
Fetal Exposure to Maternal Smoking and Neonatal Metabolite Profiles
 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Design
2.2. Fetal Tobacco Exposure
2.3. Metabolite Measurements
2.4. Covariates
2.5. Statistical Analysis
3. Results
3.1. Characteristics of the Study Population
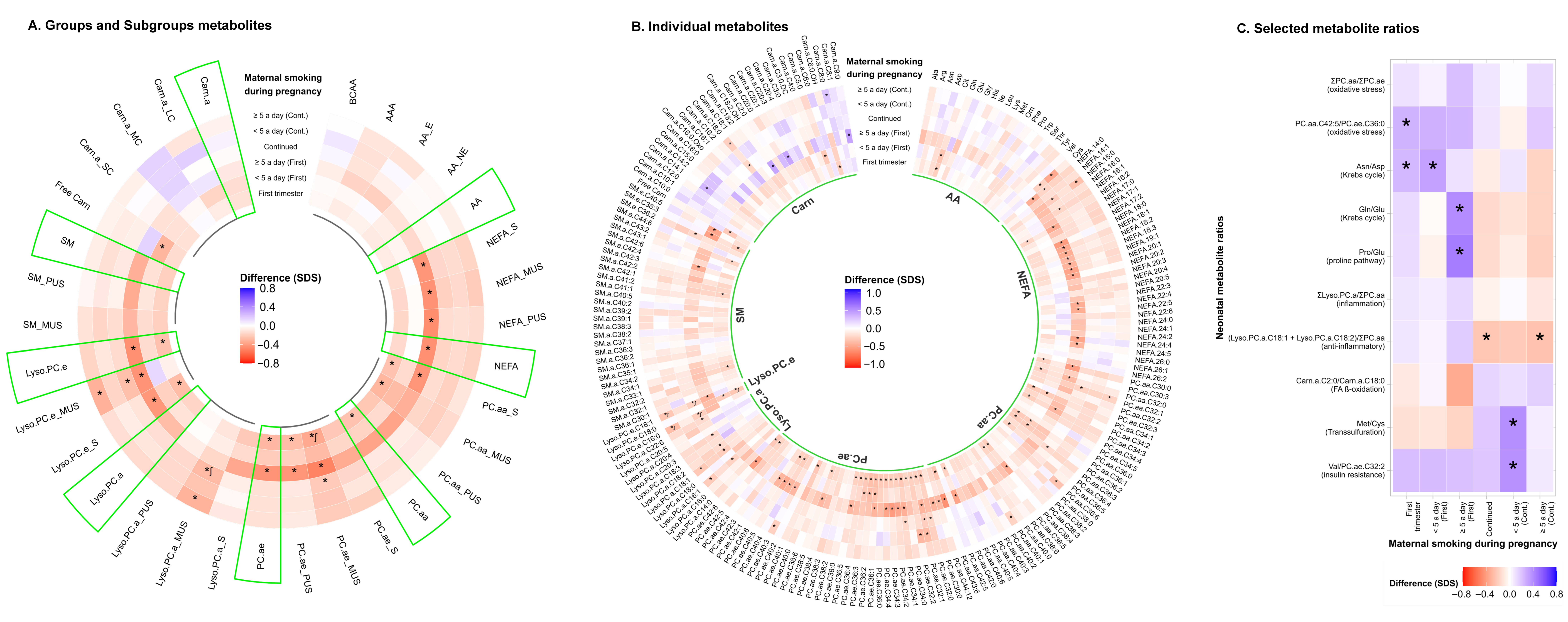
3.2. Fetal Tobacco Exposure and Neonatal Metabolite Profiles and Ratios
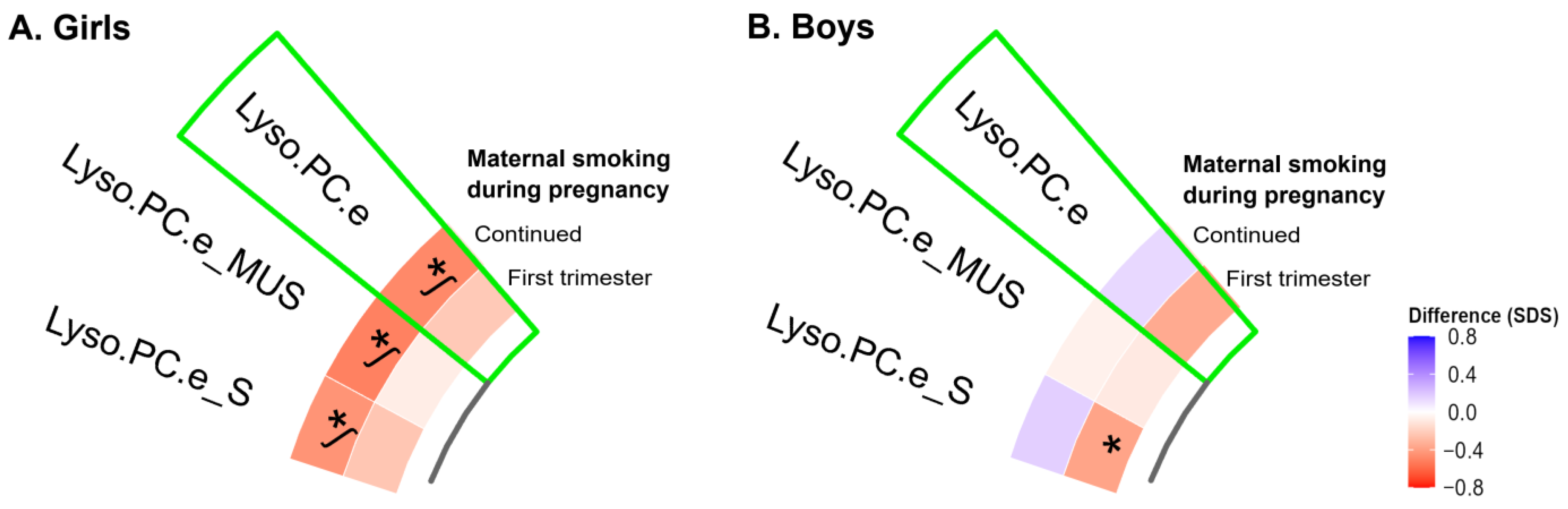
3.3. Sex-Specific Analyses
3.4. Sensitivity Analyses
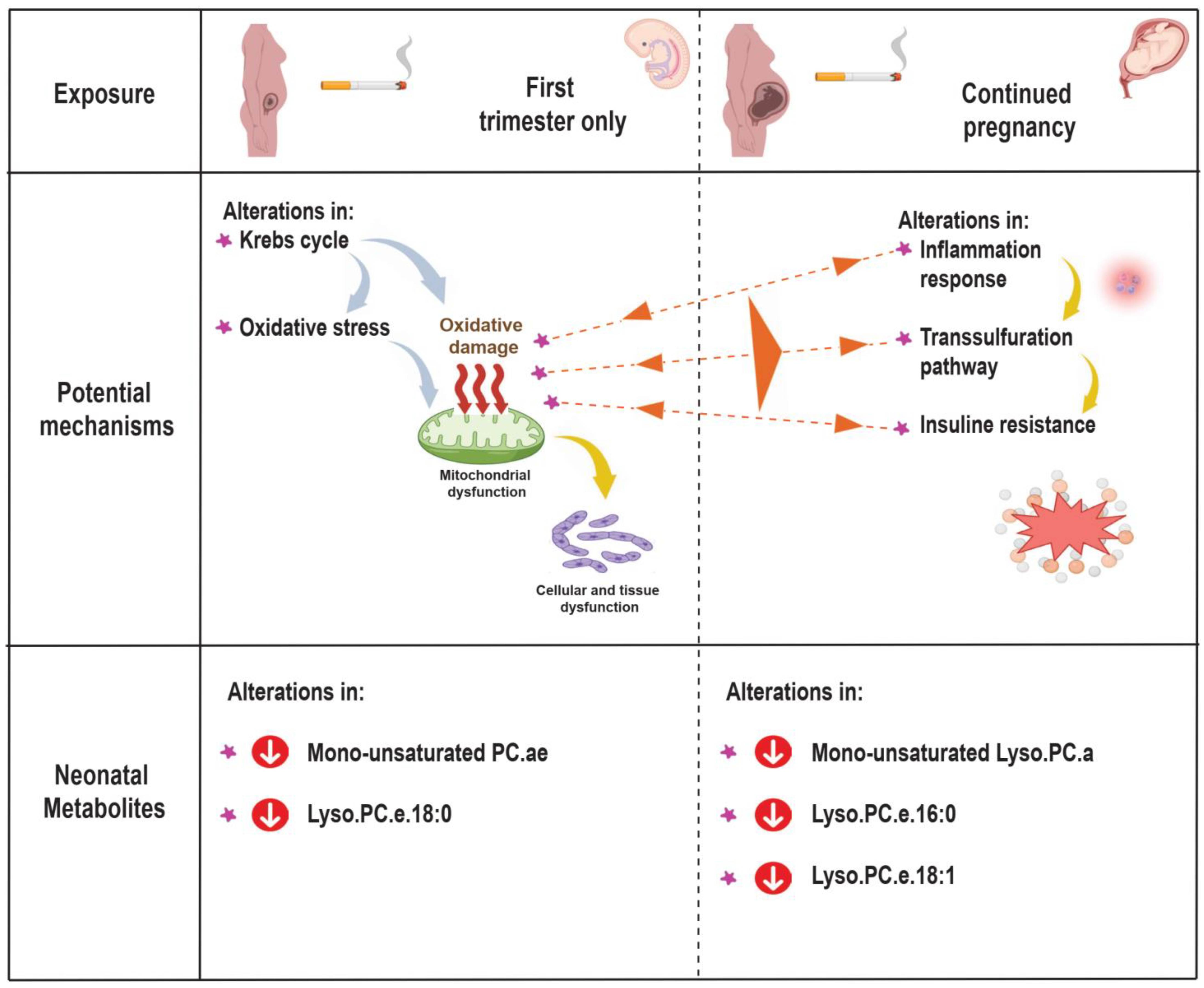
4. Discussion
Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SGA | small size for gestational age |
| PC.ae | acyl-alkyl-phosphatidylcholines |
| PC.aa | diacyl-phosphatidylcholines |
| Carn.a | acyl-carnitines |
| PC | phosphatidylcholines |
| SM | sphingomyelins |
| LC/MS | liquid chromatography/mass spectrometry |
| NEFA | non-esterified fatty acids |
| Lyso.PC.a | acyl-lysophosphatidylcholines |
| Lyso.PC.e | alkyl-lysophosphatidylcholines |
| Carn | carnitines |
| JCBN | Joint Commission on Biochemical Nomenclature |
| QC | quality control |
| CV | coefficient of variation |
| SD | standard deviation |
| BCAA | branched-chain amino acids |
| AAA | aromatic amino acids |
| SDS | standard deviations scores |
| BMI | body mass index |
| BSI | Brief Symptom Inventory |
| FDR | false discovery rate |
| IUGR | intrauterine growth restriction |
References
- Lange, S.; Probst, C.; Rehm, J.; Popova, S. National, regional, and global prevalence of smoking during pregnancy in the general population: A systematic review and meta-analysis. Lancet Glob. Health 2018, 6, e769–e776. [Google Scholar] [CrossRef] [Green Version]
- Jaddoe, V.W.V.; de Ridder, M.A.J.; van den Elzen, A.P.M.; Hofman, A.; Uiterwaal, C.; Witteman, J.C.M. Maternal smoking in pregnancy is associated with cholesterol development in the offspring: A 27-years follow-up study. Atherosclerosis 2008, 196, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Jaddoe, V.W.V.; Verburg, B.O.; de Ridder, M.A.J.; Hofman, A.; Mackenbach, J.P.; Moll, H.A.; Steegers, E.A.P.; Witteman, J.C.M. Maternal Smoking and Fetal Growth Characteristics in Different Periods of Pregnancy: The Generation R Study. Am. J. Epidemiol. 2007, 165, 1207–1215. [Google Scholar] [CrossRef] [Green Version]
- Philips, E.M.; Santos, S.; Trasande, L.; Aurrekoetxea, J.J.; Barros, H.; von Berg, A.; Bergström, A.; Bird, P.K.; Brescianini, S.; Chaoimh, N.C.; et al. Changes in parental smoking during pregnancy and risks of adverse birth outcomes and childhood overweight in Europe and North America: An individual participant data meta-analysis of 229,000 singleton births. PLoS Med. 2020, 17, e1003182. [Google Scholar] [CrossRef] [PubMed]
