

Recent Updates on Source, Biosynthesis, and Therapeutic Potential of Natural Flavonoid Luteolin: A Review
 , ,
, ,  ,
,
Abstract
:1. Introduction
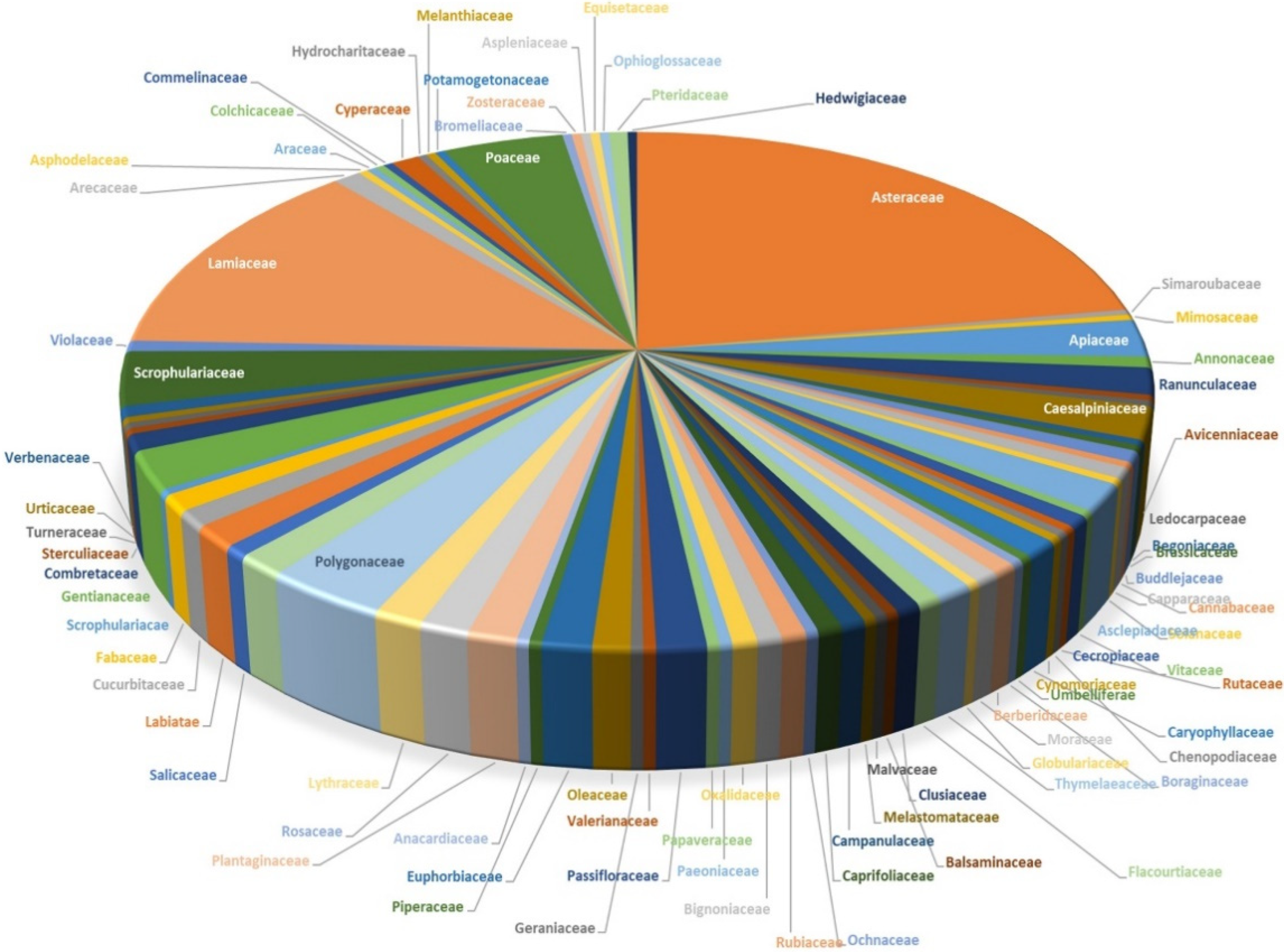
2. Source of Luteolin
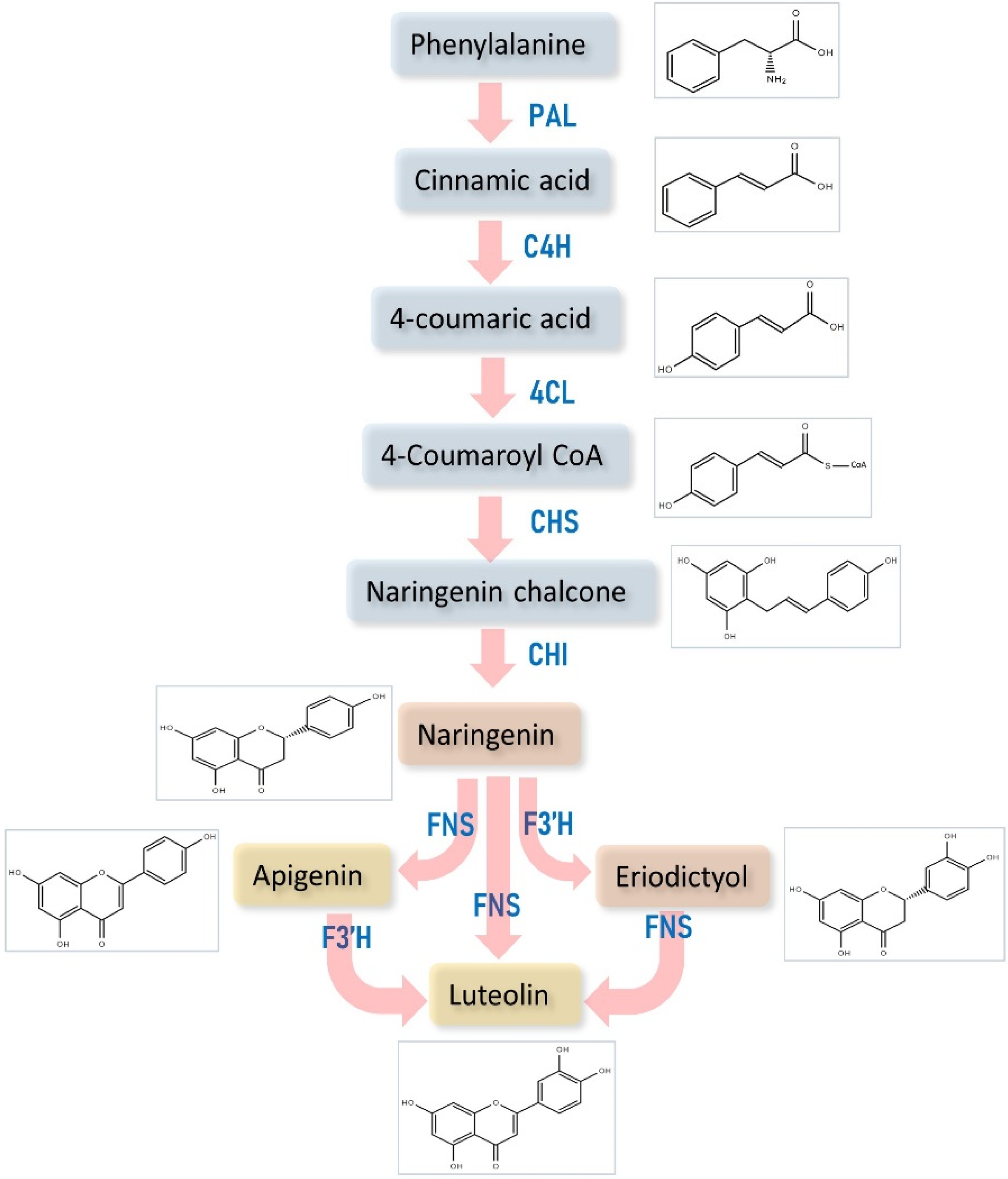
3. Luteolin Biosynthesis
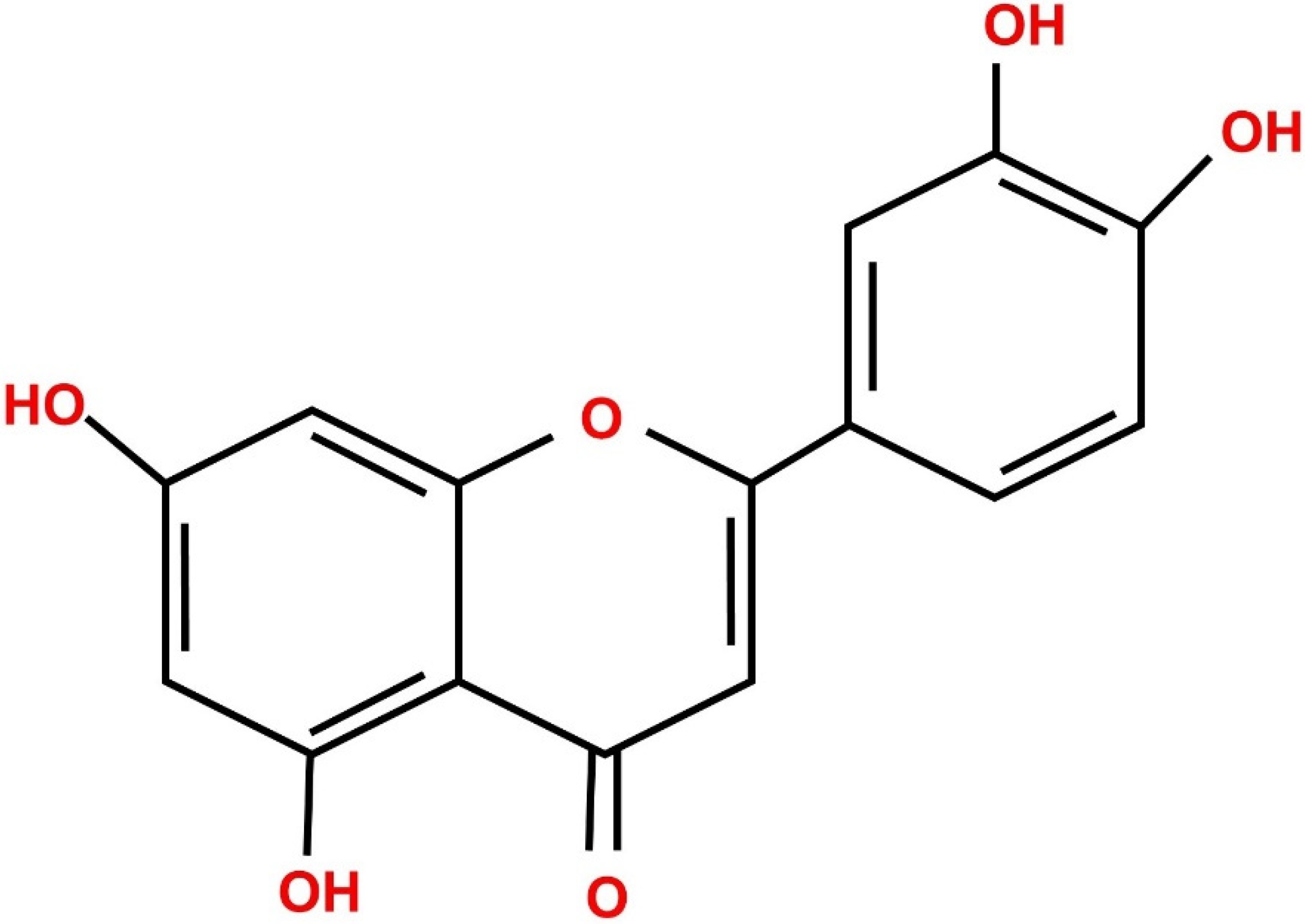
4. Physiochemical Properties of Luteolin
5. Chemopreventive Functions of Luteolin
5.1. Antioxidant Properties of Luteolin
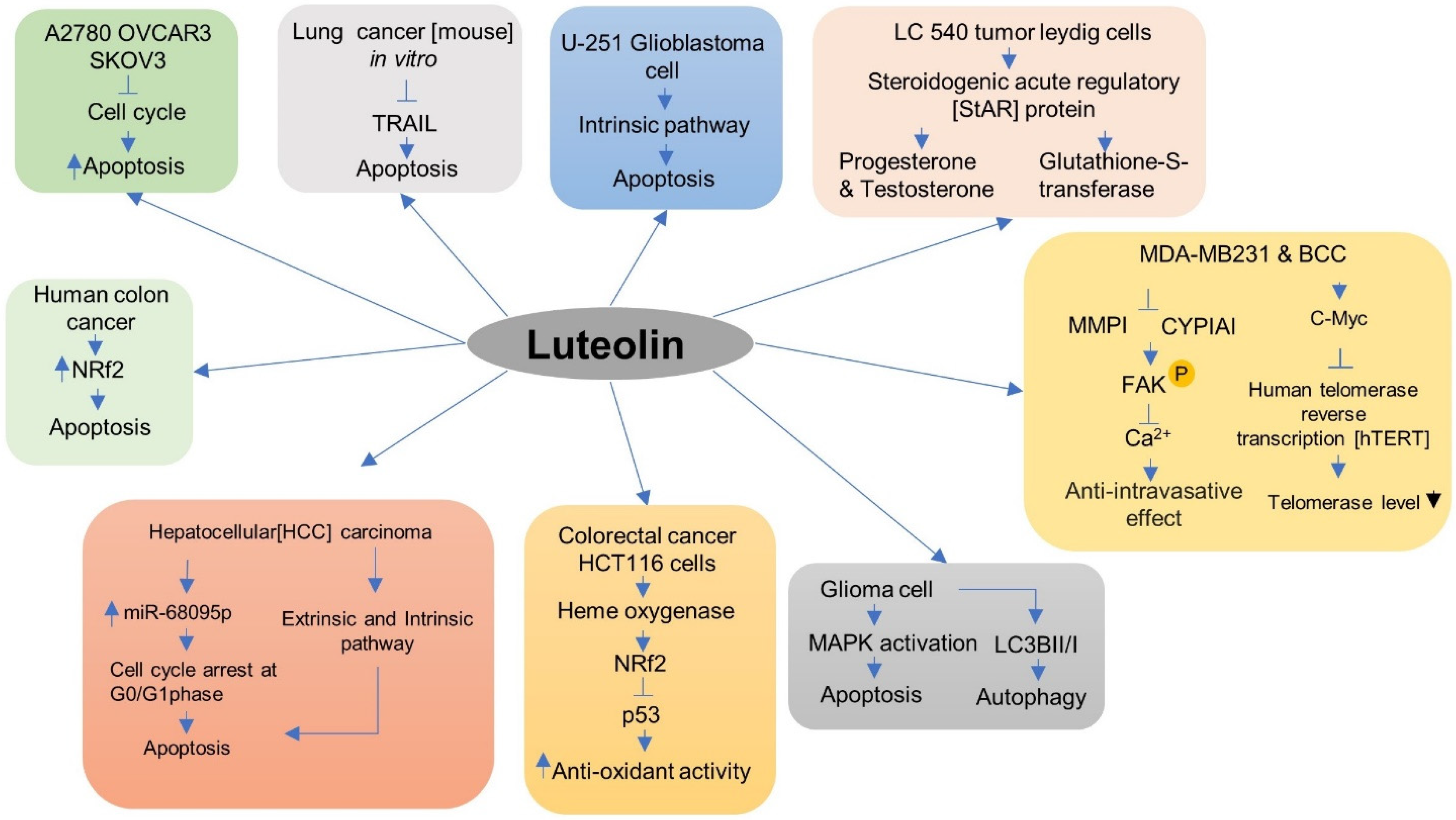
5.2. Anti-Cancer Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Cancer Cell | Mechanism | Reference |
|---|---|---|---|
| Luteolin | Colorectal cancer HCT116 cells | It increases the transcriptional activity of antioxidative response element in HCT116 cells. | [50] |
| Luteolin-7-O-glucoside and luteolin | MCF-7 cell in breast cancer | Anti-cancer activities against MC77 cells with selective index 8.0. | [51] |
| Apigenin and luteolin | MDA-MB231 breast cancer cells (BCC) immortalized lymph endothelial cell (LEC) monolayer | It suppresses pro intravasation trigger factors in MDA-MB 231 BCC, specifically MMP1 expression and CYP1A1 activity. | [52] |
| Luteolin | A2780, OVCAR3, and SKOV3 | By inducing apoptosis, arrested cell cycle thus inhibits cell invasion in ovarian cancer cells. | [53] |
| Luteolin | Myeloid leukemia cells | It triggers leukemia cells apoptosis through modulating the differential expression of PTTG1. | [54] |
| Luteolin | Lung cancer (mouse) in vitro | It enhances inhibition of tumor growth, thus decreases tumor weight and increases tumor cell apoptosis in vitro. | [33] |
| Luteolin | Tumor cells | It reduces the tumorigenic potential and inhibits the migration of U-251 glioblastoma cells. It enhances apoptosis by an intrinsic pathway. | [55] |
| Nano Luteolin | Lung cancer (H292 cell) and head and neck cancer (SSCH and TU212) cell line | Nanoluteolin inhibits the effect of tumor growth of SCCHN. | [56] |
| Luteolin | Hepatocellular (HCC) carcinoma | It represses the growth of HCC by stimulating apoptosis and cell cycle arrest at G0/G1phase in Huh7 cells at the G2/M phase; miR-68095p mediates the growth-repressive activity of luteolin in HCC. | [57] |
| Luteolin | Colon cancer cells | It induces apoptosis in doxorubicin-sensitive LoVo colon cancer cells and drug-resistant LoVo/Dx cell lines. Their cytotoxic activity in LoVo/Dx cell line was considerably lower than LoVo cell line. | [58] |
| Luetolin-7-O-glucoside | Nasopharyngeal carcinoma (NPC-039 NPC-BM) | It reduces the proliferation of NPC cell line by inducing S and G2/M cell cycle arrest by chromatin condensation at apoptosis through AKT signaling pathway. | [59] |
| Luteolin | 4TI breast cancer cell | It increases the apoptosis in 4TI BCC. | [60] |
| Luteolin | Breast cancer cell MDA-MB231 | It reduces telomerase levels in a concentration-based fashion. It inhibits phosphorylation of the NF-κB inhibitor and its target gene c-Myc to repress human telomerase reverse transcription (hTERT) expression that codes the catalytic subunit of telomerase. | [61] |
| Luteolin | Tamoxifen resistant ER (TRER) + VE Breast cancer cells | The synergistic application of luteolin and P13K, AKT, or mTOR inhibitors synergistically enhances apoptosis in TRER+VE cells. Ras gene (K-Ras, H-Ras, and N-Ras) inducer of P13K was transcriptionally suppressed by stimulation of tumor suppressor mixed-PI3K lineage leukemia 3 (MLL3) expression. | [62] |
| Luteolin | Hepatocellular cancer Hep 3B cells | It induced autophagy in p53 null Hep3B cells. | [63] |
| Luteolin | Human colon cancer | It inhibits the expression of DNA methyltransferase, a transcription repressor that enhanced the expression of the activity of ten-eleven translocation (TET) DNA methylase a transcription activator. It also increases the interaction between Nrf2 and p53, which increases the expression of antioxidative enzymes and apoptosis-related protein. | [34] |
| Luteolin | Glioma cell | It inhibits glioma cell proliferation in a time- and concentration-based fashion by glioma cell apoptosis via MAPK induction (JNK, ERK, and P38) and autophagy | [64] |
| Luteolin | LC 540 tumor Leydig cells | It activates steroidogenic acute regulatory (StAR) protein expression and increases progesterone and testosterone production. It also controls the expression of genes that participate in stress responses such as glutathione-S-transferases Gsta1 and Gstt2 and the unfolded protein response. | [65] |
