of the trichothecene mycotoxin chemotypes and associated geographical distribution and phylogenetic species of the Fusarium graminearum clade from China. Mycol. Res. 2007, 111, 967–975. [Google Scholar] [CrossRef]
- Wu, A.B.; Li, H.P.; Zhao, C.S.; Liao, Y.C. Comparative pathogenicity of Fusarium graminearum isolates from China revealed by wheat coleoptile and floret inoculations. Mycopathologia 2005, 160, 75–83. [Google Scholar] [CrossRef]
- Duvick, J.; Rood, T.; Rao, A.G.; Marshak, D.R. Purification and characterization of a novel antimicrobial peptide from maize (Zea mays L.) kernels. J. Biol. Chem. 1992, 267, 18814–18820. [Google Scholar] [CrossRef]
- Li, X.; Zhang, J.; Song, B.; Li, H.; Xu, H.; Qu, B.; Dang, F.; Liao, Y. Resistance to Fusarium head blight and seedling blight in wheat is associated with activation of a cytochrome P450 gene. Phytopathology 2010, 100, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.Y.; Li, N.; Li, H.D.; Tang, H.R. An optimized method for NMR-based plant seed metabolomic analysis with maximized polar metabolite extraction efficiency, signal-to-noise ratio, and chemical shift consistency. Analyst 2014, 139, 1769–1778. [Google Scholar] [CrossRef]
- Trygg, J.; Wold, S. Orthogonal projections to latent structures (O-PLS). J. Chemometr. 2002, 16, 119–128. [Google Scholar] [CrossRef]
- Eriksson, L.; Trygg, J.; Wold, S. CV-ANOVA for significance testing of PLS and OPLS® models. J. Chemometr. 2008, 22, 594–600. [Google Scholar] [CrossRef]
- Cloarec, O.; Dumas, M.E.; Trygg, J.; Craig, A.; Barton, R.H.; Lindon, J.C.; Nicholson, J.K.; Holmes, E. Evaluation of the orthogonal projection on latent structure model limitations caused by chemical shift variability and improved visualization of biomarker changes in 1H NMR spectroscopic metabonomic studies. Anal. Chem. 2005, 77, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.W.M. Metabolite profiling by one-and two-dimensional NMR analysis of complex mixtures. Prog. Nucl. Mag. Res. Spectrosc. 1996, 28, 161–219. [Google Scholar] [CrossRef]
- Fan, T.W.M.; Lane, A.N. Structure-based profiling of metabolites and isotopomers by NMR. Prog. Nucl. Mag. Res. Spectrosc. 2008, 52, 69–117. [Google Scholar] [CrossRef]
- Facchini, P.J.; Hagel, J.; Zulak, K.G. Hydroxycinnamic acid amide metabolism: Physiology and biochemistry. Can. J. Bot.-Rev. Can. Bot. 2002, 80, 577–589. [Google Scholar] [CrossRef]
- González-Lamothe, R.; Mitchell, G.; Gattuso, M.; Diarra, M.S.; Malouin, F.; Bouarab, K. Plant antimicrobial agents and their effects on plant and human pathogens. Int. J. Mol. Sci. 2009, 10, 3400–3419. [Google Scholar] [CrossRef]
- Atanasova-Penichon, V.; Pons, S.; Pinson-Gadais, L.; Picot, A.; Marchegay, G.; Bonnin-Verdal, M.-N.; Ducos, C.; Barreau, C.; Roucolle, J.; Sehabiague, P. Chlorogenic acid and maize ear rot resistance: A dynamic study investigating Fusarium graminearum development, deoxynivalenol production, and phenolic acid accumulation. Mol. Plant-Microbe Interact. 2012, 25, 1605–1616. [Google Scholar] [CrossRef] [Green Version]
