Chronotype and Social Jetlag: A (Self-) Critical Review


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
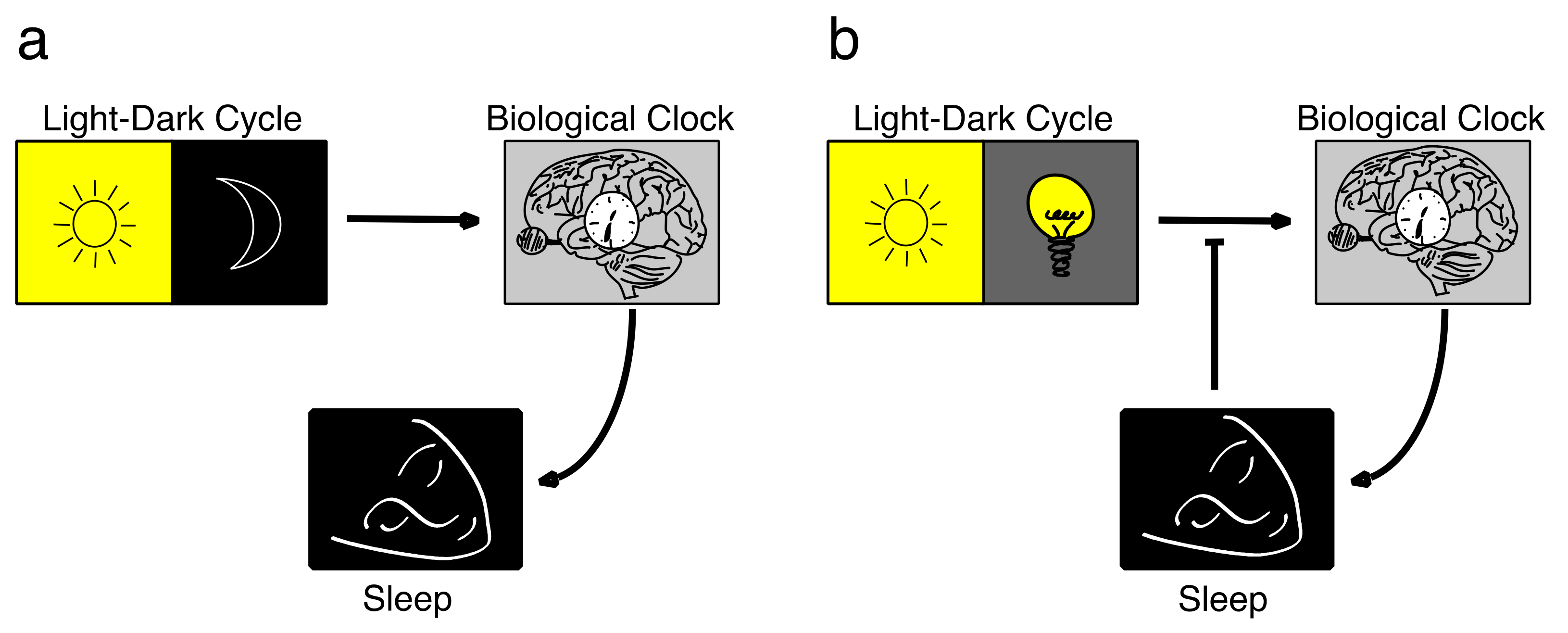
Principles of Entrainment
2. Chronotype
2.1. Concept
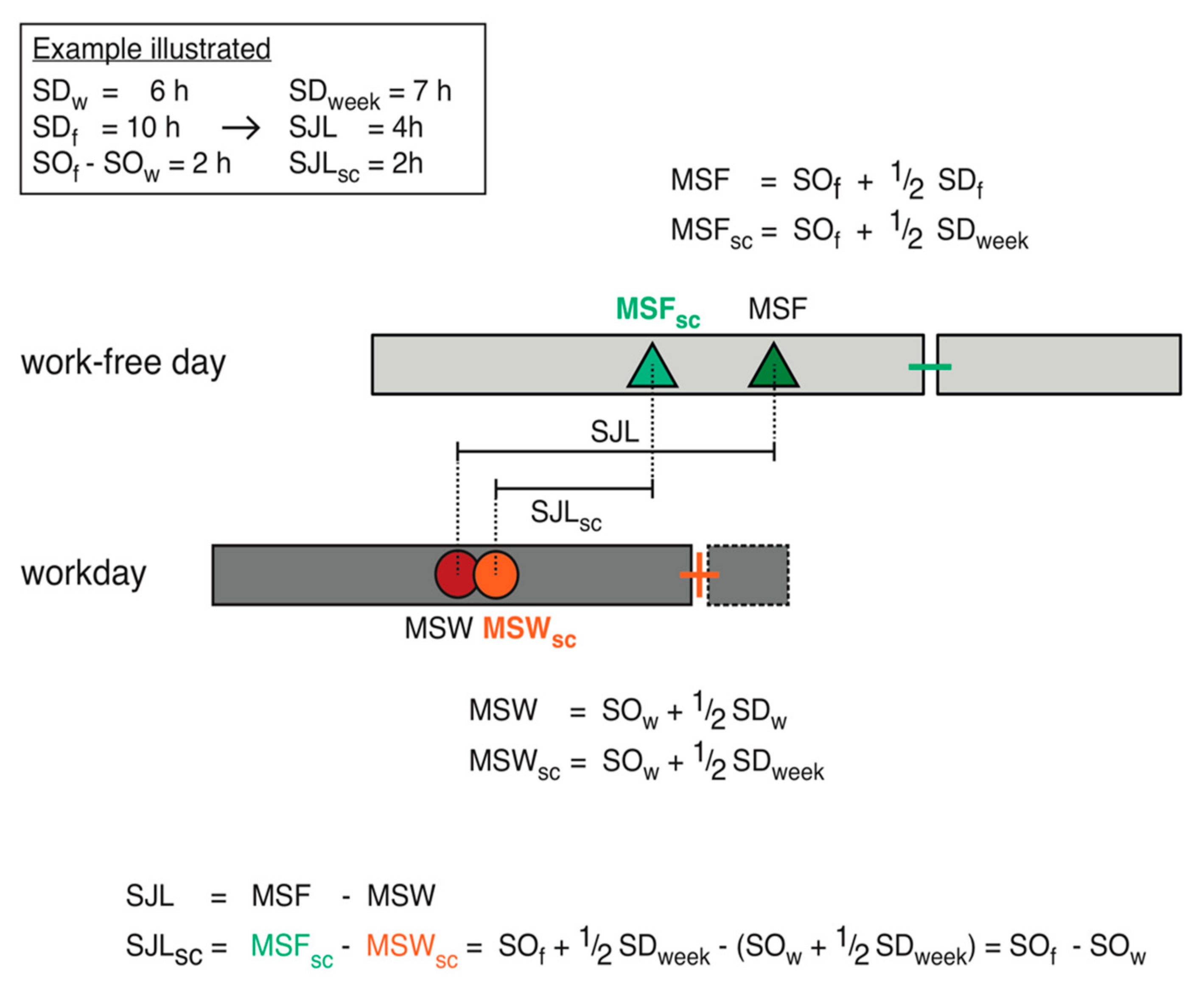
2.2. MCTQ-Estimation of Chronotype
- MSF = midsleep on free days
- MSFsc = midsleep on free days sleep corrected
- SDw = sleep duration on workdays
- SDf = sleep duration on work-free days