| Luteolin | Amelanotic melanoma C32 (CRL-1585) cells | Luteolin and its derivatives demonstrate significant cytotoxic and pro-apoptotic potential. | [66] |
| Luteolin-7-O-glucoside | Oral squamous cell carcinoma | It reduces the oral cancer cell migration and invasion, causing a decrement in cancer metastasis by decreasing p38 phosphorylation by reducing matrix metalloproteinase (MMP)-2 expression. It exerts an anti-migratory effect by inhibiting P38-induced enhanced expression of MMP-2 and also by the extracellular signal regulatory kinase pathway. | [67] |
5.3. Anti-Diabetic Activity
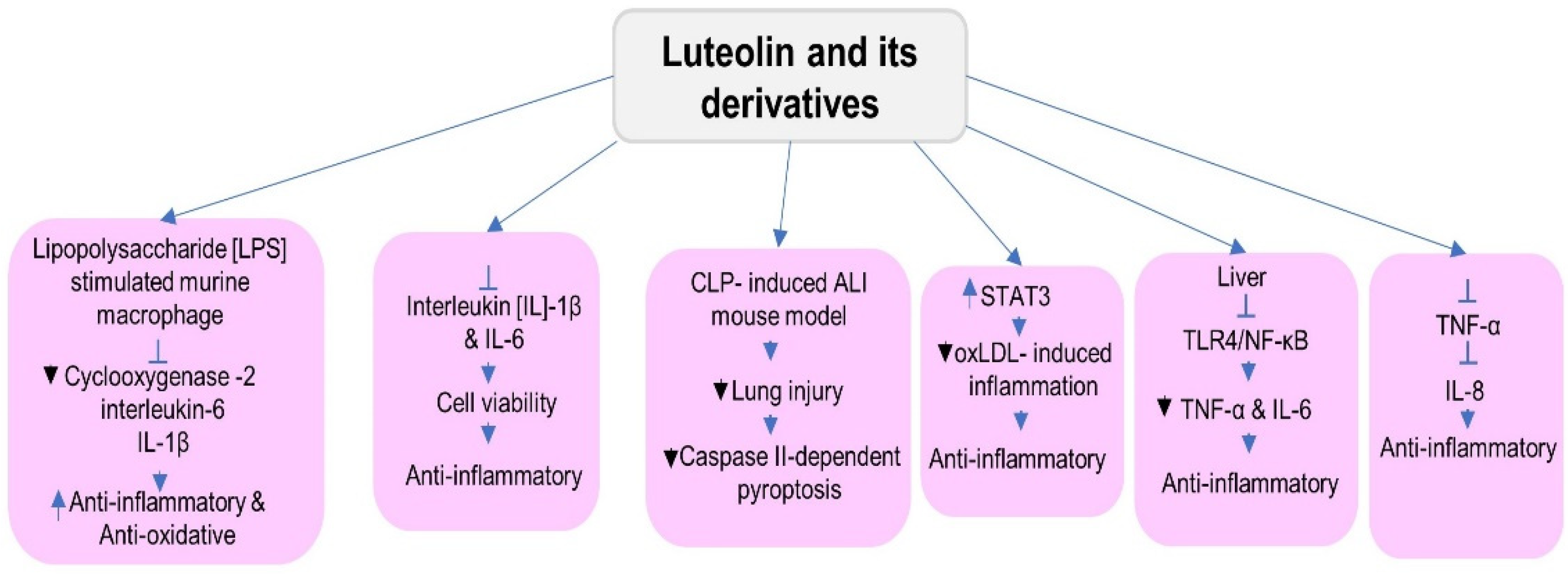
5.4. Anti-Inflammatory Activities
5.5. Protection against Alzheimer’s Disease
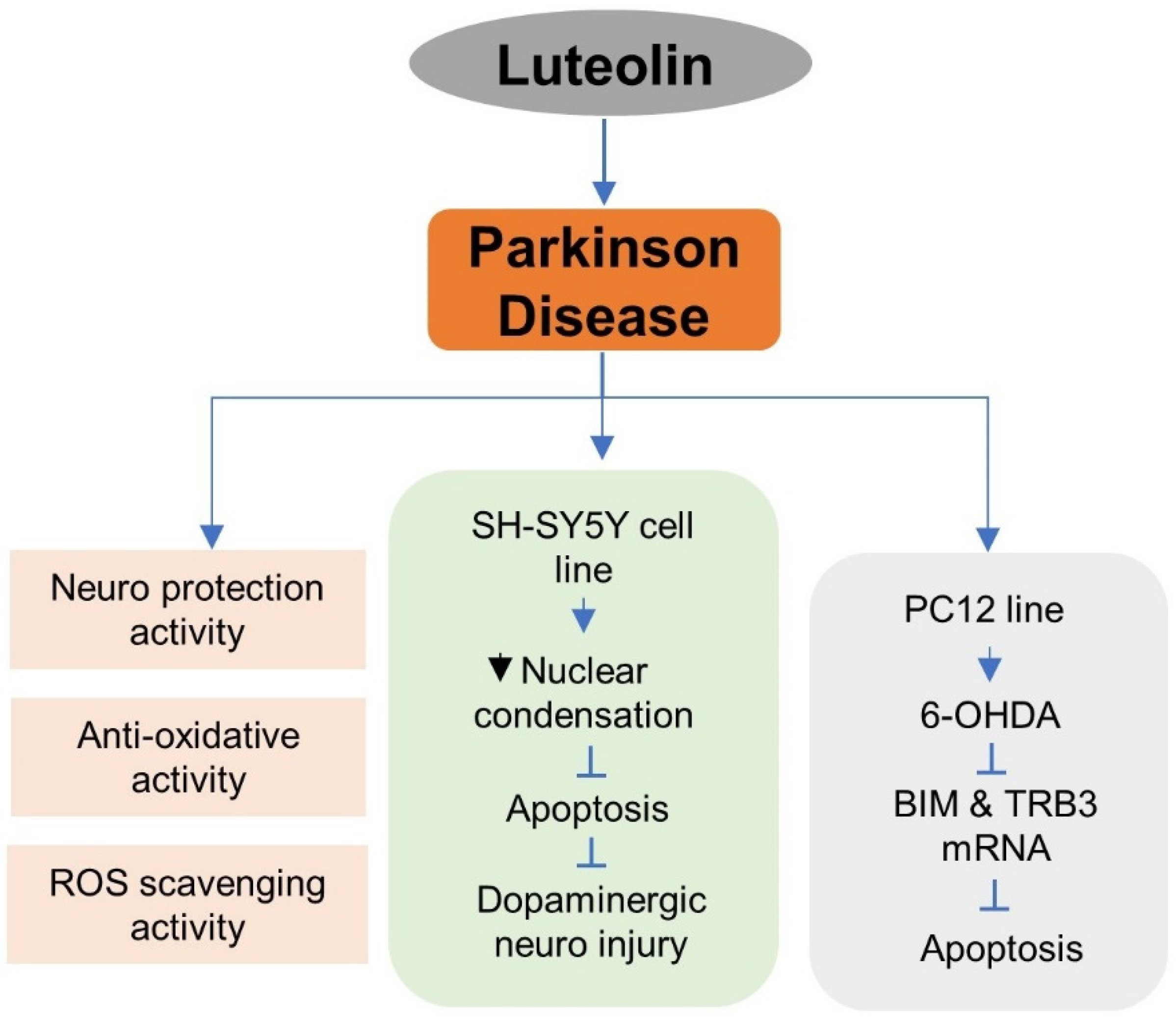
5.6. Luteolin in Parkinson’s Disease (PD) Treatment
5.7. Luteolin in Cardiac Health
5.8. Luteolin in Obesity Treatment
6. Cytotoxic Studies
7. Clinical Trials
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Imran, A.; Orhan, I.E.; Rizwan, M.; Atif, M.; et al. Luteolin, a Flavonoid, as an Anticancer Agent: A Review. Biomed. Pharmacother. 2019, 112, 108612. [Google Scholar] [CrossRef] [PubMed]
- Bravo, L. Polyphenols: Chemistry, Dietary Sources, Metabolism, and Nutritional Significance. Nutr. Rev. 2009, 56, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.F.; Braidy, N.; Gortzi, O.; Sobarzo-Sanchez, E.; Daglia, M.; Skalicka-Woźniak, K.; Nabavi, S.M. Luteolin as an Anti-Inflammatory and Neuroprotective Agent: A Brief Review. Brain Res. Bull. 2015, 119, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B.; Williams, C.A. Advances in Flavonoid Research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Hartmann, T. Diversity and Variability of Plant Secondary Metabolism: A Mechanistic View. Entomol. Exp. Appl. 1996, 80, 177–188. [Google Scholar] [CrossRef]
- Ross, J.A.; Kasum, C.M. Dietary Flavonoids: Bioavailability, Metabolic Effects, and Safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef]
- Ou, H.-C.; Pandey, S.; Hung, M.-Y.; Huang, S.-H.; Hsu, P.-T.; Day, C.-H.; Pai, P.; Viswanadha, V.P.; Kuo, W.-W.; Huang, C.-Y. Luteolin: A Natural Flavonoid Enhances the Survival of HUVECs against Oxidative Stress by Modulating AMPK/PKC Pathway. Am. J. Chin. Med. 2019, 47, 541–557. [Google Scholar] [CrossRef]
- Cook, M.T. Mechanism of Metastasis Suppression by Luteolin in Breast Cancer. Breast Cancer Targets Ther. 2018, 10, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Lazaro, M. Distribution and Biological Activities of the Flavonoid Luteolin. Mini-Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Giannasi, D.E.; Niklas, K.J. Flavonoid and Other Chemical Constituents of Fossil Miocene Celtis and Ulmus (Succor Creek Flora). Science 1977, 197, 765–767. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid Biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, K.M.; Weaver, L.M. The Shikimate Pathway. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 473–503. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, J.-L.; Austin, M.B.; Stewart, C.; Noel, J.P. Structure and Function of Enzymes Involved in the Biosynthesis of Phenylpropanoids. Plant Physiol. Biochem. 2008, 46, 356–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noel, J.P.; Ferrer, J.-L.; Jez, J.M.; Bowman, M.E.; Dixon, R.A. Structure of Chalcone Synthase and the Molecular Basis of Plant Polyketide Biosynthesis. Nat. Struct. Biol. 1999, 6, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Noel, J.P.; Jez, J.M.; Bowman, M.E.; Dixon, R.A. Structure and Mechanism of the Evolutionarily Unique Plant Enzyme Chalcone Isomerase. Nat. Struct. Biol. 2000, 7, 786–791. [Google Scholar] [CrossRef] [PubMed]
- CROFT, K.D. The Chemistry and Biological Effects of Flavonoids and Phenolic Acidsa. Ann. N. Y. Acad. Sci. 1998, 854, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Martens, S.; Mithöfer, A. Flavones and Flavone Synthases. Phytochemistry 2005, 66, 2399–2407. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.M.; Šamec, D.; Tomczyk, M.; Milella, L.; Russo, D.; Habtemariam, S.; Suntar, I.; Rastrelli, L.; Daglia, M.; Xiao, J.; et al. Flavonoid Biosynthetic Pathways in Plants: Versatile Targets for Metabolic Engineering. Biotechnol. Adv. 2020, 38, 107316. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Song, Y.; Ge, L.; Su, J.; Wen, Y.; Long, Y. Measurement and Correlation of the Solubilities of Luteolin and Rutin in Five Imidazole-Based Ionic Liquids. Fluid Phase Equilibria 2013, 344, 27–31. [Google Scholar] [CrossRef]
