Effect of Etifoxine on Locomotor Activity and Passive Learning in Rats with Diazepam-Induced Cognitive Deficit


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
- Group 1 (Control) with an equivalent volume (0.1 mL/100 g bw) of vehicle (0.1% Tween 20 in 0.9% NaCl solution);
- Group 2 with diazepam 2.5 mg/kg bw;
- Group 3 with diazepam 2.5 mg/kg bw and etifoxine in a dose of 50 mg/kg bw.
2.3. Activity Cage Test
2.4. Model of Diazepam-Induced Amnesia
2.4.1. Step-Through Test
2.4.2. Step-Down Test
2.5. Statistical Analysis
3. Results
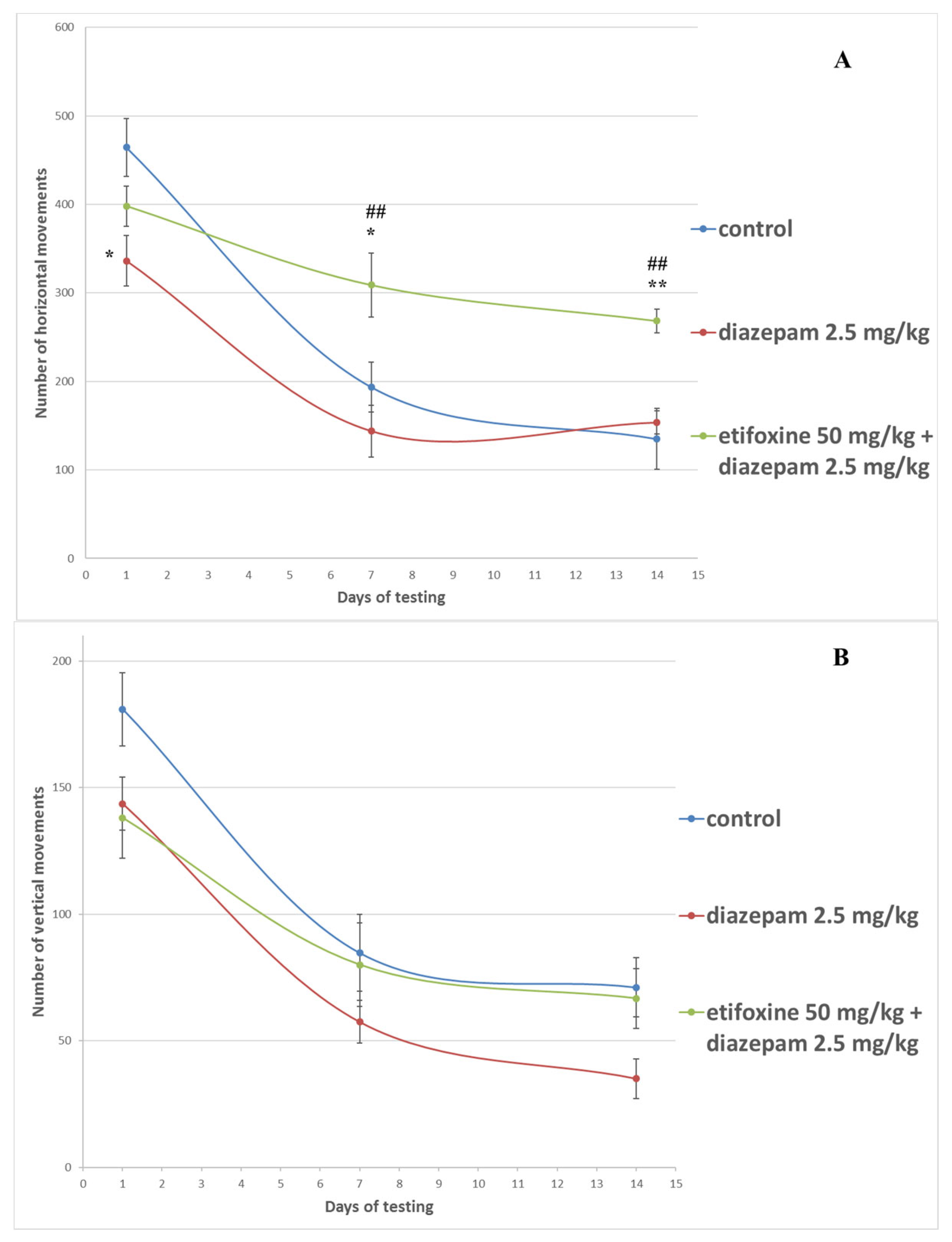
3.1. Activity Cage
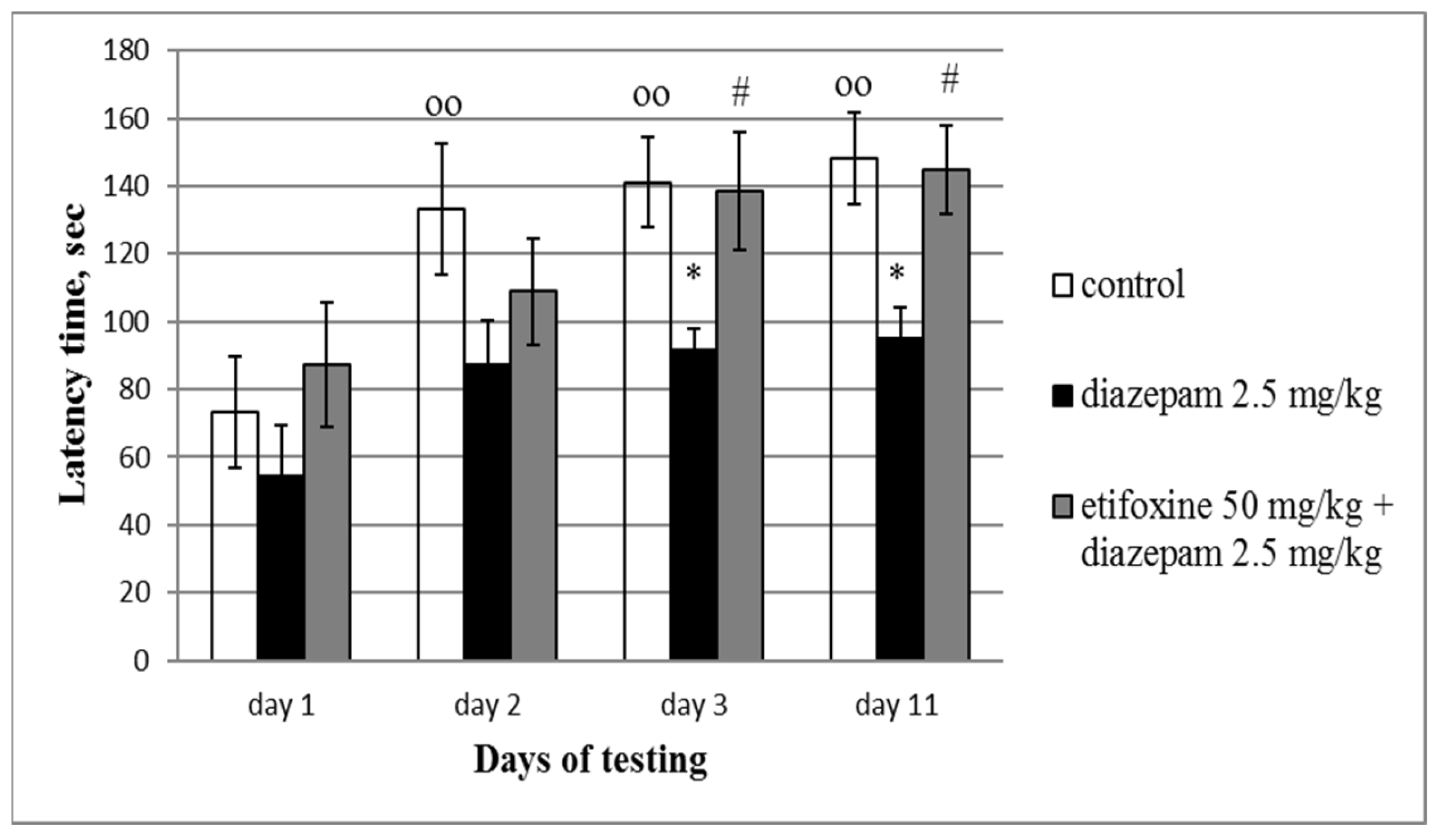
3.2. Step-Through Test
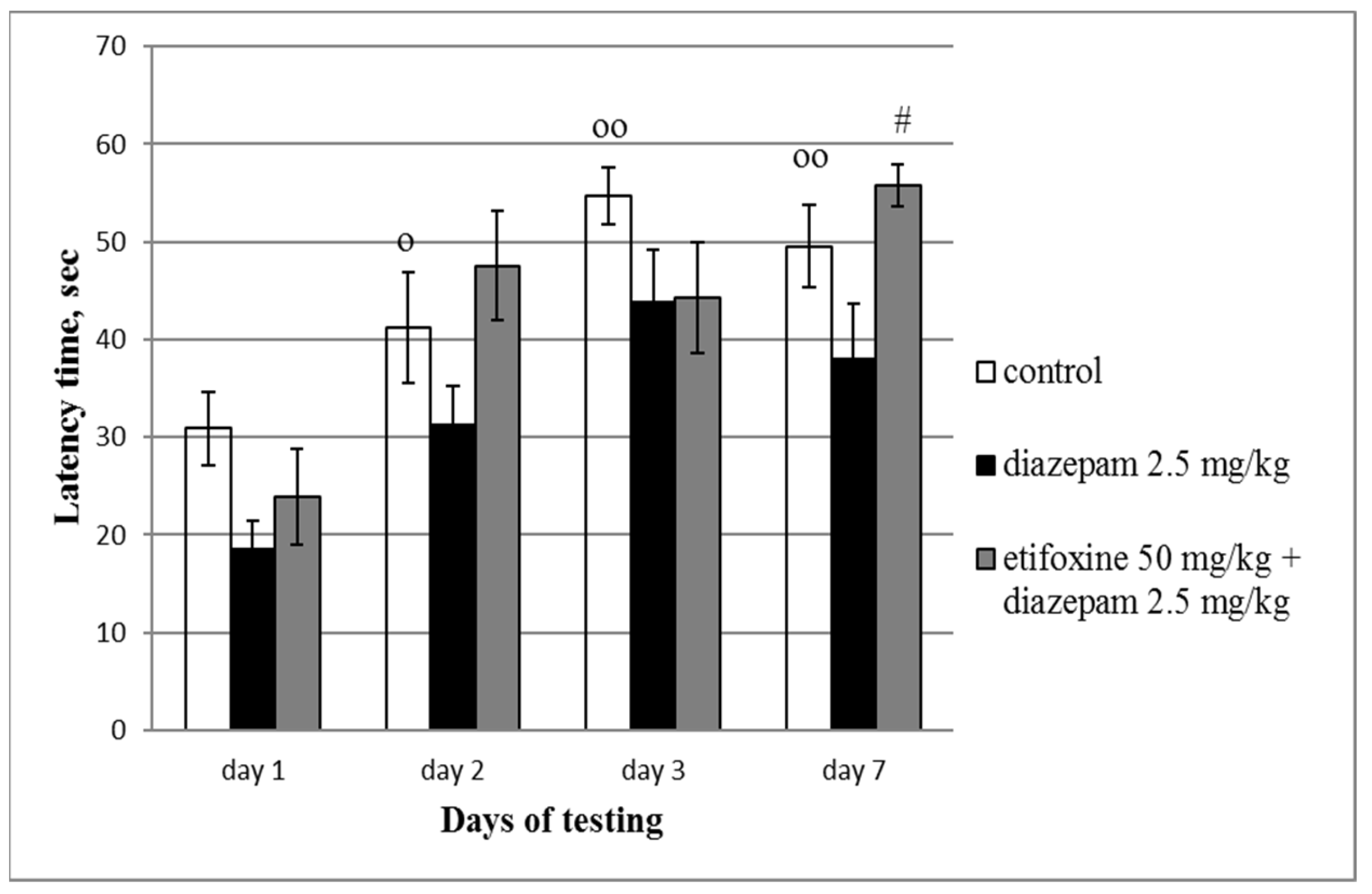
3.3. Step-Down Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Vicente, B.; Saldivia, S.; Hormazabal, N.; Bustos, C.; Rubí, P. Etifoxine is non-inferior than clonazepam for reduction of anxiety symptoms in the treatment of anxiety disorders: A randomized, double blind, non-inferiority trial. Psychopharmacology 2020, 237, 3357–3367. [Google Scholar] [CrossRef] [PubMed]
- Poisbeau, P.; Gazzo, G.; Calvel, L. Anxiolytics targeting GABAA receptors: Insights on etifoxine. World J. Biol. Psychiatry 2018, 19, S36–S45. [Google Scholar] [CrossRef] [PubMed]
- Verleye, M.; Dumas, S.; Heulard, I.; Krafft, N.; Gillardin, J.-M. Differential effects of etifoxine on anxiety-like behaviour and convulsions in BALB/cByJ and C57BL/6J mice: Any relation to overexpression of central GABAA receptor beta2 subunits? Eur. Neuropsychopharmacol. 2011, 21, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Verleye, M.; Heulard, I.; Gillardin, J.