
Assessing Anti-Adipogenic Effects of Mango Leaf Tea and Mangiferin within Cultured Adipocytes
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Mangiferin Quantification
2.3. Cell Culture and Adipocyte Differentiation
2.4. MTT Assay
2.5. Measurement of Cellular Triglyceride Content
2.6. Oil-Red O Staining
2.7. qRT-PCR
2.8. Measurement of Secretory Adiponectin Levels
2.9. Cellular Glucose Uptake
2.10. Statistical Analysis
3. Results
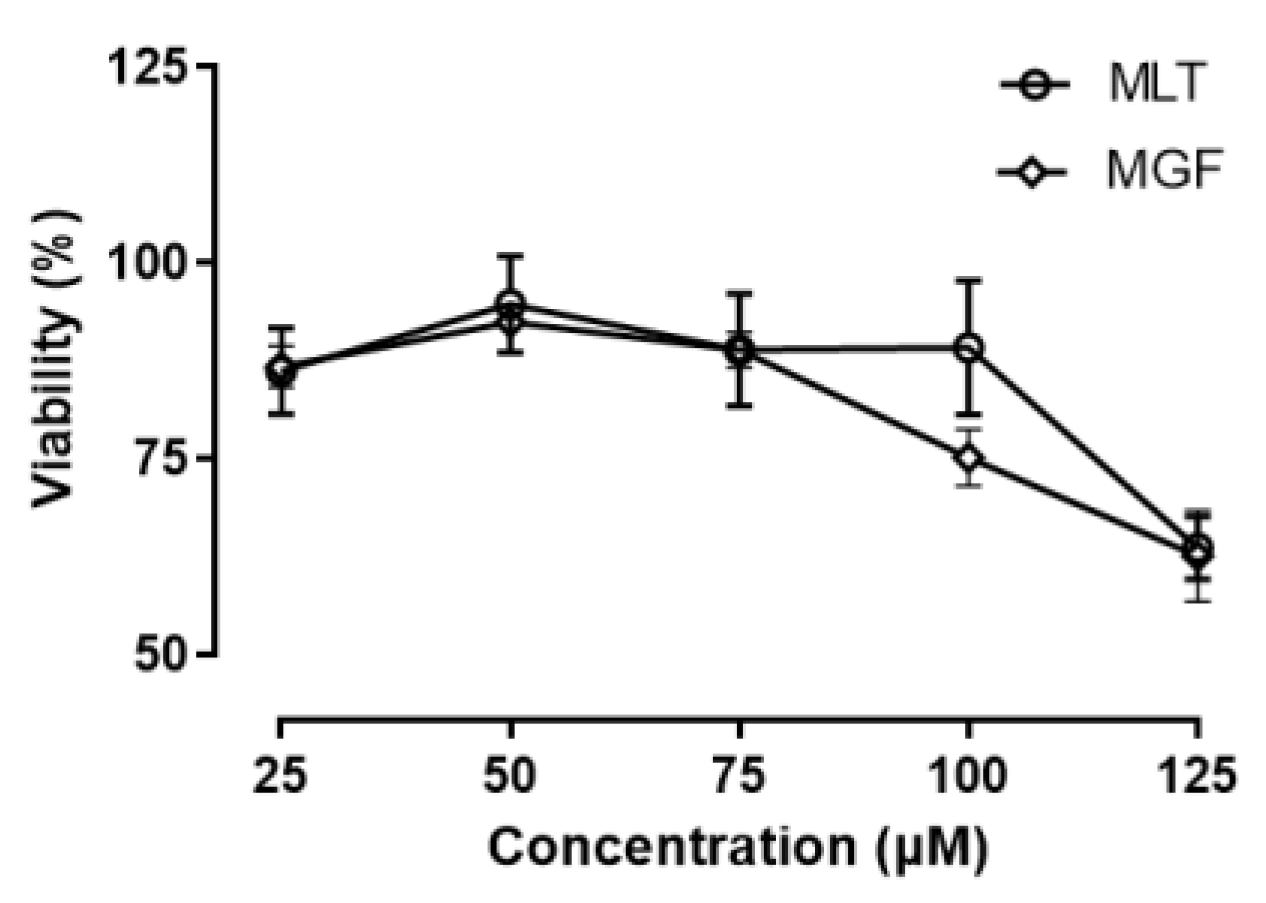
3.1. Toxicity Analysis of 3T3-L1 Cells Exposed to MGF and MLT
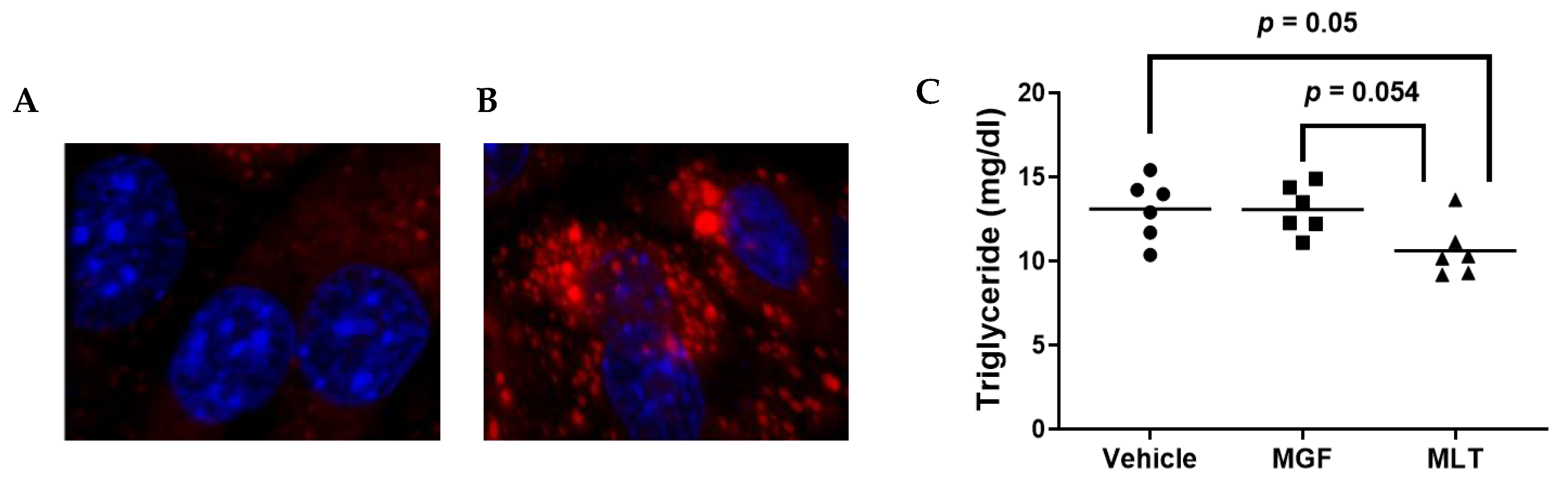
3.2. MLT-Mediated Suppression of Intracellular Triglycerides within Cultured Adipocytes
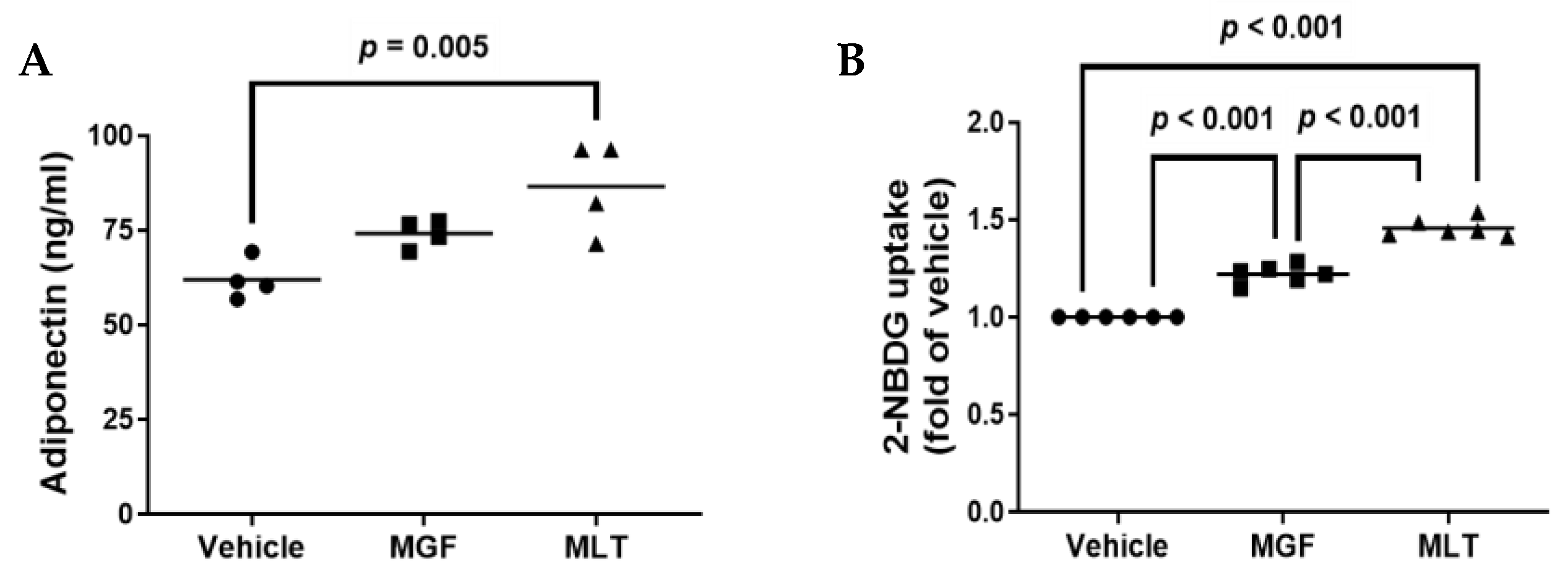
3.3. MLT Treatment Increases Secretory Adiponectin and Improves Glucose Uptake in Cultured Adipocytes
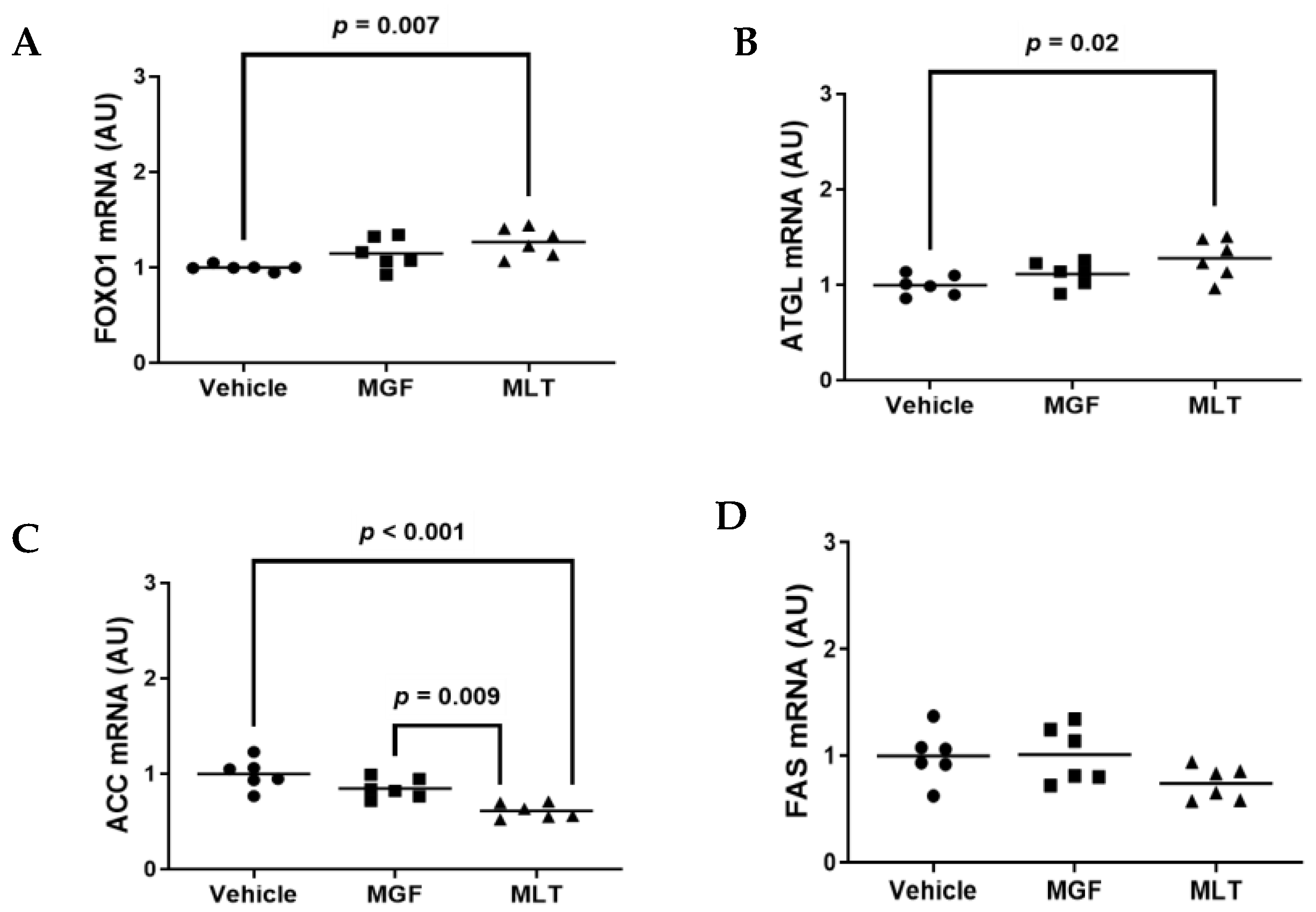
3.4. Exposing Cultured Adipocytes to MLT Alters Metabolic Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garg, S.K.; Maurer, H.; Reed, K.; Selagamsetty, R. Diabetes and cancer: Two diseases with obesity as a common risk factor. Diabetes Obes. Metab. 2014, 16, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Ko, H.; Shin, H.; Kim, H.; Hong, Y.; Lee, N.; Kim, S. Fucoxanthin exerts differing effects on 3T3-L1 cells according to the differentiation stage and inhibits glucose uptake in mature adipocytes. Biochem. Biophys. Res. Commun. 2011, 409, 769–774. Available online: https://www.sciencedirect.com/science/article/pii/S0006291X11008539 (accessed on 13 April 2023). [CrossRef] [PubMed]
- Day, C.; Bailey, C.J. Obesity in the pathogenesis of type 2 diabetes. Br. J. Diabetes Vasc. Dis. 2011, 11, 55–61. [Google Scholar] [CrossRef]
- Bhupathiraju, S.N.; Hu, F.B. Epidemiology of obesity and diabetes and their cardiovascular complications. Circ. Res. 2016, 118, 1723–1735. [Google Scholar] [CrossRef]