- Shakeel, F.; Haq, N.; Alshehri, S.; Ibrahim, M.A.; Elzayat, E.M.; Altamimi, M.A.; Mohsin, K.; Alanazi, F.K.; Alsarra, I.A. Solubility, Thermodynamic Properties and Solute-Solvent Molecular Interactions of Luteolin in Various Pure Solvents. J. Mol. Liq. 2018, 255, 43–50. [Google Scholar] [CrossRef]
- Wang, Z.; Zeng, M.; Wang, Z.; Qin, F.; Chen, J.; He, Z. Dietary Luteolin: A Narrative Review Focusing on Its Pharmacokinetic Properties and Effects on Glycolipid Metabolism. J. Agric. Food Chem. 2021, 69, 1441–1454. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free Radicals and Antioxidants in Normal Physiological Functions and Human Disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C. Flavonoid Antioxidants. Curr. Med. Chem. 2001, 8, 797–807. [Google Scholar] [CrossRef]
- Pietta, P.-G. Flavonoids as Antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.-W.; Jung, H.A.; Kang, S.S.; Choi, J.S. Antioxidant Constituents and a New Triterpenoid Glycoside From Flos Lonicerae. Arch. Pharmacal Res. 2007, 30, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-J.; Huang, C.-L.; Lian, T.-W.; Kou, M.-C.; Wang, L. Antioxidant Activity Of Glossogyne Tenuifolia. J. Agric. Food Chem. 2005, 53, 6305–6312. [Google Scholar] [CrossRef]
- Cai, Q.; Rahn, R.O.; Zhang, R. Dietary Flavonoids, Quercetin, Luteolin and Genistein, Reduce Oxidative DNA Damage and Lipid Peroxidation and Quench Free Radicals. Cancer Lett. 1997, 119, 99–107. [Google Scholar] [CrossRef]
- Horváthová, K.; Chalupa, I.; Šebová, L.; Tóthová, D.; Vachálková, A. Protective Effect of Quercetin and Luteolin in Human Melanoma HMB-2 Cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2005, 565, 105–112. [Google Scholar] [CrossRef]
- Cheng, I.F.; Breen, K. On the Ability of Four Flavonoids, Baicilein, Luteolin, Naringenin, and Quercetin, to Suppress the Fenton Reaction of the Iron-ATP Complex. BioMetals 2000, 13, 77–83. [Google Scholar] [CrossRef]
- Choi, J.S.; Islam, M.N.; Ali, M.Y.; Kim, Y.M.; Park, H.J.; Sohn, H.S.; Jung, H.A. The Effects of C-Glycosylation of Luteolin on Its Antioxidant, Anti-Alzheimer’s Disease, Anti-Diabetic, and Anti-Inflammatory Activities. Arch. Pharmacal Res. 2014, 37, 1354–1363. [Google Scholar] [CrossRef]
- Albarakati, A.J.A.; Baty, R.S.; Aljoudi, A.M.; Habotta, O.A.; Elmahallawy, E.K.; Kassab, R.B.; Abdel Moneim, A.E. Luteolin Protects against Lead Acetate-Induced Nephrotoxicity through Antioxidant, Anti-Inflammatory, Anti-Apoptotic, and Nrf2/HO-1 Signaling Pathways. Mol. Biol. Rep. 2020, 47, 2591–2603. [Google Scholar] [CrossRef]
- Boeing, T.; Souza, P.; Speca, S.; Somensi, L.B.; Mariano, L.N.B.; Cury, B.J.; Ferreira dos Anjos, M.; Quintão, N.L.M.; Dubuqoy, L.; Desreumax, P.; et al. Luteolin Prevents Irinotecan-Induced Intestinal Mucositis in Mice through Antioxidant and Anti-Inflammatory Properties. Br. J. Pharmacol. 2020, 177, 2393–2408. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Jun, C.; Lu, Y.; Jiangmei, S. Combination of Metformin and Luteolin Synergistically Protects Carbon Tetrachloride-Induced Hepatotoxicity: Mechanism Involves Antioxidant, Anti-Inflammatory, Antiapoptotic, and Nrf2/HO-1 Signaling Pathway. BioFactors 2019, 45, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.A.; Piao, M.J.; Hyun, Y.J.; Zhen, A.X.; Cho, S.J.; Ahn, M.J.; Yi, J.M.; Hyun, J.W. Luteolin Promotes Apoptotic Cell Death via Upregulation of Nrf2 Expression by DNA Demethylase and the Interaction of Nrf2 with P53 in Human Colon Cancer Cells. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; Wang, K.; Yuan, C.; Xing, R.; Ni, J.; Hu, G.; Chen, F.; Wang, X. Luteolin Protects Mice from Severe Acute Pancreatitis by Exerting HO-1-Mediated Anti-Inflammatory and Antioxidant Effects. Int. J. Mol. Med. 2016, 39, 113–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owumi, S.; Lewu, D.; Arunsi, U.; Oyelere, A. Luteolin Attenuates Doxorubicin-Induced Derangements of Liver and Kidney by Reducing Oxidative and Inflammatory Stress to Suppress apoptosis. Hum. Exp. Toxicol. 2021, 40, 1656–1672. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 6 October 2022).