-M. The anxiolytic etifoxine protects against convulsant and anxiogenic aspects of the alcohol withdrawal syndrome in mice. Alcohol 2009, 43, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Stein, D.J. Etifoxine versus alprazolam for the treatment of adjustment disorder with anxiety: A randomized controlled trial. Adv. Ther. 2015, 32, 57–68. [Google Scholar] [CrossRef]
- Nguyen, N.; Fakra, E.; Pradel, V.; Jouve, E.; Alquier, C.; Le Guern, M.-E.; Micallef, J.; Blin, O. Efficacy of etifoxine compared to lorazepam monotherapy in the treatment of patients with adjustment disorders with anxiety: A double-blind controlled study in general practice. Hum. Psychopharmacol. Clin. Exp. 2006, 21, 139–149. [Google Scholar] [CrossRef]
- Micallef, J.; Soubrouillard, C.; Guet, F.; Le Guern, M.E.; Alquier, C.; Bruguerolle, B.; Blin, O. A double blind parallel group placebo controlled comparison of sedative and amnesic effects of etifoxine and lorazepam in healthy subjects. Fundam. Clin. Pharmacol. 2001, 15, 209–216. [Google Scholar] [CrossRef]
- Servant, D.; Graziani, P.L.; Moyse, D.; Parquet, P.J. Treatment of adjustment disorder with anxiety: Efficacy and tolerance of eti-foxine in a double blind controlled study. Encephale 1998, 24, 569–574. [Google Scholar]
- Dubrovina, N.I. GABA Receptors in the Modulation of Fear Memory Extinction. Neurosci. Behav. Physiol. 2017, 47, 573–584. [Google Scholar] [CrossRef]
- Kokova, V.Y.; Zagorchev, P.I.; Apostolova, E.G.; Peychev, L.P. Etifoxine does not impair muscle tone and motor function in rats as assessed by in vivo and in vitro methods. Gen. Physiol. Biophys. 2020, 39, 179–186. [Google Scholar] [CrossRef]
- Kokova, V.; Apostolova, E.; Peychev, L. Effects of etifoxine on learning and memory of intact rats. Trakia J. Sci. 2015, 13, 40–44. [Google Scholar] [CrossRef]
