

Rapid Profiling of Metabolites Combined with Network Pharmacology to Explore the Potential Mechanism of Sanguisorba officinalis L. against Thrombocytopenia
 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Experimental Methods
2.1. Chemicals and Materials
2.2. Preparation of SO Extract
2.3. Animal Experiments
2.4. Sample Preparation
2.5. Chromatographic and MS Conditions
2.6. Metabolite Data Acquisition and Preprocessing
2.7. Network Pharmacology Analysis
3. Results and Discussion
3.1. Optimization of LC–MS Conditions
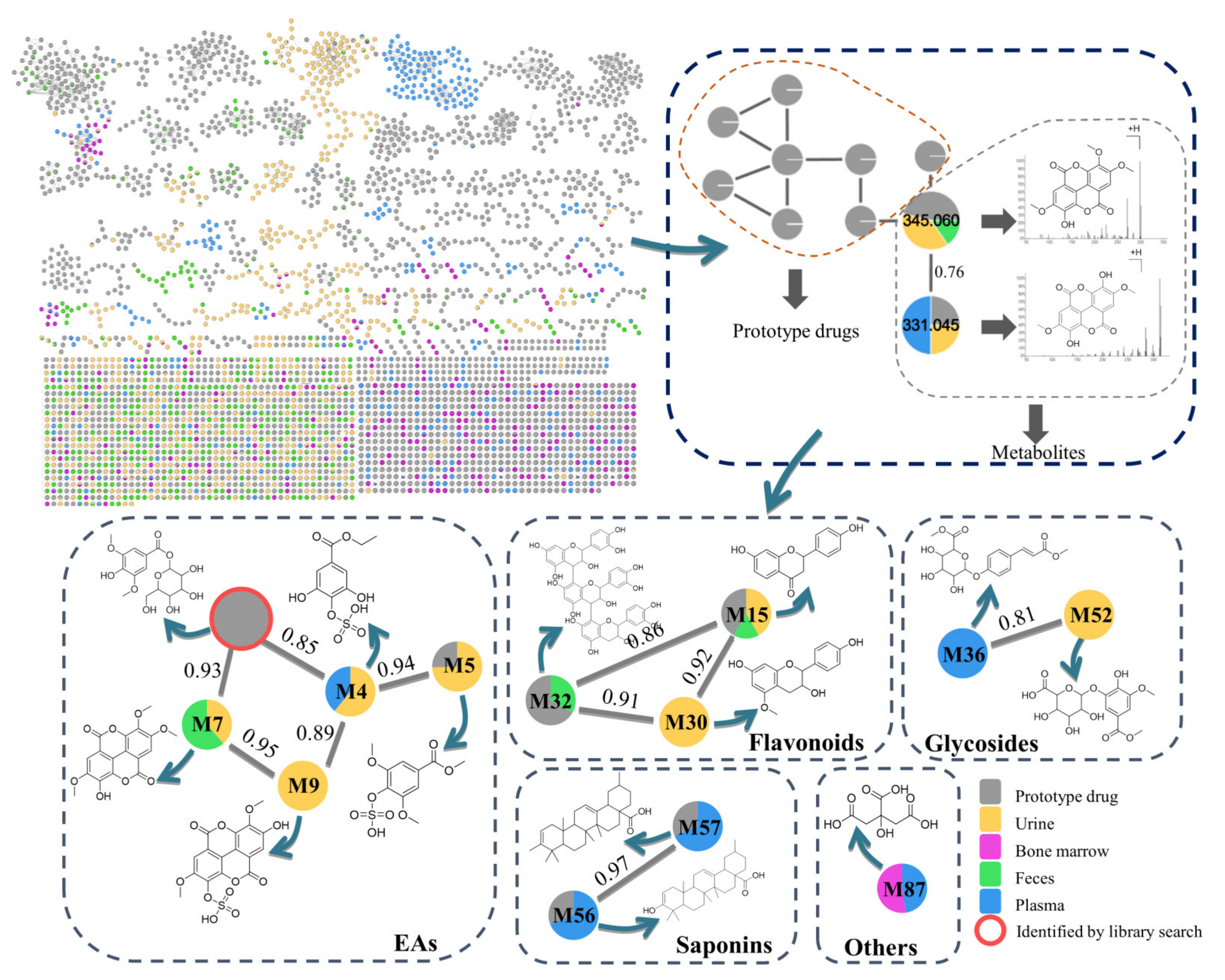
3.2. Construction of the Molecular Network of SO
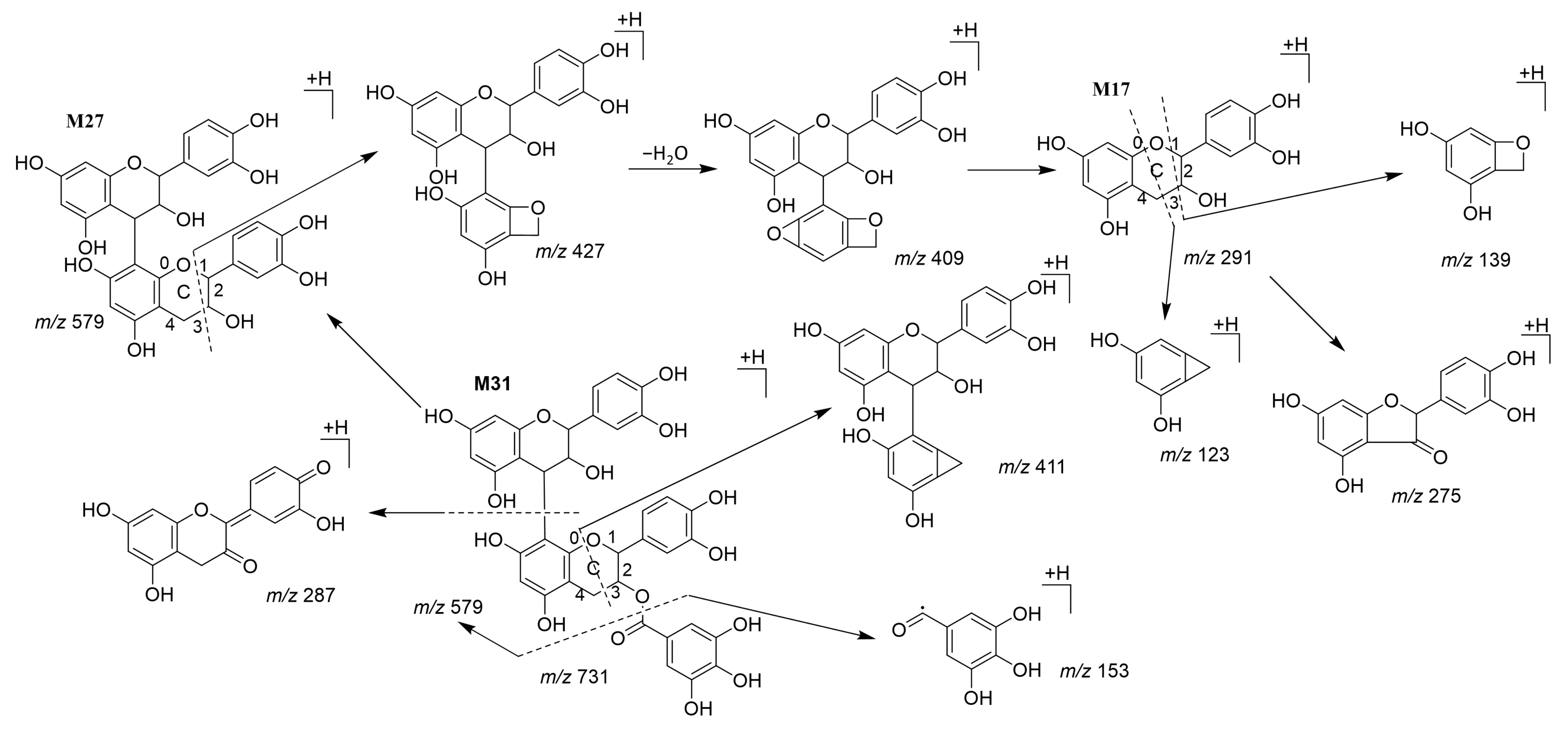
3.3. Characterization of Metabolites
3.3.1. Feces
3.3.2. Urine
3.3.3. Plasma
3.3.4. Bone Marrow
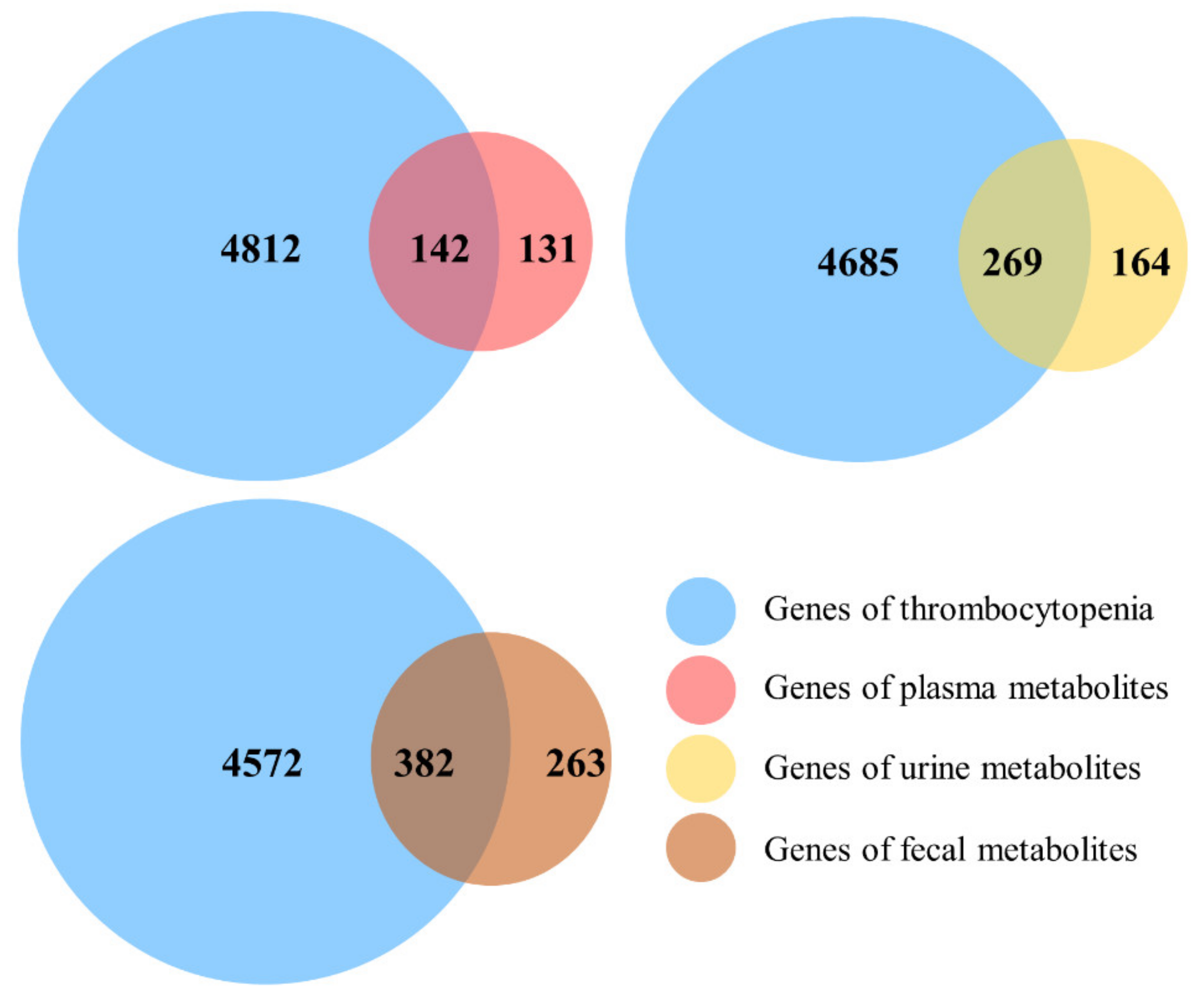
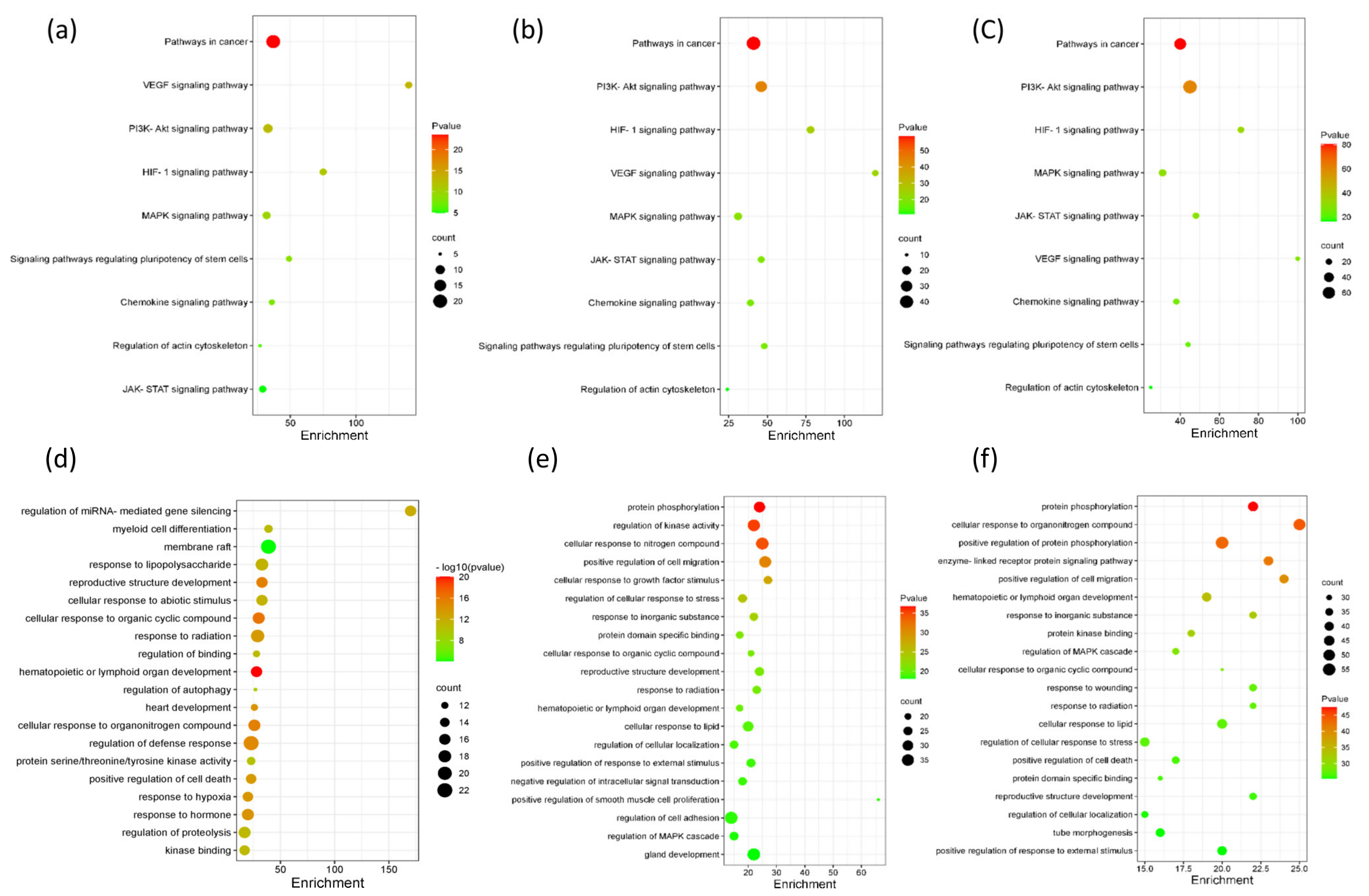
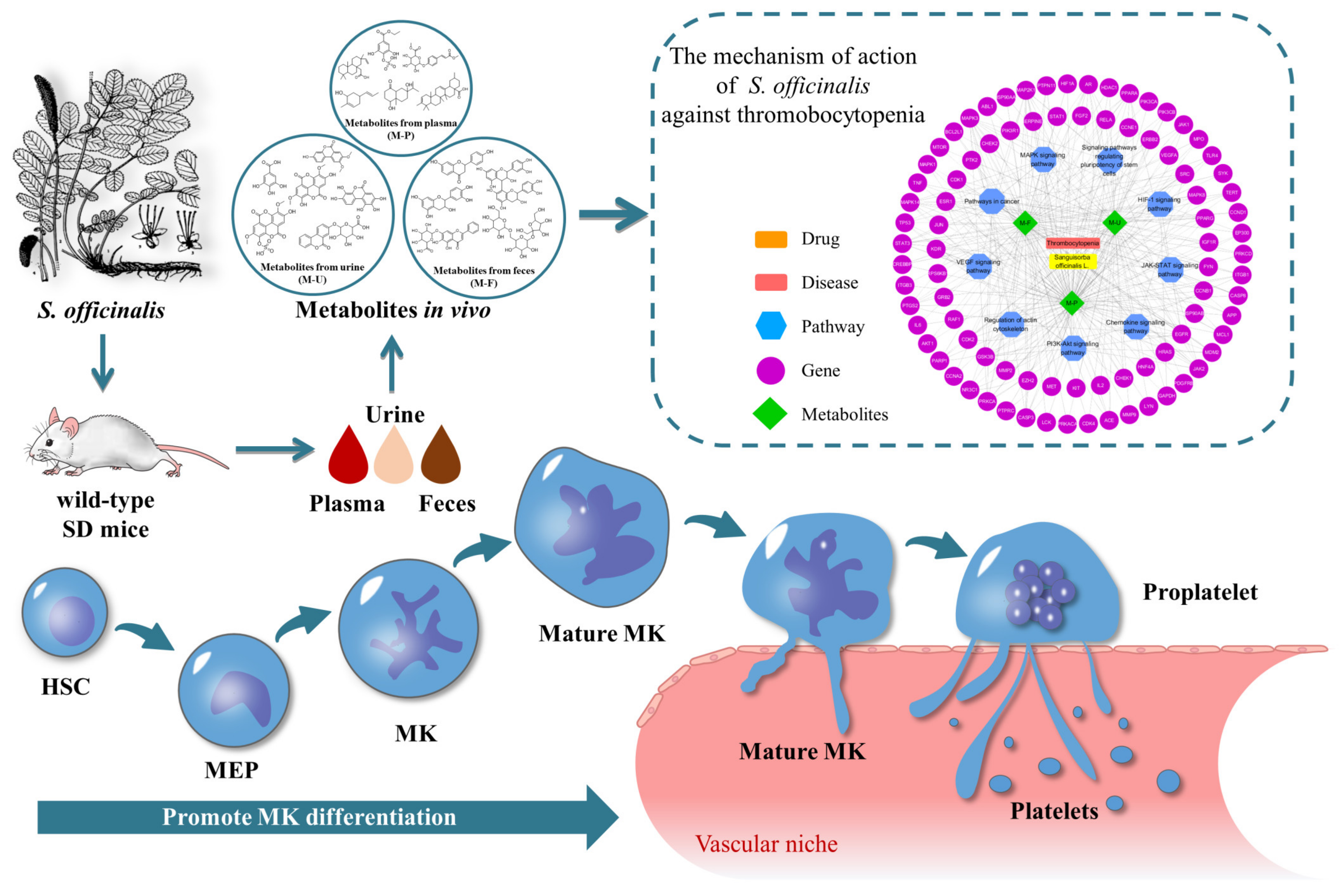
3.4. Analysis of Metabolic Pathways
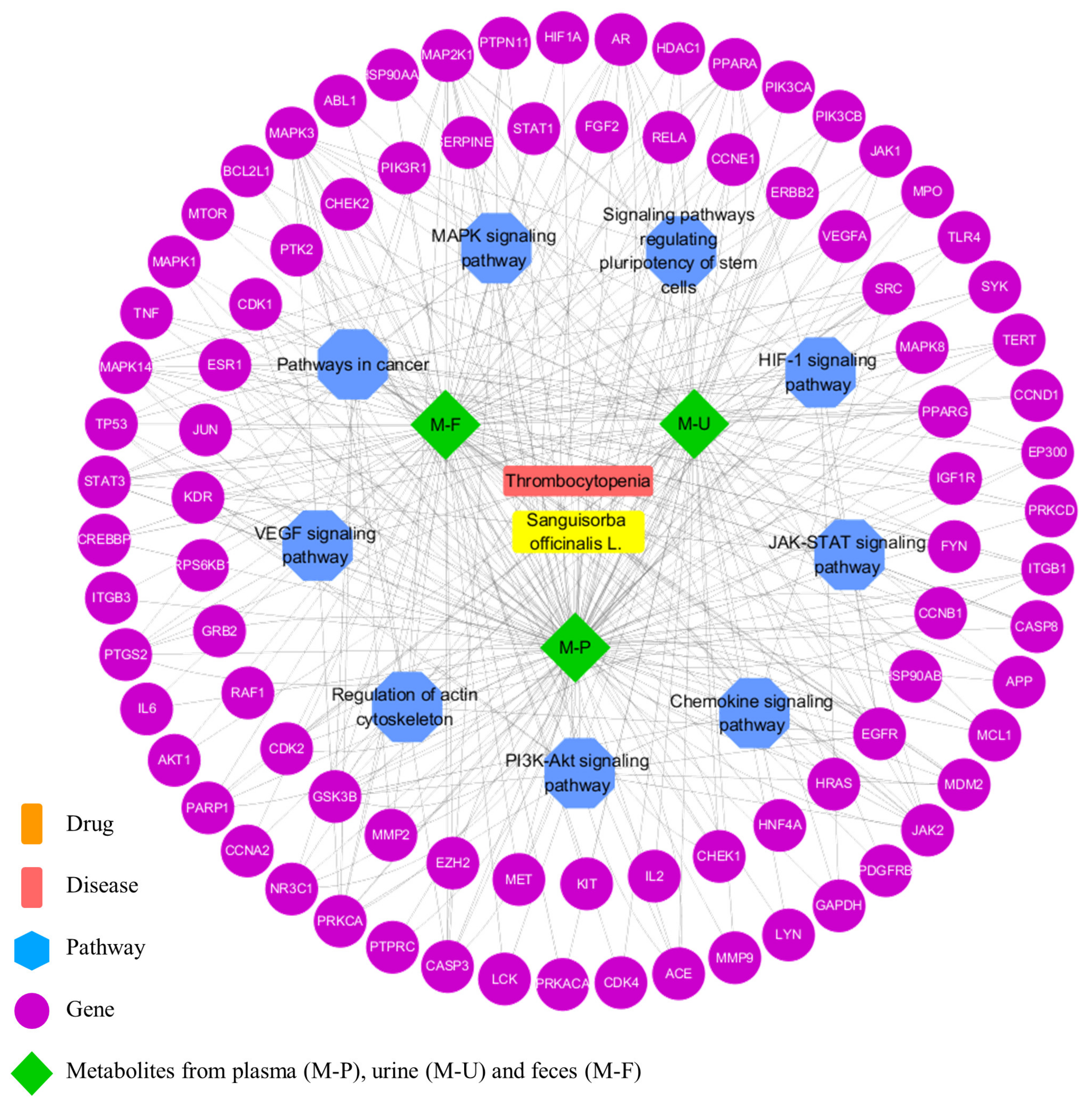
3.5. Network Pharmacology Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, L.; Cao, L.; Shi, D.; Wang, Z.; Xiao, W.; Yao, X.; Li, H.; Yu, Y. Metabolic profiles of Jin-hong tablets in rats by ultra-performance liquid chromatography coupled with quadrupole time-of-flight tandem mass spectrometry. Biomed. Chromatogr. 2021, 35, e5072. [Google Scholar] [CrossRef] [PubMed]
- Mohimani, H.; Gurevich, A.; Mikheenko, A.; Garg, N.; Nothias, L.F.; Ninomiya, A.; Takada, K.; Dorrestein, P.C.; Pevzner, P.A. Dereplication of peptidic natural products through database search of mass spectra. Nat. Chem. Biol. 2016, 13, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, H.; Li, J.; Wu, B.; Zhang, X.; Li, Y.; Yang, S.; He, M.; Feng, Y. A robust platform based on ultra-high performance liquid chromatography Quadrupole time of flight tandem mass spectrometry with a two-step data mining strategy in the investigation, classification, and identification of chlorogenic acids in Ainsliaea fragrans Champ. J. Chromatogr. A 2017, 1502, 38–50. [Google Scholar] [CrossRef]
- Zhang, D.; Xiong, L.; Fang, L.; Li, H.; Zhao, X.; Luan, R.; Zhao, P.; Zhang, X. Systematic characterization of the absorbed components of Ligustri Lucidi Fructus and their metabolic pathways in rat plasma by ultra-high-performance liquid chromatography-Q-Exactive Orbitrap tandem mass spectrometry combined with network pharmacology. J. Sep. Sci. 2021, 44, 4343–4367. [Google Scholar] [CrossRef]
