
First Evidence Indicates the Physiology- and Axial-Myopia-Dependent Profiles of Steroid Hormones in Aqueous Humor
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Clinical Samples
2.3. Extraction of SHs from AH and Plasma
2.4. Identification and Quantification of SHs Using LC MS/MS
2.5. Quantification of SHs in AH and Plasma
2.6. Quality Control of MRM Signals to Monitor SHs
2.7. Proteomic Analysis of AH and Plasma
2.8. Statistical Analysis for AH SHs
3. Results
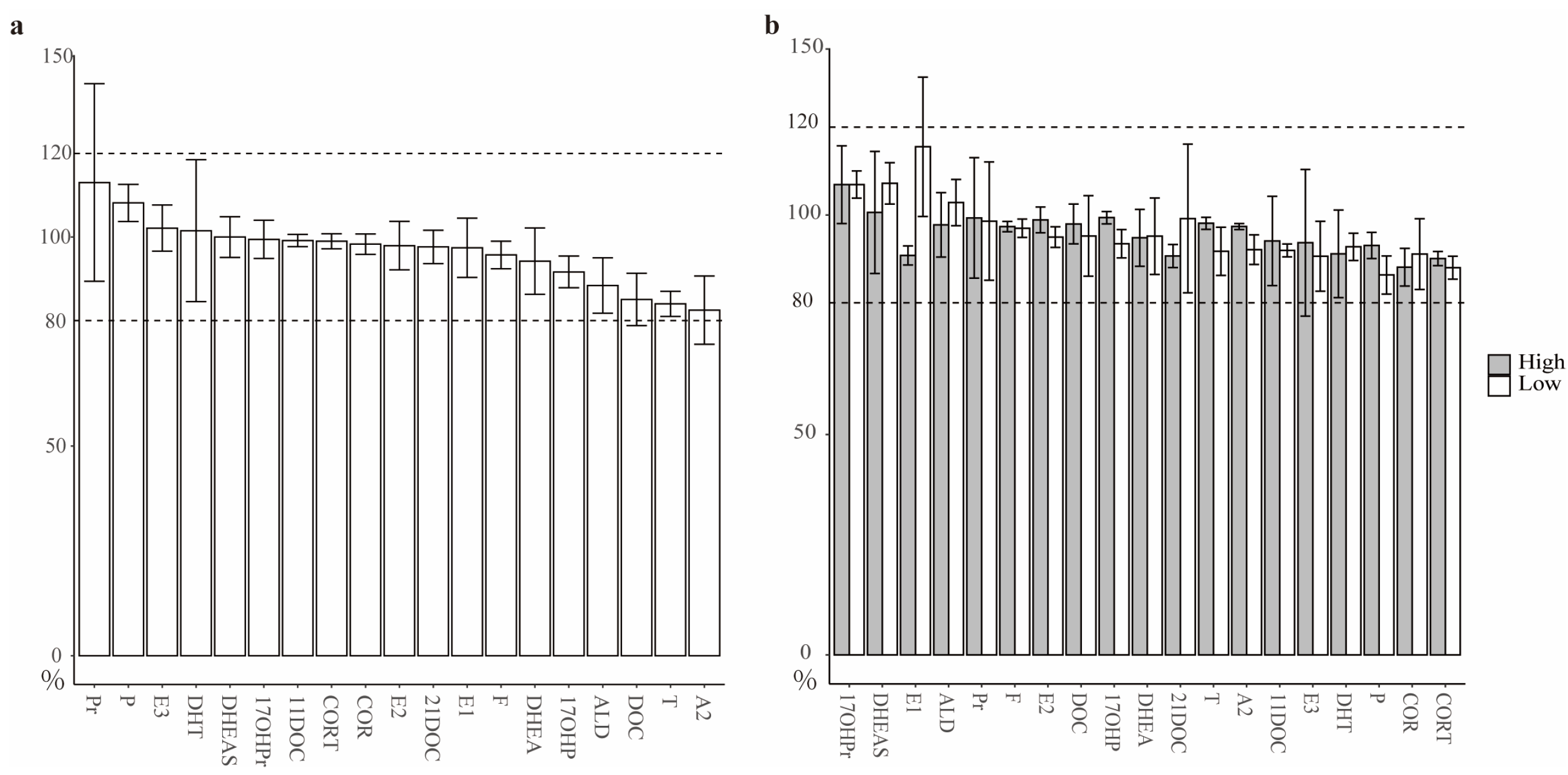
3.1. Assessments toward the Quantification of SHs in AH
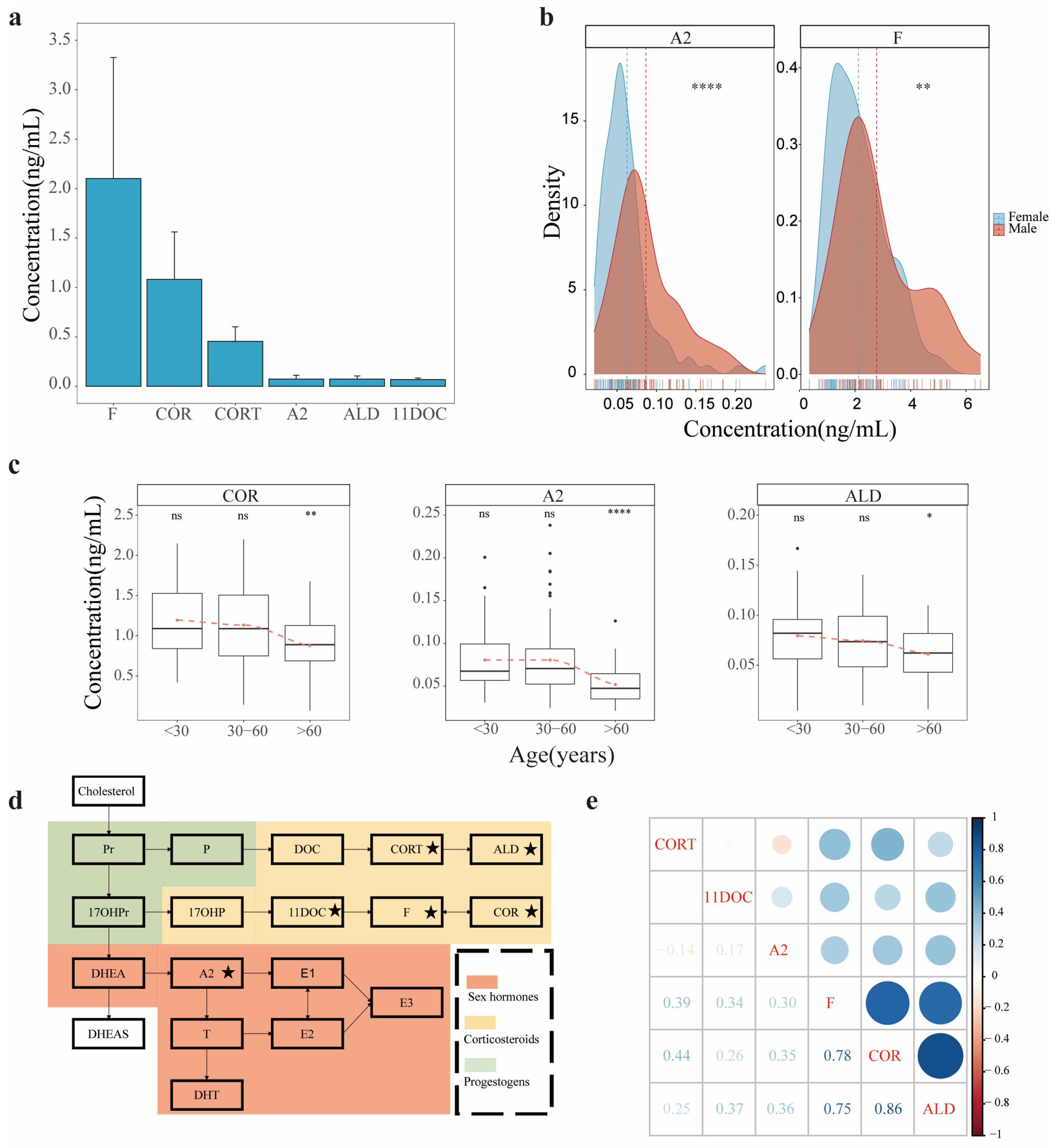
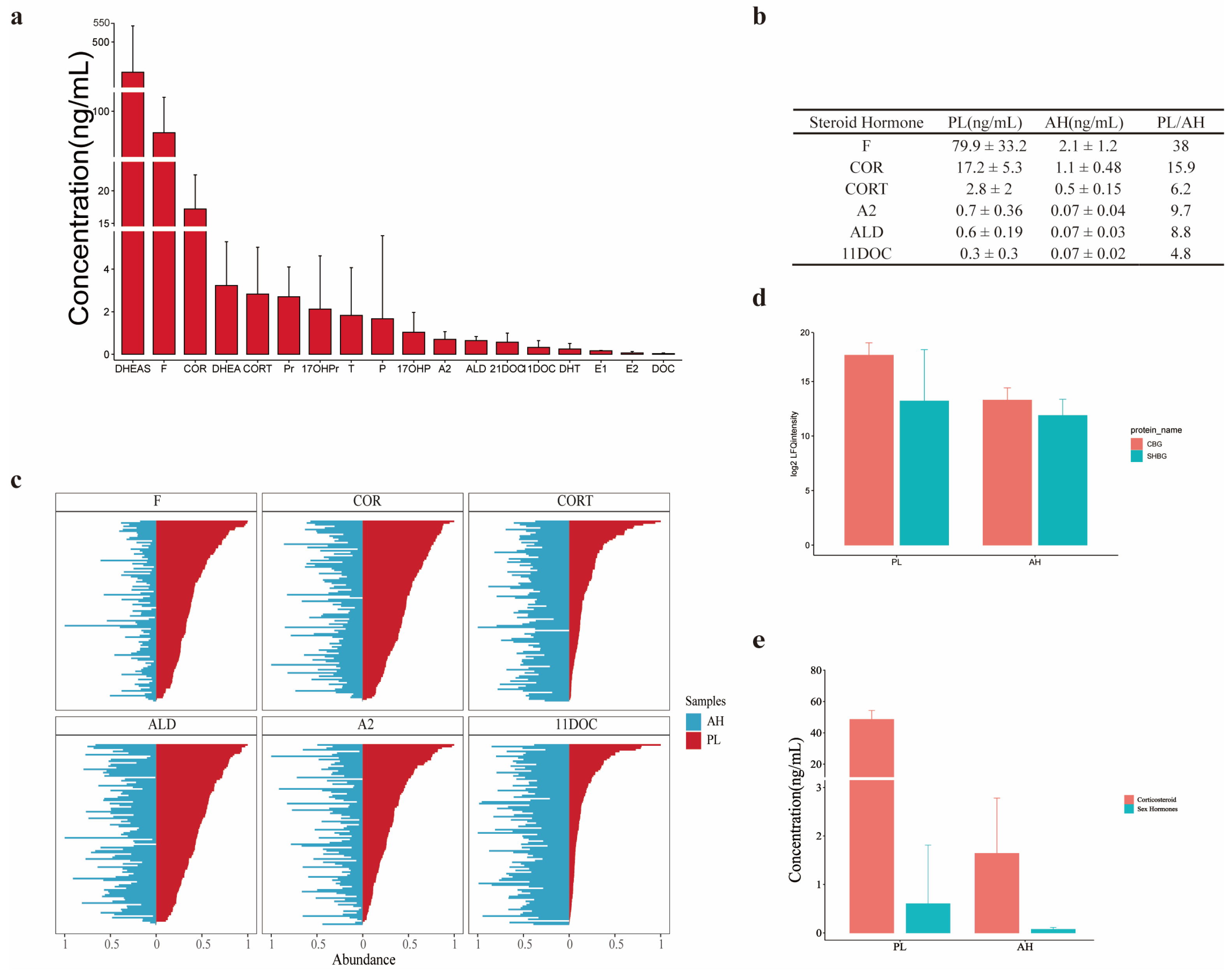
3.2. Profiles of SHs in AH
3.3. Comparison of SH Profiles between AH and Plasma
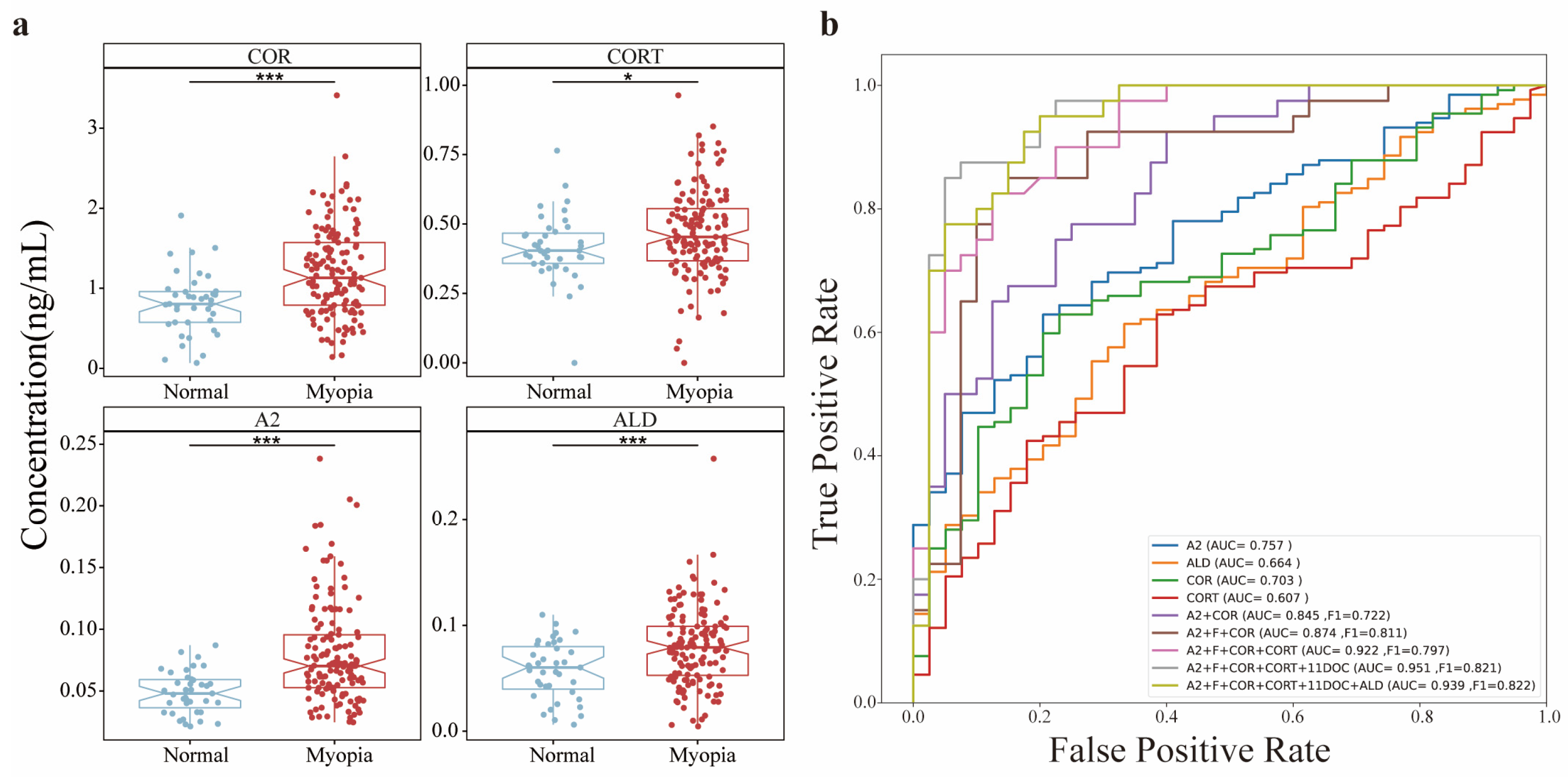
3.4. Abundance Responses of AH SHs to Axial Myopia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghayee, H.K.; Auchus, R.J. Basic concepts and recent developments in human steroid hormone biosynthesis. Rev. Endocr. Metab. Disord. 2007, 8, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Noyola-Martínez, N.; Halhali, A.; Barrera, D. Steroid hormones and pregnancy. Gynecol. Endocrinol. 2019, 35, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Rege, J.; Turcu, A.F.; Else, T.; Auchus, R.J.; Rainey, W.E. Steroid biomarkers in human adrenal disease. J. Steroid Biochem. Mol. Biol. 2019, 190, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Del Río, J.P.; Alliende, M.I.; Molina, N.; Serrano, F.G.; Molina, S.; Vigil, P. Steroid hormones and their action in women’s brains: The importance of hormonal balance. Front. Public Health 2018, 6, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdivielso, J.M.; Jacobs-Cachá, C.; Soler, M.J. Sex hormones and their influence on chronic kidney disease. Curr. Opin. Nephrol. Hypertens. 2019, 28, 1–9. [Google Scholar] [CrossRef]
- Chen, C.; Gong, X.; Yang, X.; Shang, X.; Du, Q.; Liao, Q.; Xie, R.; Chen, Y.; Xu, J. The roles of estrogen and estrogen receptors in gastrointestinal disease. Oncol. Lett. 2019, 18, 5673–5680. [Google Scholar] [CrossRef] [Green Version]
- Vegeto, E.; Villa, A.; Della Torre, S.; Crippa, V.; Rusmini, P.; Cristofani, R.; Galbiati, M.; Maggi, A.; Poletti, A. The role of sex and sex hormones in neurodegenerative diseases. Endocr. Rev. 2020, 41, 273–319. [Google Scholar] [CrossRef]
- Vaidya, A.; Carey, R.M. Evolution of the primary aldosteronism syndrome: Updating the approach. J. Clin. Endocrinol. Metab. 2020, 105, 3771–3783. [Google Scholar] [CrossRef]
- Tritos, N.A. Adrenally directed medical therapies for cushing syndrome. J. Clin. Endocrinol. Metab. 2021, 106, 16–25. [Google Scholar] [CrossRef]
- Merke, D.P.; Bornstein, S.R. Congenital adrenal hyperplasia. Lancet 2005, 365, 2125–2136. [Google Scholar] [CrossRef]
- Ye, L.; Zhao, Z.; Ren, H.; Wang, W.; Zhou, W.; Zheng, S.; Han, R.; Zhang, J.; Li, H.; Wan, Z. A Multiclassifier System to Identify and Subtype Congenital Adrenal Hyperplasia Based on Circulating Steroid Hormones. J. Clin. Endocrinol. Metab. 2022, 107, e3304–e3312. [Google Scholar] [CrossRef] [PubMed]
