
Intrapersonal Stability of Plasma Metabolomic Profiles over 10 Years among Women

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Study Population Characteristics
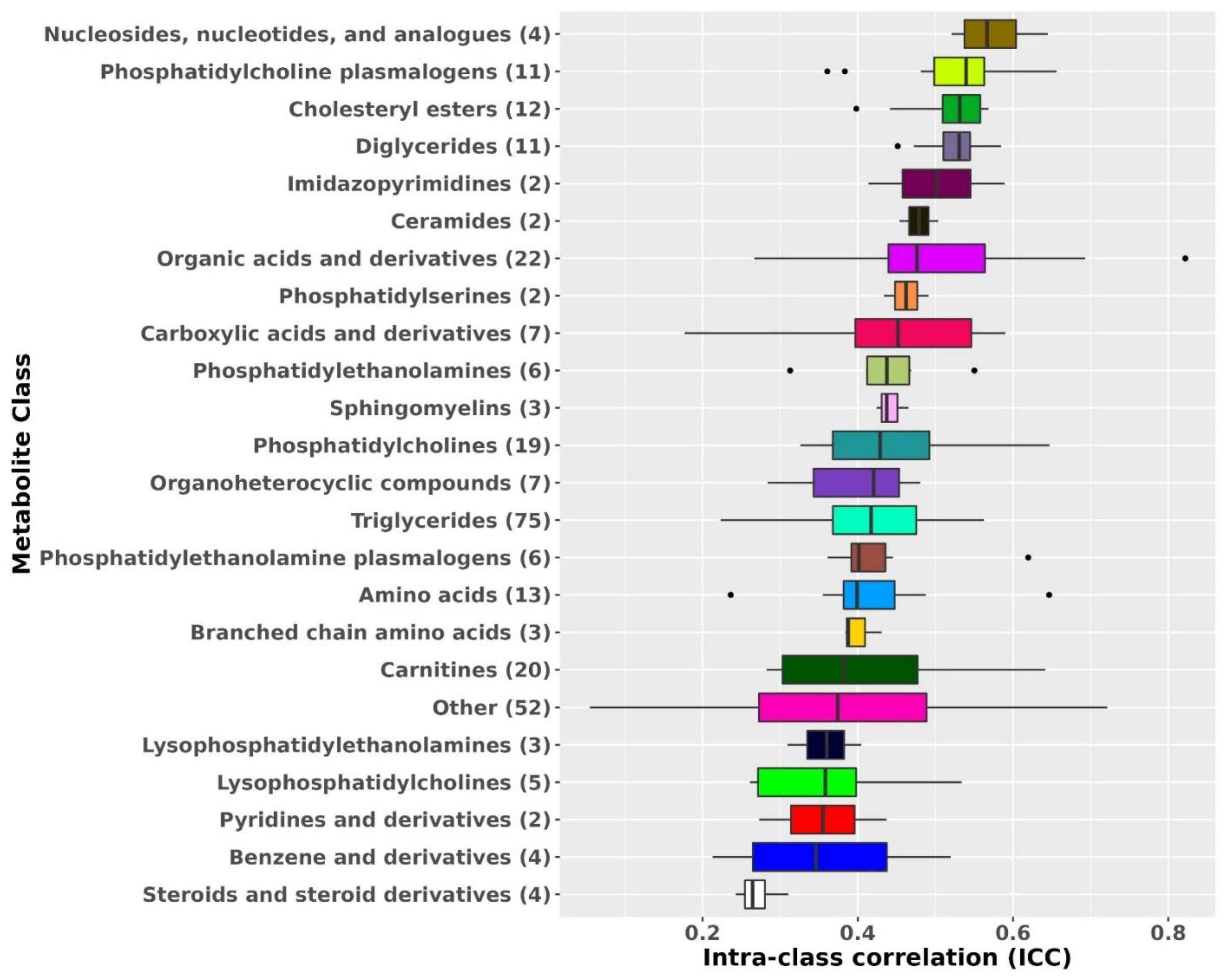
2.2. Metabolite Profile Stability over 10 Years in the Primary Dataset
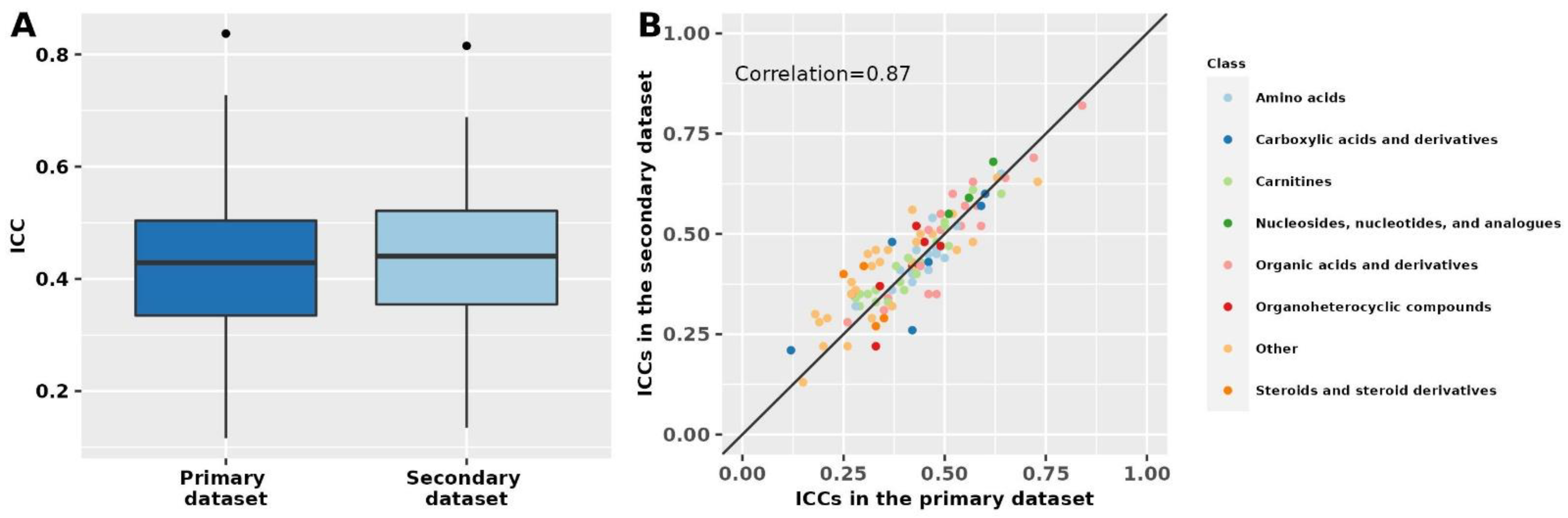
2.3. Metabolite Profile Stability over 10 Years in the Secondary Dataset
3. Discussion
4. Materials and Methods
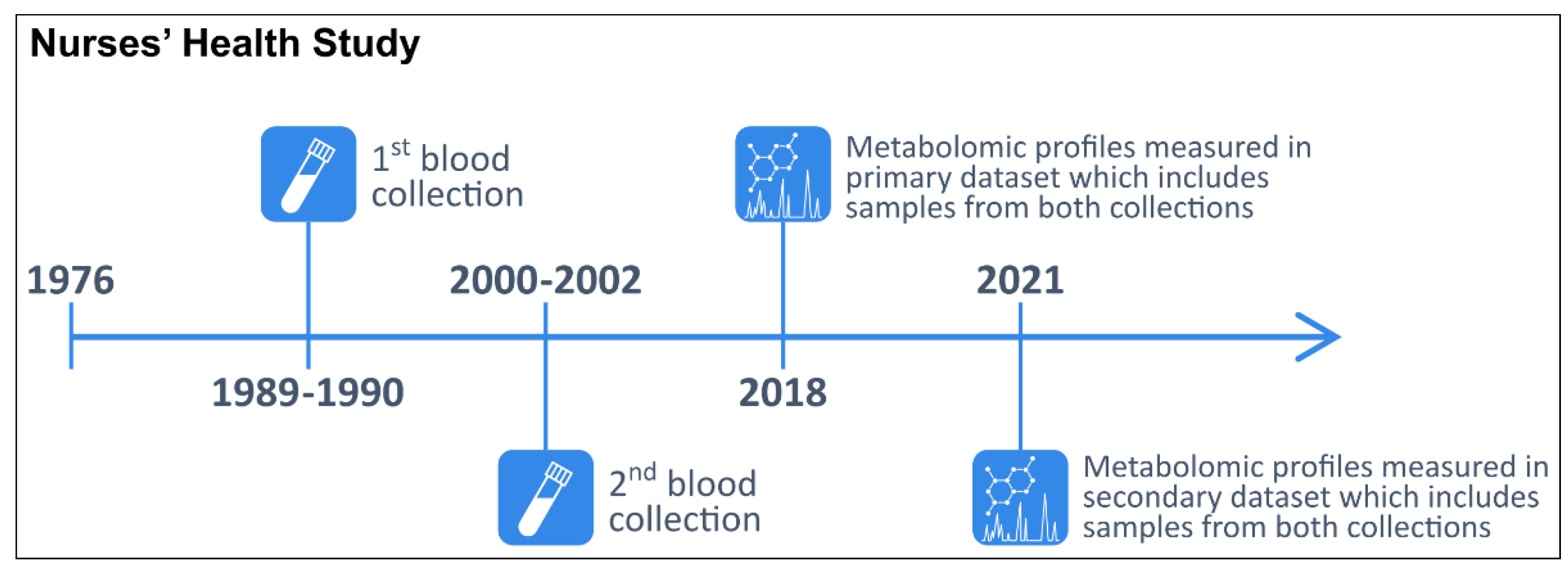
4.1. Study Population
4.2. Blood Collection Methods
4.3. Metabolite Profiling
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eliassen, A.H.; Liao, X.; Rosner, B.; Tamimi, R.M.; Tworoger, S.S.; Hankinson, S.E. Plasma carotenoids and risk of breast cancer over 20 y of follow-up. Am. J. Clin. Nutr. 2015, 101, 1197–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tworoger, S.S.; Eliassen, A.H.; Zhang, X.; Qian, J.; Sluss, P.M.; Rosner, B.A.; Hankinson, S.E. A 20-Year Prospective Study of Plasma Prolactin as a Risk Marker of Breast Cancer Development. Cancer Res. 2013, 73, 4810–4819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krumsiek, J.; Bartel, J.; Theis, F.J. Computational approaches for systems metabolomics. Curr. Opin. Biotechnol. 2016, 39, 198–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeleznik, O.A.; Clish, C.B.; Kraft, P.; Avila-Pacheco, J.; Eliassen, A.H.; Tworoger, S.S. Circulating Lysophosphatidylcholines, Phosphatidylcholines, Ceramides, and Sphingomyelins and Ovarian Cancer Risk: A 23-Year Prospective Study. JNCI J. Natl. Cancer Inst. 2019, 112, 628–636. [Google Scholar] [CrossRef]
- Zeleznik, O.A.; Eliassen, A.H.; Kraft, P.; Poole, E.M.; Rosner, B.A.; Jeanfavre, S.; Deik, A.A.; Bullock, K.; Hitchcock, D.S.; Avila-Pacheco, J.; et al. A Prospective Analysis of Circulating Plasma Metabolites Associated with Ovarian Cancer Risk. Cancer Res. 2020, 80, 1357–1367. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, D.L.S.; Maple, H.J.; Goodwin, M.; Brand, J.S.; Yip, V.; Min, J.L.; Groom, A.; Lawlor, D.A.; Ring, S. The Effect of Pre-Analytical Conditions on Blood Metabolomics in Epidemiological Studies. Metabolites 2019, 9, 64. [Google Scholar] [CrossRef] [Green Version]
- Kamlage, B.; Maldonado, S.G.; Bethan, B.; Peter, E.; Schmitz, O.; Liebenberg, V.; Schatz, P. Quality Markers Addressing Preanalytical Variations of Blood and Plasma Processing Identified by Broad and Targeted Metabolite Profiling. Clin. Chem. 2014, 60, 399–412. [Google Scholar] [CrossRef]