- Kokova, V.; Apostolova, E.; Peychev, L. P.1.j.004 Effects of etifoxine on passive avoidance tests and locomotor activity in rats. Eur. Neuropsychopharmacol. 2015, 25 (Suppl. S1), S334–S335. [Google Scholar] [CrossRef]
- Kokova, V.; Apostolova, E. Effect of etifoxine on learning and memory processes in rats with diazepam-induced amnesia. Sci. Rep. Compet. Sess. Sci. Youth 2016, 1, 399–403. [Google Scholar]
- Georgieva-Kotetarova, M.T.; Kostadinova, I. Effect of Atorvastatin and Rosuvastatin on Learning and Memory in Rats with Diazepam-Induced Amnesia. Folia Med. 2013, 55, 58–65. [Google Scholar] [CrossRef]
- Khan, Z.; Ghosh, A. Possible modulation of neurobehavioural patterns by anxiolytics drugs. Int. J. Pharm. Sci. 2010, 1, 457–464. [Google Scholar]
- Girard, P.H.; Pansart, Y.; Gillardin, J.M. Preventive and curative effects of etifoxine in a rat model of brain oedema. Clin. Exp. Pharmacol. 2009, 36, 655–661. [Google Scholar] [CrossRef]
- Shehadeh, M.; Palzur, E.; Apel, L.; Soustiel, J.F. Reduction of Traumatic Brain Damage by Tspo Ligand Etifoxine. Int. J. Mol. Sci. 2019, 20, 2639. [Google Scholar] [CrossRef]
- Palzur, E.; Edelman, D.; Sakas, R.; Soustiel, J.F. Etifoxine Restores Mitochondrial Oxidative Phosphorylation and Improves Cognitive Recovery Following Traumatic Brain Injury. Int. J. Mol. Sci. 2021, 22, 12881. [Google Scholar] [CrossRef]
- Simon-O’brien, E.; Gauthier, D.; Riban, V.; Verleye, M. Etifoxine improves sensorimotor deficits and reduces glial activation, neuronal degeneration, and neuroinflammation in a rat model of traumatic brain injury. J. Neuroinflamm. 2016, 13, 203. [Google Scholar] [CrossRef]