- Chortis, V.; Bancos, I.; Nijman, T.; Gilligan, L.C.; Taylor, A.E.; Ronchi, C.L.; O’Reilly, M.W.; Schreiner, J.; Asia, M.; Riester, A. Urine steroid metabolomics as a novel tool for detection of recurrent adrenocortical carcinoma. J. Clin. Endocrinol. Metab. 2020, 105, e307–e318. [Google Scholar] [CrossRef] [PubMed]
- Sosvorova, L.; Hill, M.; Mohapl, M.; Vitku, J.; Hampl, R. Steroid hormones in prediction of normal pressure hydrocephalus. J. Steroid Biochem. Mol. Biol. 2015, 152, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Schellevis, R.L.; Altay, L.; Kalisingh, A.; Mulders, T.W.; Sitnilska, V.; Hoyng, C.B.; Boon, C.J.; Groenewoud, J.M.; de Jong, E.K.; den Hollander, A.I. Elevated steroid hormone levels in active chronic central serous chorioretinopathy. Investig. Ophth. Vis. Sci. 2019, 60, 3407–3413. [Google Scholar] [CrossRef] [Green Version]
- McKay, T.B.; Priyadarsini, S.; Karamichos, D. Sex Hormones, Growth Hormone, and the Cornea. Cells 2022, 11, 224. [Google Scholar] [CrossRef]
- Costagliola, C.; dell’Omo, R.; Agnifili, L.; Bartollino, S.; Fea, A.M.; Uva, M.G.; Zeppa, L.; Mastropasqua, L. How many aqueous humor outflow pathways are there? Surv. Ophthalmol. 2020, 65, 144–170. [Google Scholar] [CrossRef]
- Zhang, X.-H.; Sun, H.-M.; Ji, J.; Zhang, H.; Ma, W.-J.; Jin, Z.; Yuan, J.-Q. Sex hormones and their receptors in patients with age-related cataract. J. Cataract. Refract. Surg. 2003, 29, 71–77. [Google Scholar] [CrossRef]
- Xu, G.; Zheng, J.; Cai, J.; Zhang, J.; Liu, J.; Tang, S. Corticosteroid-induced intraocular pressure elevation in the pediatric patients: A protocol for systematic review. Medicine 2021, 100, e28189. [Google Scholar] [CrossRef]
- Phulke, S.; Kaushik, S.; Kaur, S.; Pandav, S. Steroid-induced glaucoma: An avoidable irreversible blindness. J. Curr. Glaucoma. Pract. 2017, 11, 67. [Google Scholar] [CrossRef]
- Fini, M.E.; Schwartz, S.G.; Gao, X.; Jeong, S.; Patel, N.; Itakura, T.; Price, M.O.; Price, F.W., Jr.; Varma, R.; Stamer, W.D. Steroid-induced ocular hypertension/glaucoma: Focus on pharmacogenomics and implications for precision medicine. Prog. Retin. Eye Res. 2017, 56, 58–83. [Google Scholar] [CrossRef]
- Kapugi, M.; Cunningham, K. Corticosteroids. Orthop. Nurs. 2019, 38, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.C.; Silber, R.H. A quantitative color reaction for cortisone and related 17, 21-dihydroxy-20-ketosteroids. J. Biol. Chem. 1950, 185, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Yang, H.; Wei, C.C.; Xu, L.; Wei, W.B.; Jonas, J.B. High myopia as risk factor for the 10-year incidence of open-angle glaucoma in the Beijing Eye Study. Br. J. Ophthalmol. 2022, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.; Wang, H.; Jiang, Z. Axial length and its relationship to refractive error in Chinese university students. Cont. Lens Anterior Eye 2022, 45, 101470. [Google Scholar] [CrossRef]
- Brodie, R.; Hill, H. Validation issues arising from the new FDA guidance for industry on bioanalytical method validation. Chromatographia 2002, 55, S91–S94. [Google Scholar] [CrossRef]