- Nishiumi, S.; Suzuki, M.; Kobayashi, T.; Yoshida, M. Differences in metabolite profiles caused by pre-analytical blood processing procedures. J. Biosci. Bioeng. 2018, 125, 613–618. [Google Scholar] [CrossRef] [Green Version]
- Jobard, E.; Trédan, O.; Postoly, D.; André, F.; Martin, A.-L.; Elena-Herrmann, B.; Boyault, S. A Systematic Evaluation of Blood Serum and Plasma Pre-Analytics for Metabolomics Cohort Studies. Int. J. Mol. Sci. 2016, 17, 2035. [Google Scholar] [CrossRef] [Green Version]
- Breier, M.; Wahl, S.; Prehn, C.; Fugmann, M.; Ferrari, U.; Weise, M.; Banning, F.; Seissler, J.; Grallert, H.; Adamski, J.; et al. Targeted Metabolomics Identifies Reliable and Stable Metabolites in Human Serum and Plasma Samples. PLoS ONE 2014, 9, e89728. [Google Scholar] [CrossRef]
- Anton, G.; Wilson, R.; Yu, Z.-H.; Prehn, C.; Zukunft, S.; Adamski, J.; Heier, M.; Meisinger, C.; Römisch-Margl, W.; Wang-Sattler, R.; et al. Pre-Analytical Sample Quality: Metabolite Ratios as an Intrinsic Marker for Prolonged Room Temperature Exposure of Serum Samples. PLoS ONE 2015, 10, e0121495. [Google Scholar] [CrossRef] [PubMed]
- Hernandes, V.V.; Barbas, C.; Dudzik, D. A review of blood sample handling and pre-processing for metabolomics studies. Electrophoresis 2017, 38, 2232–2241. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Lehmann, R.; Xu, G. Effects of pre-analytical processes on blood samples used in metabolomics studies. Anal. Bioanal. Chem. 2015, 407, 4879–4892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner-Golbs, A.; Neuber, S.; Kamlage, B.; Christiansen, N.; Bethan, B.; Rennefahrt, U.; Schatz, P.; Lind, L. Effects of Long-Term Storage at −80 °C on the Human Plasma Metabolome. Metabolites 2019, 9, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haid, M.; Muschet, C.; Wahl, S.; Römisch-Margl, W.; Prehn, C.; Möller, G.; Adamski, J. Long-Term Stability of Human Plasma Metabolites during Storage at −80 °C. J. Proteome Res. 2017, 17, 203–211. [Google Scholar] [CrossRef]
- Yin, P.; Peter, A.; Franken, H.; Zhao, X.; Neukamm, S.S.; Rosenbaum, L.; Lucio, M.; Zell, A.; Häring, H.-U.; Xu, G.; et al. Preanalytical Aspects and Sample Quality Assessment in Metabolomics Studies of Human Blood. Clin. Chem. 2013, 59, 833–845. [Google Scholar] [CrossRef]