- Park, E.; Kim, J.; Yeo, S.; Kim, G.; Ko, E.; Lee, S.W.; Li, W.Y.; Choi, C.W.; Jeong, S. Antiadipogenic effects of loganic acid in 3T3-L1 preadipocytes and ovariectomized mice. Molecules 2018, 23, 1663. [Google Scholar] [CrossRef]
- van der Zwaal, E.M.; Janhunen, S.K.; Luijendijk, M.C.; Baclesanu, R.; Vanderschuren, L.J.; Adan, R.A.; La Fleur, S.E. Olanzapine and sibutramine have opposing effects on the motivation for palatable food. Behav. Pharmacol. 2012, 23, 198–204. [Google Scholar] [CrossRef]
- Buyukhatipoglu, H. A possibly overlooked side effect of orlistat: Gastroesophageal reflux disease. J. Natl. Med. Assoc. 2008, 100, 1207. [Google Scholar] [CrossRef]
- Yoo, S.; Seo, C.; Shin, H.; Jeong, S. Traditional herbal formula oyaksungi-san inhibits adipogenesis in 3T3-L1 adipocytes. Evid.-Based Complement. Altern. Med. 2015, 2015, 949461. [Google Scholar] [CrossRef]
- Stamatikos, A.D.; Davis, J.E.; Shay, N.F.; Ajuwon, K.M.; Deyhim, F.; Banz, W.J. Consuming Diet Supplemented with Either Red Wheat Bran or Soy Extract Changes Glucose and Insulin Levels in Female Obese Zucker Rats. Int. J. Vitam. Nutr. Res. 2019, 90, 23–32. [Google Scholar] [CrossRef]
- Choi, I.; Park, Y.; Choi, H.; Lee, E.H. Anti-adipogenic activity of rutin in 3T3-L1 cells and mice fed with high-fat diet. Biofactors 2006, 26, 273–281. [Google Scholar] [CrossRef]
- Cho, E.; Rahman, A.; Kim, S.; Baek, Y.; Hwang, H.; Oh, J.; Hwang, H.; Lee, S.; Yun, J. Chitosan oligosaccharides inhibit adipogenesis in 3T3-L1 adipocytes. J. Microbiol. Biotechnol. 2008, 18, 80–87. [Google Scholar] [PubMed]
- Tang, Q.; Otto, T.C.; Lane, M.D. Mitotic clonal expansion: A synchronous process required for adipogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 44–49. [Google Scholar] [CrossRef]
- Kim, S.; Kong, C. Anti-adipogenic effect of dioxinodehydroeckol via AMPK activation in 3T3-L1 adipocytes. Chem. Biol. Interact. 2010, 186, 24–29. [Google Scholar] [CrossRef]