- Ju, W.; Wei, Z.; Shuoping, Z.; Keqiang, Y.; Miao, J.; Huiling, H.; Zhen, M.; Fei, G.; Guangxiu, L.; Yan, R.; et al. An increase of phosphatidylcholines in follicular fluid implies attenuation of embryo quality on day 3 post-fertilization. BMC Biol. 2021, 19, 200. [Google Scholar] [CrossRef]
- Vermeulen, A. Andropause. Maturitas 2000, 34, 5–15. [Google Scholar] [CrossRef]
- Ferrari, E.; Cravello, L.; Muzzoni, B.; Casarotti, D.; Paltro, M.; Solerte, S.; Fioravanti, M.; Cuzzoni, G.; Pontiggia, B.; Magri, F. Age-related changes of the hypothalamic-pituitary-adrenal axis: Pathophysiological correlates. Eur. J. Endocrinol. 2001, 144, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Hammond, G.L. Plasma steroid-binding proteins: Primary gatekeepers of steroid hormone action. J. Endocrinol. 2016, 230, R13–R25. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.-Y.; Song, G.; Lei, F.; Li, D.; Qu, Y. Avian corticosteroid-binding globulin: Biological function and regulatory mechanisms in physiological stress responses. Front. Zool. 2021, 18, 1–11. [Google Scholar] [CrossRef]
- Simons, P.I.; Valkenburg, O.; Stehouwer, C.D.; Brouwers, M.C. Sex hormone–binding globulin: Biomarker and hepatokine? Trends Endocrinol. Metab. 2021, 32, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Asaba, H.; Hosoya, K.i.; Takanaga, H.; Ohtsuki, S.; Tamura, E.; Takizawa, T.; Terasaki, T. Blood—Brain barrier is involved in the efflux transport of a neuroactive steroid, dehydroepiandrosterone sulfate, via organic anion transporting polypeptide 2. J. Neurochem. 2000, 75, 1907–1916. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, P.; Lazon de la Jara, P.; Arumugam, B.; Bullimore, M.A. Axial length targets for myopia control. Ophthalmic Physiol. Opt. 2021, 41, 523–531. [Google Scholar] [CrossRef]
- Yue, Y.; Hsiao, Y.-W.; Zhou, J.-B. Association between MMP/TIMP levels in the aqueous humor and plasma with axial lengths in myopia patients. BioMed Res. Int. 2020, 2020, 2961742. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Rao, J.; Rong, X.; Lou, S.; Zheng, Z.; Lu, Y. Metabolic characterization of human aqueous humor in relation to high myopia. Exp. Eye Res. 2017, 159, 147–155. [Google Scholar] [CrossRef]
- Barbas-Bernardos, C.; Armitage, E.G.; García, A.; Mérida, S.; Navea, A.; Bosch-Morell, F.; Barbas, C. Looking into aqueous humor through metabolomics spectacles—Exploring its metabolic characteristics in relation to myopia. J. Pharm. Biomed. Anal. 2016, 127, 18–25. [Google Scholar] [CrossRef]
- Roberti, G.; Oddone, F.; Agnifili, L.; Katsanos, A.; Michelessi, M.; Mastropasqua, L.; Quaranta, L.; Riva, I.; Tanga, L.; Manni, G. Steroid-induced glaucoma: Epidemiology, pathophysiology, and clinical management. Surv. Ophthalmol. 2020, 65, 458–472. [Google Scholar] [CrossRef]
- Nowroozzadeh, M.H.; Thornton, S.; Watson, A.; Syed, Z.A.; Razeghinejad, R. Ocular manifestations of endocrine disorders. Clin. Exp. Optom. 2022, 105, 105–116. [Google Scholar] [CrossRef]
- Abolanle, K.A.; Akram, M.; Titilola, K.O.; Zainab, R.; Daniyal, M.; Olubuloa, A.O.; Sunday, O.O. Awareness and Current Knowledge of Cushing’s Syndrome. Biomed. J. Sci. Tech. Res. 2020, 31, 24117–24123. [Google Scholar] [CrossRef]