- Kamlage, B.; Neuber, S.; Bethan, B.; González Maldonado, S.; Wagner-Golbs, A.; Peter, E.; Schmitz, O.; Schatz, P. Impact of Prolonged Blood Incubation and Extended Serum Storage at Room Temperature on the Human Serum Metabolome. Metabolites 2018, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Chen, Y.; Xi, C.; Zhang, R.; Song, Y.; Zhan, Q.; Bi, X.; Abliz, Z. Liquid Chromatography–Tandem Mass Spectrometry-Based Plasma Metabonomics Delineate the Effect of Metabolites’ Stability on Reliability of Potential Biomarkers. Anal. Chem. 2013, 85, 2606–2610. [Google Scholar] [CrossRef]
- Abuja, P.M.; Ehrhart, F.; Schoen, U.; Schmidt, T.; Stracke, F.; Dallmann, G.; Friedrich, T.; Zimmermann, H.; Zatloukal, K. Alterations in Human Liver Metabolome during Prolonged Cryostorage. J. Proteome Res. 2015, 14, 2758–2768. [Google Scholar] [CrossRef]
- Teahan, O.; Gamble, S.; Holmes, E.; Waxman, J.; Nicholson, J.K.; Bevan, A.C.; Keun, H.C. Impact of Analytical Bias in Metabonomic Studies of Human Blood Serum and Plasma. Anal. Chem. 2006, 78, 4307–4318. [Google Scholar] [CrossRef]
- Townsend, M.K.; Bao, Y.; Poole, E.M.; Bertrand, K.; Kraft, P.; Wolpin, B.M.; Clish, C.; Tworoger, S.S. Impact of Pre-analytic Blood Sample Collection Factors on Metabolomics. Cancer Epidemiol. Biomark. Prev. 2016, 25, 823–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Zhai, G.; Singmann, P.; He, Y.; Xu, T.; Prehn, C.; Römisch-Margl, W.; Lattka, E.; Gieger, C.; Soranzo, N.; et al. Human serum metabolic profiles are age dependent. Aging Cell 2012, 11, 960–967. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, F.; Luchinat, C.; Tenori, L.; Saccenti, E. Age-and Sex-Dependent Changes of Free Circulating Blood Metabolite and Lipid Abundances, Correlations, and Ratios. J. Gerontol. Ser. A 2021, glab335. [Google Scholar] [CrossRef]
- Darst, B.F.; Koscik, R.L.; Hogan, K.J.; Johnson, S.C.; Engelman, C.D. Longitudinal plasma metabolomics of aging and sex. Aging 2019, 11, 1262–1282. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, G.; Jing, R.; Hu, T.; Likhodii, S.; Sun, G.; Randell, E.; Jia, G.; Yu, T.; Zhang, W. Metabolomics analysis of human plasma metabolites reveals the age- and sex-specific associations. J. Liq. Chromatogr. Relat. Technol. 2019, 43, 185–194. [Google Scholar] [CrossRef]
- Saito, K.; Maekawa, K.; Kinchen, J.M.; Tanaka, R.; Kumagai, Y.; Saito, Y. Gender- and Age-Associated Differences in Serum Metabolite Profiles among Japanese Populations. Biol. Pharm. Bull. 2016, 39, 1179–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trabado, S.; Al-Salameh, A.; Croixmarie, V.; Masson, P.; Corruble, E.; Fève, B.; Colle, R.; Ripoll, L.; Walther, B.; Boursier-Neyret, C.; et al. The human plasma-metabolome: Reference values in 800 French healthy volunteers; impact of cholesterol, gender and age. PLoS ONE 2017, 12, e0173615. [Google Scholar] [CrossRef] [Green Version]