- de Martel, C.; Georges, D.; Bray, F.; Ferlay, J.; Clifford, G.M. Global Burden of Cancer Attributable to Infections in 2018: A Worldwide Incidence Analysis. Lancet Glob. Health 2020, 8, e180–e190. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer Statistics for the Year 2020: An Overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef]
- Ganai, S.A.; Sheikh, F.A.; Baba, Z.A.; Mir, M.A.; Mantoo, M.A.; Yatoo, M.A. Anticancer Activity of the Plant Flavonoid Luteolin against Preclinical Models of Various Cancers and Insights on Different Signalling Mechanisms Modulated. Phytother. Res. 2021, 35, 3509–3532. [Google Scholar] [CrossRef]
- Lakhera, S.; Rana, M.; Devlal, K.; Celik, I.; Yadav, R. A Comprehensive Exploration of Pharmacological Properties, Bioactivities and Inhibitory Potentiality of Luteolin from Tridax Procumbens as Anticancer Drug by In-Silico Approach. Struct. Chem. 2022, 33, 703–719. [Google Scholar] [CrossRef] [PubMed]
- Kawaii, S.; Tomono, Y.; Katase, E.; Ogawa, K.; Yano, M. Antiproliferative Activity of Flavonoids on Several Cancer Cell Lines. Biosci. Biotechnol. Biochem. 1999, 63, 896–899. [Google Scholar] [CrossRef] [PubMed]
- Cherng, J.-M.; Shieh, D.-E.; Chiang, W.; Chang, M.-Y.; Chiang, L.-C. Chemopreventive Effects of Minor Dietary Constituents in Common Foods on Human Cancer Cells. Biosci. Biotechnol. Biochem. 2007, 71, 1500–1504. [Google Scholar] [CrossRef] [PubMed]
- Post, J.F.M.; Varma, R.S. Growth Inhibitory Effects of Bioflavonoids and Related Compounds on Human Leukemic CEM-C1 and CEM-C7 Cells. Cancer Lett. 1992, 67, 207–213. [Google Scholar] [CrossRef]
- Seelinger, G.; Merfort, I.; Wölfle, U.; Schempp, C. Anti-Carcinogenic Effects of the Flavonoid Luteolin. Molecules 2008, 13, 2628–2651. [Google Scholar] [CrossRef] [Green Version]
- Ko, W.G.; Kang, T.H.; Lee, S.J.; Kim, Y.C.; Lee, B.H. Effects of Luteolin on the Inhibition of Proliferation and Induction of Apoptosis in Human Myeloid Leukaemia Cells. Phytother. Res. 2002, 16, 295–298. [Google Scholar] [CrossRef]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.-M. Luteolin, a Flavonoid with Potential for Cancer Prevention and Therapy. Curr. Cancer Drug Targets 2008, 8, 634–646. [Google Scholar] [CrossRef]
- Gu, J.; Cheng, X.; Luo, X.; Yang, X.; Pang, Y.; Zhang, X.; Zhang, Y.; Liu, Y. Luteolin Ameliorates Cognitive Impairments by Suppressing the Expression of Inflammatory Cytokines and Enhancing Synapse-Associated Proteins GAP-43 and SYN Levels in Streptozotocin-Induced Diabetic Rats. Neurochem. Res. 2018, 43, 1905–1913. [Google Scholar] [CrossRef]
- Jang, C.H.; Moon, N.; Oh, J.; Kim, J.-S. Luteolin Shifts Oxaliplatin-Induced Cell Cycle Arrest at G0/G1 to Apoptosis in HCT116 Human Colorectal Carcinoma Cells. Nutrients 2019, 11, 770. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, S.; Tabatabaei, M.J.; Jafari, M.; Shemirani, F.; Tavakoli, S.; Mofasseri, M.; Tofighi, Z. Cuminum cyminum Fruits as Source of Luteolin-7-O-Glucoside, Potent Cytotoxic Flavonoid against Breast Cancer Cell Lines. Nat. Prod. Res. 2018, 34, 1602–1606. [Google Scholar] [CrossRef]
- Hong, J.; Fristiohady, A.; Nguyen, C.H.; Milovanovic, D.; Huttary, N.; Krieger, S.; Hong, J.; Geleff, S.; Birner, P.; Jäger, W.; et al. Apigenin and Luteolin Attenuate the Breaching of MDA-MB231 Breast Cancer Spheroids through the Lymph Endothelial Barrier in Vitro. Front. Pharmacol. 2018, 9, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavsan, Z.; Kayali, H.A. Flavonoids Showed Anticancer Effects on the Ovarian Cancer Cells: Involvement of Reactive Oxygen Species, Apoptosis, Cell Cycle and Invasion. Biomed. Pharmacother. 2019, 116, 109004. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-Y.; Tien, H.-J.; Chen, S.-F.; Horng, C.-T.; Tang, H.-L.; Jung, H.-L.; Wu, M.-J.; Yen, J.-H. Response of Myeloid Leukemia Cells to Luteolin Is Modulated by Differentially Expressed Pituitary Tumor-Transforming Gene 1 (PTTG1) Oncoprotein. Int. J. Mol. Sci. 2018, 19, 1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, Y.E.; de Lima, C.A.; Rosa, M.N.; Viviane, S.; Reis, R.M.; Priolli, D.G.; Carvalho, P.O.; do Nascimento, J.R.; da Rocha, C.Q.; Longato, G.B. Investigation of U-251 Cell Death Triggered by Flavonoid Luteolin: Towards a Better Understanding on Its Anticancer Property against Glioblastomas. Nat. Prod. Res. 2020, 35, 4807–4813. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, D.; Jung, K.-H.; Zhang, H.; Nannapaneni, S.; Wang, X.; Amin, A.R.M.R.; Chen, Z.; Chen, Z.G.; Shin, D.M. Luteolin Nanoparticle in Chemoprevention: In Vitro and In Vivo Anticancer Activity. Cancer Prev. Res. 2014, 7, 65–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.