- Deplanque, D.; Machuron, F.; Waucquier, N.; Jozefowicz, E.; Duhem, S.; Somers, S.; Colin, O.; Duhamel, A.; Bordet, R. Etifoxine impairs neither alertness nor cognitive functions of the elderly: A randomized, double-blind, placebo-controlled crossover study. Eur. Neuropsychopharmacol. 2018, 28, 925–932. [Google Scholar] [CrossRef]
- Tian, Q.; Yang, X.; Du, J.; Huang, H.; Liu, W.; Zhao, P. Translocator Protein Ligand Etifoxine Attenuates MPTP-Induced Neurotoxicity. Front. Mol. Neurosci. 2022, 15, 850904. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ma, L.; Guo, W.-Z.; Jiao, L.-B.; Zhao, H.-Y.; Ma, Y.-Q.; Hao, X.-M. TSPO ligand etifoxine attenuates LPS-induced cognitive dysfunction in mice. Brain Res. Bull. 2020, 165, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Thanapreedawat, P.; Kobayashi, H.; Inui, N.; Sakamoto, K.; Kim, M.; Yoto, A.; Yokogoshi, H. GABA Affects Novel Object Recognition Memory and Working Memory in Rats. J. Nutr. Sci. Vitaminol. 2013, 59, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Green, A.; Hainsworth, A.H.; Jackson, D.M. GABA potentiation: A logical pharmacological approach for the treatment of acute ischaemic stroke. Neuropharmacology 2000, 39, 1483–1494. [Google Scholar] [CrossRef]
- Costa, C.; Leone, G.; Saulle, E.; Pisani, F.; Bernardi, G.; Calabresi, P. Coactivation of GABA A and GABA B Receptor Results in Neuroprotection During In Vitro Ischemia. Stroke 2004, 35, 596–600. [Google Scholar] [CrossRef]
- Lee, Y.; Park, Y.; Nam, H.; Lee, J.-W.; Yu, S.-W. Translocator protein (TSPO): The new story of the old protein in neuroinflammation. BMB Rep. 2020, 53, 20–27. [Google Scholar] [CrossRef]
- Longone, P.; di Michele, F.; D’Agati, E.; Romeo, E.; Pasini, A.; Rupprecht, R. Neurosteroids as neuromodulators in the treatment of anxiety disorders. Front. Endocrinol. 2011, 2, 55. [Google Scholar] [CrossRef]