- Wei, W.; Hou, J.; Yao, C.; Bi, Q.; Wang, X.; Li, Z.; Jin, Q.; Lei, M.; Feng, Z.; Wu, W.; et al. A high-efficiency strategy integrating offline two-dimensional separation and data post-processing with dereplication: Characterization of bufadienolides in Venenum bufonis as a case study. J. Chromatogr. A 2019, 1603, 179–189. [Google Scholar] [CrossRef]
- Wakimoto, T. Toward the Dark Matter of Natural Products. Chem. Rec. 2017, 17, 1124–1134. [Google Scholar] [CrossRef]
- Vassilev, N.G.; Simova, S.D.; Dangalov, M.; Velkova, L.; Atanasov, V.; Dolashki, A.; Dolashka, P. An 1H NMR- and MS-Based Study of Metabolites Profiling of Garden Snail Helix aspersa Mucus. Metabolites 2020, 10, 360. [Google Scholar] [CrossRef]
- Maxim, N.; Lisse, D.; Guy, B.; Patrick, A.; Deirdre, C. Development of a HILIC-MS/MS Method for the Quantification of Histamine and its Main Metabolites in Human Urine Samples. Talanta 2020, 220, 121328. [Google Scholar] [CrossRef]
- Sowjanya, G.; Ganapaty, S.; Sharma, R. In vitro test methods for metabolite identification: A review. Asian J. Pharm. Pharmacol. 2019, 5, 441–450. [Google Scholar] [CrossRef]
- Duan, X.; Pan, L.; Peng, D.; Bao, Q.; Xiao, L.; Zhou, A.; Wu, H.; Peng, C.; Chen, W. Analysis of the active components and metabolites of Taohong Siwu decoction by using ultra high performance liquid chromatography quadrupole time-of-flight mass spectrometry. J. Sep. Sci. 2020, 43, 4131–4147. [Google Scholar] [CrossRef]
- Chen, L.-L.; Chen, C.-H.; Zhang, X.-X.; Wang, Y.; Wang, S.-F. Identification of constituents in Gui-Zhi-Jia-Ge-Gen-Tang by LC-IT-MS combined with LC-Q-TOF-MS and elucidation of their metabolic networks in rat plasma after oral administration. Chin. J. Nat. Med. 2019, 17, 803–821. [Google Scholar] [CrossRef]
- Fu, J.; Wu, H.; Wu, H.; Deng, R.; Li, F. Chemical and metabolic analysis of Achyranthes bidentate saponins with intestinal microflora-mediated biotransformation by ultra-performance liquid chromatography-quadrupole time-of-flight mass spectrometry coupled with metabolism platform. J. Pharm. Biomed. Anal. 2019, 170, 305–320. [Google Scholar] [CrossRef]