- Wang, N.; Wang, D.; Luo, G.; Zhou, J.; Tan, Z.; Du, Y.; Xie, H.; Liu, L.; Yang, X.; Hao, L. Lutein attenuates excessive lipid accumulation in differentiated 3T3-L1 cells and abdominal adipose tissue of rats by the SIRT1-mediated pathway. Int. J. Biochem. Cell Biol. 2021, 133, 105932. Available online: https://www.sciencedirect.com/science/article/pii/S1357272521000169 (accessed on 13 April 2023). [CrossRef] [PubMed]
- Zarasvand, S.A.; Mullins, A.P.; Arjmandi, B.; Haley-Zitlin, V. Antidiabetic properties of mango in animal models and humans: A systematic review. Nutr. Res. 2023, 111, 73–89. [Google Scholar] [CrossRef]
- Severi, J.A.; Lima, Z.P.; Kushima, H.; Monteiro Souza Brito, A.R.; Campaner dos Santos, L.; Vilegas, W.; Hiruma-Lima, C.A. Polyphenols with antiulcerogenic action from aqueous decoction of mango leaves (Mangifera indica L.). Molecules 2009, 14, 1098–1110. [Google Scholar] [CrossRef]
- Ramirez, J.E.; Zambrano, R.; Sepúlveda, B.; Simirgiotis, M.J. Antioxidant properties and hyphenated HPLC-PDA-MS profiling of Chilean Pica mango fruits (Mangifera indica L. Cv. piqueño). Molecules 2013, 19, 438–458. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Sellés, A.J.; Delgado-Hernández, R.; Garrido-Garrido, G.; García-Rivera, D.; Guevara-García, M.; Pardo-Andreu, G.L. The paradox of natural products as pharmaceuticals: Experimental evidences of a mango stem bark extract. Pharmacol. Res. 2007, 55, 351–358. [Google Scholar] [CrossRef]
- Rodeiro, I.; Hernandez, S.; Morffi, J.; Herrera, J.A.; Gómez-Lechón, M.J.; Delgado, R.; Espinosa-Aguirre, J.J. Evaluation of genotoxicity and DNA protective effects of mangiferin, a glucosylxanthone isolated from Mangifera indica L. stem bark extract. Food Chem. Toxicol. 2012, 50, 3360–3366. [Google Scholar] [CrossRef]
- Choi, E.H.; Yang, H.P.; Chun, H.S. Chitooligosaccharide ameliorates diet-induced obesity in mice and affects adipose gene expression involved in adipogenesis and inflammation. Nutr. Res. 2012, 32, 218–228. [Google Scholar] [CrossRef]