-W.; Lu, Z.-Y.; Pan, Q.; Chen, T.-T.; Feng, X.-J.; Wang, S.-M.; Pan, Y.-C.; Zhu, M.-H.; Zhang, S.-H. MicroRNA-6809-5p Mediates Luteolin-Induced Anticancer Effects against Hepatoma by Targeting Flotillin 1. Phytomedicine 2019, 57, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Palko-Labuz, A.; Sroda-Pomianek, K.; Uryga, A.; Kostrzewa-Suslow, E.; Michalak, K. Anticancer Activity of Baicalein and Luteolin Studied in Colorectal Adenocarcinoma LoVo Cells and in Drug-Resistant LoVo/Dx Cells. Biomed. Pharmacother. 2017, 88, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.; Chen, P.; Lo, Y.; Lin, C.; Chuang, Y.; Hsieh, M.; Chen, M. Luteolin-7-O-Glucoside Inhibits Cell Proliferation and Modulates apoptosis through the AKT Signaling Pathway in Human Nasopharyngeal Carcinoma. Environ. Toxicol. 2021, 36, 2013–2024. [Google Scholar] [CrossRef]
- Poppy, H.; Harahap, U.; Sitorus, P.; Satria, D. The Anticancer Activities of Vernonia Amygdalina Delile. Leaves on 4T1 Breast Cancer Cells through Phosphoinositide 3-Kinase (PI3K) Pathway. Heliyon 2020, 6, e04449. [Google Scholar] [CrossRef]
- Huang, L.; Jin, K.; Lan, H. Luteolin Inhibits Cell Cycle Progression and Induces Apoptosis of Breast Cancer Cells through Downregulation of Human Telomerase Reverse Transcriptase. Oncol. Lett. 2019, 17, 3842–3850. [Google Scholar] [CrossRef]
- Wu, H.-T.; Liu, Y.-E.; Hsu, K.-W.; Wang, Y.-F.; Chan, Y.-C.; Chen, Y.; Chen, D.-R. MLL3 Induced by Luteolin Causes Apoptosis in Tamoxifen-Resistant Breast Cancer Cells through H3K4 Monomethylation and Suppression of the PI3K/AKT/MTOR Pathway. Am. J. Chin. Med. 2020, 48, 1221–1241. [Google Scholar] [CrossRef]
- Lee, Y.; Kwon, Y.H. Regulation of Apoptosis and Autophagy by Luteolin in Human Hepatocellular Cancer Hep3B Cells. Biochem. Biophys. Res. Commun. 2019, 517, 617–622. [Google Scholar] [CrossRef]
- You, Y.; Wang, R.; Shao, N.; Zhi, F.; Yang, Y.P. Luteolin Suppresses Tumor Proliferation through Inducing Apoptosis and Autophagy via MAPK Activation in Glioma. Onco Targets Ther. 2019, 12, 2383–2396. [Google Scholar] [CrossRef] [Green Version]
- Couture, R.; Mora, N.; Bittar, A.; Najih, M.; Touaibia, M.; Martin, L.J. Luteolin Modulates Gene Expression Related to Steroidogenesis, Apoptosis, and Stress Response in Rat LC540 Tumor Leydig Cells. Cell Biol. Toxicol. 2019, 36, 31–49. [Google Scholar] [CrossRef]
- Juszczak, A.M.; Czarnomysy, R.; Strawa, J.W.; Zovko Končić, M.; Bielawski, K.; Tomczyk, M. In Vitro Anticancer Potential of Jasione Montana and Its Main Components against Human Amelanotic Melanoma Cells. Int. J. Mol. Sci. 2021, 22, 3345. [Google Scholar] [CrossRef]
- Velmurugan, B.K.; Lin, J.-T.; Mahalakshmi, B.; Chuang, Y.-C.; Lin, C.-C.; Lo, Y.-S.; Hsieh, M.-J.; Chen, M.-K. Luteolin-7-o-Glucoside Inhibits Oral Cancer Cell Migration and Invasion by Regulating Matrix Metalloproteinase-2 Expression and Extracellular Signal-Regulated Kinase Pathway. Biomolecules 2020, 10, 502. [Google Scholar] [CrossRef]
- Lin, X.; Xu, Y.; Pan, X.; Xu, J.; Ding, Y.; Sun, X.; Song, X.; Ren, Y.; Shan, P.-F. Global, Regional, and National Burden and Trend of Diabetes in 195 Countries and Territories: An Analysis from 1990 to 2025. Sci. Rep. 2020, 10, 14790. [Google Scholar] [CrossRef]
- Lin, T.Y.; Lu, C.W.; Wang, S.J. Luteolin Protects the Hippocampus against Neuron Impairments Induced by Kainic Acid in Rats. Neurotoxicology 2016, 55, 48–57. [Google Scholar] [CrossRef]
- Zhou, X.-R.; Ru, X.-C.; Xiao, C.; Pan, J.; Lou, Y.-Y.; Tang, L.-H.; Yang, J.-T.; Qian, L.-B. Sestrin2 Is Involved in the Nrf2-Regulated Antioxidative Signaling Pathway in Luteolin-Induced Prevention of the Diabetic Rat Heart from Ischemia/Reperfusion Injury. Food Funct. 2021, 12, 3562–3571. [Google Scholar] [CrossRef]
- Liu, Y.; Tian, X.; Gou, L.; Sun, L.; Ling, X.; Yin, X. Luteolin Attenuates Diabetes-Associated Cognitive Decline in Rats. Brain Res. Bull. 2013, 94, 23–29. [Google Scholar] [CrossRef]
- Davella, R.; Mamidala, E. Luteolin: A Potential Multiple Targeted Drug Effectively Inhibits Diabetes Mellitus Protein Targets. J. Pharm. Res. Int. 2021, 33, 161–171. [Google Scholar] [CrossRef]
- Kim, D.-K.; Nepali, S.; Son, J.-S.; Poudel, B.; Lee, J.-H.; Lee, Y.-M. Luteolin Is a Bioflavonoid That Attenuates Adipocyte-Derived Inflammatory Responses via Suppression of Nuclear Factor-ΚB/Mitogen-Activated Protein Kinases Pathway. Pharmacogn. Mag. 2015, 11, 627. [Google Scholar] [CrossRef] [Green Version]
