
Long-Term Paracetamol Treatment Impairs Cognitive Function and Brain-Derived Neurotrophic Factor in Adult Rat Brain
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drug Treatment
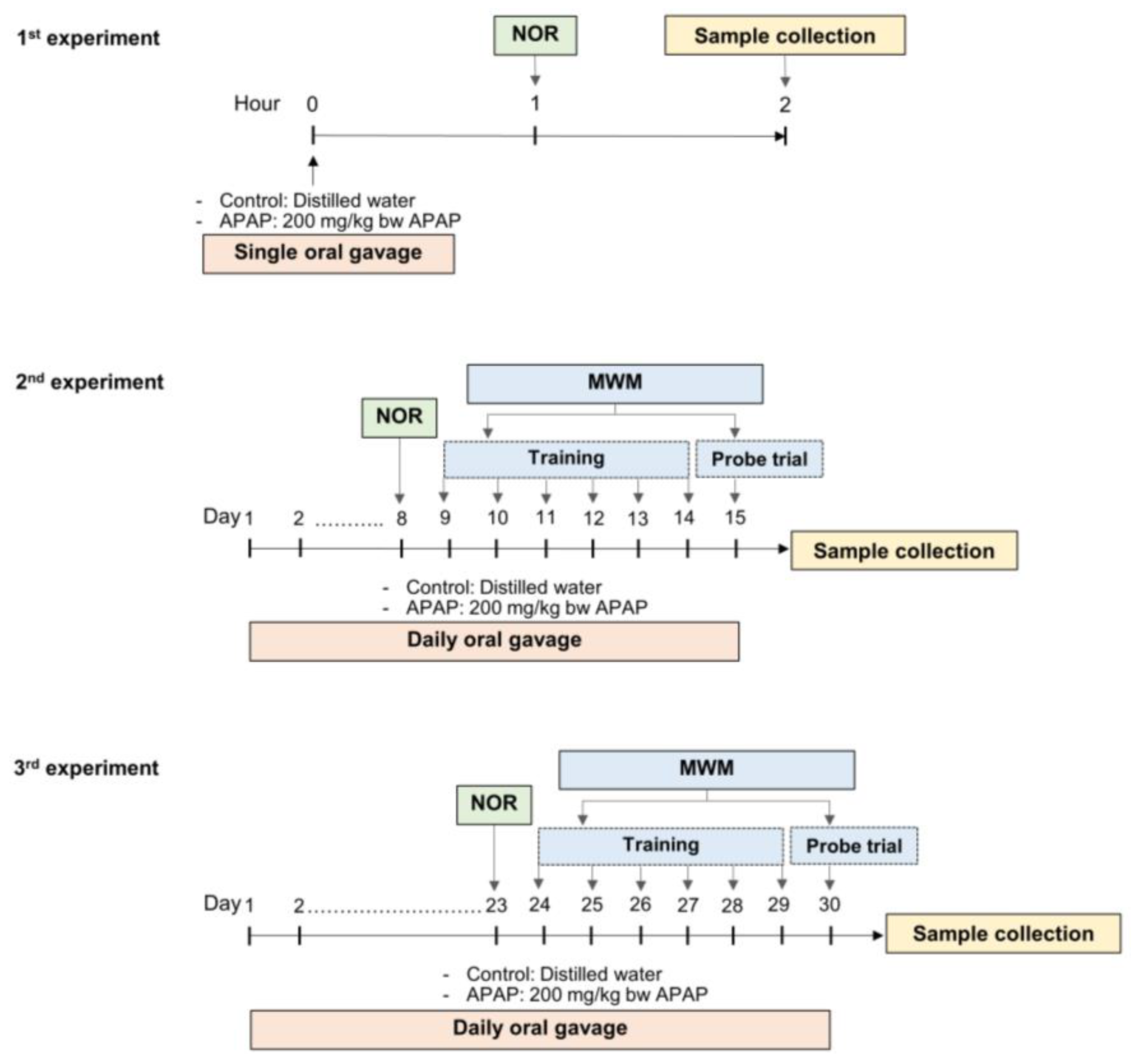
2.3. Experimental Design
2.4. Novel Object Recognition (NOR) Test
2.5. Morris Water Maze (MWM) Test
2.6. Sample Collection
2.7. Immunohistochemistry (IHC)
2.8. Western Blotting (WB)
2.9. Statistical Analysis
3. Results
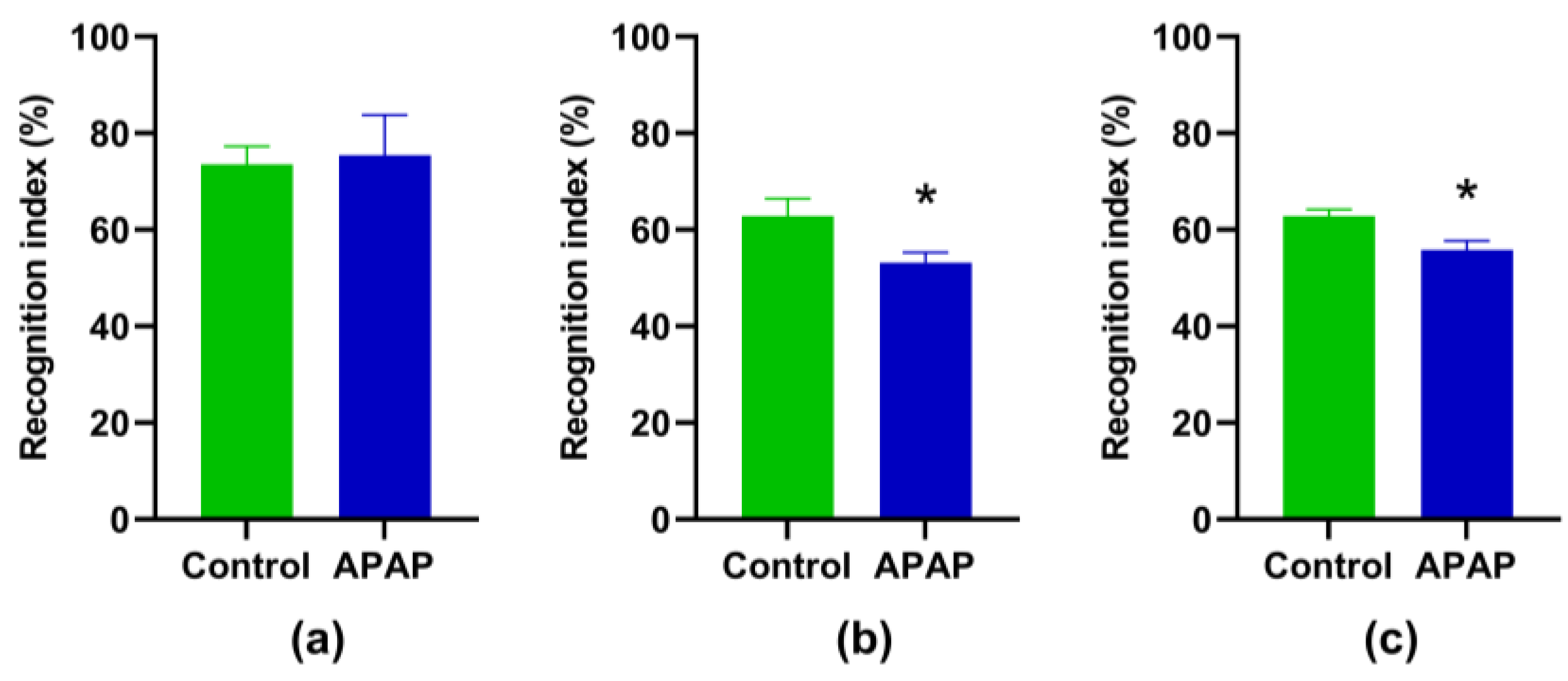
3.1. Effect of APAP Treatment on NOR Memory
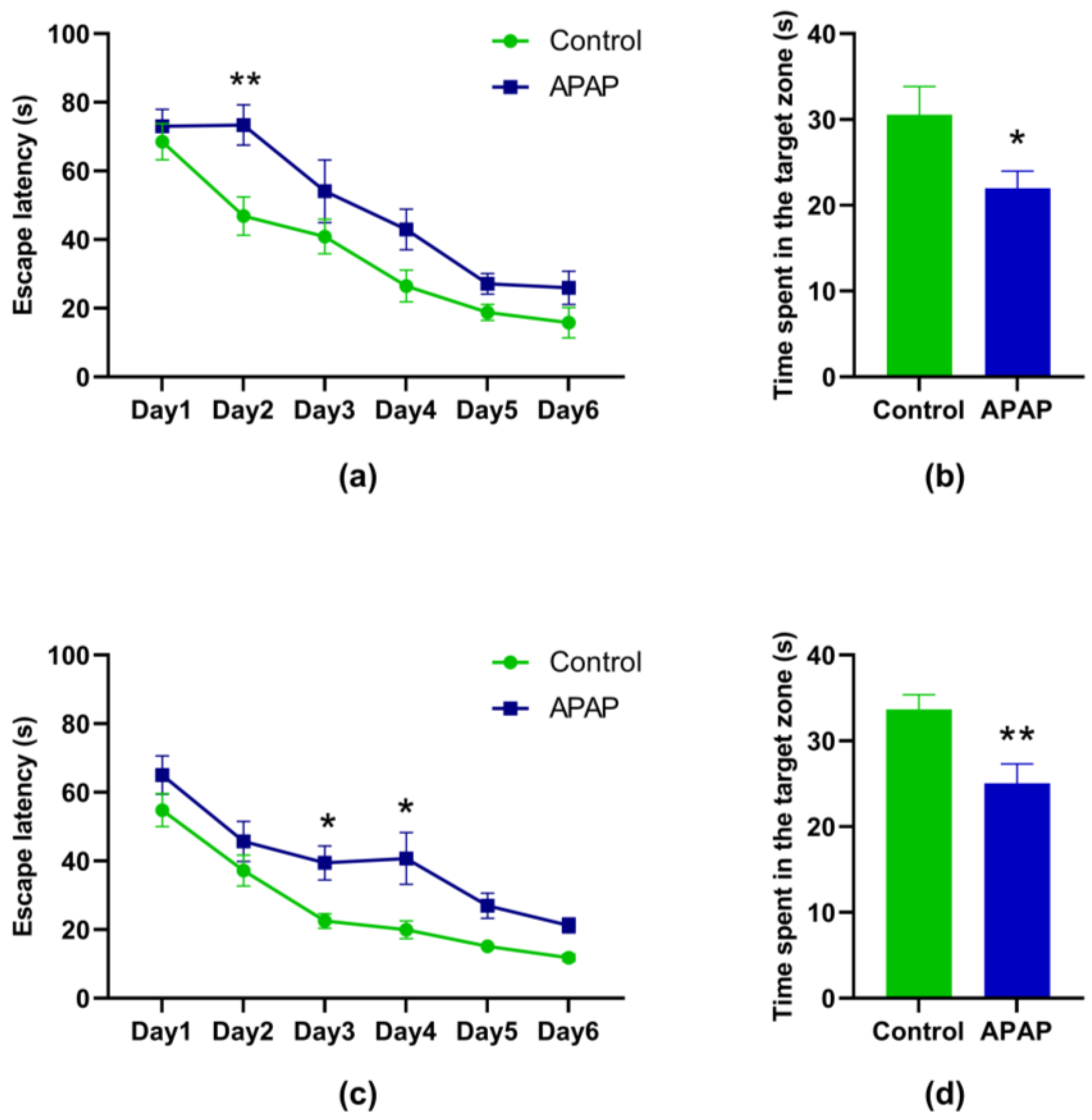
3.2. Effect of the APAP Treatment on the MWM Performance
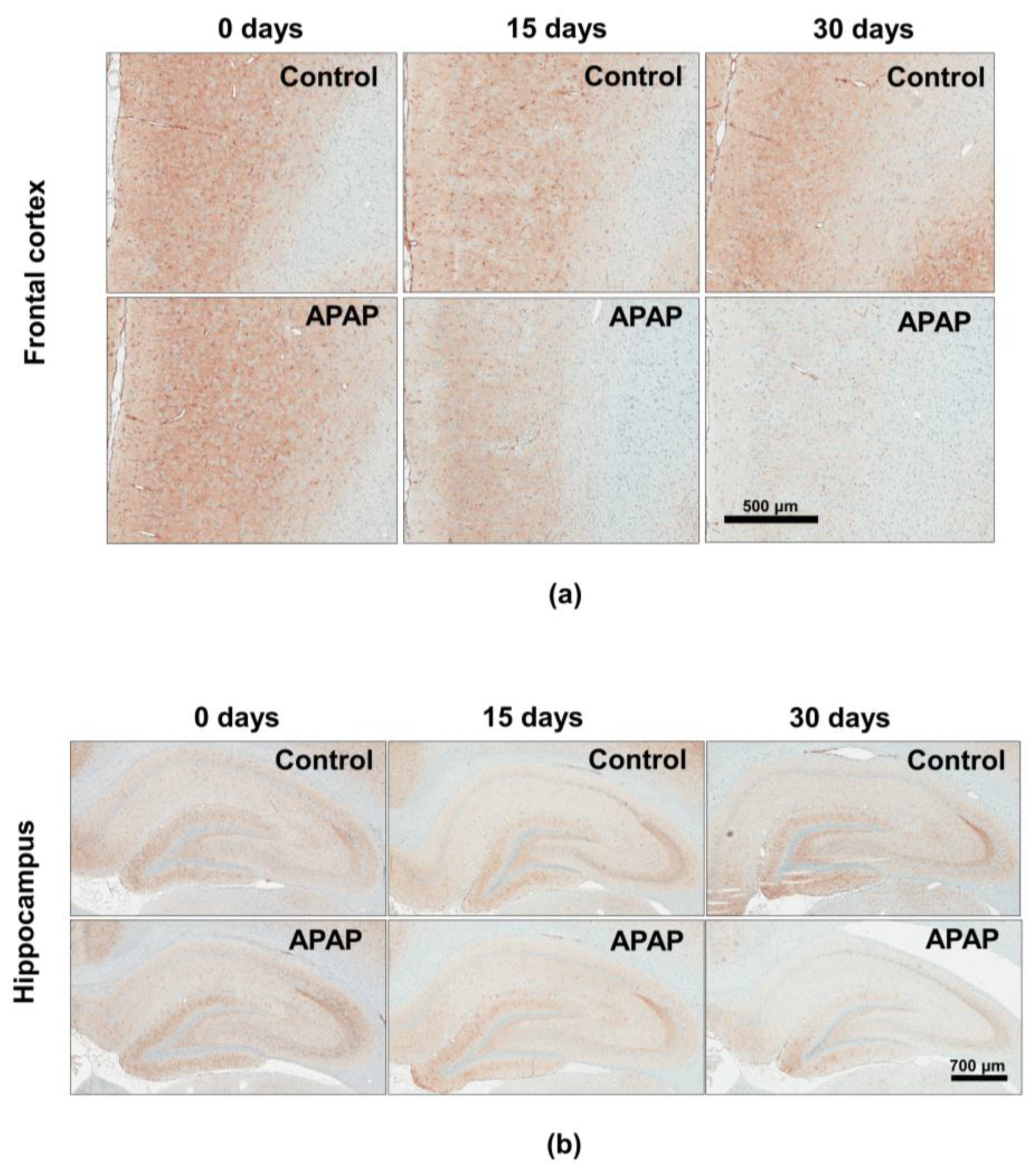
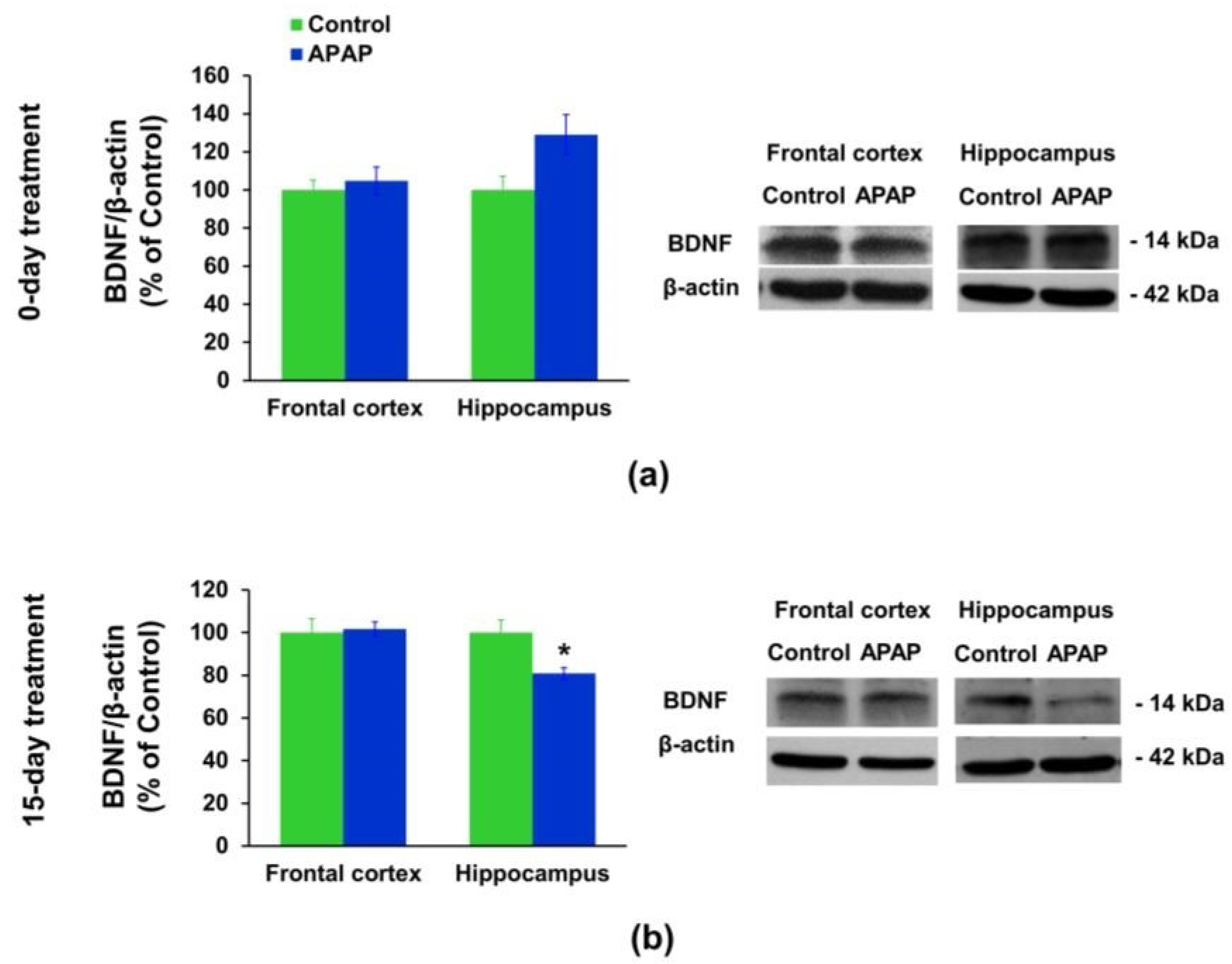
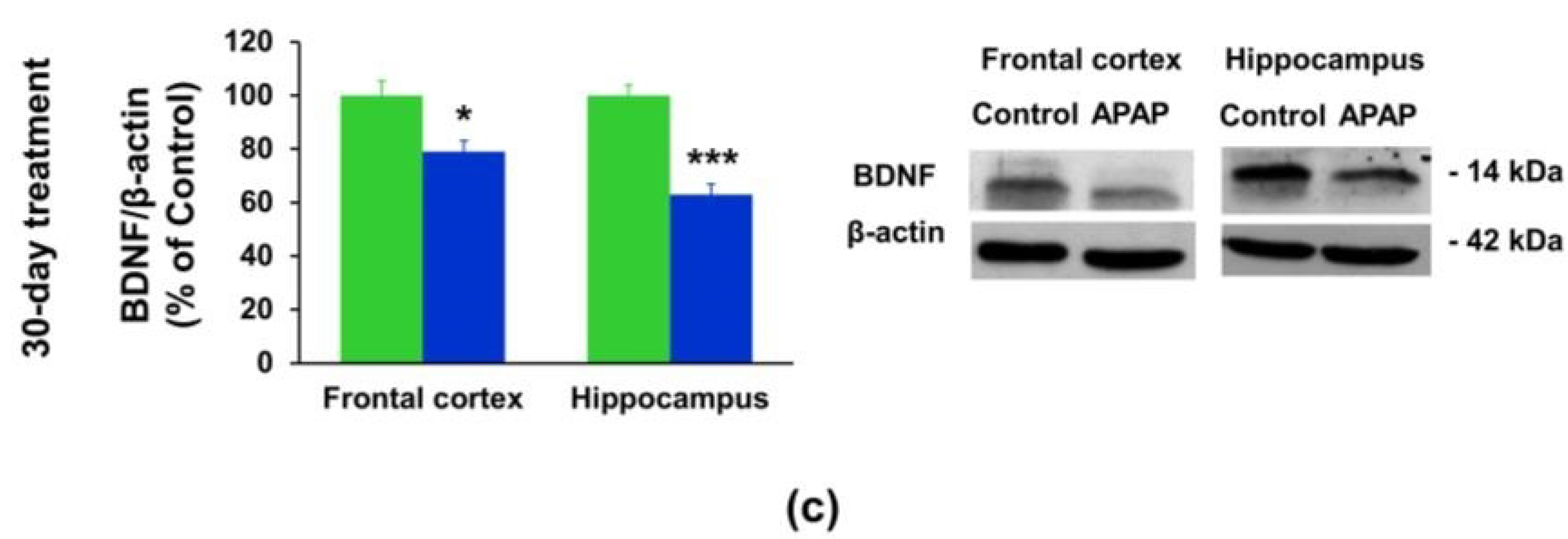