- Gao, M.-X.; Tang, X.-Y.; Zhang, F.-X.; Yao, Z.-H.; Yao, X.-S.; Dai, Y. Biotransformation and metabolic profile of Xian-Ling-Gu-Bao capsule, a traditional Chinese medicine prescription, with rat intestinal microflora by ultra-performance liquid chromatography coupled with quadrupole time-of-flight tandem mass spectrometry analysis. J. Biomed. Chromatogr. 2017, 32, e4160. [Google Scholar] [CrossRef]
- Miao, W.-J.; Wang, Q.; Bo, T.; Ye, M.; Qiao, X.; Yang, W.-Z.; Xiang, C.; Guan, X.-Y.; Guo, D.-A. Rapid characterization of chemical constituents and rats metabolites of the traditional Chinese patent medicine Gegen-Qinlian-Wan by UHPLC/DAD/qTOF-MS. J. Pharm. Biomed. Anal. 2013, 72, 99–108. [Google Scholar] [CrossRef]
- Wang, H.; Sun, H.; Zhang, A.; Li, Y.; Wang, L.; Shi, H.; Dizou, X.L.; Wang, X. Rapid identification and comparative analysis of the chemical constituents and metabolites of Phellodendri amurensis cortex and Zhibai dihuang pill by ultra-performance liquid chromatography with quadrupole TOF-MS. J. Sep. Sci. 2013, 36, 3874–3882. [Google Scholar] [CrossRef]
- Guo, X.; Lin, S.; Yang, P.; Ye, J.; Du, J.; Mu, X.; Mi, N.; Qi, X.; Lei, H.; Zhang, W.; et al. Rapid characterization and identification of the chemical constituents and rat metabolites of Deng-Zhan-Xi-Xin injection using ultra high performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry. J. Sep. Sci. 2018, 41, 3569–3582. [Google Scholar] [CrossRef]
- Wan, M.-Q.; Liu, X.-Y.; Gao, H.; Wang, T.-X.; Yang, Y.-F.; Jia, L.-Y.; Yang, X.-W.; Zhang, Y.-B. Systematic analysis of the metabolites of Angelicae Pubescentis Radix by UPLC-Q-TOF-MS combined with metabonomics approaches after oral administration to rats. J. Pharm. Biomed. Anal. 2020, 188, 113445. [Google Scholar] [CrossRef]
- Zuo, R.; Ren, W.; Bian, B.-L.; Wang, H.-J.; Wang, Y.-N.; Hu, H.; Zhao, H.-Y.; Si, N. Metabolic fate analysis of Huang–Lian–Jie–Du Decoction in rat urine and feces by LC–IT-MS combining with LC–FT-ICR-MS: A feasible strategy for the metabolism study of Chinese medical formula. Xenobiotica 2015, 46, 65–81. [Google Scholar] [CrossRef]