- Wagle, A.; Seong, S.H.; Shrestha, S.; Jung, H.A.; Choi, J.S. Korean Thistle (Cirsium Japonicum Var. Maackii (Maxim.) Matsum.): A Potential Dietary Supplement against Diabetes and Alzheimer’s Disease. Molecules 2019, 24, 649. [Google Scholar] [CrossRef] [Green Version]
- Djeujo, F.M.; Ragazzi, E.; Urettini, M.; Sauro, B.; Cichero, E.; Tonelli, M.; Froldi, G. Magnolol and Luteolin Inhibition of α-Glucosidase Activity: Kinetics and Type of Interaction Detected by in Vitro and in Silico Studies. Pharmaceuticals 2022, 15, 205. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Johnson, M.H.; Lila, M.A.; Yousef, G.; Gonzalez, E. Berry and Citrus Phenolic Compounds Inhibit Dipeptidyl Peptidase IV: Implications in Diabetes Management. Evid. Based Complement. Altern. Med. 2013, 2013, 479505. [Google Scholar] [CrossRef] [Green Version]
- Aziz, N.; Kim, M.-Y.; Cho, J.Y. Anti-Inflammatory Effects of Luteolin: A Review of in Vitro, in Vivo, and in Silico Studies. J. Ethnopharmacol. 2018, 225, 342–358. [Google Scholar] [CrossRef]
- Ding, X.; Zheng, L.; Yang, B.; Wang, X.; Ying, Y. Luteolin Attenuates Atherosclerosis via Modulating Signal Transducer and Activator of Transcription 3-Mediated Inflammatory Response. Drug Des. Dev. Ther. 2019, 13, 3899–3911. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhang, D.; Xie, K.; Wang, C.; Xu, F. Luteolin Activates Tregs to Promote IL-10 Expression and Alleviating Caspase-11-Dependent Pyroptosis in Sepsis-Induced Lung Injury. Int. Immunopharmacol. 2021, 99, 107914. [Google Scholar] [CrossRef]
- Cho, Y.-C.; Park, J.; Cho, S. Anti-Inflammatory and Anti-Oxidative Effects of Luteolin-7-O-Glucuronide in LPS-Stimulated Murine Macrophages through TAK1 Inhibition and Nrf2 Activation. Int. J. Mol. Sci. 2020, 21, 2007. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, M.; Yamashita, S.; Hashimoto, T. Enhancement of Anti-Inflammatory and Anti-Allergic Activities with Combination of Luteolin and Quercetin in in Vitro Co-Culture System. Food Sci. Technol. Res. 2017, 23, 811–818. [Google Scholar] [CrossRef]
- Stefano, D.; Caporali, S.; Daniele, D.; Rovella, V.; Cardillo, C.; Schinzari, F.; Minieri, M.; Pieri, M.; Candi, E.; Bernardini, S.; et al. Anti-Inflammatory and Proliferative Properties of Luteolin-7-O-Glucoside. Int. J. Mol. Sci. 2021, 22, 1321. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.-L.; Yang, J.-W.; Dou, H.-Y.; Li, G.-Q.; Li, X.-Y.; Shen, L.; Ji, H.-F. Anti-Inflammatory Effect of Luteolin Is Related to the Changes in the Gut Microbiota and Contributes to Preventing the Progression from Simple Steatosis to Nonalcoholic Steatohepatitis. Bioorganic Chem. 2021, 112, 104966. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Rana, M. Anti-Inflammatory Activity and Isolation of Luteolin from Plagiochasma Appendiculatum Methanol Extract. Asian Pac. J. Health Sci. 2022, 9, 76–79. [Google Scholar] [CrossRef]
- Ma, Q.; Jiang, J.-G.; Zhang, X.-M.; Zhu, W. Identification of Luteolin 7-O-β-D-Glucuronide from Cirsium Japonicum and Its Anti-Inflammatory Mechanism. J. Funct. Foods 2018, 46, 521–528. [Google Scholar] [CrossRef]
- Xie, T.; Yuan, J.; Mei, L.; Li, P.; Pan, R. Luteolin Suppresses TNF-α-Induced Inflammatory Injury and Senescence of Nucleus Pulposus Cells via the Sirt6/NF-ΚB Pathway. Exp. Ther. Med. 2022, 24, 469. [Google Scholar] [CrossRef]
- Kim, J.-H.; Park, T.-J.; Park, J.-S.; Kim, M.-S.; Chi, W.-J.; Kim, S.-Y. Luteolin-3′-O-Phosphate Inhibits Lipopolysaccharide-Induced Inflammatory Responses by Regulating NF-ΚB/MAPK Cascade Signaling in RAW 264.7 Cells. Molecules 2021, 26, 7393. [Google Scholar] [CrossRef]
- Park, C.M.; Song, Y.-S. Luteolin and Luteolin-7-O-Glucoside Protect against Acute Liver Injury through Regulation of Inflammatory Mediators and Antioxidative Enzymes in GalN/LPS-Induced Hepatitic ICR Mice. Nutr. Res. Pract. 2019, 13, 473. [Google Scholar] [CrossRef]
- Zhang, M.; He, L.; Liu, J.; Zhou, L. Luteolin Attenuates Diabetic Nephropathy through Suppressing Inflammatory Response and Oxidative Stress by Inhibiting STAT3 Pathway. Exp. Clin. Endocrinol. Diabetes 2020, 129, 729–739. [Google Scholar] [CrossRef]