- Bullimore, M.A.; Ritchey, E.R.; Shah, S.; Leveziel, N.; Bourne, R.R.; Flitcroft, D.I. The risks and benefits of myopia control. Ophthalmology 2021, 128, 1561–1579. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case Number | Case Group | Age | Gender | Eye | Diopter | Degree of Turbidity | AL (od mm) | AL (os mm) | Correspond Plasma |
|---|---|---|---|---|---|---|---|---|---|
| 20201225P0037 | ARC | 73 | female | od | −3.25DC*177 | C4N4P3 | 22 | 21.8 | yes |
| 20201225P0002 | ARC | 85 | female | od | −1.50DC*94 | C3N3P2 | 23.27 | 23.24 | yes |
| 20201225P0005 | ARC | 53 | female | od | −3.75DS − 0.50DC*92 | C2N3P4 | 22.24 | 22.11 | yes |
| 20201225P0067 | ARC | 79 | male | od | −0.25DS − 0.70DC*5 | C2N3P2 | 23.74 | 23.82 | yes |
| 20201225P0149 | ARC | 68 | female | os | −0.25DS − 0.60DC*9 | C2N3P2 | 23.07 | 23.05 | yes |
| 20201225P0041 | ARC | 69 | female | od | −0.25DS − 0.30DC*92 | C3N2P2 | 22.34 | 22.52 | yes |
| 20201225P0012 | ARC | 84 | male | od | −0.75DC*82 | C3N4P3 | 23.58 | 23.56 | yes |
| 20201225P0034 | ARC | 77 | male | od | −2.00DS − 2.50DC*48 | C2N3P2 | 24.2 | 23.84 | yes |
| 20201225P0003 | ARC | 82 | male | od | −2.00DS − 3.00DC*110 | C2N3P2 | 24.11 | 24.29 | yes |
| 20201225P0152 | ARC | 73 | male | os | +2.00DS − 3.25DC*70 | C4N3P3 | 24.16 | 23.95 | yes |
| 20201225P0137 | ARC | 91 | male | os | −4.5DC*90 | C4N4P4 | 22.54 | 22.56 | yes |
| 20201225P0121 | ARC | 65 | female | os | +1.50DC*172 | C3N2P2 | 23 | 23.12 | yes |
| 20200918P0020 | ARC | 67 | female | od | −0.25DC*132 | C2N2P2 | 24.62 | 24.28 | yes |
| 20200918P0013 | ARC | 71 | female | od | +0.75DS + 1.00DC*90 | C2N3P2 | 23.4 | 23.66 | yes |
| 20200918P0030 | ARC | 76 | male | od | −1.50DS − 2.00DC*96 | C3N3P3 | 23.12 | 23.41 | yes |
| 20200918P0019 | ARC | 73 | male | od | +0.75DS + 1.00DC*180 | C3N3P3 | 24.29 | 24.17 | yes |
| 20200918P0014 | ARC | 77 | female | os | −0.50DC*15 | C2N3P2 | 23.39 | 23.55 | yes |
| 20200918P0032 | ARC | 74 | male | od | −1.50DS | C3N3P3 | 23.39 | 23.33 | yes |
| 20200918P0044 | ARC | 78 | male | od | −0.50DC*135 | C4N4P2 | 23.56 | 23.7 | |
| 20200918P0046 | ARC | 50 | male | os | −0.75DC*77 | C3N3P3 | 23.33 | 23.58 | |
| 20201225P0015 | ARC | 61 | female | od | −-0.25DC*50 | C2N3P2 | 22.01 | 22.1 | |
| 20201225P0147 | ARC | 80 | female | os | −0.25DC*149 | C3N3P2 | 24.6 | 24.72 | |
| 20201225P0072 | ARC | 69 | female | os | +3.00DS − 3.00DC*25 | C4N4P3 | 24.54 | 24.8 | |
| 20201225P0140 | ARC | 62 | female | od | +2.00DS | C2N2P2 | 22.56 | 22.61 | |
| 20201225P0160 | ARC | 61 | male | os | −1.25DC*77 | C2N3P2 | 24.11 | 23.92 | |
| 20201225P0033 | ARC | 83 | male | os | −2.00DC*85 | C4N3P3 | 23.3 | 23.27 | |
| 20201225P0043 | ARC | 64 | female | os | −0.75DC*153 | C2N3P2 | 23.66 | 23.63 | |
| 20201225P0095 | ARC | 66 | male | od | +1.25DS + 0.25DC*75 | C3N3P2 | 23.99 | 23.88 | |
| 20201225P0009 | ARC | 63 | female | od | +3.00DS | C2N3P2 | 23.74 | 23.32 | |
| 20201225P0071 | ARC | 80 | male | od | −2.50DC*111 | C2N3P2 | 22.22 | 22.8 | yes |
| 20201225P0019 | ARC | 55 | female | od | −0.50DC*160 | C4N4P3 | 24.47 | 24.34 | yes |
| 20201225P0133 | ARC | 79 | female | od | −0.77DC*101 | C2N3P2 | 22.43 | 22.33 | yes |
| 20201225P0065 | ARC | 59 | male | od | −1.75DS − 0.75DC*85 | C2N2P2 | 23.94 | 23.93 | yes |