- Chaleckis, R.; Murakami, I.; Takada, J.; Kondoh, H.; Yanagida, M. Individual variability in human blood metabolites identifies age-related differences. Proc. Natl. Acad. Sci. USA 2016, 113, 4252–4259. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Sadanala, K.C.; Kim, A.E.-K. A Metabolomic Approach to Understanding the Metabolic Link between Obesity and Diabetes. Mol. Cells 2015, 38, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.; Sun, H.; Wang, X. Emerging role and recent applications of metabolomics biomarkers in obesity disease research. Rsc. Adv. 2017, 7, 14966–14973. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.; O′Grada, C.; Ryan, M.; Roche, H.M.; Gibney, M.J.; Gibney, E.R.; Brennan, L. The relationship between BMI and metabolomic profiles: A focus on amino acids. Proc. Nutr. Soc. 2012, 71, 634–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangel-Huerta, O.D.; Pastor-Villaescusa, B.; Gil, A. Are we close to defining a metabolomic signature of human obesity? A systematic review of metabolomics studies. Metabolomics 2019, 15, 1–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Townsend, M.K.; Clish, C.B.; Kraft, P.; Wu, C.; Souza, A.L.; Deik, A.A.; Tworoger, S.S.; Wolpin, B.M. Reproducibility of Metabolomic Profiles among Men and Women in 2 Large Cohort Studies. Clin. Chem. 2013, 59, 1657–1667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carayol, M.; Licaj, I.; Achaintre, D.; Sacerdote, C.; Vineis, P.; Key, T.J.; Moret, N.C.O.; Scalbert, A.; Rinaldi, S.; Ferrari, P. Reliability of Serum Metabolites over a Two-Year Period: A Targeted Metabolomic Approach in Fasting and Non-Fasting Samples from EPIC. PLoS ONE 2015, 10, e0135437. [Google Scholar] [CrossRef] [Green Version]
- Sampson, J.N.; Boca, S.M.; Shu, X.O.; Stolzenberg-Solomon, R.Z.; Matthews, C.E.; Hsing, A.W.; Tan, Y.T.; Ji, B.-T.; Chow, W.-H.; Cai, Q.; et al. Metabolomics in Epidemiology: Sources of Variability in Metabolite Measurements and Implications. Cancer Epidemiol. Prev. Biomark. 2013, 22, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Bennett, B.J.; de Aguiar Vallim, T.Q.; Wang, Z.; Shih, D.M.; Meng, Y.; Gregory, J.; Allayee, H.; Lee, R.; Graham, M.; Crooke, R.; et al. Trimethylamine-N-Oxide, a Metabolite Associated with Atherosclerosis, Exhibits Complex Genetic and Dietary Regulation. Cell Metab. 2013, 17, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Wan, Y.; Zeleznik, O.; Baden, M.; Wu, Y.; Clish, C.; Willett, W.; Liang, L.; Eliassen, H. Mapping the Metabolic Profiles of Long-Term Vegetable, Fruit, and Fruit Juice Consumption. Curr. Dev. Nutr. 2020, 4, 787. [Google Scholar] [CrossRef]