3.3. Effect of APAP Treatment on the BDNF Protein Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blecharz-Klin, K.; Piechal, A.; Pyrzanowska, J.; Joniec-Maciejak, I.; Kiliszek, P.; Widy-Tyszkiewicz, E. Paracetamol—The outcome on neurotransmission and spatial learning in rats. Behav. Brain Res. 2013, 253, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wei, H.; Pertovaara, A.; Wang, J.; Carlson, S. Anxiety- and activity-related effects of paracetamol on healthy and neuropathic rats. Pharmacol. Res. Perspect. 2018, 6, e00367. [Google Scholar] [CrossRef] [PubMed]
- Posadas, I.; Santos, P.; Blanco, A.; Munoz-Fernandez, M.; Cena, V. Acetaminophen induces apoptosis in rat cortical neurons. PLoS ONE 2010, 5, e15360. [Google Scholar] [CrossRef]
- Supornsilpchai, W.; le Grand, S.M.; Srikiatkhachorn, A. Cortical hyperexcitability and mechanism of medication-overuse headache. Cephalalgia 2010, 30, 1101–1109. [Google Scholar] [CrossRef]
- Yisarakun, W.; Supornsilpchai, W.; Chantong, C.; Srikiatkhachorn, A.; Maneesri-le Grand, S. Chronic paracetamol treatment increases alterations in cerebral vessels in cortical spreading depression model. Microvasc. Res. 2014, 94, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T.; Sato, T.; Irifune, M.; Tanaka, K.; Nakamura, N.; Nishikawa, T. Effect of acetaminophen, a cyclooxygenase inhibitor, on Morris water maze task performance in mice. J. Psychopharmacol. 2007, 21, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Lalert, L.; Ji-Au, W.; Srikam, S.; Chotipinit, T.; Sanguanrungsirikul, S.; Srikiatkhachorn, A.; Maneesri-le Grand, S. Alterations in Synaptic Plasticity and Oxidative Stress Following Long-Term Paracetamol Treatment in Rat Brain. Neurotox. Res. 2020, 37, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299. [Google Scholar] [CrossRef]