| 20201225P0081 | HM | 27 | male | od | −6.00DS − 2.50DC*5 | 27.99 | 24.48 | yes | |
| 20201225P0045 | HM | 30 | female | od | −8.37DS − 2.37DC*178 | 27.05 | 26.23 | yes | |
| 20200918P0015 | HM | 30 | female | od | −7.00DS − 1.25DC*172 | 25.09 | 25.31 | yes | |
| 20200918P0003 | HM | 27 | male | od | −5.75DS | 26.66 | 26.68 | yes | |
| 20200918P0037 | HM | 27 | female | od | −5.00DS − 1.25DC*80 | 26.76 | 26.67 | yes | |
| 20200918P0022 | HM | 36 | female | od | −11.62DS − 1.12DC*16 | 27.2 | 27.34 | yes | |
| 20200918P0017 | HM | 26 | female | od | −6.12DS − 0.87DC*6 | 25.56 | 25.48 | yes | |
| 20200918P0048 | HM | 21 | female | od | −7.25DS − 0.87DC*179 | 25.47 | 25.48 | ||
| 20200918P0043 | HM | 20 | male | od | −14.00DS − 4.50DC*90 | 29.92 | 28.42 | ||
| 20201225P0029 | HM | 36 | female | od | −7.25DS | 26.92 | 30.27 | ||
| 20201225P0013 | HM | 30 | female | od | −5.62DS − 0.37DC*51 | 26.16 | 26.01 | ||
| 20201225P0135 | HM | 25 | female | od | −5.87DS − 1.62DC*175 | 26.51 | 27.07 | ||
| 20201225P0134 | HM | 23 | female | od | −8.37DS − 1.00DC*4 | 27.86 | 27.77 | ||
| 20201225P0079 | HM | 45 | female | od | −6.25DS − 3.50DC*176 | 25.73 | 25.58 | ||
| 20201225P0151 | HM | 36 | male | od | −5.50DS | 26.84 | 26.77 | yes | |
| 20201225P0083 | HM | 19 | male | od | −5.87DS − 0.37DC*46 | 26 | 26 | yes | |
| 20201225P0111 | HM | 28 | female | od | −10.50DS | 27.96 | 27.19 | yes | |
| 20201225P0044 | HM | 31 | female | od | −9.00DS − 5.50DC*180 | 26.95 | 26.98 | yes | |
| 20201225P0153 | HM | 32 | female | od | −13.50DS | 30.18 | 29.81 | yes | |
| 20201225P0061 | HM | 24 | female | od | −11.25DS − 1.50DC*10 | 26 | 25.82 | yes | |
| 20201225P0105 | HM | 25 | female | od | −5.25DS | 26.32 | 26.24 | yes | |
| 20201225P0008 | HM | 26 | female | od | −6.75DS − 0.25DC*93 | 25.49 | 25.51 | yes | |
| 20201225P0051 | HM | 39 | female | od | −21.00DS − 3.00DC*10 | 25.79 | 26.35 | yes | |
| 20201225P0084 | HM | 33 | male | od | −7.00DS | 25.79 | 26.35 | yes | |
| 20201225P0040 | HM | 18 | male | od | −8.62DS − 1.87DC*180 | 26.66 | 26.73 | yes | |
| 20201225P0023 | HM | 35 | female | od | −6 | 26 | 26 | yes | |
| 20201225P0154 | HM | 28 | male | od | −12.50DS − 0.75DC*228 | 28.95 | 28.49 | yes | |
| 20201225P0128 | HM | 23 | female | od | −10.50DS | 26 | 26 | yes | |
| 20201225P0062 | HM | 23 | male | od | −12.50DS − 3.50DC*158 | 28.89 | 29.1 | yes | |
| 20201225P0110 | HM | 23 | female | od | −9.25DS − 0.62DC*2 | 27 | 27.5 | yes | |
| 20201225P0001 | HM | 28 | female | od | −6.75DS | 25.36 | 25.22 | yes | |
| 20201225P0122 | HM | 28 | male | od | −10.00DS − 2.50DC*170 | 27.67 | 27.34 | yes | |
| 20201225P0060 | HM | 19 | male | od | −9.50DS − 1.75DC*180 | 26 | 26 | yes | |
| 20201225P0119 | HM | 23 | female | od | −10.00DS − 2.00DC*180 | 28.83 | 29.19 | yes | |
| 20201225P0139 | HM | 26 | male | od | −5.25DS − 2.25DC*174 | 27.13 | 26.98 | yes | |
| 20201225P0118 | HM | 42 | female | od | −7.62DS − 0.25DC*21 | 26.43 | 26.41 | yes | |
| 20200918P0021 | HM | 24 | female | od | −7.50DS − 0.50DC*2 | 26 | 26 | yes | |
| 20200918P0012 | HM | 27 | female | od | −8.12DS − 1.00DC*5 | 26.22 | 25.63 | yes | |