- Stoffel-Wagner, B. Neurosteroid metabolism in the human brain. Eur. J. Endocrinol. 2001, 145, 669–679. [Google Scholar] [CrossRef]
- Wojtal, K.; Trojnar, M.K.; Czuczwar, S.J. Endogenous neuroprotective factors: Neurosteroids. Pharmacol. Rep. 2006, 58, 335–340. [Google Scholar]
- Frye, C.A.; Walf, A.A. Effects of progesterone administration and APPswe+PSEN1Δe9 mutation for cognitive performance of mid-aged mice. Neurobiol. Learn. Mem. 2008, 89, 17–26. [Google Scholar] [CrossRef]
- VanLandingham, J.W.; Cutler, S.M.; Virmani, S.; Hoffman, S.W.; Covey, D.F.; Krishnan, K.; Hammes, S.R.; Jamnongjit, M.; Stein, D.G. The enantiomer of progesterone acts as a molecular neuroprotectant after traumatic brain injury. Neuropharmacology 2006, 51, 1078–1085. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Chopp, M.; Stein, D.; Feit, H. Progesterone is neuroprotective after transient middle cerebral artery occlusion in male rats. Brain Res. 1996, 735, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Roof, R.L.; Duvdevani, R.; Heyburn, J.W.; Stein, D.G. Progesterone Rapidly Decreases Brain Edema: Treatment Delayed up to 24 Hours Is Still Effective. Exp. Neurol. 1996, 138, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Stein, D.G.; Hoffman, S.W. Estrogen and progesterone as neuroprotective agents in the treatment of acute brain injuries. Pediatr. Rehabil. 2003, 6, 13–22. [Google Scholar] [CrossRef]
- Wright, D.W.; Kellermann, A.L.; Hertzberg, V.S.; Clark, P.L.; Frankel, M.; Goldstein, F.C.; Salomone, J.P.; Dent, L.L.; Harris, O.A.; Ander, D.S.; et al. ProTECT: A Randomized Clinical Trial of Progesterone for Acute Traumatic Brain Injury. Ann. Emerg. Med. 2007, 49, 391–402.e2. [Google Scholar] [CrossRef]