- Tworoger, S.; Hankinson, S.E. Use of biomarkers in epidemiologic studies: Minimizing the influence of measurement error in the study design and analysis. Cancer Causes Control 2006, 17, 889–899. [Google Scholar] [CrossRef]
- Mayers, J.R.; Wu, C.; Clish, C.B.; Kraft, P.; Torrence, M.E.; Fiske, B.P.; Yuan, C.; Bao, Y.; Townsend, M.K.; Tworoger, S.S.; et al. Elevation of circulating branched-chain amino acids is an early event in human pancreatic adenocarcinoma development. Nat. Med. 2014, 20, 1193–1198. [Google Scholar] [CrossRef]
- Zeleznik, O.A.; Balasubramanian, R.; Ren, Y.; Tobias, D.K.; Rosner, B.A.; Peng, C.; Bever, A.M.; Frueh, L.; Jeanfavre, S.; Avila-Pacheco, J.; et al. Branched-Chain Amino Acids and Risk of Breast Cancer. JNCI Cancer Spectrum 2021, 5, pkab059. [Google Scholar] [CrossRef]
- Zeleznik, O.A.; Balasubramanian, R.; Zhao, Y.; Frueh, L.; Jeanfavre, S.; Avila-Pacheco, J.; Clish, C.B.; Tworoger, S.S.; Eliassen, A.H. Circulating amino acids and amino acid-related metabolites and risk of breast cancer among predominantly premenopausal women. npj Breast Cancer 2021, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dickerman, B.A.; Ebot, E.M.; Healy, B.C.; Wilson, K.M.; Eliassen, A.H.; Ascherio, A.; Pernar, C.H.; Zeleznik, O.A.; Heiden, M.G.V.; Clish, C.B.; et al. A Metabolomics Analysis of Adiposity and Advanced Prostate Cancer Risk in the Health Professionals Follow-Up Study. Metabolites 2020, 10, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, S.H.; Cui, J.; A Sparks, J.; Lu, B.; Tedeschi, S.K.; Speyer, C.B.; Moss, L.; Feser, M.L.; Kelmenson, L.B.; A Mewshaw, E.; et al. Circulating plasma metabolites and risk of rheumatoid arthritis in the Nurses’ Health Study. Rheumatology 2020, 59, 3369–3379. [Google Scholar] [CrossRef]
- Li, J.; Guasch-Ferré, M.; Chung, W.; Ruiz-Canela, M.; Toledo, E.; Corella, D.; Bhupathiraju, S.N.; Tobias, D.K.; Tabung, F.K.; Hu, J.; et al. The Mediterranean diet, plasma metabolome, and cardiovascular disease risk. Eur. Heart J. 2020, 41, 2645–2656. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Tworoger, S.S.; Eliassen, A.H.; Hankinson, S.E. Postmenopausal plasma sex hormone levels and breast cancer risk over 20 years of follow-up. Breast Cancer Res. Treat. 2013, 137, 883–892. [Google Scholar] [CrossRef] [Green Version]