| 20200918P0008 | HM | 24 | female | od | −6.87DS − 2.87DC*6 | 25.97 | 26.13 | yes | |
| 20200918P0039 | HM | 27 | female | od | −5.12DS − 0.37DC*39 | 26.18 | 26.11 | yes | |
| 20200918P0036 | HMC | 51 | female | od | −8.00DS − 0.75DC*110 | C2N3P2 | 26.79 | 26.68 | yes |
| 20200918P0004 | HMC | 52 | male | od | −16.00DS − 0.75DC*4 | C2N2P2 | 30.61 | 29.17 | yes |
| 20200918P0035 | HMC | 53 | male | os | −8.50DS − 1.50DC*95 | C2N2P2 | 30.32 | 28.9 | yes |
| 20200918P0026 | HMC | 53 | male | od | −11.00DS − 2.00DC*70 | C2N3P2 | 28.4 | 27.98 | yes |
| 20200918P0007 | HMC | 53 | female | od | −7.50DS − 1.00DC*90 | C2N2P2 | 26.8 | 26.22 | yes |
| 20200918P0018 | HMC | 31 | male | os | −9.25DS | C2N2P1 | 25.95 | 25.89 | yes |
| 20200918P0011 | HMC | 51 | male | os | −10.00DS − 0.50DC*25 | C4N4P3 | 28.16 | 28.82 | yes |
| 20200918P0001 | HMC | 46 | female | os | −9.00DS/ − 0.50DC*60 | C2N2P2 | 26.63 | 25.77 | yes |
| 20200918P0005 | HMC | 50 | female | od | −10.00DS | C2N2P2 | 28.21 | 28.67 | yes |
| 20200918P0028 | HMC | 42 | male | os | −9.00DS − 0.750DC*25 | C3N2P3 | 25.36 | 27.7 | yes |
| 20200918P0034 | HMC | 47 | male | os | −11.25DS − 1.50DC*143 | C2N2P2 | 28.9 | 28.15 | yes |
| 20200918P0038 | HMC | 44 | female | od | −2.75DS | C2N2P2 | 25.89 | 25.7 | yes |
| 20200918P0002 | HMC | 54 | male | os | −7.75DS − 2.25DC*174 | C2N2P2 | 27.22 | 27.6 | yes |
| 20200918P0016 | LM | 22 | female | od | −5.87DS − 0.50DC*2 | 25.89 | 25.83 | yes | |
| 20200918P0047 | LM | 30 | female | od | −5.50DS − 0.25DC*2 | 24.89 | 24.88 | ||
| 20201225P0066 | LM | 26 | female | od | −4.50DS − 1.00DC*10 | 24.78 | 25.02 | ||
| 20201225P0014 | LM | 35 | female | od | −4.75DS − 1.12DC*150 | 24.6 | 24.16 | ||
| 20201225P0097 | LM | 26 | female | od | −7.00DS | 24.45 | 24.46 | ||
| 20201225P0059 | LM | 34 | male | od | −4.50DS − 0.37DC*136 | 25.32 | 25.37 | ||
| 20201225P0021 | LM | 30 | female | od | −5.00DS − 1.50DC*60 | 24.96 | 24.4 | ||
| 20201225P0098 | LM | 27 | female | od | −4.12DS − 0.87DC*13 | 24.91 | 24.38 | ||
| 20201225P0126 | LM | 24 | female | od | −5.50DS − 1.00DC*175 | 26.08 | 25.83 | ||
| 20201225P0068 | LM | 26 | female | od | −4.50DS | 25.14 | 25.09 | ||
| 20201225P0017 | LM | 31 | female | od | −5.25DS | 25.31 | 25.34 | ||
| 20201225P0082 | LM | 37 | female | od | −3.75DS | 23.99 | 23.93 | ||
| 20201225P0016 | LM | 18 | female | od | −5.50DS − 1.50DC*2 | 25.72 | 26.12 | ||
| 20201225P0091 | LM | 26 | female | od | −3.50DS − 1.00DC*171 | 24.08 | 24.65 | yes | |
| 20201225P0093 | LM | 29 | female | od | −5.50DS − 0.62DC*87 | 25.27 | 25.77 | yes | |
| 20201225P0094 | LM | 22 | male | od | −7.75DS − 1.25DC*8 | 24.99 | 24.61 | yes | |
| 20200918P0027 | LM | 33 | female | od | −5.00DS | 23 | 23 | yes | |
| 20200918P0031 | ARC | 68 | male | od | −0.25DC*45 | C2N3P3 | 23.78 | 23.33 | yes |
| 20200918P0040 | ARC | 60 | male | os | −0.50DC*50 | C2N2P2 | 24.63 | 24.78 | yes |
| 20201225P0030 | ARC | 66 | male | od | +0.90DC*104 | C3N3P2 | 24.03 | 23.6 | yes |
| 20201225P0052 | ARC | 73 | female | od | −0.50DC*86 | C3N4P2 | 22.67 | 22.86 | yes |
| 20201225P0116 | ARC | 83 | male | os | −1.75DC*91 | C2N3P3 | 23.58 | 23.58 | yes |