- García-Rivera, D.; Delgado, R.; Bougarne, N.; Haegeman, G.; Berghe, W.V. Gallic acid indanone and mangiferin xanthone are strong determinants of immunosuppressive anti-tumour effects of Mangifera indica L. bark in MDA-MB231 breast cancer cells. Cancer Lett. 2011, 305, 21–31. [Google Scholar] [CrossRef]
- Alasvand, S.; Crane, J.; Haley-Zitlin, V. Chemical Characteristics of Mango Leaf Tea Prepared from Different Varieties of Mangifera indica L. Leaves. Curr. Dev. Nutr. 2022, 6, 263. [Google Scholar] [CrossRef]
- Alasvand Zarasvand, S. Chemical and Sensory Analyses of Teas from Mangifera indica L. Leaves and Evaluation of Their Anti-Adipogenic Effect in 3T3-L1 Adipocytes; Publication No. 3213; TigerPrints Publishing, Clemson University: Clemson, SC, USA, 2022. [Google Scholar]
- Zebisch, K.; Voigt, V.; Wabitsch, M.; Brandsch, M. Protocol for effective differentiation of 3T3-L1 cells to adipocytes. Anal. Biochem. 2012, 425, 88–90. [Google Scholar] [CrossRef]
- Kumar, B.D.; Krishnakumar, K.; Jaganathan, S.K.; Mandal, M. Effect of Mangiferin and Mahanimbine on Glucose Utilization in 3T3-L1 cells. Pharmacogn. Mag. 2013, 9, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Jo, H.; Echesabal-Chen, J.; Stamatikos, A. Combined LXR and RXR Agonist Therapy Increases ABCA1 Protein Expression and Enhances ApoAI-Mediated Cholesterol Efflux in Cultured Endothelial Cells. Metabolites 2021, 11, 640. [Google Scholar] [CrossRef]
- Esobi, I.C.; Barksdale, C.; Heard-Tate, C.; Reigers Powell, R.; Bruce, T.F.; Stamatikos, A. MOVAS Cells: A Versatile Cell Line for Studying Vascular Smooth Muscle Cell Cholesterol Metabolism. Lipids 2021, 56, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Haslam, D.W.; James, W.P. Obesity. Lancet 2005, 366, 1197–1209. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Hirose, H.; Saito, I.; Tomita, M.; Taniyama, M.; Matsubara, K.; Okazaki, Y.; Ishii, T.; Nishikai, K.; Saruta, T. Correlation of the adipocyte-derived protein adiponectin with insulin resistance index and serum high-density lipoprotein-cholesterol, independent of body mass index, in the Japanese population. Clin. Sci. 2002, 103, 137–142. [Google Scholar] [CrossRef]
- Ronti, T.; Lupattelli, G.; Mannarino, E. The endocrine function of adipose tissue: An update. Clin. Endocrinol. 2006, 64, 355–365. [Google Scholar] [CrossRef]
- Sferrazzo, G.; Palmeri, R.; Vanella, L.; Parafati, L.; Ronsisvalle, S.; Biondi, A.; Basile, F.; Li Volti, G.; Barbagallo, I. Mangifera indica L. leaf extract induces adiponectin and regulates adipogenesis. Int. J. Mol. Sci. 2019, 20, 3211. [Google Scholar] [CrossRef] [PubMed]
- Cowherd, R.M.; Lyle, R.E.; McGehee, R.E., Jr. Molecular Regulation of Adipocyte Differentiation, Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 1999; pp. 3–10. [Google Scholar]
- Kozak, L.P.; Jensen, J.T. Genetic and developmental control of multiple forms of L-glycerol 3-phosphate dehydrogenase. J. Biol. Chem. 1974, 249, 7775–7781. [Google Scholar] [CrossRef] [PubMed]
- Siersbæk, R.; Nielsen, R.; Mandrup, S. PPARγ in adipocyte differentiation and metabolism–novel insights from genome-wide studies. FEBS Lett. 2010, 584, 3242–3249. [Google Scholar] [CrossRef] [PubMed]
- Hafidi, M.E.; Buelna-Chontal, M.; Sánchez-Muñoz, F.; Carbó, R. Adipogenesis: A necessary but harmful strategy. Int. J. Mol. Sci. 2019, 20, 3657. [Google Scholar] [CrossRef]
- Song, Z.; Xiaoli, A.M.; Yang, F. Regulation and metabolic significance of de novo lipogenesis in adipose tissues. Nutrients 2018, 10, 1383. [Google Scholar] [CrossRef]
- Rosen, E.D.; Spiegelman, B.M. Adipocytes as regulators of energy balance and glucose homeostasis. Nature 2006, 444, 847–853. [Google Scholar] [CrossRef]