- Uwishema, O.; Mahmoud, A.; Sun, J.; Correia, I.F.S.; Bejjani, N.; Alwan, M.; Nicholas, A.; Oluyemisi, A.; Dost, B. Is Alzheimer’s Disease an Infectious Neurological Disease? A Review of the Literature. Brain Behav. 2022, 12, e2728. [Google Scholar] [CrossRef]
- Ali, F.; Jyoti, S.; Naz, F.; Ashafaq, M.; Shahid, M.; Siddique, Y.H. Therapeutic Potential of Luteolin in Transgenic Drosophila Model of Alzheimer’s Disease. Neurosci. Lett. 2019, 692, 90–99. [Google Scholar] [CrossRef]
- Kou, J.; Shi, J.; He, Y.; Hao, J.; Zhang, H.; Luo, D.; Song, J.; Yan, Y.; Xie, X.; Du, G.; et al. Luteolin Alleviates Cognitive Impairment in Alzheimer’s Disease Mouse Model via Inhibiting Endoplasmic Reticulum Stress-Dependent Neuroinflammation. Acta Pharmacol. 2021, 43, 840–849. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-R.; Pei, S.-Y.; Fan, D.-X.; Liu, Y.-H.; Pan, X.-F.; Song, F.-X.; Deng, S.-H.; Qiu, H.-B.; Zhang, N. Luteolin Protects Pheochromocytoma (PC-12) Cells against Aβ25-35-Induced Cell Apoptosis through the ER/ERK/MAPK Signalling Pathway. Evid. Based Complement. Altern. Med. 2020, 2020, 2861978. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, D.S.; Kang, S.; Kim, H.J. The Combination of Luteolin and L-Theanine Improved Alzheimer Disease–like Symptoms by Potentiating Hippocampal Insulin Signaling and Decreasing Neuroinflammation and Norepinephrine Degradation in Amyloid-β–Infused Rats. Nutr. Res. 2018, 60, 116–131. [Google Scholar] [CrossRef]
- Zhao, G.; Yao-Yue, C.; Qin, G.-W.; Guo, L.-H. Luteolin from Purple Perilla Mitigates ROS Insult Particularly in Primary Neurons. Neurobiol. Aging 2012, 33, 176–186. [Google Scholar] [CrossRef]
- Qin, L.; Chen, Z.; Yang, L.; Shi, H.; Wu, H.; Zhang, B.; Zhang, W.; Xu, Q.; Huang, F.; Wu, X. Luteolin-7-O-Glucoside Protects Dopaminergic Neurons by Activating Estrogen-Receptor-Mediated Signaling Pathway in MPTP-Induced Mice. Toxicology 2019, 426, 152256. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.-W.; Yen, J.-H.; Shen, Y.-T.; Wu, K.-Y.; Wu, M.-J. Luteolin Modulates 6-Hydroxydopamine-Induced Transcriptional Changes of Stress Response Pathways in PC12 Cells. PLoS ONE 2014, 9, e97880. [Google Scholar] [CrossRef]
- Guo, D.-J.; Li, F.; Yu, P.H.-F.; Chan, S.-W. Neuroprotective Effects of Luteolin against Apoptosis Induced by 6-Hydroxydopamine on Rat Pheochromocytoma PC12 Cells. Pharm. Biol. 2012, 51, 190–196. [Google Scholar] [CrossRef]
- Reudhabibadh, R.; Binlateh, T.; Chonpathompikunlert, P.; Nonpanya, N.; Prommeenate, P.; Chanvorachote, P.; Hutamekalin, P. Suppressing Cdk5 Activity by Luteolin Inhibits MPP+-Induced Apoptotic of Neuroblastoma through Erk/Drp1 and Fak/Akt/GSK3β Pathways. Molecules 2021, 26, 1307. [Google Scholar] [CrossRef]
- Liu, D.; Luo, H.; Qiao, C. SHP-1/STAT3 Interaction Is Related to Luteolin-Induced Myocardial Ischemia Protection. Inflammation 2021, 45, 88–99. [Google Scholar] [CrossRef]
- Zhao, L.; Zhou, Z.; Zhu, C.; Fu, Z.; Yu, D. Luteolin Alleviates Myocardial Ischemia Reperfusion Injury in Rats via Siti1/NLRP3/NF-ΚB Pathway. Int. Immunopharmacol. 2020, 85, 106680. [Google Scholar] [CrossRef]
- Du, Y.; Liu, P.; Xu, T.; Pan, D.; Zhu, H.; Zhai, N.; Zhang, Y.; Li, D. Luteolin Modulates SERCA2a Leading to Attenuation of Myocardial Ischemia/ Reperfusion Injury via Sumoylation at Lysine 585 in Mice. Cell. Physiol. Biochem. 2018, 45, 883–898. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Zhang, C.; Zhu, H.; Wang, S.; Zhou, Y.; Zhao, J.; Xia, Y.; Li, D. Luteolin Modulates SERCA2a via Sp1 Upregulation to Attenuate Myocardial Ischemia/Reperfusion Injury in Mice. Sci. Rep. 2020, 10, 15407. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Xu, T.; Wu, P.; Pan, D.; Chen, J.; Chen, J.; Zhang, B.; Zhu, H.; Li, D. Luteolin Improves Cardiac Dysfunction in Heart Failure Rats by Regulating Sarcoplasmic Reticulum Ca2+-ATPase 2a. Sci. Rep. 2017, 7, 41017. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.; Li, D.; Pan, H.; Chen, D.; Qi, L.; Zhang, R.; Sun, H. Luteolin Inhibits Apoptosis and Improves Cardiomyocyte Contractile Function through the PI3K/Akt Pathway in Simulated Ischemia/Reperfusion. Pharmacology 2011, 88, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Xu, T.; Luo, Y.; Zhang, Y.; Xuan, H.; Ma, Y.; Pan, D.; Li, D.; Zhu, H. Luteolin Enhances Sarcoplasmic Reticulum Ca2+-ATPase Activity through P38 MAPK Signaling Thus Improving Rat Cardiac Function after Ischemia/Reperfusion. Cell. Physiol. Biochem. 2017, 41, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 6 October 2022).