- Eliassen, A.H.; Warner, E.T.; Rosner, B.; Collins, L.C.; Beck, A.H.; Quintana, L.M.; Tamimi, R.M.; Hankinson, S.E. Plasma 25-Hydroxyvitamin D and Risk of Breast Cancer in Women Followed over 20 Years. Cancer Res. 2016, 76, 5423–5430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortner, R.T.; Tworoger, S.S.; Wu, T.; Eliassen, A.H. Plasma florescent oxidation products and breast cancer risk: Repeated measures in the Nurses’ Health Study. Breast Cancer Res. Treat. 2013, 141, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Cahill, L.E.; Sacks, F.M.; Rimm, E.B.; Jensen, M.K. Cholesterol efflux capacity, HDL cholesterol, and risk of coronary heart disease: A nested case-control study in men. J. Lipid Res. 2019, 60, 1457–1464. [Google Scholar] [CrossRef]
- Jensen, M.K.; Rimm, E.B.; Furtado, J.D.; Sacks, F.M. Apolipoprotein C-III as a Potential Modulator of the Association Between HDL-Cholesterol and Incident Coronary Heart Disease. J. Am. Heart Assoc. 2012, 1, e000232. [Google Scholar] [CrossRef] [Green Version]
- Mendivil, C.O.; Rimm, E.B.; Furtado, J.; Chiuve, S.; Sacks, F.M. Low-Density Lipoproteins Containing Apolipoprotein C-III and the Risk of Coronary Heart Disease. Circulation 2011, 124, 2065–2072. [Google Scholar] [CrossRef] [Green Version]
- Hankinson, S.E.; Willett, W.C.; Michaud, M.S.; Manson, J.E.; Colditz, G.A.; Longcope, C.; Rosner, B.; Speizer, F.E. Plasma Prolactin Levels and Subsequent Risk of Breast Cancer in Postmenopausal Women. JNCI J. Natl. Cancer Inst. 1999, 91, 629–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tworoger, S.S.; Sluss, P.; Hankinson, S.E. Association between Plasma Prolactin Concentrations and Risk of Breast Cancer among Predominately Premenopausal Women. Cancer Res. 2006, 66, 2476–2482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittenbecher, C.; Guasch-Ferré, M.; Haslam, D.E.; Dennis, C.; Li, J.; Bhupathiraju, S.N.; Lee, C.H.; Qi, Q.; Liang, L.; Eliassen, A.H.; et al. Changes in metabolomics profiles over ten years and subsequent risk of developing type 2 diabetes: Results from the Nurses′ Health Study. EBioMedicine 2022, 75, 103799. [Google Scholar] [CrossRef] [PubMed]
