
Improvement of Glycemic Control by a Functional Food Mixture Containing Maltodextrin, White Kidney Bean Extract, Mulberry Leaf Extract, and Niacin-Bound Chromium Complex in Obese Diabetic db/db Mice


Abstract
:1. Introduction
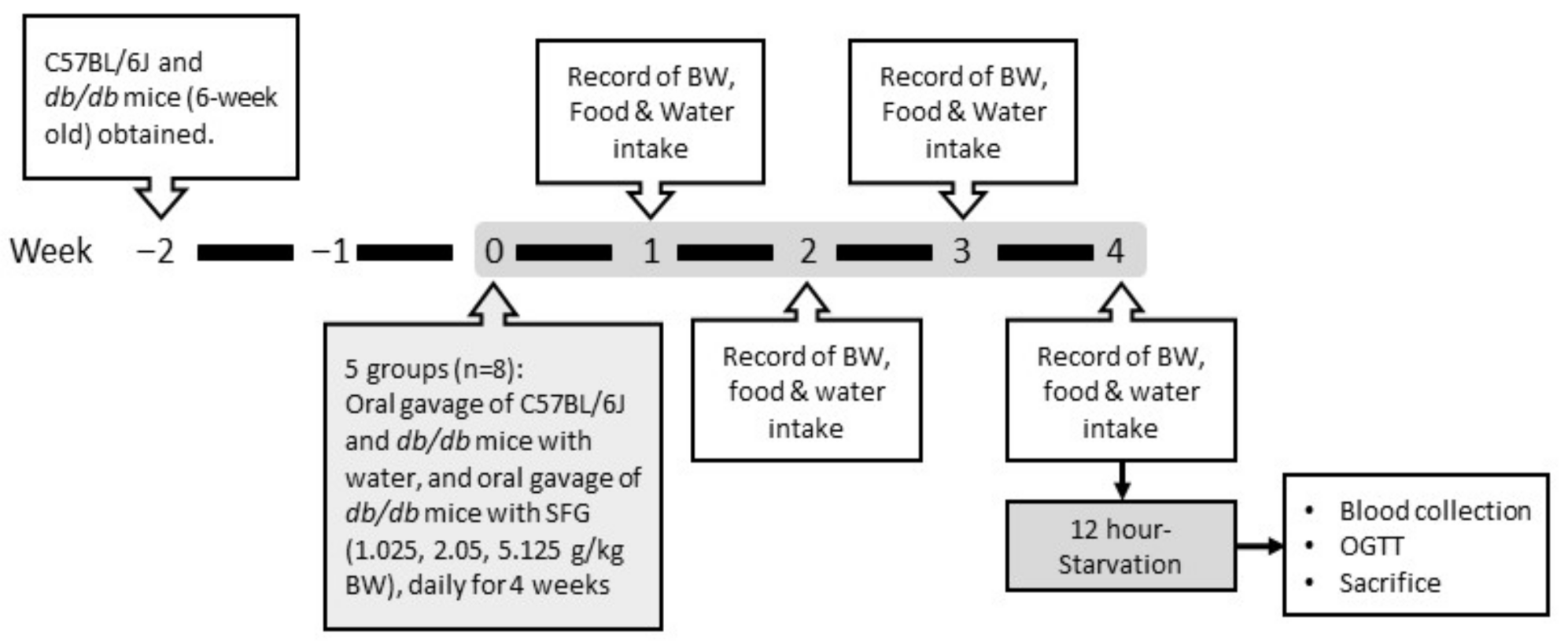
2. Materials and Methods
2.1. Animals and Diets
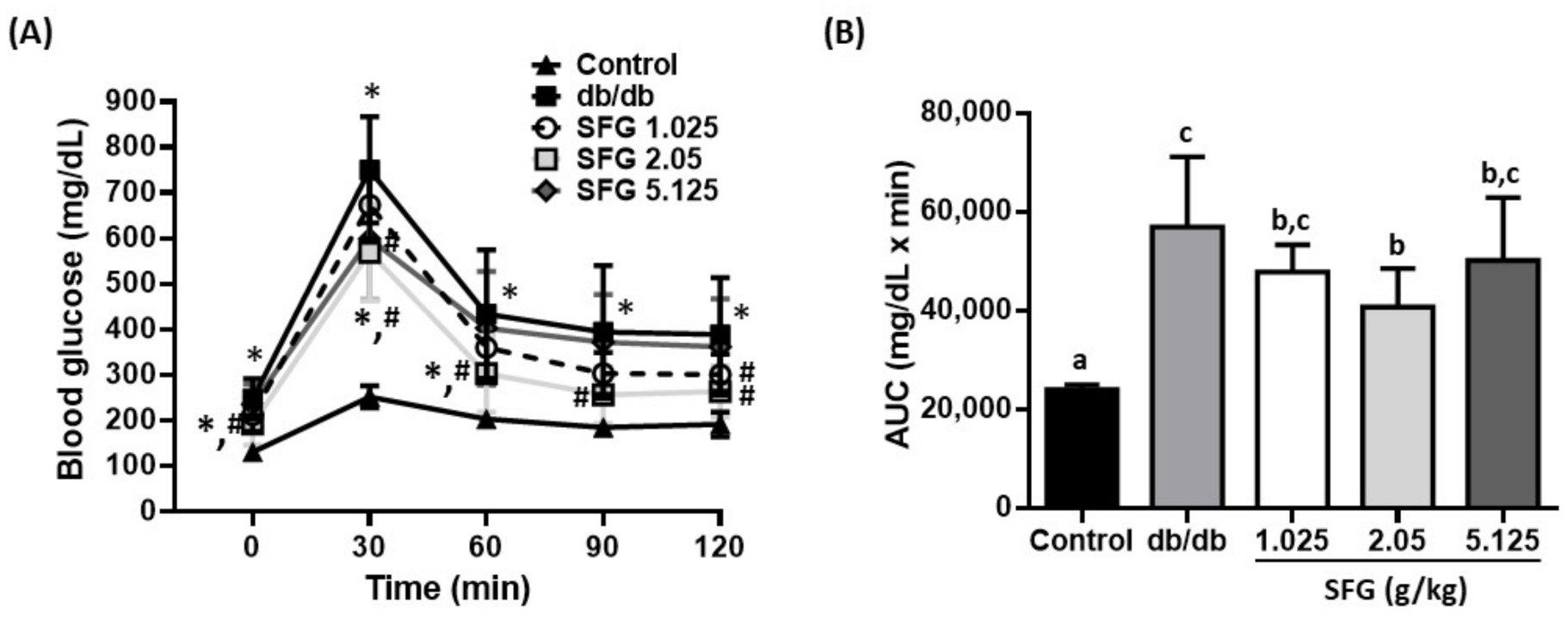
2.2. Measurement of Fasting Plasma Glucose and Oral Glucose Tolerance Test (OGTT)
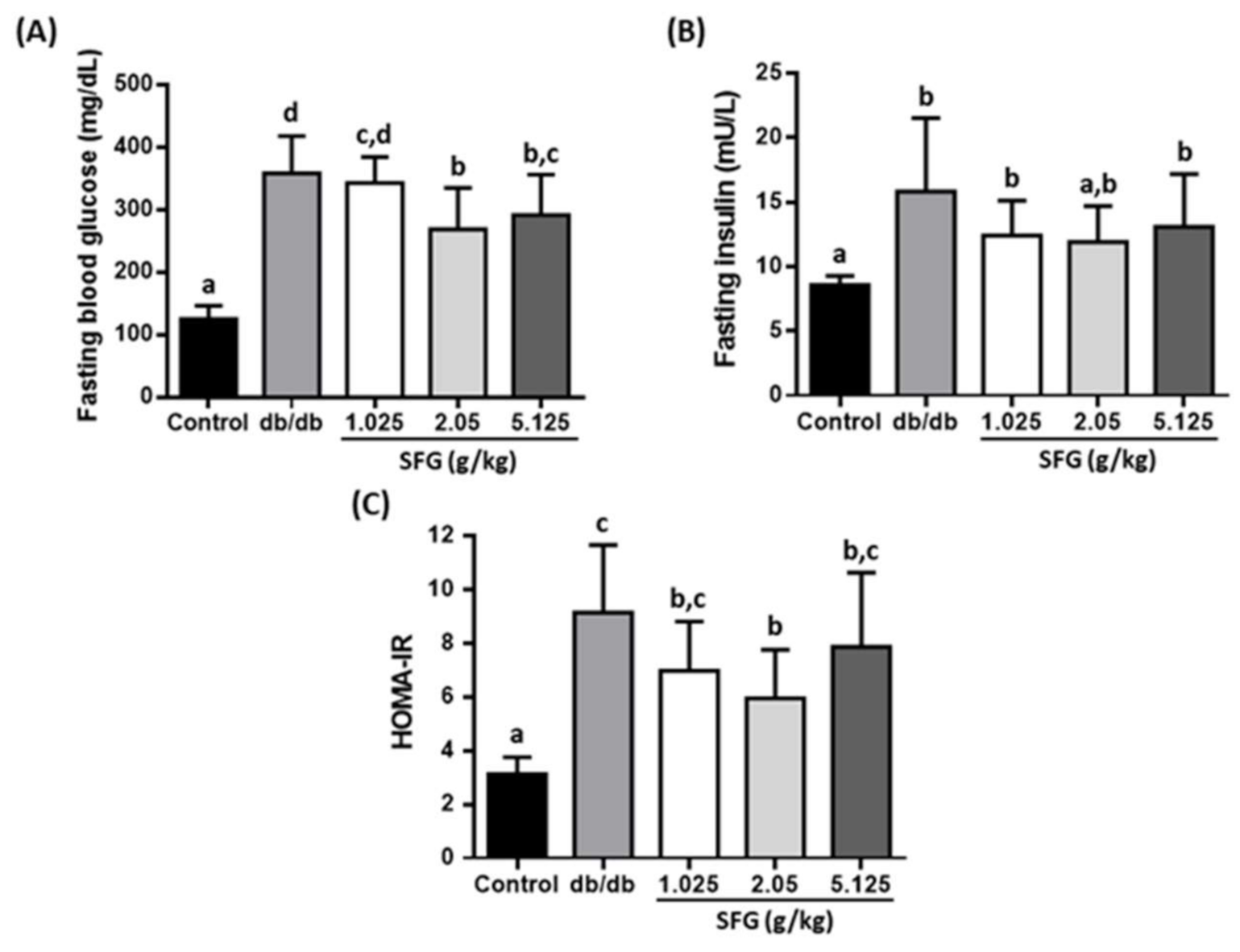
2.3. Measurement of Insulin and Homeostatic Model Assessment for Insulin Resistance (HOMA-IR)
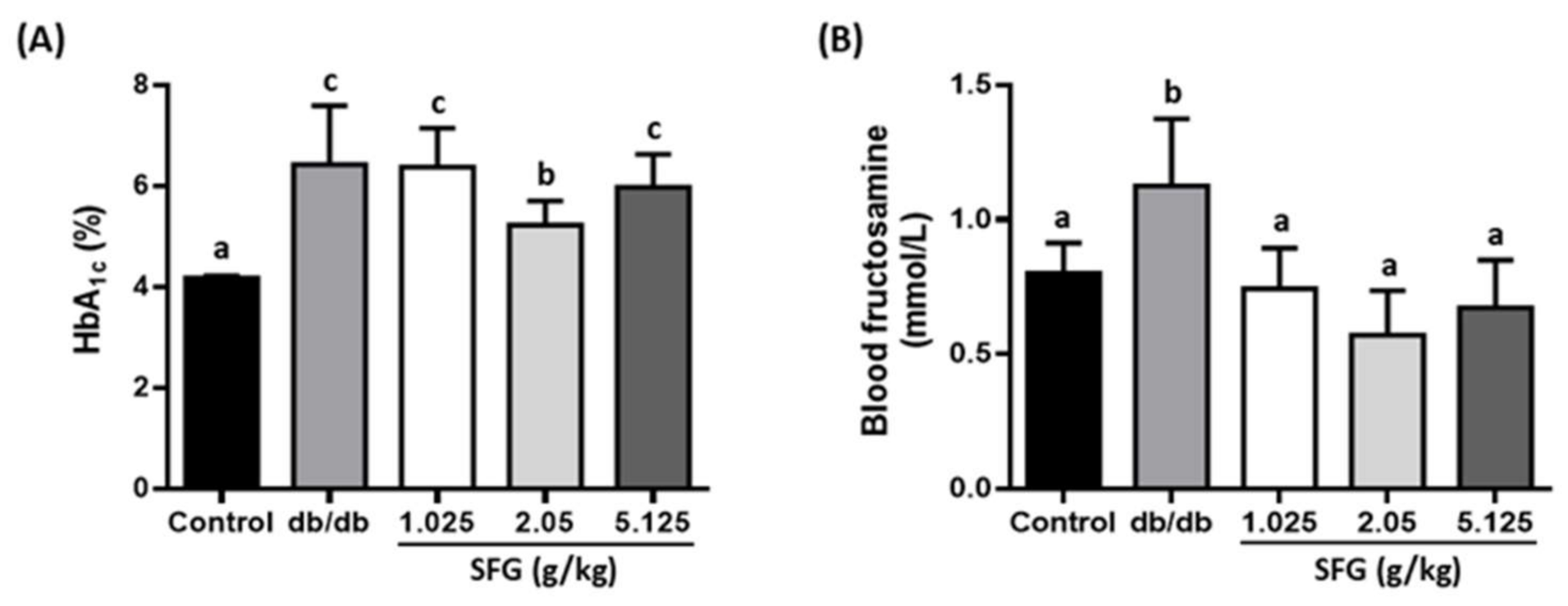