- Cajachagua-Torres, K.N.; El Marroun, H.; Reiss, I.K.M.; Jaddoe, V.W.V. Maternal preconception and pregnancy tobacco and cannabis use in relation to placental developmental markers: A population-based study. Reprod. Toxicol. 2022, 110, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Geelhoed, J.J.M.; El Marroun, H.; Verburg, B.O.; Van Osch-Gevers, L.; Hofman, A.; Huizink, A.C.; Moll, H.A.; Verhulst, F.C.; Helbing, W.A.; Steegers, E.A.P.; et al. Maternal smoking during pregnancy, fetal arterial resistance adaptations and cardiovascular function in childhood. BJOG Int. J. Obstet. Gynaecol. 2011, 118, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Harris, H.R.; Willett, W.C.; Michels, K.B. Parental smoking during pregnancy and risk of overweight and obesity in the daughter. Int. J. Obes. 2013, 37, 1356–1363. [Google Scholar] [CrossRef] [Green Version]
- de Jonge, L.L.; Harris, H.R.; Rich-Edwards, J.W.; Willett, W.C.; Forman, M.R.; Jaddoe, V.W.V.; Michels, K.B. Parental smoking in pregnancy and the risks of adult-onset hypertension. Hypertension 2013, 61, 494–500. [Google Scholar] [CrossRef] [Green Version]
- Jaddoe, V.W.; de Jonge, L.L.; van Dam, R.M.; Willett, W.C.; Harris, H.; Stampfer, M.J.; Hu, F.B.; Michels, K.B. Fetal exposure to parental smoking and the risk of type 2 diabetes in adult women. Diabetes Care 2014, 37, 2966–2973. [Google Scholar] [CrossRef] [Green Version]
- Cajachagua-Torres, K.N.; El Marroun, H.; Reiss, I.K.M.; Santos, S.; Jaddoe, V.W.V. Foetal tobacco and cannabis exposure, body fat and cardio-metabolic health in childhood. Pediatr. Obes. 2022, 17, e12863. [Google Scholar] [CrossRef]
- Wang-Sattler, R.; Yu, Y.; Mittelstrass, K.; Lattka, E.; Altmaier, E.; Gieger, C.; Ladwig, K.H.; Dahmen, N.; Weinberger, K.M.; Hao, P.; et al. Metabolic Profiling Reveals Distinct Variations Linked to Nicotine Consumption in Humans—First Results from the KORA Study. PLoS ONE 2008, 3, e3863. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Holzapfel, C.; Dong, X.; Bader, E.; Yu, Z.; Prehn, C.; Perstorfer, K.; Jaremek, M.; Roemisch-Margl, W.; Rathmann, W.; et al. Effects of smoking and smoking cessation on human serum metabolite profile: Results from the KORA cohort study. BMC Med. 2013, 11, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolle-Kampczyk, U.E.; Krumsiek, J.; Otto, W.; Röder, S.W.; Kohajda, T.; Borte, M.; Theis, F.; Lehmann, I.; von Bergen, M. Metabolomics reveals effects of maternal smoking on endogenous metabolites from lipid metabolism in cord blood of newborns. Metabolomics 2016, 12, 76. [Google Scholar] [CrossRef] [Green Version]