- Mascanfroni, I.D.; Takenaka, M.C.; Yeste, A.; Patel, B.; Wu, Y.; Kenison, J.E.; Siddiqui, S.; Basso, A.S.; Otterbein, L.E.; Pardoll, D.M.; et al. Metabolic control of type 1 regulatory T cell differentiation by AHR and HIF1-α. Nat. Med. 2015, 21, 638–646. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.F.; Morningstar, J.E.; Yang, Q.; Zheng, B.; Gao, Y.; Jeanfavre, S.; Scott, J.; Fernandez, C.; Zheng, Z.; O′Connor, S.; et al. Dimethylguanidino valeric acid is a marker of liver fat and predicts diabetes. J. Clin. Investig. 2017, 127, 4394–4402. [Google Scholar] [CrossRef] [PubMed]
- Paynter, N.P.; Balasubramanian, R.; Giulianini, F.; Wang, D.D.; Tinker, L.F.; Gopal, S.; Deik, A.A.; Bullock, K.; Pierce, K.A.; Scott, J.; et al. Metabolic Predictors of Incident Coronary Heart Disease in Women. Circulation 2018, 137, 841–853. [Google Scholar] [CrossRef]
- Rosner, B.; Glynn, R.J. Interval estimation for rank correlation coefficients based on the probit transformation with extension to measurement error correction of correlated ranked data. Stat. Med. 2007, 26, 633–646. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Metabolites | Median | Quartile 1−Quartile 3 |
|---|---|---|
| All metabolites | 0.43 | 0.36–0.50 |
| Lipids and lipid-related metabolites | 0.44 | 0.38–0.51 |
| Polar metabolites | 0.42 | 0.33–0.49 |
| Metabolites with CV < 25% | 0.44 | 0.38–0.51 |
| Metabolites with CV ≥ 25% | 0.34 | 0.28–0.42 |
| Participants | Median | Quartile 1–Quartile 3 |
|---|---|---|
| All women | 0.43 | 0.36–0.50 |
| Fasting women | 0.45 | 0.37–0.52 |
| Women with stable BMI | 0.43 | 0.36–0.51 |
| Women with a change in BMI * | 0.41 | 0.33–0.48 |
| Postmenopausal women not using hormone therapy | 0.44 | 0.36–0.53 |
| Control women | 0.44 | 0.37–0.51 |
| Intra-Class Correlation | ||||||
|---|---|---|---|---|---|---|
| Metabolite Name | Metabolite Class | All | Fasting | Stable BMI | Unstable BMI | |
| Most stable metabolites | N6,N6-dimethyllysine | Organic acids and derivatives | 0.82 | 0.84 | 0.83 | 0.83 |
| Dimethylguanidino valerate | Other | 0.72 | 0.73 | 0.73 | 0.71 | |
| N-acetylornithine | Organic acids and derivatives | 0.69 | 0.72 | 0.70 | 0.68 | |
| C34:2 PC plasmalogen | Phosphatidylcholine plasmalogens | 0.66 | 0.67 | 0.66 | 0.61 | |
| C38:4 PC | Phosphatidylcholines | 0.65 | 0.66 | 0.66 | 0.63 | |
| Glycine | Amino acids | 0.65 | 0.64 | 0.65 | 0.62 | |
| C5-DC carnitine | Carnitines | 0.64 | 0.63 | 0.66 | 0.62 | |
| N4-acetylcytidine | Nucleosides, nucleotides, and analogues | 0.64 | 0.62 | 0.68 | 0.62 | |
| (A)Symmetric dimethylarginine | Organic acids and derivatives | 0.62 | 0.63 | 0.66 | 0.65 | |
| C36:1 PE plasmalogen | Phosphatidylethanolamine plasmalogens | 0.62 | 0.62 | 0.64 | 0.59 | |
