
Enhancement of Seawater Stress Tolerance in Barley by the Endophytic Fungus Aspergillus ochraceus

 ,
,  , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Results
2.1. Screening of Salt-Tolerant Endophytic Fungi
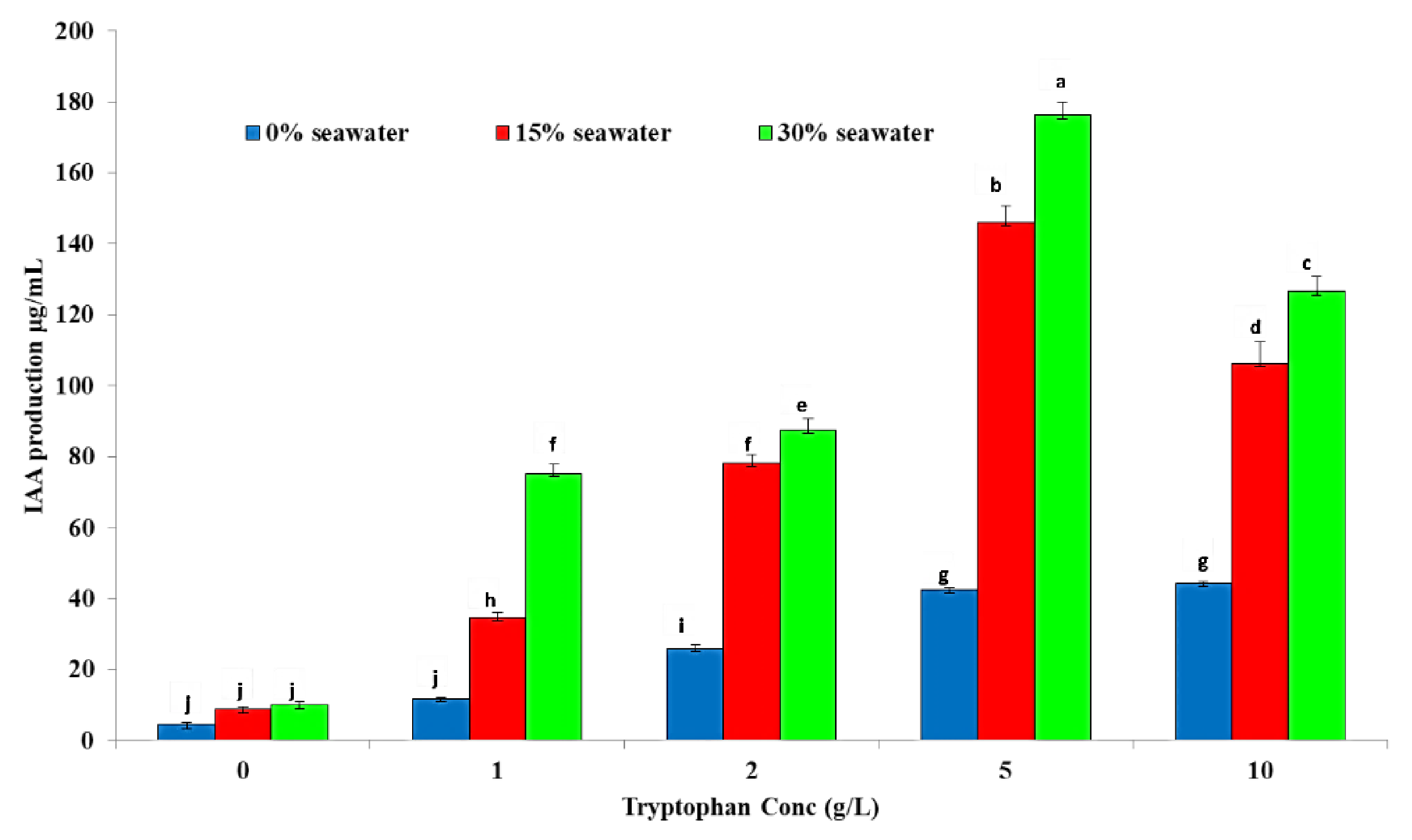
2.2. Indole Acetic Acid (IAA) Production
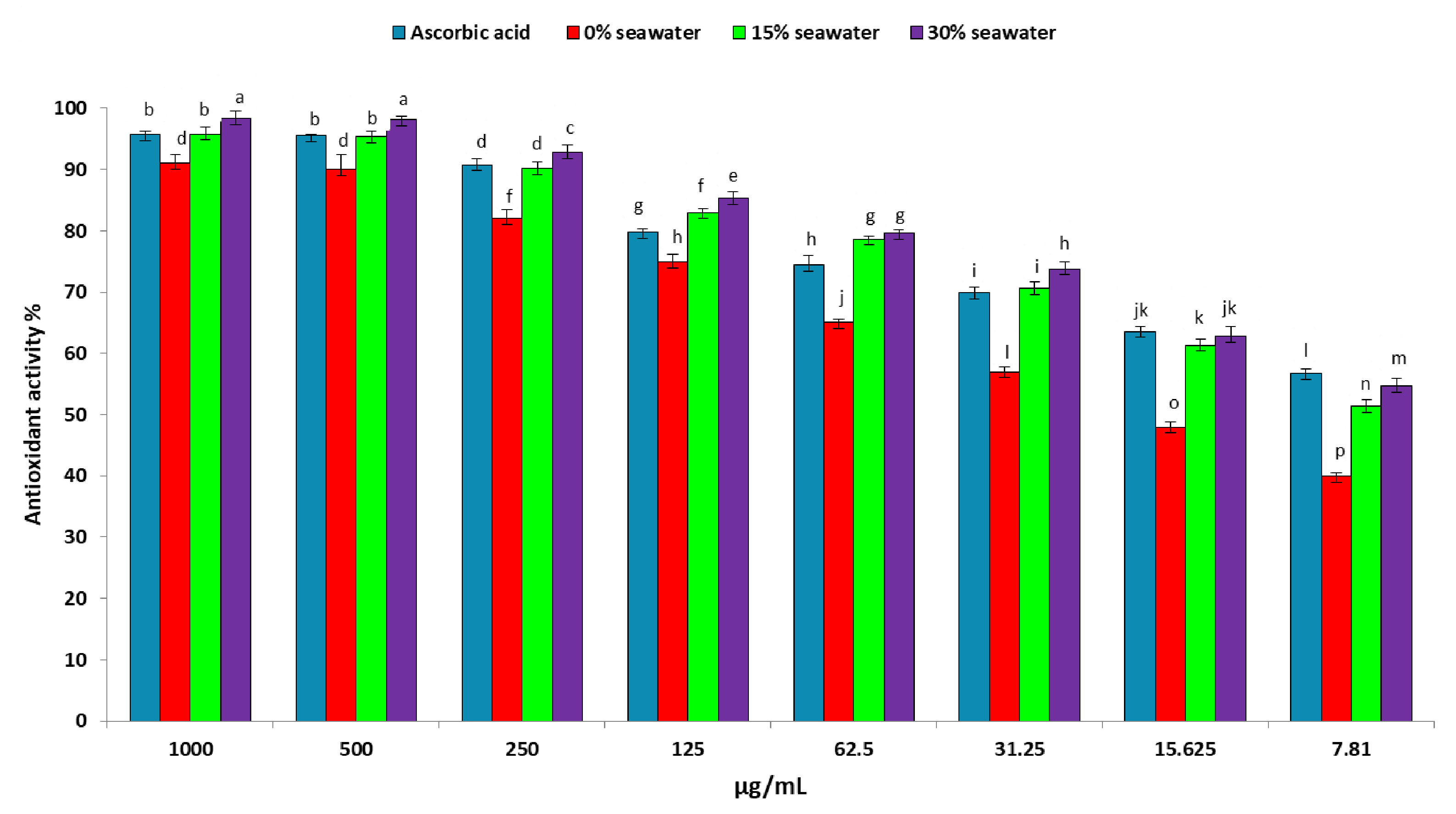
2.3. Antioxidant Production
2.4. Antifungal Activity
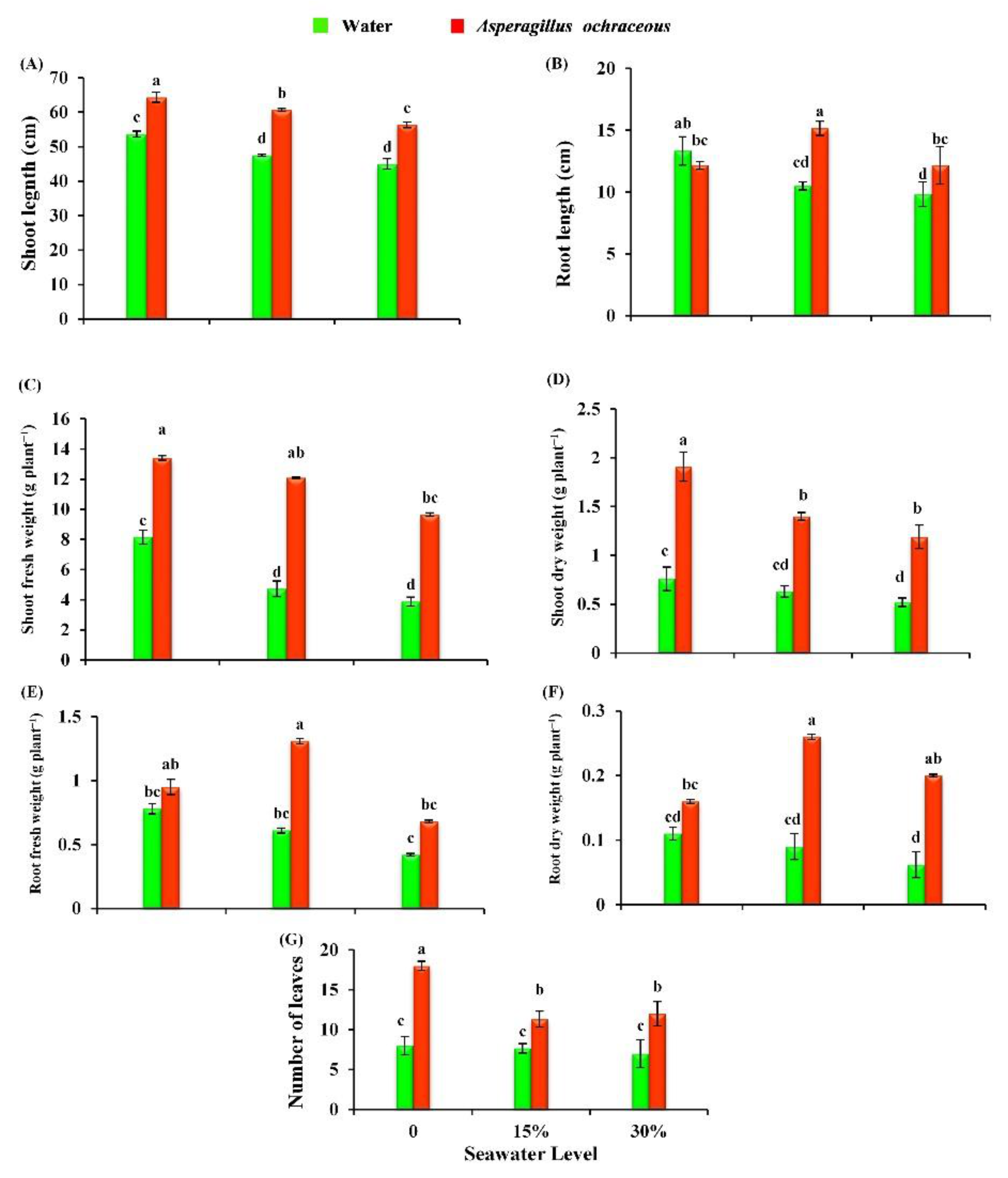
2.5. Aspergillus Ochraceus Application Promoted the Growth of Barley Plants under Salinity Stress Conditions
2.6. Aspergillus Ochraceus Application Enhanced Leaf Pigments of Barley Plants under Salinity Stress Conditions
2.7. Aspergillus Ochraceus Application Affected the Soluble Sugar, Protein, and Proline Contents of Barley Plants under Salinity Stress Conditions
2.7.1. Soluble Sugars
2.7.2. Soluble Proteins
2.7.3. Free Proline
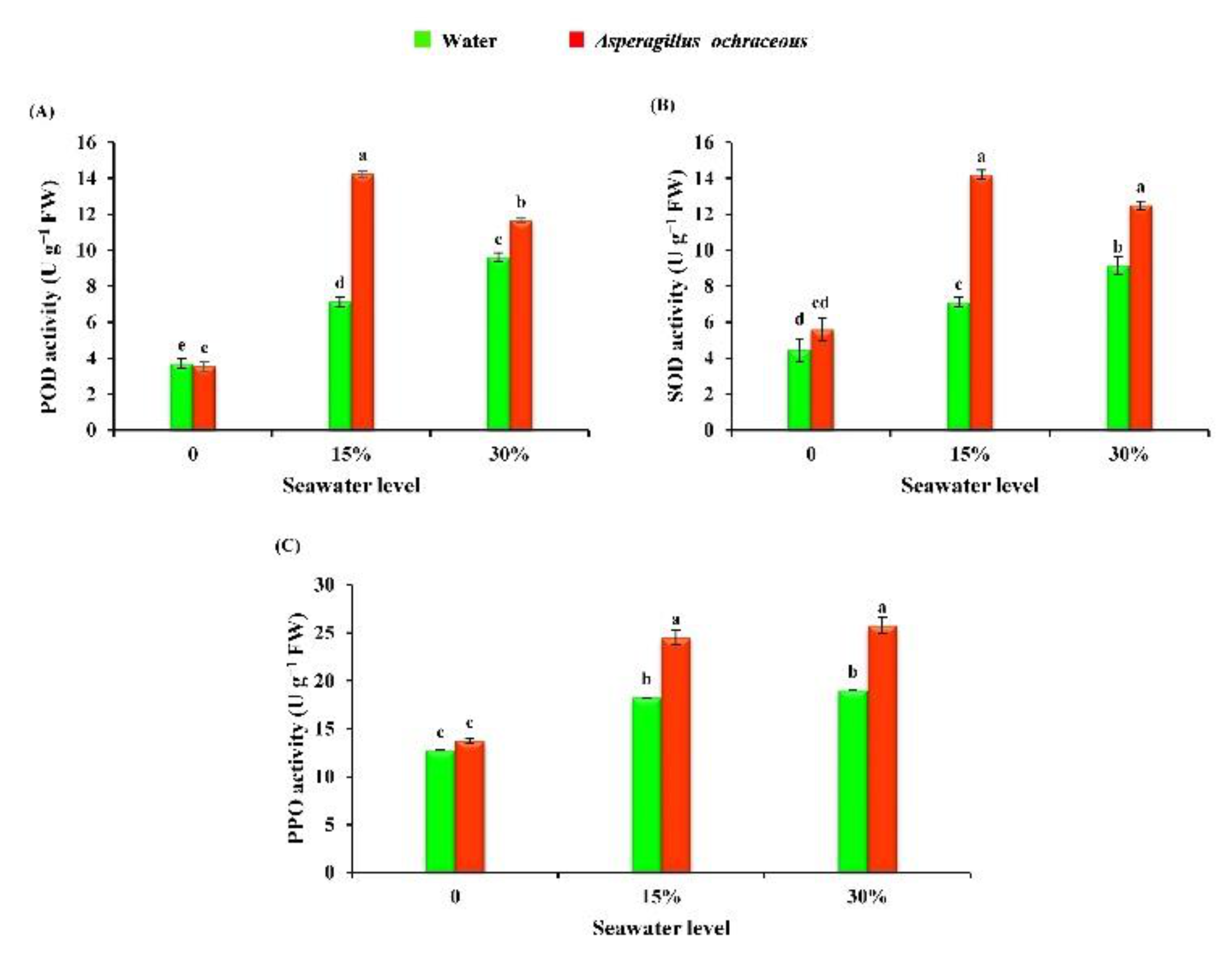
2.8. Aspergillus Ochraceus Application Boosted the Activity of the Antioxidant Enzymes of Barley Plants under Salinity Stress Conditions
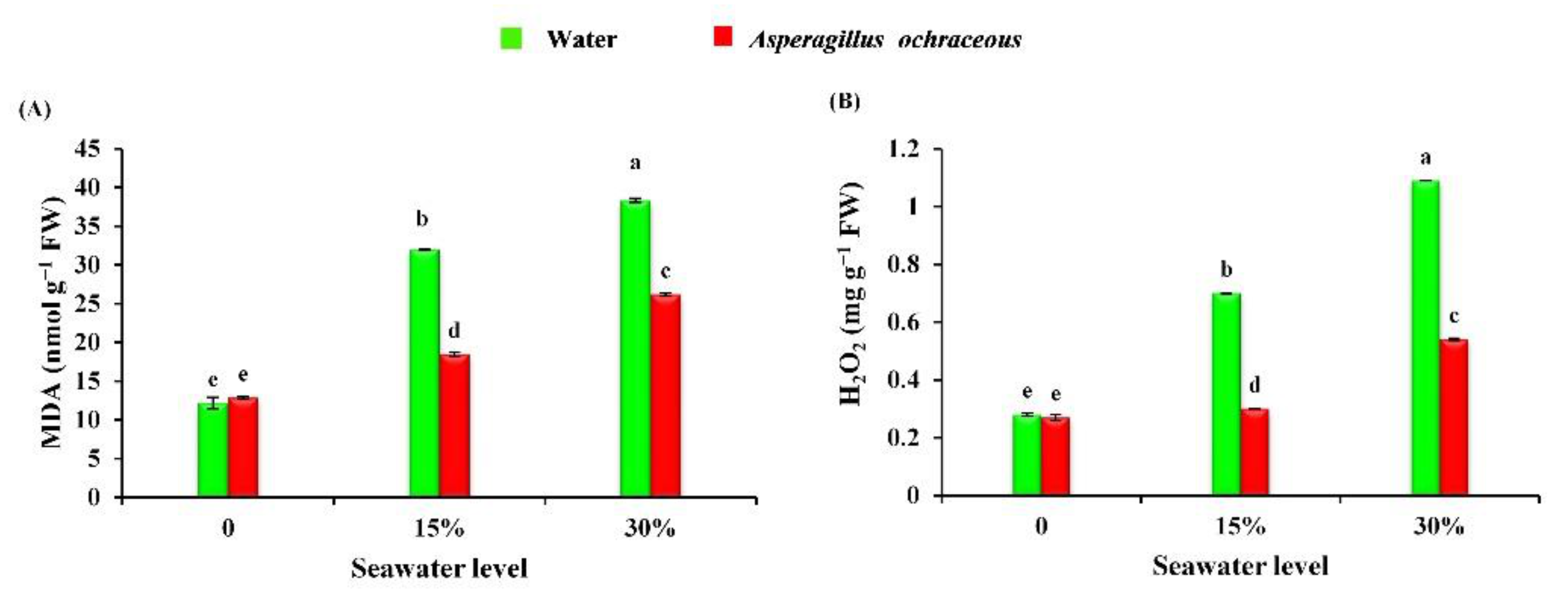
2.9. Aspergillus Ochraceus Application Suppressed the Malondialdehyde and Hydrogen Peroxide Contents of Barley Plants under Salinity Stress Conditions
2.10. Aspergillus Ochraceus Application Increased the Yield Attributes of Barley Plants under Salinity Stress Conditions
3. Discussion
4. Materials and Methods
4.1. Fungal Endophytes and Growth Condition
4.2. Screening of Fungal Endophyte Strains according to Salt Tolerance
4.3. Indole Acetic Acid Production
4.4. Antioxidant Activity of Crude Extracts of A. ochraceus
4.5. Antifungal Activity
4.6. Pot Experiment
4.6.1. Biochemical Parameters
Estimation of Photosynthetic Pigments
Determination of Osmolyte Contents
Extraction and Determination of Antioxidant Enzymes
Estimation of Malondialdehyde Content
Determination of Hydrogen Peroxide (H2O2) Content
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bybordi, A. Effect of different ratios of nitrate and ammonium on photosynthesis, and fatty acid composition of canola under saline conditions. Int. J. Agric. Crop Sci. 2012, 4, 622–626. [Google Scholar]
- Ewais, E.A.; Ismail, M.A.; Amin, M.A.; Badawy, A.A. Efficiency of salicylic acid and glycine on sugar beet plants grown under heavy metals pollution. Egypt. J. Biotechnol. 2015, 48, 112–126. [Google Scholar]
- De Oliveira, A.B.; Alencar, N.L.M.; Gomes-Filho, E. Comparison between the water and salt stress effects on plant growth and development. Responses Org. Water Stress 2013, 67. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, Z.S.; Khan, M.A. Some physiological attributes of dimorphic seeds of Halopyrum mucronatum (L.) Stapf. Pakistan J. Bot. 2013, 45, 1975–1979. [Google Scholar]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Osman, M.S.; Badawy, A.A.; Osman, A.I.; Abdel Latef, A.A.H. Ameliorative impact of an extract of the halophyte Arthrocnemum macrostachyum on growth and biochemical parameters of soybean under salinity stress. J. Plant Growth Regul. 2021, 40, 1245–1256. [Google Scholar] [CrossRef]
- Ren, S.; Lyle, C.; Jiang, G.; Penumala, A. Soybean salt tolerance 1 (GmST1) reduces ROS production, enhances ABA sensitivity, and abiotic stress tolerance in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 445. [Google Scholar] [CrossRef] [Green Version]
- Atzori, G.; de Vos, A.C.; van Rijsselberghe, M.; Vignolini, P.; Rozema, J.; Mancuso, S.; van Bodegom, P.M. Effects of increased seawater salinity irrigation on growth and quality of the edible halophyte Mesembryanthemum crystallinum L. under field conditions. Agric. Water Manag. 2017, 187, 37–46. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Mostofa, M.G.; Rahman, M.M.; Abdel-Farid, I.B.; Tran, L.P. Extracts from yeast and carrot roots enhance maize performance under seawater-induced salt stress by altering physio-biochemical characteristics of stressed plants. J. Plant Growth Regul. 2019, 38, 966–979. [Google Scholar] [CrossRef]