- Cheong, J.N.; Wlodek, M.E.; Moritz, K.M.; Cuffe, J.S.M. Programming of maternal and offspring disease: Impact of growth restriction, fetal sex and transmission across generations. J. Physiol. 2016, 594, 4727–4740. [Google Scholar] [CrossRef] [Green Version]
- Zarén, B.; Lindmark, G.; Bakketeig, L. Maternal smoking affects fetal growth more in the male fetus. Paediatr. Perinat. Epidemiol. 2000, 14, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Sandovici, I.; Fernandez-Twinn, D.S.; Hufnagel, A.; Constância, M.; Ozanne, S.E. Sex differences in the intergenerational inheritance of metabolic traits. Nat. Metab. 2022, 4, 507–523. [Google Scholar] [CrossRef]
- Clifton, V.L. Review: Sex and the Human Placenta: Mediating Differential Strategies of Fetal Growth and Survival. Placenta 2010, 31, S33–S39. [Google Scholar] [CrossRef]
- Manikkam, M.; Crespi, E.J.; Doop, D.D.; Herkimer, C.; Lee, J.S.; Yu, S.; Brown, M.B.; Foster, D.L.; Padmanabhan, V. Fetal Programming: Prenatal Testosterone Excess Leads to Fetal Growth Retardation and Postnatal Catch-Up Growth in Sheep. Endocrinology 2004, 145, 790–798. [Google Scholar] [CrossRef] [Green Version]
- Jaddoe, V.W.V.; van Duijn, C.M.; Franco, O.H.; van der Heijden, A.J.; van Iizendoorn, M.H.; de Jongste, J.C.; van der Lugt, A.; Mackenbach, J.P.; Moll, H.A.; Raat, H.; et al. The Generation R Study: Design and cohort update 2012. Eur. J. Epidemiol. 2012, 27, 739–756. [Google Scholar] [CrossRef] [Green Version]
- Jaddoe, V.W.V.; van Duijn, C.M.; van der Heijden, A.J.; Mackenbach, J.P.; Moll, H.A.; Steegers, E.A.P.; Tiemeier, H.; Uitterlinden, A.G.; Verhulst, F.C.; Hofman, A. The Generation R Study: Design and cohort update until the age of 4 years. Eur. J. Epidemiol. 2008, 23, 801. [Google Scholar] [CrossRef]
- Statistics Netherlands. Allochtonen in Nederland 2004; Statistics Netherlands: Voorbug, Heerlen, The Netherlands, 2004. [Google Scholar]
- Jaddoe, V.W.V.; Mackenbach, J.P.; Moll, H.A.; Steegers, E.A.; Tiemeier, H.; Verhulst, F.C.; Witteman, J.C.; Hofman, A. The Generation R Study: Design and cohort profile. Eur. J. Epidemiol. 2006, 21, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Voerman, E.; Jaddoe, V.W.V.; Uhl, O.; Shokry, E.; Horak, J.; Felix, J.F.; Koletzko, B.; Gaillard, R. A population-based resource for intergenerational metabolomics analyses in pregnant women and their children: The Generation R Study. Metabolomics 2020, 16, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellmuth, C.; Weber, M.; Koletzko, B.; Peissner, W. Nonesterified Fatty Acid Determination for Functional Lipidomics: Comprehensive Ultrahigh Performance Liquid Chromatography–Tandem Mass Spectrometry Quantitation, Qualification, and Parameter Prediction. Anal. Chem. 2012, 84, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- Harder, U.; Koletzko, B.; Peissner, W. Quantification of 22 plasma amino acids combining derivatization and ion-pair LC–MS/MS. J. Chromatogr. B 2011, 879, 495–504. [Google Scholar] [CrossRef]