- Cirulli, F.; Berry, A.; Chiarotti, F.; Alleva, E. Intrahippocampal administration of BDNF in adult rats affects short-term behavioral plasticity in the Morris water maze and performance in the elevated plus-maze. Hippocampus 2004, 14, 802–807. [Google Scholar] [CrossRef]
- Heldt, S.A.; Stanek, L.; Chhatwal, J.P.; Ressler, K.J. Hippocampus-specific deletion of BDNF in adult mice impairs spatial memory and extinction of aversive memories. Mol. Psychiatry 2007, 12, 656–670. [Google Scholar] [CrossRef] [Green Version]
- Minichiello, L. TrkB signalling pathways in LTP and learning. Nat. Rev. Neurosci. 2009, 10, 850. [Google Scholar] [CrossRef] [PubMed]
- Montkowski, A.; Holsboer, F. Intact spatial learning and memory in transgenic mice with reduced BDNF. Neuroreport 1997, 8, 779–782. [Google Scholar] [CrossRef]
- Hofer, M.; Pagliusi, S.R.; Hohn, A.; Leibrock, J.; Barde, Y.A. Regional distribution of brain-derived neurotrophic factor mRNA in the adult mouse brain. EMBO J. 1990, 9, 2459–2464. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Nagappan, G.; Lu, Y. BDNF and Synaptic Plasticity, Cognitive Function, and Dysfunction. In Neurotrophic Factors; Lewin, G.R., Carter, B.D., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 223–250. [Google Scholar]
- Blecharz-Klin, K.; Wawer, A.; Jawna-Zboinska, K.; Pyrzanowska, J.; Piechal, A.; Mirowska-Guzel, D.; Widy-Tyszkiewicz, E. Early paracetamol exposure decreases brain-derived neurotrophic factor (BDNF) in striatum and affects social behaviour and exploration in rats. Pharmacol. Biochem. Behav. 2018, 168, 25–32. [Google Scholar] [CrossRef]
- Klein, R.M.; Rigobello, C.; Vidigal, C.B.; Moura, K.F.; Barbosa, D.S.; Gerardin, D.C.C.; Ceravolo, G.S.; Moreira, E.G. Gestational exposure to paracetamol in rats induces neurofunctional alterations in the progeny. Neurotoxicol. Teratol. 2019, 77, 106838. [Google Scholar] [CrossRef] [PubMed]
- Viberg, H.; Eriksson, P.; Gordh, T.; Fredriksson, A. Paracetamol (acetaminophen) administration during neonatal brain development affects cognitive function and alters its analgesic and anxiolytic response in adult male mice. Toxicol. Sci. 2014, 138, 139–147. [Google Scholar] [CrossRef]
- FDA. Guidance for Industry: Estimating the Maximum Safe Starting Dose in Initial Clinical Trials for Therapeutics in Adult Healthy Volunteers: U.S. Food and Drug Administration. 2005. Available online: https://www.fda.gov/ucm/groups/fdagov-public/@fdagov-drugs-gen/documents/document/ucm078932.pdf (accessed on 15 December 2022).