- Asaf, S.; Hamayun, M.; Khan, A.L.; Waqas, M.; Khan, M.A.; Jan, R.; Lee, I.J.; Hussain, A. Salt tolerance of Glycine max. L induced by endophytic fungus Aspergillus flavus CSH1, via regulating its endogenous hormones and antioxidative system. Plant Physiol. Biochem. 2018, 128, 13–23. [Google Scholar]
- Powthong, P.; Jantrapanukorn, B.; Thongmee, A.; Suntornthiticharoen, P. Screening of antimicrobial activities of the endophytic fungi isolated from Sesbania grandiflora (L.) pers. J. Agric. Sci. Technol. 2013, 15, 1513–1522. [Google Scholar]
- Pavithra, G.; Bindal, S.; Rana, M.; Srivastava, S. Role of endophytic microbes against plant pathogens: A review. Asian J. Plant Sci. 2020, 19, 54–62. [Google Scholar]
- Rana, K.L.; Kour, D.; Kaur, T.; Devi, R.; Yadav, A.N.; Yadav, N.; Dhaliwal, H.S.; Saxena, A.K. Endophytic microbes: Biodiversity, plant growth-promoting mechanisms and potential applications for agricultural sustainability. Antonie Van Leeuwenhoek 2020, 113, 1075–1107. [Google Scholar] [CrossRef]
- Abro, M.A.; Sun, X.; Li, X.; Jatoi, G.H.; Guo, L.-D. Biocontrol potential of fungal endophytes against Fusarium oxysporum f. sp. cucumerinum causing wilt in cucumber. Plant Pathol. J. 2019, 35, 598–608. [Google Scholar] [CrossRef] [PubMed]
- González, V.; Armijos, E.; Garcés-Claver, A. Fungal endophytes as biocontrol agents against the main soil-borne diseases of melon and watermelon in Spain. Agronomy 2020, 10, 820. [Google Scholar] [CrossRef]
- Wei, F.; Zhang, Y.; Shi, Y.; Feng, H.; Zhao, L.; Feng, Z.; Zhu, H. Evaluation of the biocontrol potential of endophytic fungus Fusarium solani CEF559 against Verticillium dahliae in cotton plant. Biomed. Res. Int. 2019, 2019, 3187943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasoni, L.; de Gurfinkel, S. Biocontrol of Rhizoctonia solani by the endophytic fungus Cladorrhinum foecundissimum in cotton plants. Australas. Plant Pathol. 2009, 38, 389–391. [Google Scholar] [CrossRef]
- Chowdhary, K.; Sharma, S. Plant Growth Promotion and Biocontrol Potential of Fungal Endophytes in the Inflorescence of Aloe vera L. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2020, 90, 1045–1055. [Google Scholar] [CrossRef]
- Gurulingappa, P.; Sword, G.A.; Murdoch, G.; McGee, P.A. Colonization of crop plants by fungal entomopathogens and their effects on two insect pests when in planta. Biol. Control. 2010, 55, 34–41. [Google Scholar] [CrossRef]
- Salas-Marina, M.A.; Silva-Flores, M.A.; Cervantes-Badillo, M.G.; Rosales-Saavedra, M.T.; Islas-Osuna, M.A.; Casas-Flores, S. The plant growth-promoting fungus Aspergillus ustus promotes growth and induces resistance against different lifestyle pathogens in Arabidopsis thaliana. J. Microbiol Biotechnol. 2011, 21, 686–696. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.; Shin, D.J.; Won, H.Y.; Song, J.; Kyung, M. Aspergillus terreus JF27 promotes the growth of tomato plants and induces resistance against Pseudomonas syringae pv. tomato. Mycobiology 2018, 46, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Janardan, Y.; Verma, J.P.; Tiwari, K.N. Plant growth promoting activities of fungi and their effect on chickpea plant growth. Asian J. Biol. Sci. 2011, 4, 291–299. [Google Scholar]
- Richardson, A.E.; Barea, J.-M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Schulte, D.; Close, T.J.; Graner, A.; Langridge, P.; Matsumoto, T.; Muehlbauer, G.; Sato, K.; Schulman, A.H.; Waugh, R.; Wise, R.P.; et al. The international barley sequencing consortium—At the threshold of efficient access to the barley genome. Plant Physiol. 2009, 149, 142–147. [Google Scholar] [CrossRef] [Green Version]
- Visioni, A.; Al-Abdallat, A.; Elenien, J.A.; Verma, R.P.S.; Gyawali, S.; Baum, M. Genomics and molecular breeding for improving tolerance to abiotic stress in barley (Hordeum vulgare L.). In Genomics Assisted Breeding of Crops for Abiotic Stress Tolerance, Vol. II.; Springer: Berlin, Germany, 2019; pp. 49–68. [Google Scholar]