- Kwon, E.-Y.; Choi, M.-S. Luteolin Targets the Toll-like Receptor Signaling Pathway in Prevention of Hepatic and Adipocyte Fibrosis and Insulin Resistance in Diet-Induced Obese Mice. Nutrients 2018, 10, 1415. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, Q.-X.; Wang, X.; Zhang, L.; Qu, W.; Bao, B.; Liu, C.-A.; Liu, J. Dietary Luteolin Activates Browning and Thermogenesis in Mice through an AMPK/PGC1α Pathway-Mediated Mechanism. Int. J. Obes. 2016, 40, 1841–1849. [Google Scholar] [CrossRef]
- Park, H.-S.; Kim, S.-H.; Kim, Y.S.; Ryu, S.Y.; Hwang, J.-T.; Yang, H.J.; Kim, G.-H.; Kwon, D.Y.; Kim, M.-S. Luteolin Inhibits Adipogenic Differentiation by Regulating PPARγ Activation. BioFactors 2009, 35, 373–379. [Google Scholar] [CrossRef]
- Kwon, E.-Y.; Kim, S.; Choi, M.-S. Luteolin-Enriched Artichoke Leaf Extract Alleviates the Metabolic Syndrome in Mice with High-Fat Diet-Induced Obesity. Nutrients 2018, 10, 979. [Google Scholar] [CrossRef] [Green Version]
- Ando, C.; Takahashi, N.; Hirai, S.; Nishimura, K.; Lin, S.; Uemura, T.; Goto, T.; Yu, R.; Nakagami, J.; Murakami, S.; et al. Luteolin, a Food-Derived Flavonoid, Suppresses Adipocyte-Dependent Activation of Macrophages by Inhibiting JNK Activation. FEBS Lett. 2009, 583, 3649–3654. [Google Scholar] [CrossRef]
- Park, H.; Lee, K.; Kim, S.; Hong, M.J.; Jeong, N.; Kim, M. Luteolin Improves Hypercholesterolemia and Glucose Intolerance through LXRα-Dependent Pathway in Diet-Induced Obese Mice. J. Food Biochem. 2020, 44, e13358. [Google Scholar] [CrossRef] [PubMed]
- Caporali, S.; De Stefano, A.; Calabrese, C.; Giovannelli, A.; Pieri, M.; Savini, I.; Tesauro, M.; Bernardini, S.; Minieri, M.; Terrinoni, A. Anti-Inflammatory and Active Biological Properties of the Plant-Derived Bioactive Compounds Luteolin and Luteolin 7-Glucoside. Nutrients 2022, 14, 1155. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Cho, C.S.; Jun, H.O.; Ryu, N.H.; Kim, J.H.; Yu, Y.S.; Kim, J.S.; Kim, J.H. Anti-Angiogenic Effect of Luteolin on Retinal Neovascularization via Blockade of Reactive Oxygen Species Production. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7718–7726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; He, X.; Chen, S.; Le, Y.; Bryant, M.S.; Guo, L.; Witt, K.L.; Mei, N. The Genotoxicity Potential of Luteolin Is Enhanced by CYP1A1 and CYP1A2 in Human Lymphoblastoid TK6 Cells. Toxicol. Lett. 2021, 344, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Fateh, A.H.; Mohamed, Z.; Chik, Z.; Alsalahi, A.; Md Zin, S.R.; Alshawsh, M.A. Prenatal Developmental Toxicity Evaluation of Verbena Officinalis during Gestation Period in Female Sprague-Dawley Rats. Chem. Biol. Interact. 2019, 304, 28–42. [Google Scholar] [CrossRef]
- Cariddi, L.N.; Sabini, M.C.; Escobar, F.M.; Montironi, I.; Mañas, F.; Iglesias, D.; Comini, L.R.; Sabini, L.I.; Dalcero, A.M. Polyphenols as Possible Bioprotectors against Cytotoxicity and DNA Damage Induced by Ochratoxin A. Environ. Toxicol. Pharmacol. 2015, 39, 1008–1018. [Google Scholar] [CrossRef]
- D’Ascanio, L.; Vitelli, F.; Cingolani, C.; Maranzano, M.; Brenner, M.J.; Di Stadio, A. Randomized clinical trial “olfactory dysfunction after COVID-19: Olfactory rehabilitation therapy vs. intervention treatment with Palmitoylethanolamide and Luteolin”: Preliminary results. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 4156–4162. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Feng, Y.; Liu, Y.; Ye, X.; Ji, X.; Sun, L.; Gao, F.; Zhang, Q.; Li, Y.; Zhu, B.; et al. FuzhengJieduXiaoji Formulation Inhibits Hepatocellular Carcinoma Progression in Patients by Targeting the AKT/CyclinD1/P21/P27 Pathway. Phytomedicine 2021, 87, 153575. [Google Scholar] [CrossRef]
- Lunardelli, M.L.; Crupi, R.; Siracusa, R.; Cocuzza, G.; Cordaro, M.; Martini, E.; Impellizzeri, D.; Di Paola, R.; Cuzzocrea, S. Co-UltraPEALut: Role in Preclinical and Clinical Delirium Manifestations. CNS Neurol. Disord. Drug Targets 2019, 18, 530–554. [Google Scholar] [CrossRef]
- Castellino, G.; Nikolic, D.; Magán-Fernández, A.; Malfa, G.A.; Chianetta, R.; Patti, A.M.; Amato, A.; Montalto, G.; Toth, P.P.; Banach, M.; et al. Altilix® Supplement Containing Chlorogenic Acid and Luteolin Improved Hepatic and Cardiometabolic Parameters in Subjects with Metabolic Syndrome: A 6 Month Randomized, Double-Blind, Placebo-Controlled Study. Nutrients 2019, 11, 2580. [Google Scholar] [CrossRef]
- Gelabert-Rebato, M.; Wiebe, J.C.; Martin-Rincon, M.; Galvan-Alvarez, V.; Curtelin, D.; Perez-Valera, M.; Juan Habib, J.; Pérez-López, A.; Vega, T.; Morales-Alamo, D.; et al. Enhancement of Exercise Performance by 48 Hours, and 15-Day Supplementation with Mangiferin and Luteolin in Men. Nutrients 2019, 11, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taliou, A.; Zintzaras, E.; Lykouras, L.; Francis, K. An Open-Label Pilot Study of a Formulation Containing the Anti-Inflammatory Flavonoid Luteolin and Its Effects on Behavior in Children with Autism Spectrum Disorders. Clin. Ther. 2013, 35, 592–602. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muruganathan, N.; Dhanapal, A.R.; Baskar, V.; Muthuramalingam, P.; Selvaraj, D.; Aara, H.; Shiek Abdullah, M.Z.; Sivanesan, I. Recent Updates on Source, Biosynthesis, and Therapeutic Potential of Natural Flavonoid Luteolin: A Review. Metabolites 2022, 12, 1145. https://doi.org/10.3390/metabo12111145
Muruganathan N, Dhanapal AR, Baskar V, Muthuramalingam P, Selvaraj D, Aara H, Shiek Abdullah MZ, Sivanesan I. Recent Updates on Source, Biosynthesis, and Therapeutic Potential of Natural Flavonoid Luteolin: A Review. Metabolites. 2022; 12(11):1145. https://doi.org/10.3390/metabo12111145
Chicago/Turabian StyleMuruganathan, Nandakumar, Anand Raj Dhanapal, Venkidasamy Baskar, Pandiyan Muthuramalingam, Dhivya Selvaraj, Husne Aara, Mohamed Zubair Shiek Abdullah, and Iyyakkannu Sivanesan. 2022. "Recent Updates on Source, Biosynthesis, and Therapeutic Potential of Natural Flavonoid Luteolin: A Review" Metabolites 12, no. 11: 1145. https://doi.org/10.3390/metabo12111145