- Cao, J.L.; Wang, S.S.; Hu, H.; He, C.W.; Wan, J.B.; Su, H.X.; Wang, Y.T.; Li, P. Online comprehensive two-dimensional hydrophilic interaction chromatographyxreversed-phase liquid chromatography coupled with hybrid linear ion trap Orbitrap mass spectrometry for the analysis of phenolic acids in Salvia miltiorrhiza. J. Chromatogr. A. 2018, 1536, 216–227. [Google Scholar] [CrossRef]
- Xu, T.; Li, S.; Sun, Y.; Pi, Z.; Liu, S.; Song, F.; Liu, Z. Systematically characterize the absorbed effective substances of Wutou Decoction and their metabolic pathways in rat plasma using UHPLC-Q-TOF-MS combined with a target network pharmacological analysis. J. Pharm. Biomed. Anal. 2017, 141, 95–107. [Google Scholar] [CrossRef]
- Feng, G.; Liu, Z.; Liu, S.; Xing, J.; Song, F.; Pi, Z. A target integration strategy for analyzing multidimensional chemical and metabolic substance groups of Ding-Zhi-Xiao-Wan prescription by using ultra-high performance liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2019, 1608, 460412. [Google Scholar] [CrossRef]
- Lin, P.; Qin, Z.F.; Yao, Z.H.; Wang, L.; Zhang, W.Y.; Yu, Y.; Dai, Y.; Zhou, H.; Yao, X.S. Metabolites profile of Gualou Xiebai Baijiu decoction (a classical traditional Chinese medicine prescription) in rats by ultra-performance liquid chromatography coupled with quadrupole time-of-flight tandem mass spectrometry. J. Chromatogr. B 2018, 1085, 72–88. [Google Scholar] [CrossRef]
- Li, Z.H.; Guo, X.M.; Cao, Z.L.; Liu, X.J.; Liao, X.N.; Huang, C.; Xu, W.Q.; Liu, L.; Yang, P. New MS network analysis pattern for the rapid identification of constituents from traditional Chinese medicine prescription Lishukang capsules in vitro and in vivo based on UHPLC/Q-TOF-MS. Talanta 2018, 189, 606–621. [Google Scholar] [CrossRef]
- Hu, L.; Yao, Z.; Qin, Z.; Liu, L.; Song, X.; Dai, Y.; Kiyohara, H.; Yamada, H.; Yao, X. In vivo metabolic profiles of Bu-Zhong-Yi-Qi-Tang, a famous traditional Chinese medicine prescription, in rats by ultra-high-performance liquid chromatography coupled with quadrupole time-of-flight tandem mass spectrometry. J. Pharm. Biomed. Anal. 2019, 171, 81–98. [Google Scholar] [CrossRef]
- Xing, S.; Hu, Y.; Yin, Z.; Liu, M.; Tang, X.; Fang, M.; Huan, T. Retrieving and Utilizing Hypothetical Neutral Losses from Tandem Mass Spectra for Spectral Similarity Analysis and Unknown Metabolite Annotation. Anal. Chem. 2020, 92, 14476–14483. [Google Scholar] [CrossRef]