- Mwando, E.; Angessa, T.T.; Han, Y.; Li, C. Salinity tolerance in barley during germination—Homologs and potential genes. J. Zhejiang Univ. B 2020, 21, 93–121. [Google Scholar] [CrossRef] [PubMed]
- Omar, M.N.A.; Osman, M.E.H.; Kasim, W.A.; Abd El-Daim, I.A. Improvement of salt tolerance mechanisms of barley cultivated under salt stress using Azospirillum brasilense. In Salinity and Water Stress; Springer: Berlin, Germany, 2009; pp. 133–147. [Google Scholar]
- Jalili, B.; Bagheri, H.; Azadi, S.; Soltani, J. Identification and salt tolerance evaluation of endophyte fungi isolates from halophyte plants. Int. J. Environ. Sci. Technol. 2020, 17, 3459–3466. [Google Scholar] [CrossRef]
- Morsy, M.; Cleckler, B.; Armuelles-Millican, H. Fungal endophytes promote tomato growth and enhance drought and salt tolerance. Plants 2020, 9, 877. [Google Scholar] [CrossRef] [PubMed]
- Biango-Daniels, M.N.; Hodge, K.T. Sea salts as a potential source of food spoilage fungi. Food Microbiol. 2018, 69, 89–95. [Google Scholar] [CrossRef] [PubMed]
- González-Martinez, S.; Soria, I.; Ayala, N.; Portillo-López, A. Culturable halotolerant fungal isolates from Southern California Gulf sediments. Open Agric. 2017, 2, 292–299. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, L.; Zalar, P.; Gunde-Cimerman, N.; Azua-Bustos, A.; Sterflinger, K.; Piñar, G. Aspergillus atacamensis and A. salisburgensis: Two new halophilic species from hypersaline/arid habitats with a phialosimplex-like morphology. Extremophiles 2017, 21, 755–773. [Google Scholar] [CrossRef] [PubMed]
- Dastogeer, K.M.G.; Li, H.; Sivasithamparam, K.; Wylie, S.J. In vitro salt and thermal tolerance of fungal endophytes of Nicotiana spp. growing in arid regions of north-western Australia. Arch. Phytopathol. Plant Prot. 2018, 51, 602–616. [Google Scholar] [CrossRef]
- Kunčič, M.K.; Kogej, T.; Drobne, D.; Gunde-Cimerman, N. Morphological response of the halophilic fungal genus Wallemia to high salinity. Appl. Environ. Microbiol. 2010, 76, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikram, M.; Ali, N.; Jan, G.; Jan, F.G.; Rahman, I.U.; Iqbal, A.; Hamayun, M. IAA producing fungal endophyte Penicillium roqueforti Thom., enhances stress tolerance and nutrients uptake in wheat plants grown on heavy metal contaminated soils. PLoS ONE 2018, 13, e0208150. [Google Scholar] [CrossRef]
- Kuswinanti, T.; Syam’un, E.; Masniawati, A. The potency of endophytic fungal isolates collected from local aromatic rice as indole acetic acid (IAA) producer. Procedia Food Sci. 2015, 3, 96–103. [Google Scholar]
- Mehmood, A.; Hussain, A.; Irshad, M.; Hamayun, M.; Iqbal, A.; Khan, N. In vitro production of IAA by endophytic fungus Aspergillus awamori and its growth promoting activities in Zea mays. Symbiosis 2019, 77, 225–235. [Google Scholar] [CrossRef]
- Ahmad, F.; Ahmad, I.; KHAN, M.S. Indole acetic acid production by the indigenous isolates of Azotobacter and fluorescent Pseudomonas in the presence and absence of tryptophan. Turk. J. Biol. 2005, 29, 29–34. [Google Scholar]
- Turbat, A.; Rakk, D.; Vigneshwari, A.; Kocsubé, S.; Thu, H.; Szepesi, Á.; Bakacsy, L.; Škrbić, B.D.; Jigjiddorj, E.-A.; Vágvölgyi, C.; et al. Characterization of the plant growth-promoting activities of endophytic fungi isolated from sophora flavescens. Microorganisms 2020, 8, 683. [Google Scholar] [CrossRef]
- Fouda, A.H.; Hassan, S.E.-D.; Eid, A.M.; Ewais, E.E.-D. Biotechnological applications of fungal endophytes associated with medicinal plant Asclepias sinaica (Bioss.). Ann. Agric. Sci. 2015, 60, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, C.E.; Gundel, P.E.; Helander, M.; Saikkonen, K. Endophytic mediation of reactive oxygen species and antioxidant activity in plants: A review. Fungal Divers. 2012, 54, 1–10. [Google Scholar] [CrossRef]