- Hellmuth, C.; Uhl, O.; Standl, M.; Demmelmair, H.; Heinrich, J.; Koletzko, B.; Thiering, E. Cord Blood Metabolome Is Highly Associated with Birth Weight, but Less Predictive for Later Weight Development. Obes. Facts 2017, 10, 85–100. [Google Scholar] [CrossRef]
- Hellmuth, C.; Lindsay, K.L.; Uhl, O.; Buss, C.; Wadhwa, P.D.; Koletzko, B.; Entringer, S. Association of maternal prepregnancy BMI with metabolomic profile across gestation. Int. J. Obes. 2017, 41, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Blaauwendraad, S.M.; Voerman, E.; Trasande, L.; Kannan, K.; Santos, S.; Ruijter, G.J.G.; Sol, C.M.; Marchioro, L.; Shokry, E.; Koletzko, B.; et al. Associations of maternal bisphenol urine concentrations during pregnancy with neonatal metabolomic profiles. Metabolomics 2021, 17, 84. [Google Scholar] [CrossRef]
- IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN). Nomenclature and Symbolism for Amino Acids and Peptides. Eur. J. Biochem. 1984, 138, 9–37. [Google Scholar] [CrossRef]
- Uhl, O.; Fleddermann, M.; Hellmuth, C.; Demmelmair, H.; Koletzko, B. Phospholipid Species in Newborn and 4 Month Old Infants after Consumption of Different Formulas or Breast Milk. PLoS ONE 2016, 11, e0162040. [Google Scholar] [CrossRef] [Green Version]
- Shokry, E.; Marchioro, L.; Uhl, O.; Bermúdez, M.G.; García-Santos, J.A.; Segura, M.T.; Campoy, C.; Koletzko, B. Impact of maternal BMI and gestational diabetes mellitus on maternal and cord blood metabolome: Results from the PREOBE cohort study. Acta Diabetol. 2019, 56, 421–430. [Google Scholar] [CrossRef]
- Lindsay, K.L.; Hellmuth, C.; Uhl, O.; Buss, C.; Wadhwa, P.D.; Koletzko, B.; Entringer, S. Longitudinal Metabolomic Profiling of Amino Acids and Lipids across Healthy Pregnancy. PLoS ONE 2016, 10, e0145794. [Google Scholar] [CrossRef] [PubMed]
- Rauschert, S.; Mori, T.A.; Beilin, L.J.; Jacoby, P.; Uhl, O.; Koletzko, B.; Oddy, W.H.; Hellmuth, C. Early Life Factors, Obesity Risk, and the Metabolome of Young Adults. Obesity 2017, 25, 1549–1555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, R.; Wang, J.; Su, M.; Jia, E.; Chen, S.; Chen, T.; Ni, Y. Missing Value Imputation Approach for Mass Spectrometry-based Metabolomics Data. Sci. Rep. 2018, 8, 663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnos, S.; Wahl, S.; Haid, M.; Eekhoff, E.M.W.; Pool, R.; Floegel, A.; Deelen, J.; Much, D.; Prehn, C.; Breier, M.; et al. Metabolite ratios as potential biomarkers for type 2 diabetes: A DIRECT study. Diabetologia 2018, 61, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Hocher, B.; Adamski, J. Metabolomics for clinical use and research in chronic kidney disease. Nat. Rev. Nephrol. 2017, 13, 269–284. [Google Scholar] [CrossRef]