2.4. Measurement of Blood Biochemical Parameters
2.5. Statistical Analysis
3. Results
3.1. Effects of SFG on Weight Gain, Food Intake, and Water Consumption in db/db Mice
3.2. SFG Improved Glycemic Parameters in db/db Mice
3.3. SFG Improved Glucose Uptake by Enhancing Insulin Sensitivity in db/db Mice
3.4. SFG Improved Plasma Triglyceride (TG) Levels in db/db Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bhupathiraju, S.N.; Hu, F.B. Epidemiology of Obesity and Diabetes and Their Cardiovascular Complications. Circ. Res. 2016, 118, 1723–1735. [Google Scholar] [CrossRef] [PubMed]
- International Diabetes Federation. IDF Diabetes Atlas, 2017, 8th ed.; Suvi, K., Joao da Rocha Fernandes, B.M., Eds.; International Diabetes Federation: Brussels, Belgium, 2017; ISBN 978-2-930229-87-4. Available online: http://www.diabetesatlas.org/ (accessed on 3 February 2020).
- Centers for Disease Control and Prevention. National Diabetes Statistics Report website. 2022. Available online: https://www.cdc.gov/diabetes/data/statistics-report/index.html (accessed on 9 May 2022).
- Sattar, N.; Gill, J.M. Type 2 diabetes as a disease of ectopic fat? BMC Med. 2014, 12, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef]
- Ley, S.H.; Hamdy, O.; Mohan, V.; Hu, F.B. Prevention and management of type 2 diabetes: Dietary components and nutritional strategies. Lancet 2014, 383, 1999–2007. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Te Morenga, L. Carbohydrate quality and human health: A series of systematic reviews and meta-analyses. Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef] [Green Version]
- Institute of Medicine. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids (Macronutrients); The National Academies Press: Washington, DC, USA, 2002; Available online: https://www8.nationalacademies.org/onpinews/newsitem.aspx?RecordID=s10490 (accessed on 13 May 2022).