- FDA. FDA Drug Safety Communication: Prescription Acetaminophen Products to Be Limited to 325 mg Per Dosage Unit; Boxed Warning Will Highlight Potential for Severe Liver Failure: U.S. Food and Drug Administration; 2018 [updated 02/07/2018]. Available online: https://www.fda.gov/drugs/drug-safety-and-availability/fda-drug-safety-communication-prescription-acetaminophen-products-be-limited-325-mg-dosage-unit (accessed on 15 December 2022).
- Morley, K.C.; Gallate, J.E.; Hunt, G.E.; Mallet, P.E.; McGregor, I.S. Increased anxiety and impaired memory in rats 3 months after administration of 3,4-methylenedioxymethamphetamine (“ecstasy”). Eur. J. Pharmacol. 2001, 433, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.G.M. Spatial localization does not require the presence of local cues. Learn. Motiv. 1981, 12, 239–260. [Google Scholar] [CrossRef]
- Ghanem, C.I.; Pérez, M.J.; Manautou, J.E.; Mottino, A.D. Acetaminophen from liver to brain: New insights into drug pharmacological action and toxicity. Pharmacol. Res. 2016, 109, 119–131. [Google Scholar] [CrossRef]
- Manyike, P.T.; Kharasch, E.D.; Kalhorn, T.F.; Slattery, J.T. Contribution of CYP2E1 and CYP3A to acetaminophen reactive metabolite formation. Clin. Pharmacol. Ther. 2000, 67, 275–282. [Google Scholar] [CrossRef]
- Pitchaimani, V.; Arumugam, S.; Thandavarayan, R.A.; Thiyagarajan, M.K.; Aiyalu, R.; Sreedhar, R.; Nakamura, T.; Watanabe, K. Nootropic activity of acetaminophen against colchicine induced cognitive impairment in rats. J. Clin. Biochem. Nutr. 2012, 50, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.X.; Zhang, J.H.; Cao, J.B.; Wang, W.; Wang, D.X.; Zhang, X.Y.; Yu, J.; Zhang, Y.Y.; Zhang, Y.Z.; Mi, W.D. Acetaminophen attenuates lipopolysaccharide-induced cognitive impairment through antioxidant activity. J. Neuroinflamm. 2017, 14, 17. [Google Scholar] [CrossRef] [PubMed]
- Garrone, B.; Durando, L.; Prenderville, J.; Sokolowska, E.; Milanese, C.; Di Giorgio, F.P.; Callaghan, C.; Bianchi, M. Paracetamol (acetaminophen) rescues cognitive decline, neuroinflammation and cytoskeletal alterations in a model of post-operative cognitive decline (POCD) in middle-aged rats. Sci. Rep. 2021, 11, 10139. [Google Scholar] [CrossRef]
- Upadhya, S.C.; Tirumalai, P.S.; Boyd, M.R.; Mori, T.; Ravindranath, V. Cytochrome P4502E (CYP2E) in brain: Constitutive expression, induction by ethanol and localization by fluorescence in situ hybridization. Arch. Biochem. Biophys. 2000, 373, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Posadas, I.; Santos, P.; Cena, V. Acetaminophen induces human neuroblastoma cell death through NFKB activation. PLoS ONE 2012, 7, e50160. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M. The importance of free radicals and catalytic metal ions in human diseases. Mol. Asp. Med. 1985, 8, 89–193. [Google Scholar] [CrossRef]
- Ramachandran, A.; Jaeschke, H. Oxidative Stress and Acute Hepatic Injury. Curr. Opin. Toxicol. 2018, 7, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Iwata, E.; Asanuma, M.; Nishibayashi, S.; Kondo, Y.; Ogawa, N. Different effects of oxidative stress on activation of transcription factors in primary cultured rat neuronal and glial cells. Brain Res. Mol. Brain Res. 1997, 50, 213–220. [Google Scholar] [CrossRef]
- Light, K.E.; Ge, Y.; Belcher, S.M. Early postnatal ethanol exposure selectively decreases BDNF and truncated TrkB-T2 receptor mRNA expression in the rat cerebellum. Brain Res. Mol. Brain Res. 2001, 93, 46–55. [Google Scholar] [CrossRef]
- Lu, W.; Man, H.; Ju, W.; Trimble, W.S.; MacDonald, J.F.; Wang, Y.T. Activation of synaptic NMDA receptors induces membrane insertion of new AMPA receptors and LTP in cultured hippocampal neurons. Neuron 2001, 29, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Högestätt, E.D.; Jönsson, B.A.; Ermund, A.; Andersson, D.A.; Björk, H.; Alexander, J.P.; Cravatt, B.F.; Basbaum, A.I.; Zygmunt, P.M. Conversion of acetaminophen to the bioactive N-acylphenolamine AM404 via fatty acid amide hydrolase-dependent arachidonic acid conjugation in the nervous system. J. Biol. Chem. 2005, 280, 31405–31412. [Google Scholar] [CrossRef]