- Nakae, J.; Kitamura, T.; Kitamura, Y.; Biggs, W.H., III; Arden, K.C.; Accili, D. The forkhead transcription factor Foxo1 regulates adipocyte differentiation. Dev. Cell 2003, 4, 119–129. [Google Scholar] [CrossRef]
- Schweiger, M.; Schreiber, R.; Haemmerle, G.; Lass, A.; Fledelius, C.; Jacobsen, P.; Tornqvist, H.; Zechner, R.; Zimmermann, R. Adipose triglyceride lipase and hormone-sensitive lipase are the major enzymes in adipose tissue triacylglycerol catabolism. J. Biol. Chem. 2006, 281, 40236–40241. [Google Scholar] [CrossRef]
- Denechaud, P.; Bossard, P.; Lobaccaro, J.A.; Millatt, L.; Staels, B.; Girard, J.; Postic, C. ChREBP, but not LXRs, is required for the induction of glucose-regulated genes in mouse liver. J. Clin. Investig. 2008, 118, 956–964. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Han, L.; Gao, X.; Liu, E.; Wang, T. Regulation of lipid and glucose homeostasis by mango tree leaf extract is mediated by AMPK and PI3K/AKT signaling pathways. Food Chem. 2013, 141, 2896–2905. [Google Scholar] [CrossRef]
- Lafontan, M.; Langin, D. Lipolysis and lipid mobilization in human adipose tissue. Prog. Lipid Res. 2009, 48, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Tang, T.; Abbott, M.; Viscarra, J.A.; Wang, Y.; Sul, H.S. AMPK phosphorylates desnutrin/ATGL and hormone-sensitive lipase to regulate lipolysis and fatty acid oxidation within adipose tissue. Mol. Cell. Biol. 2016, 36, 1961–1976. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, E.; Goldberg, E.B.; Makarova, K.S.; Lin, L.; Brown, W.J.; Jackson, C.L. ATGL has a key role in lipid droplet/adiposome degradation in mammalian cells. EMBO Rep. 2006, 7, 106–113. [Google Scholar] [CrossRef]
- Kershaw, E.E.; Hamm, J.K.; Verhagen, L.A.; Peroni, O.; Katic, M.; Flier, J.S. Adipose triglyceride lipase: Function, regulation by insulin, and comparison with adiponutrin. Diabetes 2006, 55, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, P.; English, T.; Karki, S.; Qiang, L.; Tao, R.; Kim, J.; Luo, Z.; Farmer, S.R.; Kandror, K.V. SIRT1 controls lipolysis in adipocytes via FOXO1-mediated expression of ATGL. J. Lipid Res. 2011, 52, 1693–1701. [Google Scholar] [CrossRef]