- Shetty, R.; Fretté, X.; Jensen, B.; Shetty, N.P.; Jensen, J.D.; Jorgensen, H.J.L.; Newman, M.-A.; Christensen, L.P. Silicon-induced changes in antifungal phenolic acids, flavonoids, and key phenylpropanoid pathway genes during the interaction between Miniature roses and the biotrophic pathogen Podosphaera pannosa. Plant Physiol. 2011, 157, 2194–2205. [Google Scholar] [CrossRef] [Green Version]
- Sabino, A.R.; Tavares, S.S.; Riffel, A.; Li, J.V.; Oliveira, D.J.; Feres, C.I.; Henrique, L.; Oliveira, J.S.; Correia, G.D.; Sabino, A.R.; et al. 1H NMR metabolomic approach reveals chlorogenic acid as a response of sugarcane induced by exposure to diatraea saccharalis. Ind. Crops Prod. 2019, 140, 111651. [Google Scholar] [CrossRef]
- Gunnaiah, R.; Kushalappa, A.C. Metabolomics deciphers the host resistance mechanisms in wheat cultivar Sumai-3, against trichothecene producing and non-producing isolates of Fusarium graminearum. Plant Physiol. Biochem. 2014, 83, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Arisan-Atac, I.; Wolschek, M.F.; Kubicek, C.P. Trehalose-6-phosphate synthase A affects citrate accumulation by Aspergillus niger under conditions of high glycolytic flux. FEMS Microbiol. Lett. 1996, 140, 77–83. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coefficient (r) a | |||||||
|---|---|---|---|---|---|---|---|
| Metabolites (No) | Sumai 3 (Resistant) | Annong 8455 (Susceptible) | |||||
| Sugars | TPS1− vs. H2O | TPS1− vs. WT | TPS2− vs. H2O | TPS2− vs. WT | TPS1− vs. H2O | TPS2− vs. H2O | TPS2− vs. TPS1− |
| sucrose (17) c | −0.849 b | −0.882 | |||||
| α-glucose (18) | −0.826 | ||||||
| β-glucose (19) | −0.816 | ||||||
| fructose (21) | −0.857 | ||||||
| myo-inositol (22) | −0.788 | −0.821 | |||||
| Amino acids | |||||||
| Ala (5) | −0.781 | ||||||
| Gln (10) | −0.762 | ||||||
| Asp (11) | −0.857 | ||||||
| Phe (13) | 0.835 | 0.756 | |||||
| Organic acids | |||||||
| pyruvate (24) | −0.841 | ||||||
| formate (27) | −0.823 | ||||||
| fumarate (28) | 0.794 | −0.904 | |||||
| a-KG (30) | −0.836 | −0.782 | −0.769 | ||||
| malate (31) | −0.963 | ||||||
| Choline metabolites | |||||||
| phosphocholine (33) | −0.859 | −0.877 | −0.871 | ||||
| glycine betaine (34) | −0.837 | 0.814 | −0.892 | ||||
| Nucleotide metabolites | |||||||
| adenosine (37) | 0.824 | −0.805 | −0.917 | ||||
| uridine (38) | 0.825 | ||||||
| guanosine (39) | 0.929 | 0.758 | 0.799 | −0.826 | −0.799 | ||
| hypoxanthine (40) | 0.856 | ||||||
| thymidine (45) | 0.821 | 0.834 | −0.843 | ||||
| Secondary metabolites | |||||||
| p-hydroxy cinnamaic acid (43) | 0.854 | 0.753 | |||||
| chlorogenic acid (44) | 0.922 | 0.826 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Chen, F.; Liu, L.; Fan, X.; Liu, H.; Zeng, D.; Zhang, X. The Different Metabolic Responses of Resistant and Susceptible Wheats to Fusarium graminearum Inoculation. Metabolites 2022, 12, 727. https://doi.org/10.3390/metabo12080727
Liu C, Chen F, Liu L, Fan X, Liu H, Zeng D, Zhang X. The Different Metabolic Responses of Resistant and Susceptible Wheats to Fusarium graminearum Inoculation. Metabolites. 2022; 12(8):727. https://doi.org/10.3390/metabo12080727
Chicago/Turabian StyleLiu, Caixiang, Fangfang Chen, Laixing Liu, Xinyu Fan, Huili Liu, Danyun Zeng, and Xu Zhang. 2022. "The Different Metabolic Responses of Resistant and Susceptible Wheats to Fusarium graminearum Inoculation" Metabolites 12, no. 8: 727. https://doi.org/10.3390/metabo12080727