- Lynn, K.-S.; Cheng, M.-L.; Chen, Y.-R.; Hsu, C.; Chen, A.; Lih, T.M.; Chang, H.-Y.; Huang, C.-J.; Shiao, M.-S.; Pan, W.-H.; et al. Metabolite Identification for Mass Spectrometry-Based Metabolomics Using Multiple Types of Correlated Ion Information. Anal. Chem. 2015, 87, 2143–2151. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Hautbergue, T.; Jamin, E.L.; Costantino, R.; Tadrist, S.; Meneghetti, L.; Tabet, J.-C.; Debrauwer, L.; Oswald, I.P.; Puel, O. Combination of Isotope Labeling and Molecular Networking of Tandem Mass Spectrometry Data To Reveal 69 Unknown Metabolites Produced by Penicillium nordicum. Anal. Chem. 2019, 91, 12191–12202. [Google Scholar] [CrossRef]
- Wishart, D.S. Emerging applications of metabolomics in drug discovery and precision medicine. Nat. Rev. Drug Discov. 2016, 15, 473–484. [Google Scholar] [CrossRef]
- Bonneau, N.; Chen, G.; Lachkar, D.; Boufridi, A.; Gallard, J.-F.; Retailleau, P.; Petek, S.; Debitus, C.; Evanno, L.; Beniddir, M.A.; et al. An Unprecedented Blue Chromophore Found in Nature using a “Chemistry First” and Molecular Networking Approach: Discovery of Dactylocyanines A-H. Chem. A Eur. J. 2017, 23, 14454–14461. [Google Scholar] [CrossRef]
- Carriot, N.; Paix, B.; Greff, S.; Viguier, B.; Briand, J.-F.; Culioli, G. Integration of LC/MS-based molecular networking and classical phytochemical approach allows in-depth annotation of the metabolome of non-model organisms—The case study of the brown seaweed Taonia atomaria. Talanta 2020, 225, 121925. [Google Scholar] [CrossRef]
- Dai, Y.; Zhang, K.; Xiong, L.; Wang, L.; Guo, Z.; Yang, J.; Wu, A.; Wu, J.; Zeng, J. Comprehensive profiling of Sanguisorba officinalis using offline two-dimensional mixed-mode liquid chromatography × reversed-phase liquid chromatography, tandem high-resolution mass spectrometry, and molecular network. J. Sep. Sci. 2022, 45, 1727–1736. [Google Scholar] [CrossRef]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682–690. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, C.; Su, X.; Zhu, J.; Chen, X.; Fu, Y.; Wang, Z.; Zhou, J.; Xiao, W.; Zheng, C.; et al. Deciphering the multicomponent synergy mechanism from a systems pharmacology perspective: Application to Gualou Xiebai Decoction for coronary heart disease. J. Funct. Foods 2018, 47, 143–155. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, X.; Zhong, Y.; Zhou, L.; Liang, J.; Zeng, J.; Zhou, L.; Yuan, E.; Zhu, J.; Wang, C.-Z.; et al. Active microbial metabolites study on antitussive and expectorant effects and metabolic mechanisms of platycosides fraction of Platycodonis Radix. J. Chromatogr. B 2022, 1195, 123171. [Google Scholar] [CrossRef]
- Lin, J.; Zeng, J.; Liu, S.; Shen, X.; Jiang, N.; Wu, Y.-S.; Li, H.; Wang, L.; Wu, J.-M. DMAG, a novel countermeasure for the treatment of thrombocytopenia. Mol. Med. 2021, 27, 149. [Google Scholar] [CrossRef]
- Mu, X.; Xu, X.; Guo, X.; Yang, P.; Du, J.; Mi, N.; Cheng, T.; Lu, L.; Qi, X.; Wang, X.; et al. Identification and characterization of chemical constituents in Dengzhan Shengmai Capsule and their metabolites in rat plasma by ultra-performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry. J. Chromatogr. B 2019, 1108, 54–64. [Google Scholar] [CrossRef]
- Wu, J.; Zhong, Q.; Wang, T.; Wang, C.; Du, Y.; Ji, S.; Wang, L.; Guo, M.; Tang, D. MS-based metabolite analysis of two licorice chalcones in mice plasma, bile, feces, and urine after oral administration. Biomed. Chromatogr. 2020, 35, e4998. [Google Scholar] [CrossRef]
- Li, C.-X.; Liang, J.; Song, Y.; Chai, J.-H.; Kuang, H.-X.; Xia, Y.-G. Structural characterization of the metabolites of orally ingested hederasaponin B, a natural saponin that is isolated from Acanthopanax senticosus leaves by liquid chromatography–mass spectrometry. J. Pharm. Biomed. Anal. 2021, 197, 113929. [Google Scholar] [CrossRef]
- Zani, C.L.; Carroll, A.R. Database for Rapid Dereplication of Known Natural Products Using Data from MS and Fast NMR Experiments. J. Nat. Prod. 2017, 80, 1758–1766. [Google Scholar] [CrossRef]
- Liu, R.; Meng, C.; Zhang, Z.; Ma, H.; Lv, T.; Xie, S.; Liu, Y.; Wang, C. Comparative metabolism of schaftoside in healthy and calcium oxalate kidney stone rats by UHPLC-Q-TOF-MS/MS method. Anal. Biochem. 2020, 597, 113673. [Google Scholar] [CrossRef]
- Zhang, F.-X.; Li, Z.-T.; Li, C.; Li, M.; Yao, Z.-H.; Yao, X.-S.; Dai, Y. Characterization of lignans in Forsythiae Fructus and their metabolites in rats by ultra-performance liquid chromatography coupled time-of-flight mass spectrometry. J. Pharm. Pharmacol. 2020, 72, 1879–1892. [Google Scholar] [CrossRef]