- Zhang, Y.; Qian, Q.; Ge, D.; Li, Y.; Wang, X.; Chen, Q.; Gao, X.; Wang, T. Identification of benzophenone C-glucosides from mango tree leaves and their inhibitory effect on triglyceride accumulation in 3T3-L1 adipocytes. J. Agric. Food Chem. 2011, 59, 11526–11533. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Sequence (5′-3′) | Amplicon (bp) | Accession Number |
|---|---|---|---|
| GAPDH | Forward: AACTTTGGCATTGTGGAAGG | 132 | NM_001289726 |
| Reverse: GGATGCAGGGATGATGTTCT | |||
| FOXO1 | Forward: GATCTACGAGTGGATGGT | 523 | NM_019739 |
| Reverse: CAGCGTAGACGCCATCTT | |||
| ATGL | Forward: CACTTTAGCTCCAAGGATGA | 380 | NM_001163689 |
| Reverse: TGGTTCAGTAGGCCATTCCT | |||
| ACC | Forward: GGGCTACCTCTAATGGTCTT | 439 | NM_133360 |
| Reverse: CTACCTGATGGTAAATGGGA | |||
| FAS | Forward: TTGCTGGCACTACAGAATGC | 192 | NM_007988 |
| Reverse: AACAGCCTCAGAGCGACAAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zarasvand, S.A.; Haley-Zitlin, V.; Oladosu, O.; Esobi, I.; Powell, R.R.; Bruce, T.; Stamatikos, A. Assessing Anti-Adipogenic Effects of Mango Leaf Tea and Mangiferin within Cultured Adipocytes. Diseases 2023, 11, 70. https://doi.org/10.3390/diseases11020070
Zarasvand SA, Haley-Zitlin V, Oladosu O, Esobi I, Powell RR, Bruce T, Stamatikos A. Assessing Anti-Adipogenic Effects of Mango Leaf Tea and Mangiferin within Cultured Adipocytes. Diseases. 2023; 11(2):70. https://doi.org/10.3390/diseases11020070
Chicago/Turabian StyleZarasvand, Sepideh Alasvand, Vivian Haley-Zitlin, Olanrewaju Oladosu, Ikechukwu Esobi, Rhonda Reigers Powell, Terri Bruce, and Alexis Stamatikos. 2023. "Assessing Anti-Adipogenic Effects of Mango Leaf Tea and Mangiferin within Cultured Adipocytes" Diseases 11, no. 2: 70. https://doi.org/10.3390/diseases11020070