- Fayet-Moore, F.; George, A.; Cassettari, T.; Yulin, L.; Tuck, K.; Pezzullo, L. Healthcare Expenditure and Productivity Cost Savings from Reductions in Cardiovascular Disease and Type 2 Diabetes Associated with Increased Intake of Cereal Fibre among Australian Adults: A Cost of Illness Analysis. Nutrients 2018, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Stephen, A.M.; Champ, M.M.-J.; Cloran, S.J.; Fleith, M.; Van Lieshout, L.; Mejborn, H.; Burley, V.J. Dietary fibre in Europe: Current state of knowledge on definitions, sources, recommendations, intakes and relationships to health. Nutr. Res. Rev. 2017, 30, 149–190. [Google Scholar] [CrossRef]
- Murphy, M.M.; Douglass, J.S.; Birkett, A. Resistant Starch Intakes in the United States. J. Am. Diet. Assoc. 2008, 108, 67–78. [Google Scholar] [CrossRef]
- Birt, D.F.; Boylston, T.; Hendrich, S.; Jane, J.-L.; Hollis, J.; Li, L.; McClelland, J.; Moore, S.; Phillips, G.J.; Rowling, M.; et al. Resistant Starch: Promise for Improving Human Health. Adv. Nutr. Int. Rev. J. 2013, 4, 587–601. [Google Scholar] [CrossRef] [Green Version]
- Slavin, J.L. Dietary fiber and body weight. Nutrition 2005, 21, 411–418. [Google Scholar] [CrossRef]
- Astina, J.; Sapwarobol, S. Resistant Maltodextrin and Metabolic Syndrome: A Review. J. Am. Coll. Nutr. 2018, 38, 380–385. [Google Scholar] [CrossRef]
- Hashizume, C.; Kishimoto, Y.; Kanahori, S.; Yamamoto, T.; Okuma, K.; Yamamoto, K. Improvement Effect of Resistant Maltodextrin in Humans with Metabolic Syndrome by Continuous Administration. J. Nutr. Sci. Vitaminol. 2012, 58, 423–430. [Google Scholar] [CrossRef] [Green Version]
- Barrett, M.L.; Udani, J.K. A proprietary alpha-amylase inhibitor from white bean (Phaseolus vulgaris): A review of clinical studies on weight loss and glycemic control. Nutr. J. 2011, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Nolan, R.; Shannon, O.M.; Robinson, N.; Joel, A.; Houghton, D.; Malcomson, F.C. It’s No Has Bean: A Review of the Effects of White Kidney Bean Extract on Body Composition and Metabolic Health. Nutrients 2020, 12, 1398. [Google Scholar] [CrossRef]
- Chan, E.W.C.; Lye, P.-Y.; Wong, S.-K. Phytochemistry, pharmacology, and clinical trials of Morus alba. Chin. J. Nat. Med. 2016, 14, 17–30. [Google Scholar] [CrossRef]
- Jozefczuk, J.; Malikowska, K.; Glapa, A.; Stawinska-Witoszynska, B.; Nowak, J.K.; Bajerska, J.; Lisowska, A.; Walkowiak, J. Mulberry leaf extract decreases digestion and absorption of starch in healthy subjects—A randomized, placebo-controlled, crossover study. Adv. Med. Sci. 2017, 62, 302–306. [Google Scholar] [CrossRef]
- Riche, D.M.; Riche, K.D.; East, H.E.; Barrett, E.K.; May, W.L. Impact of mulberry leaf extract on type 2 diabetes (Mul-DM): A randomized, placebo-controlled pilot study. Complement. Ther. Med. 2017, 32, 105–108. [Google Scholar] [CrossRef]
- Kim, G.-N.; Kwon, Y.-I.; Jang, H.-D. Mulberry Leaf Extract Reduces Postprandial Hyperglycemia with Few Side Effects by Inhibiting α-Glucosidase in Normal Rats. J. Med. Food 2011, 14, 712–717. [Google Scholar] [CrossRef]
- Shara, M.; Yasmin, T.; Kincaid, A.; Limpach, A.L.; Bartz, J.; Brenneman, K.A.; Chatterjee, A.; Bagchi, M.; Stohs, S.J.; Bagchi, D. Safety and toxicological evaluation of a novel niacin-bound chromium (III) complex. J. Inorg. Biochem. 2005, 99, 2161–2183. [Google Scholar] [CrossRef]