- Liang, J.; Xu, F.; Zhang, Y.-Z.; Huang, S.; Zang, X.-Y.; Zhao, X.; Zhang, L.; Shang, M.-Y.; Yang, D.-H.; Wang, X.; et al. The profiling and identification of the absorbed constituents and metabolites of Paeoniae Radix Rubra decoction in rat plasma and urine by the HPLC–DAD–ESI-IT-TOF-MSn technique: A novel strategy for the systematic screening and identification of absorbed constituents and metabolites from traditional Chinese medicines. J. Pharm. Biomed. Anal. 2013, 83, 108–121. [Google Scholar] [CrossRef]
- Shahraki, A.; Ebrahimi, A. Ellagitannin derivatives and some conjugated metabolites: Aqueous-DMSO proton affinities and acidity constants. Struct. Chem. 2019, 30, 1343–1351. [Google Scholar] [CrossRef]
- Ma, J.-Y.; Zhou, X.; Fu, J.; He, C.-Y.; Feng, R.; Huang, M.; Shou, J.-W.; Zhao, Z.-X.; Li, X.-Y.; Zhang, L.; et al. In Vivo Metabolite Profiling of a Purified Ellagitannin Isolated from Polygonum capitatum in Rats. Molecules 2016, 21, 1110. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, H.; Kishino, S.; Kudoh, M.; Yamamoto, H.; Ogawa, J. Evaluation of electron-transferring cofactor mediating enzyme systems involved in urolithin dehydroxylation in Gordonibacter urolithinfaciens DSM 27213. J. Biosci. Bioeng. 2020, 129, 552–557. [Google Scholar] [CrossRef]
- Chen, X.; Li, B.; Gao, Y.; Ji, J.; Wu, Z.; Chen, S. Saponins from Sanguisorba officinalis Improve Hematopoiesis by Promoting Survival through FAK and Erk1/2 Activation and Modulating Cytokine Production in Bone Marrow. Front. Pharmacol. 2017, 8, 130. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-H.; Zhou, S.-Y.; Feng, R.; Wang, Y.-Z.; Kong, Y.; Zhou, Y.; Zhang, J.-M.; Wang, M.; Zhao, J.-Z.; Wang, Q.-M.; et al. Increased prostacyclin levels inhibit the aggregation and activation of platelets via the PI3K–AKT pathway in prolonged isolated thrombocytopenia after allogeneic hematopoietic stem cell transplantation. Thromb. Res. 2016, 139, 1–9. [Google Scholar] [CrossRef]
- Farsani, S.S.M.; Sadeghizadeh, M.; Gholampour, M.A.; Safari, Z.; Najafi, F. Nanocurcumin as a novel stimulator of megakaryopoiesis that ameliorates chemotherapy-induced thrombocytopenia in mice. Life Sci. 2020, 256, 117840. [Google Scholar] [CrossRef]