- Cui, J.-L.; Guo, T.-T.; Ren, Z.-X.; Zhang, N.-S.; Wang, M.-L. Diversity and antioxidant activity of culturable endophytic fungi from alpine plants of Rhodiola crenulata, R. angusta, and R. sachalinensis. PLoS ONE 2015, 10, e0118204. [Google Scholar] [CrossRef] [Green Version]
- Rahmawati, S.I.; Izzati, F.N.; Hapsari, Y.; Septiana, E.; Rachman, F.; Simanjuntak, P. Endophytic microbes and antioxidant activities of secondary metabolites from mangroves Avicennia marina and Xylocarpus granatum. IOP Conf. Ser. Earth Environ. Sci. 2019, 278, 12065. [Google Scholar] [CrossRef]
- Ravindran, C.; Naveenan, T.; Varatharajan, G.R.; Rajasabapathy, R.; Meena, R.M. Antioxidants in mangrove plants and endophytic fungal associations. Bot. Mar. 2012, 55, 269–279. [Google Scholar] [CrossRef]
- Khalil, A.M.A.; Abdelaziz, A.M.; Khaleil, M.M.; Hashem, A.H. Fungal endophytes from leaves of Avicennia marina growing in semi-arid environment as a promising source for bioactive compounds. Lett. Appl. Microbiol. 2020, 72, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Schillaci, M.; Walker, R.; Smith, P.M.C.; Watt, M.; Roessner, U. Alleviation of salinity stress in plants by endophytic plant-fungal symbiosis: Current knowledge, perspectives and future directions. Plant Soil 2020, 461, 219–244. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, Y.; Liu, C.; Chen, F.; Ge, H.; Tian, F.; Yang, T.; Ma, K.; Zhang, Y. Trichoderma harzianum mitigates salt stress in cucumber via multiple responses. Ecotoxicol. Environ. Saf. 2019, 170, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Firáková, S.; Šturdiková, M.; Múčková, M. Bioactive secondary metabolites produced by microorganisms associated with plants. Biologia 2007, 62, 251–257. [Google Scholar] [CrossRef]
- Rashmi, M.; Kushveer, J.S.; Sarma, V.V. Secondary metabolites produced by endophytic fungi from marine environments. In Endophytes and Secondary Metabolites; Jha, S., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–36. ISBN 978-3-319-76900-4. [Google Scholar]
- Yasmeen, R.; Siddiqui, Z.S. Physiological responses of crop plants against Trichoderma harzianum in saline environment. Acta Bot. Croat. 2017, 76, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Ikram, M.; Ali, N.; Jan, G.; Iqbal, A.; Hamayun, M.; Jan, F.G.; Hussain, A.; Lee, I.-J.; Ikram, M.; Ali, N.; et al. Trichoderma reesei improved the nutrition status of wheat crop under salt stress. J. Plant Interact. 2019, 14, 590–602. [Google Scholar] [CrossRef] [Green Version]
- Jan, F.G.; Hamayun, M.; Hussain, A.; Iqbal, A.; Jan, G.; Khan, S.A.; Khan, H.; Lee, I.-J.; Iqbal, A.; Jan, G.; et al. A promising growth promoting Meyerozyma caribbica from Solanum xanthocarpum alleviated stress in maize plants. Biosci. Rep. 2019, 39, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Gan, Y.; Xu, B. Application of plant-growth-promoting fungi Trichoderma longibrachiatum T6 enhances tolerance of wheat to salt stress through improvement of antioxidative defense system and gene expression. Front. Plant Sci. 2016, 7, 1405. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.S.; Kim, K.-S.; Hamayun, M.; Kim, Y. Silicon confers soybean resistance to salinity stress through regulation of reactive oxygen and reactive nitrogen species. Front. Plant Sci. 2019, 10, 1725. [Google Scholar] [CrossRef]
- Pandya, N.D.; Desai, P.V.; Jadhav, H.P.; Sayyed, R.Z. Plant growth promoting potential of Aspergillus sp. NPF7, isolated from wheat rhizosphere in South Gujarat, India. Environ. Sustain. 2018, 1, 245–252. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Shoresh, M.; Harman, G.E.; Mastouri, F. Induced systemic resistance and plant responses to fungal biocontrol agents. Annu. Rev. Phytopathol. 2010, 48, 21–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.L.; Hamayun, M.; Ahmad, N.; Hussain, J.; Kang, S.-M.; Kim, Y.-H.; Adnan, M.; Tang, D.-S.; Waqas, M.; Radhakrishnan, R.; et al. Salinity stress resistance offered by endophytic fungal interaction between Penicillium minioluteum LHL09 and Glycine max. J. Microbiol. Biotechnol. 2011, 21, 893–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada, B.; Aroca, R.; Barea, J.M.; Ruiz-Lozano, J.M. Native arbuscular mycorrhizal fungi isolated from a saline habitat improved maize antioxidant systems and plant tolerance to salinity. Plant Sci. 2013, 201, 42–51. [Google Scholar] [CrossRef]