- Cappelletti, P.; Tallarita, E.; Rabattoni, V.; Campomenosi, P.; Sacchi, S.; Pollegioni, L. Proline oxidase controls proline, glutamate, and glutamine cellular concentrations in a U87 glioblastoma cell line. PLoS ONE 2018, 13, e0196283. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Sun, G.; Aitken, D.; Likhodii, S.; Liu, M.; Martin, G.; Furey, A.; Randell, E.; Rahman, P.; Jones, G.; et al. Lysophosphatidylcholines to phosphatidylcholines ratio predicts advanced knee osteoarthritis. Rheumatology 2016, 55, 1566–1574. [Google Scholar] [CrossRef] [Green Version]
- Belalcázar, A.D.; Ball, J.G.; Frost, L.M.; Valentovic, M.A.; Wilkinson, J. Transsulfuration Is a Significant Source of Sulfur for Glutathione Production in Human Mammary Epithelial Cells. ISRN Biochem. 2013, 2013, 637897. [Google Scholar] [CrossRef] [Green Version]
- Campesi, I.; Carru, C.; Zinellu, A.; Occhioni, S.; Sanna, M.; Palermo, M.; Tonolo, G.; Mercuro, G.; Franconi, F. Regular cigarette smoking influences the transsulfuration pathway, endothelial function, and inflammation biomarkers in a sex-gender specific manner in healthy young humans. Am. J. Transl. Res. 2013, 5, 497–509. [Google Scholar]
- Pickens, C.A.; Vazquez, A.I.; Jones, A.D.; Fenton, J.I. Obesity, adipokines, and C-peptide are associated with distinct plasma phospholipid profiles in adult males, an untargeted lipidomic approach. Sci. Rep. 2017, 7, 6335. [Google Scholar] [CrossRef] [Green Version]
- Kirchberg, F.F.; Brandt, S.; Moß, A.; Peissner, W.; Koenig, W.; Rothenbacher, D.; Brenner, H.; Koletzko, B.; Hellmuth, C.; Wabitsch, M. Metabolomics reveals an entanglement of fasting leptin concentrations with fatty acid oxidation and gluconeogenesis in healthy children. PLoS ONE 2017, 12, e0183185. [Google Scholar] [CrossRef]
- Yong-Ping, L.; Reichetzeder, C.; Prehn, C.; Yin, L.-H.; Chu, C.; Elitok, S.; Krämer, B.K.; Adamski, J.; Hocher, B. Impact of maternal smoking associated lyso-phosphatidylcholine 20:3 on offspring brain development. J. Steroid Biochem. Mol. Biol. 2020, 199, 105591. [Google Scholar] [CrossRef] [PubMed]
- Statistics Netherlands. Standaard Onderwijsindeling 2003; Statistics Netherlands: Voorbug, Heerlen, The Netherlands, 2004. [Google Scholar]