- Muramatsu, S.; Shiraishi, S.; Miyano, K.; Sudo, Y.; Toda, A.; Mogi, M.; Hara, M.; Yokoyama, A.; Kawasaki, Y.; Taniguchi, M.; et al. Metabolism of AM404 From Acetaminophen at Human Therapeutic Dosages in the Rat Brain. Anesth. Pain Med. 2016, 6, e32873. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, D.C.; Pittman, B.; Perry, E.; Simen, A. Preliminary evidence of cannabinoid effects on brain-derived neurotrophic factor (BDNF) levels in humans. Psychopharmacology 2009, 202, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Bamji, S.X.; Rico, B.; Kimes, N.; Reichardt, L.F. BDNF mobilizes synaptic vesicles and enhances synapse formation by disrupting cadherin-beta-catenin interactions. J. Cell Biol. 2006, 174, 289–299. [Google Scholar] [CrossRef]
- Bramham, C.R.; Messaoudi, E. BDNF function in adult synaptic plasticity: The synaptic consolidation hypothesis. Prog. Neurobiol. 2005, 76, 99–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liu, I.Y.; Bi, X.; Thompson, R.F.; Doctrow, S.R.; Malfroy, B.; Baudry, M. Reversal of age-related learning deficits and brain oxidative stress in mice with superoxide dismutase/catalase mimetics. Proc. Natl. Acad. Sci. USA 2003, 100, 8526–8531. [Google Scholar] [CrossRef] [PubMed]
- Luikart, B.W.; Nef, S.; Virmani, T.; Lush, M.E.; Liu, Y.; Kavalali, E.T.; Parada, L.F. TrkB has a cell-autonomous role in the establishment of hippocampal Schaffer collateral synapses. J. Neurosci. 2005, 25, 3774–3786. [Google Scholar] [CrossRef] [PubMed]
- Martinez, A.; Alcantara, S.; Borrell, V.; Del Rio, J.A.; Blasi, J.; Otal, R.; Campos, N.; Boronat, A.; Barbacid, M.; Silos-Santiago, I.; et al. TrkB and TrkC signaling are required for maturation and synaptogenesis of hippocampal connections. J. Neurosci. 1998, 18, 7336–7350. [Google Scholar] [CrossRef] [PubMed]
- Otal, R.; Martinez, A.; Soriano, E. Lack of TrkB and TrkC signaling alters the synaptogenesis and maturation of mossy fiber terminals in the hippocampus. Cell Tissue Res. 2005, 319, 349–358. [Google Scholar] [CrossRef]
- Hou, Y.; Zhou, L.; Yang, Q.D.; Du, X.P.; Li, M.; Yuan, M.; Zhou, Z.W. Changes in hippocampal synapses and learning-memory abilities in a streptozotocin-treated rat model and intervention by using fasudil hydrochloride. Neuroscience 2012, 200, 120–129. [Google Scholar] [CrossRef]
- Xiao, Y.; Fu, H.; Han, X.; Hu, X.; Gu, H.; Chen, Y.; Wei, Q.; Hu, Q. Role of Synaptic Structural Plasticity in Impairments of Spatial Learning and Memory Induced by Developmental Lead Exposure in Wistar Rats. PLoS ONE 2014, 9, e115556. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lalert, L.; Tantarungsee, N.; Chotipinit, T.; Ji-au, W.; Srikiatkhachorn, A.; Maneesri-le Grand, S. Long-Term Paracetamol Treatment Impairs Cognitive Function and Brain-Derived Neurotrophic Factor in Adult Rat Brain. Sci. Pharm. 2023, 91, 11. https://doi.org/10.3390/scipharm91010011
Lalert L, Tantarungsee N, Chotipinit T, Ji-au W, Srikiatkhachorn A, Maneesri-le Grand S. Long-Term Paracetamol Treatment Impairs Cognitive Function and Brain-Derived Neurotrophic Factor in Adult Rat Brain. Scientia Pharmaceutica. 2023; 91(1):11. https://doi.org/10.3390/scipharm91010011
Chicago/Turabian StyleLalert, Laddawan, Nutnicha Tantarungsee, Tipthanan Chotipinit, Wilawan Ji-au, Anan Srikiatkhachorn, and Supang Maneesri-le Grand. 2023. "Long-Term Paracetamol Treatment Impairs Cognitive Function and Brain-Derived Neurotrophic Factor in Adult Rat Brain" Scientia Pharmaceutica 91, no. 1: 11. https://doi.org/10.3390/scipharm91010011