- Sairam, R.K.; Rao, K.V.; Srivastava, G.C. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Aftab, T.; Khan, M.M.A.; da Silva, J.A.T.; Idrees, M.; Naeem, M. Role of salicylic acid in promoting salt stress tolerance and enhanced artemisinin production in Artemisia annua L. J. Plant Growth Regul. 2011, 30, 425–435. [Google Scholar] [CrossRef]
- Siddiqui, Z.; Cho, J.-I.; Park, S.-H.; Kwon, T.-R.; Lee, G.-S.; Jeong, M.-J.; Kim, K.-W.; Lee, S.-K.; Park, S.-C. Phenotyping of rice in salt stress environment using high-throughput infrared imaging. Acta Bot. Croat. 2014, 73, 312–321. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Salokhe, V.M. Rice root growth and physiological responses to SRI water management and implications for crop productivity. Paddy Water Environ. 2011, 9, 41–52. [Google Scholar] [CrossRef]
- Vafadar, F.; Amooaghaie, R.; Otroshy, M. Effects of plant-growth-promoting rhizobacteria and arbuscular mycorrhizal fungus on plant growth, stevioside, NPK, and chlorophyll content of Stevia rebaudiana. J. Plant Interact. 2014, 9, 128–136. [Google Scholar] [CrossRef]
- Rawat, L.; Singh, Y.; Shukla, N.; Kumar, J. Alleviation of the adverse effects of salinity stress in wheat (Triticum aestivum L.) by seed biopriming with salinity tolerant isolates of Trichoderma harzianum. Plant Soil 2011, 347, 387. [Google Scholar] [CrossRef]
- Zhang, F.; Yuan, J.; Yang, X.; Cui, Y.; Chen, L.; Ran, W.; Shen, Q. Putative Trichoderma harzianum mutant promotes cucumber growth by enhanced production of indole acetic acid and plant colonization. Plant Soil 2013, 368, 433–444. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Al Huqail, A.A.; Egamberdieva, D. Alleviation of abiotic salt stress in Ochradenus baccatus (Del.) by Trichoderma hamatum (Bonord.) Bainier. J. Plant Interact. 2014, 9, 857–868. [Google Scholar] [CrossRef]
- Younesi, O.; Moradi, A. Effects of plant growth-promoting rhizobacterium (PGPR) and arbuscular mycorrhizal fungus (AMF) on antioxidant enzyme activities in salt-stressed bean (Phaseolus vulgaris L.). Agriculture 2014, 60, 10–21. [Google Scholar] [CrossRef] [Green Version]
- Bouzouina, M.; Kouadria, R.; Lotmani, B. Fungal endophytes alleviate salt stress in wheat in terms of growth, ion homeostasis and osmoregulation. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Goicoechea, N.; Merino, S.; Sánchez-Díaz, M. Arbuscular mycorrhizal fungi can contribute to maintain antioxidant and carbon metabolism in nodules of Anthyllis cytisoides L. subjected to drought. J. Plant Physiol. 2005, 162, 27–35. [Google Scholar] [CrossRef]
- Hamayun, M.; Hussain, A.; Iqbal, A.; Khan, S.A.; Lee, I.-J. Endophytic fungus Aspergillus japonicus mediates host plant growth under normal and heat stress conditions. Biomed. Res. Int. 2018. [Google Scholar] [CrossRef]
- Xie, Y.; Luo, H.; Du, Z.; Hu, L.; Fu, J. Identification of cadmium-resistant fungi related to Cd transportation in bermudagrass [Cynodon dactylon (L.) Pers.]. Chemosphere 2014, 117, 786–792. [Google Scholar] [CrossRef]
- Li, X.; Han, S.; Wang, G.; Liu, X.; Amombo, E. The fungus aspergillus aculeatus enhances salt-stress tolerance, metabolite accumulation, and improves forage quality in perennial ryegrass. Front. Microbiol. 2017, 8, 1664. [Google Scholar] [CrossRef] [Green Version]
- Robert-Seilaniantz, A.; Navarro, L.; Bari, R.; Jones, J.D.G. Pathological hormone imbalances. Curr. Opin. Plant Biol. 2007, 10, 372–379. [Google Scholar] [CrossRef]