- SDweek = weekly average sleep duration
- SOf = sleep onset on work-free days
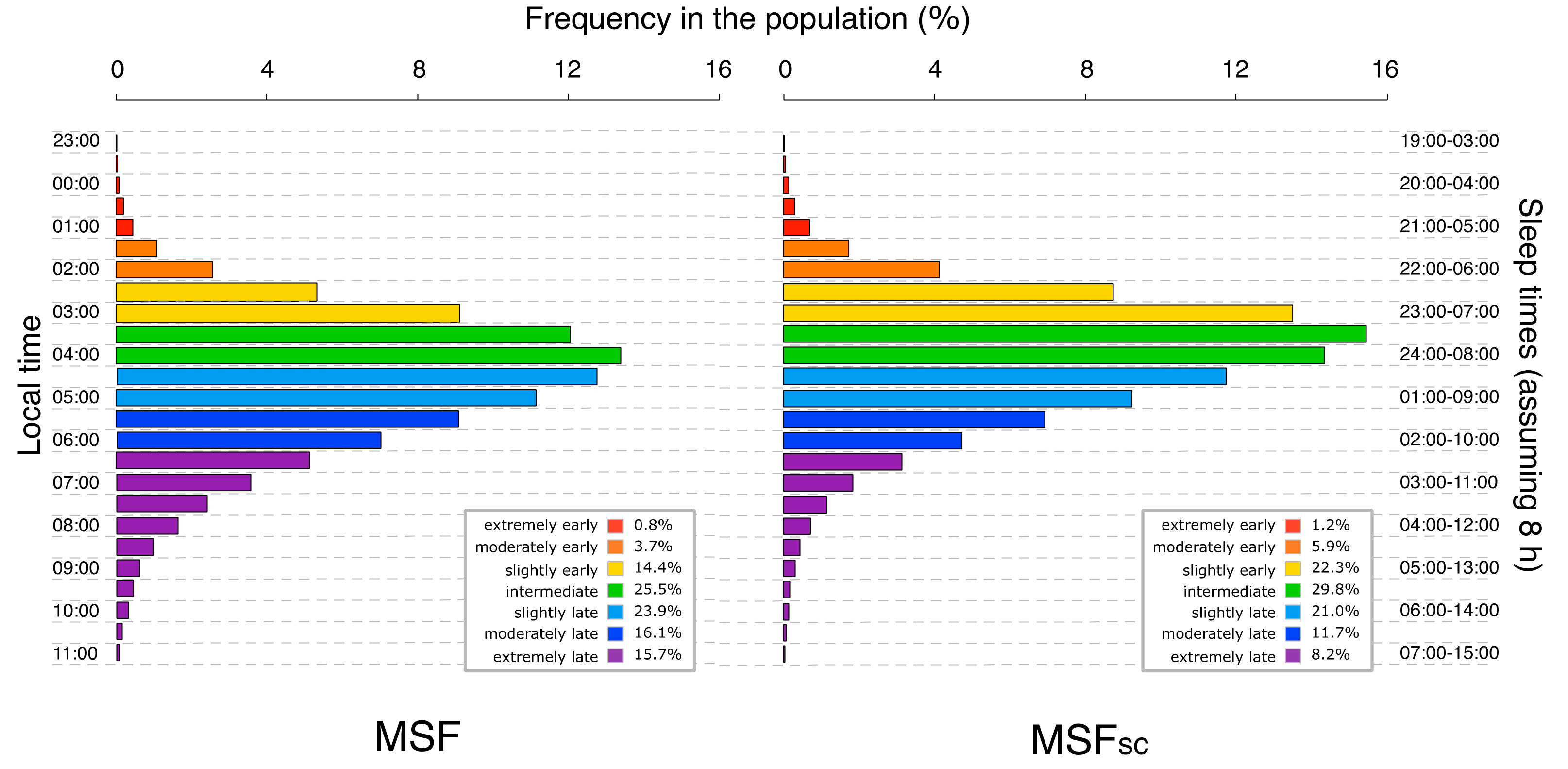
2.3. Characteristics of MCTQ-Chronotype
2.4. Discussion
2.4.1. MCTQ-Chronotyping: Pros and Cons
2.4.2. The Stability of a Chronotype: Trait or State?
2.4.3. How is Circadian State Related to Health and Disease?
3. Social Jetlag
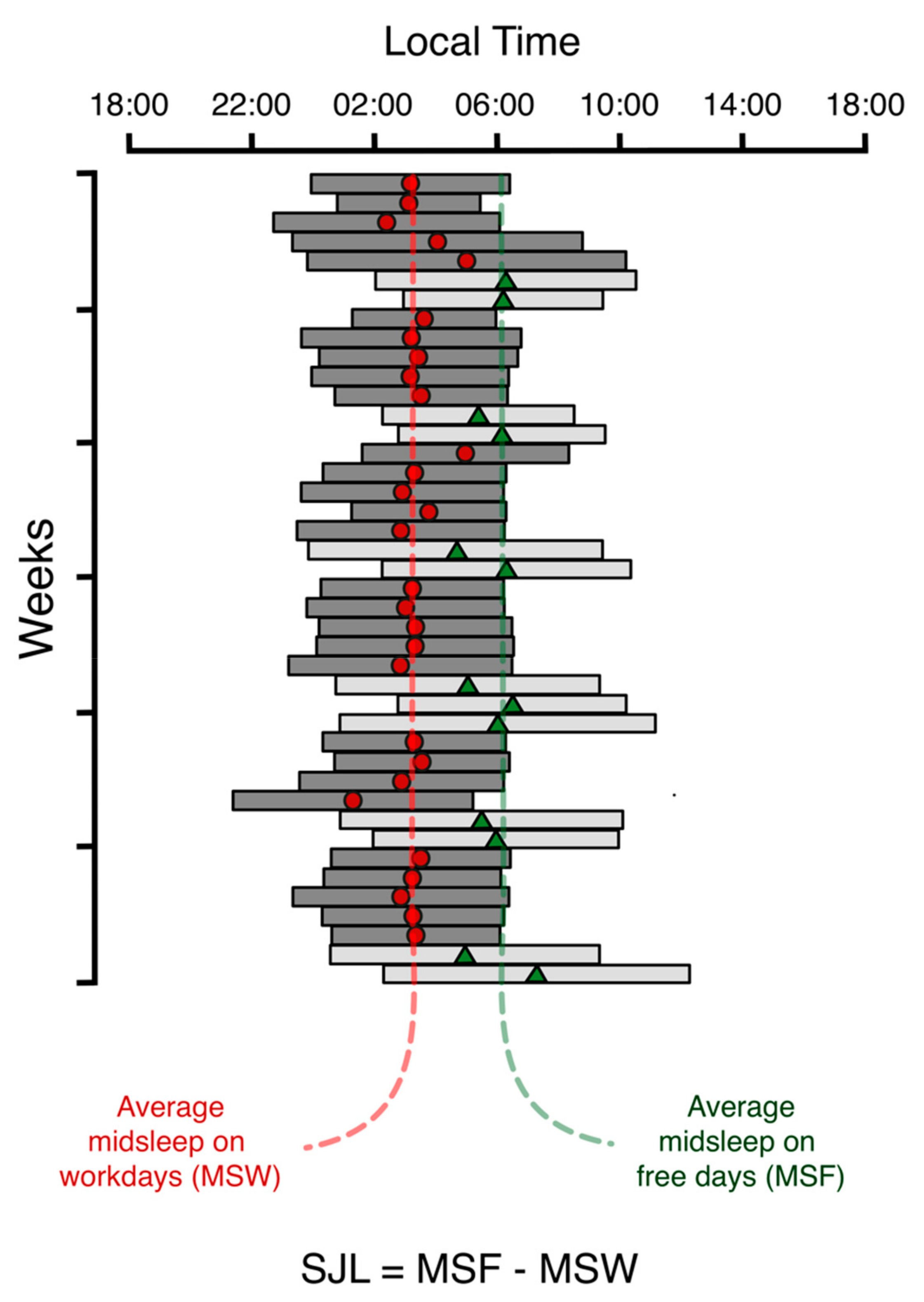
3.1. Concept
3.2. Social Jetlag Computation