- Perricone, N.V.; Bagchi, D.; Echard, B.; Preuss, H.G. Blood pressure lowering effects of niacin-bound chromium (III) (NBC) in sucrose-fed rats: Renin–angiotensin system. J. Inorg. Biochem. 2008, 102, 1541–1548. [Google Scholar] [CrossRef]
- Lau, F.C.; Bagchi, M.; Sen, C.K.; Bagchi, D. Nutrigenomic basis of beneficial effects of chromium (III) on obesity and diabetes. Mol. Cell. Biochem. 2008, 317, 1–10. [Google Scholar] [CrossRef]
- Röder, P.V.; Wu, B.; Liu, Y.; Han, W. Pancreatic regulation of glucose homeostasis. Exp. Mol. Med. 2016, 48, e219. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-T.; Chiu, C.-Y.; Huang, C.-F.; Peng, F.-C.; Liu, S.-H. Genotoxicity and 28-day oral toxicity studies of a functional food mixture containing maltodextrin, white kidney bean extract, mulberry leaf extract, and niacin-bound chromium complex. Regul. Toxicol. Pharmacol. 2018, 92, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Suriano, F.; Vieira-Silva, S.; Falony, G.; Roumain, M.; Paquot, A.; Pelicaen, R.; Régnier, M.; Delzenne, N.M.; Raes, J.; Muccioli, G.G.; et al. Novel insights into the genetically obese (ob/ob) and diabetic (db/db) mice: Two sides of the same coin. Microbiome 2021, 9, 147. [Google Scholar] [CrossRef]
- Wang, B.; Chandrasekera, P.C.; Pippin, J.J. Leptin- and Leptin Receptor-Deficient Rodent Models: Relevance for Human Type 2 Diabetes. Curr. Diabetes Rev. 2014, 10, 131–145. [Google Scholar] [CrossRef] [Green Version]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrinello, C.M.; Selvin, E. Beyond HbA1c and Glucose: The Role of Nontraditional Glycemic Markers in Diabetes Diagnosis, Prognosis, and Management. Curr. Diabetes Rep. 2014, 14, 548. [Google Scholar] [CrossRef] [PubMed]
- Gounden, V.; Ngu, M.; Anastasopoulou, C.; Jialal, I. Fructosamine; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Antuna-Puente, B.; Disse, E.; Rabasa-Lhoret, R.; Laville, M.; Capeau, J.; Bastard, J.P. How can we measure insulin sensitivity/resistance? Diabetes Metab. 2011, 37, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Bergman, M.; Abdul-Ghani, M.; DeFronzo, R.A.; Manco, M.; Sesti, G.; Fiorentino, T.V.; Ceriello, A.; Rhee, M.; Phillips, L.S.; Chung, S.; et al. Review of methods for detecting glycemic disorders. Diabetes Res. Clin. Pract. 2020, 165, 108233. [Google Scholar] [CrossRef]
- Portincasa, P.; Bonfrate, L.; Vacca, M.; De Angelis, M.; Farella, I.; Lanza, E.; Khalil, M.; Wang, D.Q.-H.; Sperandio, M.; Di Ciaula, A. Gut Microbiota and Short Chain Fatty Acids: Implications in Glucose Homeostasis. Int. J. Mol. Sci. 2022, 23, 1105. [Google Scholar] [CrossRef]
- Meenu, M.; Xu, B. A critical review on anti-diabetic and anti-obesity effects of dietary resistant starch. Crit. Rev. Food Sci. Nutr. 2018, 59, 3019–3031. [Google Scholar] [CrossRef]
- Colantonio, A.G.; Werner, S.L.; Brown, M. The Effects of Prebiotics and Substances with Prebiotic Properties on Metabolic and Inflammatory Biomarkers in Individuals with Type 2 Diabetes Mellitus: A Systematic Review. J. Acad. Nutr. Diet. 2019, 120, 587–607. [Google Scholar] [CrossRef] [Green Version]