- Manchanda, G.; Garg, N. Alleviation of salt-induced ionic, osmotic and oxidative stresses in Cajanus cajan nodules by AM inoculation. Plant Biosyst. 2011, 145, 88–97. [Google Scholar] [CrossRef]
- Gundel, P.E.; Martínez-Ghersa, M.A.; Omacini, M.; Cuyeu, R.; Pagano, E.; Ríos, R.; Ghersa, C.M. Mutualism effectiveness and vertical transmission of symbiotic fungal endophytes in response to host genetic background. Evol. Appl. 2012, 5, 838–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagheri, A.A.; Saadatmand, S.; Niknam, V.; Nejadsatari, T.; Babaeizad, V. Effect of endophytic fungus, Piriformospora indica, on growth and activity of antioxidant enzymes of rice (Oryza sativa L.) under salinity stress. Int. J. Adv. Biol. Biomed. Res. 2013, 1, 1337–1350. [Google Scholar]
- Pessarakli, M.; Huber, J.T. Biomass production and protein synthesis by alfalfa under salt stress. J. Plant Nutr. 1991, 14, 283–293. [Google Scholar] [CrossRef]
- Mohamed, H.I.; Gomaa, E.Z. Effect of plant growth promoting Bacillus subtilis and Pseudomonas fluorescens on growth and pigment composition of radish plants (Raphanus sativus) under NaCl stress. Photosynthetica 2012, 50, 263–272. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Kang, S.; Baek, I.; Lee, I. Characterization of plant growth-promoting traits of Penicillium species against the effects of high soil salinity and root disease. J. Plant Interact. 2014, 9, 754–762. [Google Scholar] [CrossRef]
- Devi, R.G.; Pandiyarajan, V.; Gurusaravanan, P. Alleviating effect of IAA on salt stressed Phaseolus mungo (L.) with reference to growth and biochemical characteristics. Recent Res. Sci. Technol. 2012, 4, 22–24. [Google Scholar]
- Bertrand, A.; Bipfubusa, M.; Dhont, C.; Chalifour, F.-P.; Drouin, P.; Beauchamp, C.J. Rhizobial strains exert a major effect on the amino acid composition of alfalfa nodules under NaCl stress. Plant Physiol. Biochem. 2016, 108, 344–352. [Google Scholar] [CrossRef]
- Sampangi-Ramaiah, M.H.; Dey, P.; Jambagi, S.; Kumari, M.M.V.; Oelmüller, R.; Nataraja, K.N.; Ravishankar, K.V.; Ravikanth, G.; Shaanker, R.U. An endophyte from salt-adapted Pokkali rice confers salt-tolerance to a salt-sensitive rice variety and targets a unique pattern of genes in its new host. Sci. Rep. 2020, 10, 3237. [Google Scholar] [CrossRef]
- Baltruschat, H.; Fodor, J.; Harrach, B.D.; Niemczyk, E.; Barna, B.; Gullner, G.; Janeczko, A.; Kogel, K.K.; Schäfer, P.; Schwarczinger, I.; et al. Salt tolerance of barley induced by the root endophyte Piriformospora indica is associated with a strong increase in antioxidants. New Phytol. 2008, 180, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Dardanelli, M.S.; de Cordoba, F.J.F.; Espuny, M.R.; Carvajal, M.A.R.; Díaz, M.E.S.; Serrano, A.M.G.; Okon, Y.; Megías, M. Effect of Azospirillum brasilense coinoculated with Rhizobium on Phaseolus vulgaris flavonoids and Nod factor production under salt stress. Soil Biol. Biochem. 2008, 40, 2713–2721. [Google Scholar] [CrossRef]
- Kohler, J.; Hernández, J.A.; Caravaca, F.; Roldán, A. Induction of antioxidant enzymes is involved in the greater effectiveness of a PGPR versus AM fungi with respect to increasing the tolerance of lettuce to severe salt stress. Environ. Exp. Bot. 2009, 65, 245–252. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.N. The multifarious PGPR Serratia marcescens CDP-13 augments induced systemic resistance and enhanced salinity tolerance of wheat (Triticum aestivum L.). PLoS ONE 2016, 11, e0155026. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.J.; Xu, W.; Liu, A.; Chen, S. Endogenous melatonin deficiency aggravates high temperature-induced oxidative stress in Solanum lycopersicum L. Environ. Exp. Bot. 2019, 161, 303–311. [Google Scholar] [CrossRef]
- Waller, F.; Achatz, B.; Baltruschat, H.; Fodor, J.; Becker, K.; Fischer, M.; Heier, T.; Hückelhoven, R.; Neumann, C.; Von Wettstein, D.; et al. The endophytic fungus Piriformospora indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield. Proc. Natl. Acad. Sci. USA 2005, 102, 13386–13391. [Google Scholar] [CrossRef] [Green Version]