- Derogatis, L.R.; Melisaratos, N. The Brief Symptom Inventory: An introductory report. Psychol Med. 1983, 13, 595–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaddoe, V.W.V.; Bakker, R.; van Duijn, C.M.; van der Heijden, A.J.; Lindemans, J.; Mackenbach, J.P.; Moll, H.A.; Steegers, E.A.P.; Tiemeier, H.; Uitterlinden, A.G.; et al. The Generation R Study Biobank: A resource for epidemiological studies in children and their parents. Eur. J. Epidemiol. 2007, 22, 917–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niklasson, A.; Ericson, A.; Fryer, J.G.; Karlberg, J.; Lawrence, C.; Karlberg, P. An update of the Swedish reference standards for weight, length and head circumference at birth for given gestational age (1977–1981). Acta Paediatr. Scand. 1991, 80, 756–762. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- van Buuren, S.; Groothuis-Oudshoorn, K. Mice: Multivariate Imputation by Chained Equations in R. J. Stat. Softw. 2011, 45, 1–67. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Barker, D.J. Fetal origins of coronary heart disease. BMJ (Clin. Res. Ed.) 1995, 311, 171–174. [Google Scholar] [CrossRef]
- McMillen, I.C.; Robinson, J.S. Developmental Origins of the Metabolic Syndrome: Prediction, Plasticity, and Programming. Physiol. Rev. 2005, 85, 571–633. [Google Scholar] [CrossRef]
- Lu, Y.P.; Reichetzeder, C.; Prehn, C.; Yin, L.H.; Yun, C.; Zeng, S.; Chu, C.; Adamski, J.; Hocher, B. Cord Blood Lysophosphatidylcholine 16: 1 is Positively Associated with Birth Weight. Cell Physiol. Biochem. 2018, 45, 614–624. [Google Scholar] [CrossRef]
- Feng, J.-H.; Yan, Y.-E.; Liang, G.; Liu, Y.-S.; Li, X.-J.; Zhang, B.-J.; Chen, L.-B.; Yu, H.; He, X.-H.; Wang, H. Maternal and fetal metabonomic alterations in prenatal nicotine exposure-induced rat intrauterine growth retardation. Mol. Cell Endocrinol. 2014, 394, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Liang, G.; Yan, Y.E.; He, W.W.; Liu, Y.S.; Chen, L.B.; Magdalou, J.; Wang, H. Nicotine-induced over-exposure to maternal glucocorticoid and activated glucocorticoid metabolism causes hypothalamic–pituitary–adrenal axis-associated neuroendocrine metabolic alterations in fetal rats. Toxicol. Lett. 2012, 209, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Sastry, B.V.R.; Horst, M.A.; Naukam, R.J. Maternal tobacco smoking and changes in amino acid uptake by human placental villi: Induction of uptake systems, gammaglutamyltranspeptidase and membrane fluidity. Placenta 1989, 10, 345–358. [Google Scholar] [CrossRef]
- Lucendo-Villarin, B.; Filis, P.; Swortwood, M.J.; Huestis, M.A.; Meseguer-Ripolles, J.; Cameron, K.; Iredale, J.P.; O’Shaughnessy, P.J.; Fowler, P.A.; Hay, D.C. Modelling foetal exposure to maternal smoking using hepatoblasts from pluripotent stem cells. Arch. Toxicol. 2017, 91, 3633–3643. [Google Scholar] [CrossRef] [Green Version]