| 20201225P0108 | ARC | 84 | female | od | −1.00DC*133 | C3N4P2 | 22.62 | 22.52 | yes |
| 20201225P0085 | ARC | 73 | female | od | −1.00DC*78 | C2N3P2 | 22.6 | 22.69 | yes |
| 20201225P0109 | ARC | 89 | male | od | −1.50DC*78 | C3N3P3 | 24.35 | 24.04 | yes |
| 20201225P0039 | ARC | 53 | female | os | −0.25DS − 0.34DC*100 | C1N3P3 | 23.9 | 23.66 | yes |
| 20201225P0022 | ARC | 75 | female | os | +1.25DS + 1.50DC*170 | C2N3P2 | 23.26 | 23.29 | yes |
| 20201225P0101 | ARC | 65 | male | od | −0.25DS − 0.25DC*67 | C2N4P2 | 23.46 | 23.2 | yes |
| 20200918P0051 | ARC | 68 | female | od | −1.50DC*45 | C4N4P4 | 23.71 | 23.49 | |
| 20201225P0115 | ARC | 91 | female | os | −1.50DC*92 | C4N4P4 | 23.44 | 22.95 | |
| 20201225P0114 | ARC | 82 | female | od | −0.50DC*45 | C4N3P3 | 23.4 | 23.38 | |
| 20201225P0027 | HM | 28 | female | od | −6.37DS − 0.87DC*2 | 26.64 | 26.56 | ||
| 20201225P0092 | HM | 27 | male | od | −11.00DS − 3.00DC*5 | 29.61 | 28.36 | ||
| 20201225P0130 | HM | 31 | female | od | −8.37DS − 1.25DC*2 | 28.64 | 27.67 | ||
| 20201225P0078 | HM | 21 | male | od | −14.50DS − 0.25DC*7 | 29.32 | 28.78 | ||
| 20201225P0018 | HM | 26 | female | od | −10.00DS − 0.25DC*25 | 28.38 | 28.17 | ||
| 20201225P0035 | HM | 23 | male | od | −8.00DS − 3.00DC*175 | 29.12 | 28.76 | yes | |
| 20200918P0029 | HM | 49 | female | od | −10.25DS | 26 | 26 | yes | |
| 20200918P0041 | HM | 26 | male | od | −7.50DS − 1.75DC*175 | 27.31 | 27.43 | yes | |
| 20201225P0049 | HM | 20 | female | od | −7.25DS − 1.25DC*38 | 27.92 | 25.17 | yes | |
| 20201225P0020 | HM | 39 | female | od | −4.75DS | 26.71 | 26.74 | yes | |
| 20201225P0090 | HM | 40 | male | od | −4.75DS | 26.49 | 26.67 | yes | |
| 20201225P0142 | HMC | 58 | male | od | −5.50DS − 0.50DC*90 | C2N2P2 | 25.64 | 25.67 | yes |
| 20201225P0129 | HMC | 54 | male | od | −4.25DS − 3.00DC*170 | C2N3P3 | 26.53 | 26.15 | yes |
| 20200918P0045 | HMC | 52 | male | os | −5.00DS | C1N1P4 | 26.74 | 26.51 | |
| 20200918P0050 | HMC | 72 | female | od | −15.00DS − 0.50DC*85 | C3N4P2 | 29.72 | 26.94 | |
| 20201225P0155 | HMC | 57 | male | os | −7.00DS − 1.00DC*75 | C2N2P2 | 27.14 | 26.92 | |
| 20201225P0103 | HMC | 37 | male | od | −25.75DS | C2N2P3 | 33.8 | 25.74 | |
| 20201225P0031 | HMC | 59 | female | os | −9.25DS − 0.50DC*16 | C3N2P2 | 28 | 29.17 | |
| 20201225P0024 | HMC | 55 | female | od | −9.00DS − 0.25DC*25 | C3N4P2 | 28.05 | 25.34 | |
| 20201225P0074 | HMC | 52 | male | od | −8.50DS − 0.75DC 167 | C2N2P2 | 27.75 | 24.6 | |
| 20201225P0010 | HMC | 41 | male | od | −8.00DS − 1.00DC*5 | C2N2P2 | 26.05 | 26.12 | |
| 20201225P0146 | HMC | 58 | female | os | −16.00DS − 1.50DC*121 | C2N4P4 | 27.17 | 30.06 | |
| 20201225P0080 | HMC | 62 | male | os | −7.50DS − 1.00DC*75 | C2N2P2 | 25.79 | 26.32 | |
| 20201225P0026 | HMC | 68 | female | os | −7.00DS − 1.50DC*175 | C3N2P2 | 27.06 | 25.73 | |
| 20201225P0050 | HMC | 74 | male | os | −20.00DS − 1.50DC*155 | C2N2P2 | 31.31 | 31.31 | |
| 20201225P0028 | HMC | 48 | male | os | −14.25DS − 2.25DC*80 | C2N3P2 | 27.42 | 27.48 | |
| 20201225P0025 | HMC | 76 | female | os | −7.50DS − 0.75DC*80 | C2N2P2 | 27.6 | 28.15 | |