- SJL: social jetlag
- MSF: midsleep on free days
- MSW: midsleep on workdays
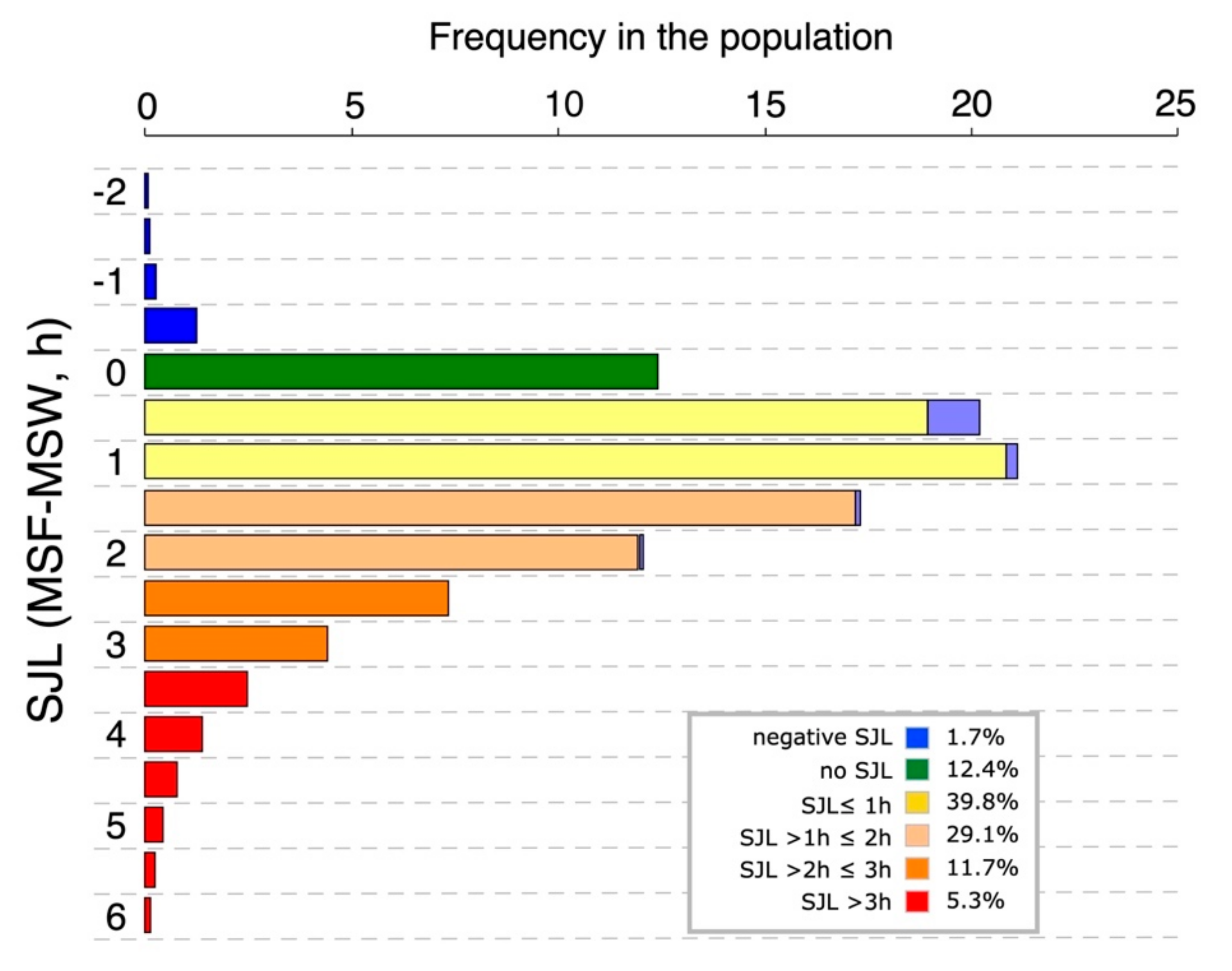
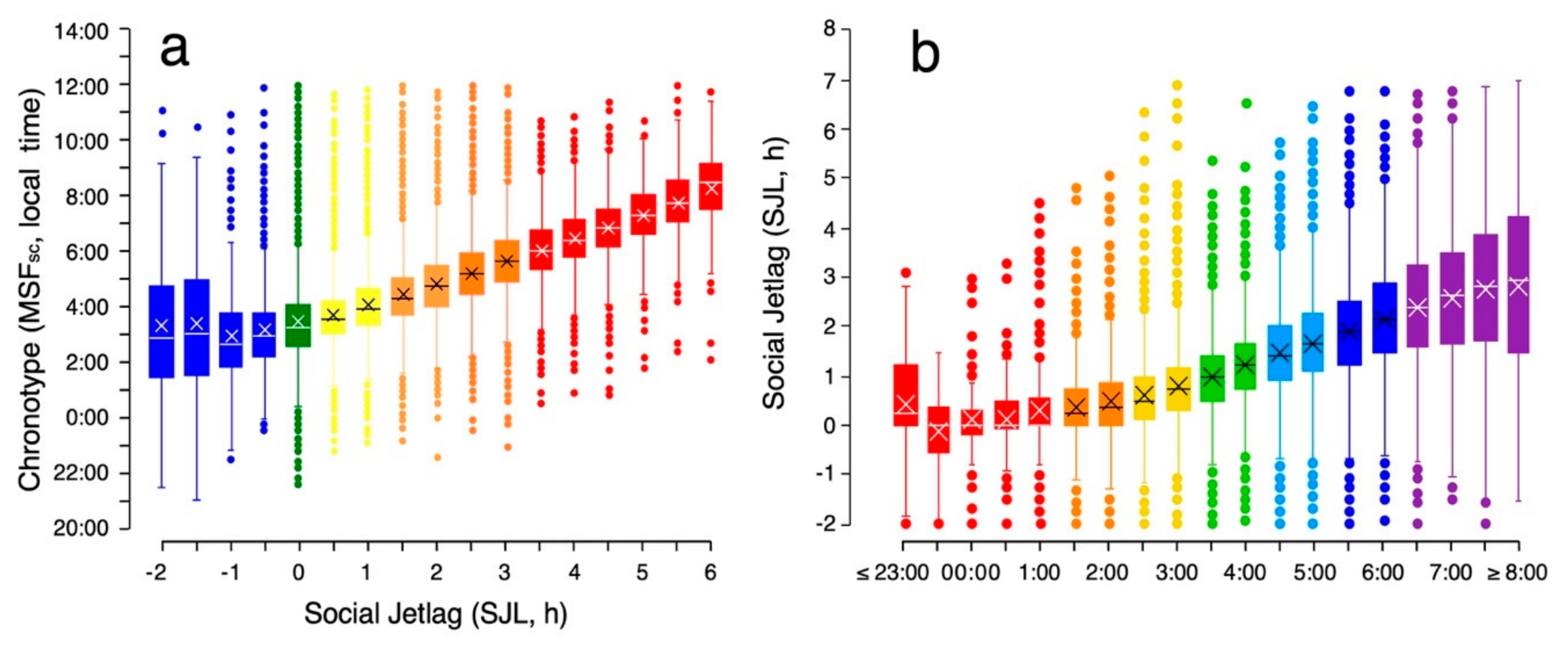
3.3. Characteristics of Social Jetlag
3.4. Discussion
3.4.1. What Does Social Jetlag Quantify?
3.4.2. Social Jetlag and Sleep Debt
3.4.3. Misunderstandings About Social Jetlag and Conundrums to be Solved
4. Outlook
Author Contributions
Funding