- Dessì, A.; Corona, L.; Pintus, R.; Fanos, V. Exposure to tobacco smoke and low birth weight: From epidemiology to metabolomics. Expert Rev. Proteom. 2018, 15, 647–656. [Google Scholar] [CrossRef]
- Fischer, S.T.; Lili, L.N.; Li, S.; Tran, V.T.; Stewart, K.B.; Schwartz, C.E.; Jones, D.P.; Sherman, S.L.; Fridovich-Keil, J.L. Low-level maternal exposure to nicotine associates with significant metabolic perturbations in second-trimester amniotic fluid. Environ. Int. 2017, 107, 227–234. [Google Scholar] [CrossRef]
- Tan, Y.; Barr, D.B.; Ryan, P.B.; Fedirko, V.; Sarnat, J.A.; Gaskins, A.J.; Chang, C.-J.; Tang, Z.; Marsit, C.J.; Corwin, E.J.; et al. High-resolution metabolomics of exposure to tobacco smoke during pregnancy and adverse birth outcomes in the Atlanta African American maternal-child cohort. Environ. Pollut. 2022, 292, 118361. [Google Scholar] [CrossRef]
- O’Shaughnessy, P.J.; Monteiro, A.; Bhattacharya, S.; Fowler, P.A. Maternal Smoking and Fetal Sex Significantly Affect Metabolic Enzyme Expression in the Human Fetal Liver. J. Clin. Endocrinol. Metab. 2011, 96, 2851–2860. [Google Scholar] [CrossRef] [Green Version]
- Bruin, J.E.; Petre, M.A.; Raha, S.; Morrison, K.M.; Gerstein, H.C.; Holloway, A.C. Fetal and Neonatal Nicotine Exposure in Wistar Rats Causes Progressive Pancreatic Mitochondrial Damage and Beta Cell Dysfunction. PLoS ONE 2008, 3, e3371. [Google Scholar] [CrossRef] [Green Version]
- Kooijman, M.N.; Kruithof, C.J.; van Duijn, C.M.; Duijts, L.; Franco, O.H.; van, I.M.H.; de Jongste, J.C.; Klaver, C.C.; van der Lugt, A.; Mackenbach, J.P.; et al. The Generation R Study: Design and cohort update 2017. Eur. J. Epidemiol. 2016, 31, 1243–1264. [Google Scholar] [CrossRef] [Green Version]
- Hellmuth, C.; Lindsay, K.L.; Uhl, O.; Buss, C.; Wadhwa, P.D.; Koletzko, B.; Entringer, S. Maternal Metabolomic Profile and Fetal Programming of Offspring Adiposity: Identification of Potentially Protective Lipid Metabolites. Mol. Nutr. Food Res. 2019, 63, 1700889. [Google Scholar] [CrossRef] [PubMed]
- Rauschert, S.; Uhl, O.; Koletzko, B.; Kirchberg, F.; Mori, T.A.; Huang, R.-C.; Beilin, L.J.; Hellmuth, C.; Oddy, W.H. Lipidomics Reveals Associations of Phospholipids with Obesity and Insulin Resistance in Young Adults. J. Clin. Endocrinol. Metab. 2016, 101, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Uhl, O.; Glaser, C.; Demmelmair, H.; Koletzko, B. Reversed phase LC/MS/MS method for targeted quantification of glycerophospholipid molecular species in plasma. J. Chromatogr. B 2011, 879, 3556–3564. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Maternal Smoking During | ||||