- Jogawat, A.; Saha, S.; Bakshi, M.; Dayaman, V.; Kumar, M.; Dua, M.; Varma, A.; Oelmüller, R.; Tuteja, N.; Johri, A.K. Piriformospora indica rescues growth diminution of rice seedlings during high salt stress. Plant Signal. Behav. 2013, 8, e26891. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, P.; Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; John, R.; Egamberdieva, D.; Gucel, S. Role of Trichoderma harzianum in mitigating NaCl stress in Indian mustard (Brassica juncea L) through antioxidative defense system. Front. Plant Sci. 2015, 6, 868. [Google Scholar] [CrossRef] [Green Version]
- Abdel Latef, A.A.H.; Omer, A.M.; Badawy, A.A.; Osman, M.S.; Ragaey, M.M. Strategy of salt tolerance and interactive impact of Azotobacter chroococcum and/or Alcaligenes faecalis inoculation on canola (Brassica napus L.) plants grown in saline soil. Plants 2021, 10, 110. [Google Scholar] [CrossRef]
- Darvishan, M.; Tohidi-Moghadam, H.R.; Zahedi, H. The effects of foliar application of ascorbic acid (vitamin C) on physiological and biochemical changes of corn (Zea mays L) under irrigation withholding in different growth stages. Maydica 2013, 58, 195–200. [Google Scholar]
- Yousuf, P.Y.; Ahmad, A.; Hemant Ganie, A.H.; Aref, I.M.; Iqbal, M. Potassium and calcium application ameliorates growth and oxidative homeostasis in salt-stressed Indian mustard (Brassica juncea) plants. Pak. J. Bot 2015, 47, 1629–1639. [Google Scholar]
- Nessim, A.; Kasim, W. Physiological impact of seed priming with CaCl2 or carrot root extract on lupinus termis plants fully grown under salinity stress. Egypt. J. Bot. 2019, 59, 763–777. [Google Scholar] [CrossRef]
- Ripa, F.A.; Cao, W.; Tong, S.; Sun, J. Assessment of plant growth promoting and abiotic stress tolerance properties of wheat endophytic fungi. Biomed. Res. Int. 2019, 2019, 6105865. [Google Scholar] [CrossRef]
- Zhang, Z.; Mao, B.; Li, H.; Zhou, W.; Takeuchi, Y.; Yoneyama, K. Effect of salinity on physiological characteristics, yield and quality of microtubers in vitro in potato. Acta Physiol. Plant 2005, 27, 481–489. [Google Scholar] [CrossRef]
- Stavridou, E.; Hastings, A.; Webster, R.J.; Robson, P.R.H. The impact of soil salinity on the yield, composition and physiology of the bioenergy grass Miscanthus × giganteus. Gcb Bioenergy 2017, 9, 92–104. [Google Scholar] [CrossRef] [Green Version]
- Zein, F.I.; Gaiza, E.A.; EL-Sanafawy, H.M.; Talha, N.I. Effect of specific ions, salinity and alkalinity on yield and quality of some Egyptian cotton genotypes. Egypt. J. Soil Sci. 2020, 60, 183–194. [Google Scholar] [CrossRef]
- Mohamed, A.S.; Mahmoud, M.G.; Sharaf, A.E.-M.M. Interactive effect of Trichoderma viride on broad bean (cv. Vicia faba L.) genotypes grown under different salinity stress conditions. Int. J. Ecotoxicol. Ecobiol. 2016, 1, 141–151. [Google Scholar]
- Khalid, A.; Aftab, F. Effect of exogenous application of IAA and GA 3 on growth, protein content, and antioxidant enzymes of Solanum tuberosum L. grown in vitro under salt stress. Cell. Dev. Biol. 2020, 56, 377–389. [Google Scholar]
- Hashem, A.H.; Hasanin, M.S.; Khalil, A.M.A.; Suleiman, W.B. Eco-green conversion of watermelon peels to single cell oils using a unique oleaginous fungus: Lichtheimia corymbifera AH13. Waste Biomass Valorization 2019, 11, 5721–5732. [Google Scholar] [CrossRef]
- Mohite, B. Isolation and characterization of indole acetic acid ( IAA ) producing bacteria from rhizospheric soil and its effect on plant growth. J. Soil Sci. Plant Nutr. 2013, 13, 638–649. [Google Scholar] [CrossRef]
- De Sousa, E.S.O.; Cortez, A.C.A.; de Melhem, M.S.C.; Frickmann, H.; de Souza, J.V.B. Factors influencing susceptibility testing of antifungal drugs: A critical review of document M27-A4 from the Clinical and Laboratory Standards Institute (CLSI). Braz. J. Microbiol. 2020, 51, 1791–1800. [Google Scholar] [CrossRef] [PubMed]