- Schumacher, M.; Akwa, Y.; Guennoun, R.; Robert, F.; Labombarda, F.; Désarnaud, F.; Robel, P.; De Nicola, A.F.; Baulieu, E.-E. Steroid synthesis and metabolism in the nervous system: Trophic and protective effects. J. Neurocytol. 2000, 29, 307–326. [Google Scholar] [CrossRef]
- Schumacher, M.; Guennoun, R.; Mercier, G.; Désarnaud, F.; Lacor, P.; Bénavides, J.; Ferzaz, B.; Robert, F.; Baulieu, E.E. Progesterone synthesis and myelin formation in peripheral nerves. Brain Res. Rev. 2001, 37, 343–359. [Google Scholar] [CrossRef]
- Verleye, M.; Akwa, Y.; Liere, P.; Ladurelle, N.; Pianos, A.; Eychenne, B.; Schumacher, M.; Gillardin, J.-M. The anxiolytic etifoxine activates the peripheral benzodiazepine receptor and increases the neurosteroid levels in rat brain. Pharmacol. Biochem. Behav. 2005, 82, 712–720. [Google Scholar] [CrossRef]
- Christensen, A.; Pike, C.J. TSPO ligand PK11195 improves Alzheimer-related outcomes in aged female 3xTg-AD mice. Neurosci. Lett. 2018, 683, 7–12. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kokova, V.; Apostolova, E. Effect of Etifoxine on Locomotor Activity and Passive Learning in Rats with Diazepam-Induced Cognitive Deficit. Sci. Pharm. 2023, 91, 25. https://doi.org/10.3390/scipharm91020025
Kokova V, Apostolova E. Effect of Etifoxine on Locomotor Activity and Passive Learning in Rats with Diazepam-Induced Cognitive Deficit. Scientia Pharmaceutica. 2023; 91(2):25. https://doi.org/10.3390/scipharm91020025
Chicago/Turabian StyleKokova, Vesela, and Elisaveta Apostolova. 2023. "Effect of Etifoxine on Locomotor Activity and Passive Learning in Rats with Diazepam-Induced Cognitive Deficit" Scientia Pharmaceutica 91, no. 2: 25. https://doi.org/10.3390/scipharm91020025