|---|---|---|---|---|
| Total N = 828 | Non-Smoking n = 631 | First Trimester Only n = 77 | Continued during Pregnancy n = 120 | |
| Maternal characteristic | ||||
| Age, years, mean (±SD) | 31.4 (4.1) | 31.7 (3.9) | 30.9 (4.1) | 30.2 (5.2) ** |
| Educational level | ||||
| None/Primary (%) | 2.3 | 1.3 | 5.2 * | 5.8 ** |
| Secondary (%) | 35.4 | 31.5 | 33.8 | 56.7 |
| Higher (%) | 62.3 | 67.2 | 61.0 | 37.5 |
| Pre-pregnancy body mass index, kg/m2, median (95% range) | 22.5 (18.4–33.7) | 22.6 (18.5–33.4) | 22.1 (18.4–30.3) | 22.3 (18.4–34.8) |
| Folate concentrations, nmol/L, median (95% range) | 19.5 (6.1–39.6) | 20.7 (6.3–40.5) | 18.4 (7.3–35.0) | 12.6 (5.7–33.3) ** |
| Psychopathology score, median (95% range) | 0.12 (0.00–0.79) | 0.10 (0–0.63) | 0.14 (0–0.71) | 0.19 (0–1.69) ** |
| Alcohol use | ||||
| Never drank in pregnancy (%) | 32.2 | 34.1 | 15.6 ** | 33.3 |
| First trimester only (%) | 15.7 | 14.1 | 29.9 | 15.0 |
| Continued drinking (%) | 52.1 | 51.8 | 54.5 | 51.7 |
| Paternal tobacco use, yes (%) | 41.6 | 32.3 | 58.4 ** | 75.8 ** |
| Environmental smoking | ||||
| No (%) | 66.5 | 70.0 | 62.3 | 38.3 ** |
| Occasionally (%) | 21.3 | 19.8 | 28.6 | 20.8 |
| Daily (%) | 12.2 | 8.9 | 9.1 | 29.2 |
| Birth characteristics | ||||
| Gestational age, weeks, median (95% range) | 40.3 (36.7–42.3) | 40.3 (37.0–42.3) | 40.3 (36.7–42.3) | 40.3 (36.6–42.4) |
| Premature birth (%) | 2.9 | 2.5 | 3.9 | 4.2 |
| Birth weight, grams, mean (±SD) | 3555.0 (506.8) | 3569.7 (488.8) | 3601.8 (573.4) | 3402.3 (532.8) ** |
| Small for gestational age (%) | 10.0 | 8.1 | 7.8 | 21.7 ** |
| Low birth weight (%) | 2.2 | 1.7 | 2.6 | 4.2 * |
| Female sex, yes (%) | 46.1 | 46.8 | 51.9 | 39.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cajachagua-Torres, K.N.; Blaauwendraad, S.M.; El Marroun, H.; Demmelmair, H.; Koletzko, B.; Gaillard, R.; Jaddoe, V.W.V. Fetal Exposure to Maternal Smoking and Neonatal Metabolite Profiles. Metabolites 2022, 12, 1101. https://doi.org/10.3390/metabo12111101
Cajachagua-Torres KN, Blaauwendraad SM, El Marroun H, Demmelmair H, Koletzko B, Gaillard R, Jaddoe VWV. Fetal Exposure to Maternal Smoking and Neonatal Metabolite Profiles. Metabolites. 2022; 12(11):1101. https://doi.org/10.3390/metabo12111101
Chicago/Turabian StyleCajachagua-Torres, Kim N., Sophia M. Blaauwendraad, Hanan El Marroun, Hans Demmelmair, Berthold Koletzko, Romy Gaillard, and Vincent W. V. Jaddoe. 2022. "Fetal Exposure to Maternal Smoking and Neonatal Metabolite Profiles" Metabolites 12, no. 11: 1101. https://doi.org/10.3390/metabo12111101