- Ye, Z.; Arumugam, V.; Haugabrooks, E.; Williamson, P.; Hendrich, S. Soluble dietary fiber (Fibersol-2) decreased hunger and increased satiety hormones in humans when ingested with a meal. Nutr. Res. 2015, 35, 393–400. [Google Scholar] [CrossRef]
- Astina, J.; Sapwarobol, S. Attenuation of glycaemic and insulin responses following tapioca resistant maltodextrin consumption in healthy subjects: A randomised cross-over controlled trial. J. Nutr. Sci. 2020, 9, e29. [Google Scholar] [CrossRef]
- Goodman, B.E. Insights into digestion and absorption of major nutrients in humans. Adv. Physiol. Educ. 2010, 34, 44–53. [Google Scholar] [CrossRef]
- Phimarn, W.; Wichaiyo, K.; Silpsavikul, K.; Sungthong, B.; Saramunee, K. A meta-analysis of efficacy of Morus alba Linn. to improve blood glucose and lipid profile. Eur. J. Nutr. 2016, 56, 1509–1521. [Google Scholar] [CrossRef]
- Horne, G.; Wilson, F.X.; Tinsley, J.; Williams, D.H.; Storer, R. Iminosugars past, present and future: Medicines for tomorrow. Drug Discov. Today 2011, 16, 107–118. [Google Scholar] [CrossRef]
- Bae, U.-J.; Jung, E.-S.; Jung, S.-J.; Chae, S.-W.; Park, B.-H. Mulberry leaf extract displays antidiabetic activity in db/db mice via Akt and AMP-activated protein kinase phosphorylation. Food Nutr. Res. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Naowaboot, J.; Pannangpetch, P.; Kukongviriyapan, V.; Prawan, A.; Kukongviriyapan, U.; Itharat, A. Mulberry Leaf Extract Stimulates Glucose Uptake and GLUT4 Translocation in Rat Adipocytes. Am. J. Chin. Med. 2012, 40, 163–175. [Google Scholar] [CrossRef]
- Naowaboot, J.; Pannangpetch, P.; Kukongviriyapan, V.; Kukongviriyapan, U.; Nakmareong, S.; Itharat, A. Mulberry leaf extract restores arterial pressure in streptozotocin-induced chronic diabetic rats. Nutr. Res. 2009, 29, 602–608. [Google Scholar] [CrossRef]
- Gao, X.-H.; Zhang, S.-D.; Wang, L.-T.; Yu, L.; Zhao, X.-L.; Ni, H.-Y.; Wang, Y.-Q.; Wang, J.-D.; Shan, C.-H.; Fu, Y.-J. Anti-Inflammatory Effects of Neochlorogenic Acid Extract from Mulberry Leaf (Morus alba L.) Against LPS-Stimulated Inflammatory Response through Mediating the AMPK/Nrf2 Signaling Pathway in A549 Cells. Molecules 2020, 25, 1385. [Google Scholar] [CrossRef] [Green Version]
- Anderson, A.R. Chromium as an Essential Nutrient for Humans. Regul. Toxicol. Pharmacol. 1997, 26, S35–S41. [Google Scholar] [CrossRef] [Green Version]
- FDA. Food labeling: Reference Daily Intakes, Final Rule; Department of Health and Human Services, Food and Drug Administration: Washington, DC, USA, 1995; 2022, pp. 67164–67175.
- Institute of Medicine. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; The National Academies Press: Washington, DC, USA, 2001; pp. 197–223. ISBN 978-0-309-07290-8. [Google Scholar]
- Hua, Y.; Clark, S.; Ren, J.; Sreejayan, N. Molecular mechanisms of chromium in alleviating insulin resistance. J. Nutr. Biochem. 2012, 23, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Batsis, J.A.; Apolzan, J.W.; Bagley, P.J.; Blunt, H.B.; Divan, V.; Gill, S.; Golden, A.; Gundumraj, S.; Heymsfield, S.B.; Kahan, S.; et al. A Systematic Review of Dietary Supplements and Alternative Therapies for Weight Loss. Obesity 2021, 29, 1102–1113. [Google Scholar] [CrossRef]