- Vernon, L.P.; Seely, G.R. The Chlorophylls; Academic Press: New York, NY, USA, 1966. [Google Scholar]
- Umbreit, W.W.; Burris, R.H.; Stauffer, J.F. Manometric Techniques: A Manual Describing Methods Applicable to the Study of Tissue Metabolism; Burgess Publishing Co.: Minneapolis, MN, USA, 1964. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent J Biol Chem 193: 265–275. Find. this Artic. Online 1951, 193, 265–275. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Bergmeyer, H.U. Methods of Enzymatic Analysis, 2nd ed.; Academic Press: New York, NY, USA, 1974; Volume 1, p. 974. [Google Scholar]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Matta, A.; Dimond, A.E. Symptoms of Fusarium wilt in relation to quantity of fungus and enzyme activity in tomato stems. Phytopathology 1963, 53, 574–575. [Google Scholar]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Endophyte Name | Accession Number | Fungal Growth at Different Concentrations of NaCl g L−1 | ||||
|---|---|---|---|---|---|---|
| 50 | 100 | 150 | 200 | 250 | ||
| Aspergillus hiratsukae | MT089951 | ++ | + | − | − | − |
| Alternaria tenuissima | MT089952 | +++ | ++ | + | − | − |
| Chaetomium sp. | MT089953 | ++ | + | − | − | − |
| Curvularia lunata | MT089955 | +++ | ++ | + | − | − |
| Chaetomium sp. | MT089956 | ++ | + | − | − | − |
| Chaetomium globosum | MT089957 | ++ | + | − | − | − |
| Aspergillus ochraceus | MT089958 | +++ | +++ | ++ | + | − |
| Treatment no. | Inhibition Zone (mm) (1000 µg/mL) | MIC (µg/mL) |
|---|---|---|
| 0% seawater | 0 c | Not Detected |
| 15% seawater | 17.3 ± 0.57 b | 250 |
| 30% seawater | 23.6 ± 1.15 a | 62.5 |
| Treatments | Soluble Sugars | Soluble Protein | Free Proline | ||
|---|---|---|---|---|---|
| Salinity | 0% S.W. | 93.09 ± 2.96 b | 44.89 ± 0.73 b | 0.38 ± 0.01 d | |
| 15% S.W. | 78.72 ± 0.90 d | 30.67 ± 0.29 e | 0.88 ± 0.03 b | ||
| 30% S.W. | 70.65 ± 2.37 e | 25.52 ± 0.57 f | 1.00 ± 0.02 a | ||
| Salinity + A. ochraceus | 0% S.W. | 104.08 ± 2.01 a | 73.55 ± 0.38 a | 0.39 ± 0.002 d | |
| 15% S.W. | 86.81 ± 2.46 bc | 41.34 ± 0.32 c | 0.55 ± 0.001 c | ||
| 30% S.W. | 80.72 ± 3.76 cd | 39.00 ± 0.40 d | 0.59 ± 0.003 c | ||
| Treatments | Spike Length (cm) | Spike Weight (g plant−1) | Number of Grains | Grains Weight (g plant−1) | 1000 Grains Weight (g plant−1) | |
|---|---|---|---|---|---|---|
| Salinity | 0% S.W. | 15.93 ± 0.17 a | 2.95 ± 0.25 ab | 28.33 ± 1.02 a | 2.18 ± 0.15 ab | 73.33 ± 1.20 ab |
| 15% S.W. | 12.7 ± 0.24 bc | 1.82 ± 0.23 d | 16.66 ± 1.20 b | 1.34 ± 0.17 c | 56.66 ± 1.20 c | |
| 30% S.W. | 9.96 ± 0.44 d | 1.33 ± 0.23 d | 9.33 ± 1.45 c | 1.09 ± 0.20 c | 34.33 ± 2.40 e | |
| Salinity + A. ochraceus | 0% S.W. | 16.3 ± 0.62 a | 3.64 ± 0.15 a | 32.33 ± 1.47 a | 2.7 ± 0.04 a | 78.66 ± 1.85 a |
| 15% S.W. | 15.16 ± 0.57 ab | 2.74 ± 0.28 bc | 26.66 ± 1.21 a | 2.12 ± 0.21 b | 68.33 ± 2.84 b | |
| 30% S.W. | 12.5 ± 1.04 c | 1.98 ± 0.41 cd | 14.33 ± 1.51 bc | 1.46 ± 0.06 c | 45 ± 1.73 e | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badawy, A.A.; Alotaibi, M.O.; Abdelaziz, A.M.; Osman, M.S.; Khalil, A.M.A.; Saleh, A.M.; Mohammed, A.E.; Hashem, A.H. Enhancement of Seawater Stress Tolerance in Barley by the Endophytic Fungus Aspergillus ochraceus. Metabolites 2021, 11, 428. https://doi.org/10.3390/metabo11070428
Badawy AA, Alotaibi MO, Abdelaziz AM, Osman MS, Khalil AMA, Saleh AM, Mohammed AE, Hashem AH. Enhancement of Seawater Stress Tolerance in Barley by the Endophytic Fungus Aspergillus ochraceus. Metabolites. 2021; 11(7):428. https://doi.org/10.3390/metabo11070428
Chicago/Turabian StyleBadawy, Ali A., Modhi O. Alotaibi, Amer M. Abdelaziz, Mahmoud S. Osman, Ahmed M. A. Khalil, Ahmed M. Saleh, Afrah E. Mohammed, and Amr H. Hashem. 2021. "Enhancement of Seawater Stress Tolerance in Barley by the Endophytic Fungus Aspergillus ochraceus" Metabolites 11, no. 7: 428. https://doi.org/10.3390/metabo11070428