| 20201225P0100 | HMC | 54 | female | os | −23.00DS | C2N2P2 | 32.49 | 31.5 | yes |
| 20201225P0124 | HMC | 75 | male | os | −12.00DS − 2.50DC*65 | C3N3P2 | 28.98 | 28.99 | yes |
| 20201225P0104 | LM | 31 | female | od | −2.75DS | 24.57 | 24.47 | yes | |
| 20201225P0148 | LM | 27 | female | od | −5.00DS | 25.5 | 25.37 | yes | |
| 20201225P0141 | LM | 28 | female | od | −5.25DS − 2.00DC*5 | 25.07 | 25.96 | yes | |
| 20201225P0058 | LM | 24 | male | od | −3.25DS | 25.36 | 25.39 | yes | |
| 20201225P0159 | LM | 36 | female | od | −5.00DS | 25.61 | 25.47 | yes | |
| 20201225P0088 | LM | 30 | female | od | −4.75DS | 24.81 | 24.85 | yes | |
| 20201225P0145 | LM | 23 | female | os | −3.75DS | 25.86 | 25.86 | yes | |
| 20201225P0158 | LM | 24 | female | od | −5.25DS | 24.41 | 24.51 | yes | |
| 20201225P0138 | LM | 24 | female | od | −5.00DS | 23 | 23 | yes | |
| 20200918P0025 | LM | 31 | female | od | −5.25DS − 1.00DC*12 | 24.68 | 24.12 | yes | |
| 20200918P0006 | HM | 20 | male | od | −10.87DS − 4.37DC*1 | 30.9 | 27.88 | yes | |
| 20200918P0024 | HMC | 37 | female | od | −23.00DS | C1N1P3 | 32.04 | 30.84 | yes |
| 20200918P0023 | HMC | 41 | female | od | −25.00DS − 1.50DC*162 | C3N4P2 | 33.64 | 32.41 | yes |
| 20200918P0033 | HMC | 60 | male | od | −21.50DS | C3N3P2 | 28.68 | 29.31 | |
| 20200918P0042 | HMC | 49 | female | od | −24.00DS − 2.25DC*135 | C2N2P2 | 32.6 | 31.87 | |
| 20201225P0069 | HMC | 51 | male | od | −29.00DS | C2N2P2 | 31.48 | 30.97 | |
| 20201225P0156 | HMC | 53 | female | os | −20.00DS − 1.00DC*70 | C2N2P2 | 31.49 | 31.17 | |
| 20201225P0064 | HMC | 46 | male | od | −14.00DS | C2N2P2 | 28.8 | 28.56 | |
| 20201225P0113 | HMC | 58 | female | od | −6.00DS | C3N2P3 | 26.34 | 25.74 | |
| 20201225P0125 | HMC | 69 | male | os | −8.5DS − 1.50DC*85 | C2N2P2 | 31.42 | 30.18 | |
| 20201225P0099 | HMC | 54 | male | od | −15.50DS − 1.50DC*20 | C2N3P2 | 25.93 | 25.96 | |
| 20201225P0004 | HMC | 52 | female | os | −15.00DS − 0.50DC*7 | C2N2P2 | 29.07 | 30.06 | |
| 20201225P0144 | HMC | 50 | female | os | −3.00DS | C2N3P2 | 26.83 | 26.73 | |
| 20201225P0057 | HMC | 55 | male | od | −11.00DS | C2N2P2 | 31.18 | 32.42 | |
| 20201225P0165 | HMC | 63 | male | os | −22.50DS | C5N4P2 | 28.94 | 28.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, T.; Huang, X.; Liu, J.; Liu, X.; Zeng, K.; Yan, Z.; Mei, S.; Sun, L.; Xi, W.; Ni, J.; et al. First Evidence Indicates the Physiology- and Axial-Myopia-Dependent Profiles of Steroid Hormones in Aqueous Humor. Metabolites 2022, 12, 1220. https://doi.org/10.3390/metabo12121220
Chou T, Huang X, Liu J, Liu X, Zeng K, Yan Z, Mei S, Sun L, Xi W, Ni J, et al. First Evidence Indicates the Physiology- and Axial-Myopia-Dependent Profiles of Steroid Hormones in Aqueous Humor. Metabolites. 2022; 12(12):1220. https://doi.org/10.3390/metabo12121220
Chicago/Turabian StyleChou, Tiansheng, Xiaosheng Huang, Jiawei Liu, Xinhua Liu, Kun Zeng, Zonghui Yan, Shaoyi Mei, Liangnan Sun, Wenqun Xi, Jinglan Ni, and et al. 2022. "First Evidence Indicates the Physiology- and Axial-Myopia-Dependent Profiles of Steroid Hormones in Aqueous Humor" Metabolites 12, no. 12: 1220. https://doi.org/10.3390/metabo12121220