- Gerds, A.T. Beyond JAK-STAT: Novel therapeutic targets in Ph-negative MPN. Am. Soc. Hematol. Educ. Program. 2019, 2019, 407–414. [Google Scholar] [CrossRef]
- Huang, R.; Hayashi, Y.; Yan, X.; Bu, J.; Wang, J.; Zhang, Y.; Zhou, Y.; Tang, Y.; Wu, L.; Xu, Z.; et al. HIF1A is a critical downstream mediator for hemophagocytic lymphohistiocytosis. Haematologica 2017, 102, 1956–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramasz, B.; Krüger, A.; Reinhardt, J.; Sinha, A.; Gerlach, M.; Gerbaulet, A.; Reinhardt, S.; Dahl, A.; Chavakis, T.; Wielockx, B.; et al. Hematopoietic stem cell response to acute thrombocytopenia requires signaling through distinct receptor tyrosine kinases. Blood 2019, 134, 1046–1058. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Li, Y.; Li, H.; Lyu, M.; Zhang, D.; Fu, R.; Guan, Y.; Wang, S.; Sun, B.; Dou, X.; et al. Increased plasma sCXCL16 levels may have a relationship with Th1/Th2 imbalance in primary immune thrombocytopenia. Cytokine 2017, 99, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Poulter, N.S.; Thomas, S.G. Cytoskeletal regulation of platelet formation: Coordination of F-actin and microtubules. Int. J. Biochem. Cell Biol. 2015, 66, 69–74. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, Y.; Zhang, K.; Wang, L.; Xiong, L.; Huang, F.; Huang, Q.; Wu, J.; Zeng, J. Rapid Profiling of Metabolites Combined with Network Pharmacology to Explore the Potential Mechanism of Sanguisorba officinalis L. against Thrombocytopenia. Metabolites 2022, 12, 1074. https://doi.org/10.3390/metabo12111074
Dai Y, Zhang K, Wang L, Xiong L, Huang F, Huang Q, Wu J, Zeng J. Rapid Profiling of Metabolites Combined with Network Pharmacology to Explore the Potential Mechanism of Sanguisorba officinalis L. against Thrombocytopenia. Metabolites. 2022; 12(11):1074. https://doi.org/10.3390/metabo12111074
Chicago/Turabian StyleDai, Yubei, Kailian Zhang, Long Wang, Ling Xiong, Feihong Huang, Qianqian Huang, Jianming Wu, and Jing Zeng. 2022. "Rapid Profiling of Metabolites Combined with Network Pharmacology to Explore the Potential Mechanism of Sanguisorba officinalis L. against Thrombocytopenia" Metabolites 12, no. 11: 1074. https://doi.org/10.3390/metabo12111074