| Least stable metabolites | 1-methylhistidine | Other | 0.21 | 0.18 | 0.21 | 0.25 |
| 4-hydroxyhippurate | Other | 0.21 | 0.19 | 0.18 | 0.26 | |
| Acetaminophen * | Other | 0.2 | 0.18 | 0.21 | 0.21 | |
| Guanosine | Other | 0.2 | 0.20 | 0.17 | 0.23 | |
| Allantoin | Other | 0.18 | 0.21 | 0.15 | 0.20 | |
| Hydroxyproline | Carboxylic acids and derivatives | 0.18 | 0.12 | 0.17 | 0.17 | |
| Methyl N-methylanthranilate | Other | 0.17 | 0.13 | 0.14 | 0.22 | |
| Trimethylamine-N-oxide | Other | 0.16 | 0.15 | 0.12 | 0.24 | |
| Ectoine | Other | 0.09 | 0.08 | 0.08 | 0.10 | |
| Palmitoylethanolamide | Other | 0.05 | 0.03 | 0.05 | 0.06 | |
| First Collection | Second Collection | |
|---|---|---|
| n | 1880 | 1184 |
| Age, y | 55.57 (6.92) | 66.46 (6.87) |
| BMI, kg/m2 | 25.37 (4.53) | 26.57 (5.11) |
| Physical activity, MET-hrs/wk | 16.34 (20.00) | 19.58 (20.78) |
| Alcohol consumption, g/day | 6.71 (10.95) | 5.81 (9.45) |
| AHEI ^ | 47.31 (10.67) | 50.16 (9.98) |
| Menopausal status, % | ||
| Premenopausal | 479 (25.5) | 8 (0.7) |
| Postmenopausal, no PMH # use | 577 (30.7) | 374 (31.6) |
| Postmenopausal, PMH # use | 587 (31.2) | 788 (66.6) |
| Missing/Dubious | 237 (12.6) | 14 (1.2) |
| Fasting (>8 h), % | 1309 (69.6) | 1062 (89.7) |
| Smoking, % | ||
| Never | 888 (47.4) | 551 (46.7) |
| Past | 748 (39.9) | 575 (48.7) |
| Current | 238 (12.7) | 55 (4.7) |
| Race, % | ||
| White | 1853 (98.6) | 1173 (99.1) |
| Black | 14 (0.7) | 4 (0.3) |
| Asian | 10 (0.5) | 5 (0.4) |
| Other/missing | 3 (0.2) | 2 (0.2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeleznik, O.A.; Wittenbecher, C.; Deik, A.; Jeanfavre, S.; Avila-Pacheco, J.; Rosner, B.; Rexrode, K.M.; Clish, C.B.; Hu, F.B.; Eliassen, A.H. Intrapersonal Stability of Plasma Metabolomic Profiles over 10 Years among Women. Metabolites 2022, 12, 372. https://doi.org/10.3390/metabo12050372
Zeleznik OA, Wittenbecher C, Deik A, Jeanfavre S, Avila-Pacheco J, Rosner B, Rexrode KM, Clish CB, Hu FB, Eliassen AH. Intrapersonal Stability of Plasma Metabolomic Profiles over 10 Years among Women. Metabolites. 2022; 12(5):372. https://doi.org/10.3390/metabo12050372
Chicago/Turabian StyleZeleznik, Oana A., Clemens Wittenbecher, Amy Deik, Sarah Jeanfavre, Julian Avila-Pacheco, Bernard Rosner, Kathryn M. Rexrode, Clary B. Clish, Frank B. Hu, and A. Heather Eliassen. 2022. "Intrapersonal Stability of Plasma Metabolomic Profiles over 10 Years among Women" Metabolites 12, no. 5: 372. https://doi.org/10.3390/metabo12050372