- Willis, H.J.; Thomas, W.; Eldridge, A.L.; Harkness, L.; Green, H.; Slavin, J.L. Increasing doses of fiber do not influence short-term satiety or food intake and are inconsistently linked to gut hormone levels. Food Nutr. Res. 2010, 54, 5135. [Google Scholar] [CrossRef]
- Nirmala Prasadi, P.V.; Joye, I.J. Dietary Fibre from Whole Grains and Their Benefits on Metabolic Health. Nutrients 2020, 12, 3045. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SFG (g/kg) | |||||
|---|---|---|---|---|---|
| Control | db/db | 1.025 | 2.05 | 5.125 | |
| Body Weight (g) | |||||
| Week 1 | 22.98 ± 0.73 a | 35.25 ± 2.87 b | 36.48 ± 1.78 b | 36.53 ± 2.05 b | 36.80 ± 1.65 b |
| Week 2 | 24.18 ± 1.09 a | 37.13 ± 2.79 b | 37.60 ± 1.85 b | 38.45 ± 2.31 b | 38.38 ± 2.32 b |
| Week 3 | 24.35 ± 1.03 a | 38.95 ± 2.17 b | 38.50 ± 1.96 b | 39.48 ± 1.08 b | 39.10 ± 2.64 b |
| Week 4 | 24.28 ± 1.06 a | 39.95 ± 1.33 b,c | 38.35 ± 2.33 b | 41.83 ± 2.17 c | 39.58 ± 2.99 b |
| Feed Intake (g/day) | |||||
| Week 1 | 5.71 ± 0.97 a | 11.28 ± 2.18 b | 10.08 ± 1.10 b | 12.42 ± 1.96 b | 10.68 ± 1.48 b |
| Week 2 | 6.84 ± 1.65 a | 9.00 ± 1.04 a,b | 7.82 ± 0.94 a,b | 9.67 ± 0.96 b | 8.73 ± 2.02 a,b |
| Week 3 | 6.89 ± 1.74 a | 9.51 ± 0.25 b | 7.51 ± 0.58 a,b | 9.17 ± 1.05 a,b | 8.15 ± 2.60 a,b |
| Week 4 | 7.44 ± 0.63 a | 9.08 ± 0.56 b | 6.82 ± 0.50 a,b | 7.80 ± 0.53 a,b | 8.20 ± 2.17 a,b |
| Water Consumption (g/day) | |||||
| Week 1 | 7.05 ± 0.61 a | 17.43 ± 10.11 b | 16.62 ± 1.80 b | 15.07 ± 1.96 b | 15.78 ± 1.07 b |
| Week 2 | 6.29 ± 0.32 a | 17.05 ± 16.36 b | 17.25 ± 13.69 b | 13.32 ± 0.90 b | 16.27 ± 2.06 b |
| Week 3 | 6.11 ± 0.14 a | 11.85 ± 6.64 b | 12.42 ± 5.28 b | 12.58 ± 1.33 b | 12.03 ± 6.57 b |
| Week 4 | 5.74 ± 1.79 a | 14.44 ± 10.64 b | 14.21 ± 1.14 b | 12.10 ± 0.77 b | 14.95 ± 10.03 b |
| SFG (g/kg) | |||||
|---|---|---|---|---|---|
| Control | db/db | 1.025 | 2.05 | 5.125 | |
| TC (mg/dL) | 89.3 ± 14.3 a | 129.0 ± 25.2 b | 108.8 ± 8.8 c | 133.0 ± 22.3 b,c | 111.4 ± 28.1 a,b,c |
| TG (mg/dL) | 119.1 ± 6.4 a | 192.3 ± 21.1 c | 158.5 ± 31.0 b | 168.8 ± 23.7 b | 160.0 ± 21.2 b |
| AST (U/L) | 168.0 ± 59.4 a | 300.9 ± 184.1 a | 266.9 ± 138.9 a | 282.0 ± 218.2 a | 202.8 ± 131.7 a |
| ALT (U/L) | 39.1 ± 17.6 a | 84.0 ± 27.5 b | 93.5 ± 60.8 b | 75.5 ± 36.3 a,b | 69.9 ± 30.2 a,b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tzeng, H.-P.; Chiu, C.-Y.; Liu, S.-H.; Chiang, M.-T. Improvement of Glycemic Control by a Functional Food Mixture Containing Maltodextrin, White Kidney Bean Extract, Mulberry Leaf Extract, and Niacin-Bound Chromium Complex in Obese Diabetic db/db Mice. Metabolites 2022, 12, 693. https://doi.org/10.3390/metabo12080693
Tzeng H-P, Chiu C-Y, Liu S-H, Chiang M-T. Improvement of Glycemic Control by a Functional Food Mixture Containing Maltodextrin, White Kidney Bean Extract, Mulberry Leaf Extract, and Niacin-Bound Chromium Complex in Obese Diabetic db/db Mice. Metabolites. 2022; 12(8):693. https://doi.org/10.3390/metabo12080693
Chicago/Turabian StyleTzeng, Huei-Ping, Chen-Yuan Chiu, Shing-Hwa Liu, and Meng-Tsan Chiang. 2022. "Improvement of Glycemic Control by a Functional Food Mixture Containing Maltodextrin, White Kidney Bean Extract, Mulberry Leaf Extract, and Niacin-Bound Chromium Complex in Obese Diabetic db/db Mice" Metabolites 12, no. 8: 693. https://doi.org/10.3390/metabo12080693