Conflicts of Interest
References
- Williams, G.E. Geological constraints on the Precambrian history of Earth’s rotation and the Moon’s orbit. Rev. Geophys. 2000, 38, 37–59. [Google Scholar] [CrossRef]
- Roenneberg, T.; Daan, S.; Merrow, M. The art of entrainment. J. Biol. Rhythms 2003, 18, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Wirz-Justice, A.; Merrow, M. Life between clocks: Daily temporal patterns of human chronotypes. J. Biol. Rhythms 2003, 18, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.P.; McHill, A.W.; Birks, B.R.; Griffin, B.R.; Rusterholz, T.; Chinoy, E.D. Entrainment of the human circadian clock to the natural light-dark cycle. Curr. Biol. 2013, 23, 1554–1558. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, M.; Dinich, J.; Merrow, M.; Roenneberg, T. Social jetlag: Misalignment of biological and social time. Chronobiol. Int. 2006, 23, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Horne, J.A.; Ostberg, O. Individual differences in human circadian rhythms. Biol. Psychol. 1977, 5, 179–190. [Google Scholar] [CrossRef]
- Kandeger, A.; Selvi, Y.; Tanyer, D.K. The effects of individual circadian rhythm differences on insomnia, impulsivity, and food addiction. Eat. Weight Disord. EWD 2019, 24, 47–55. [Google Scholar] [CrossRef]
- Randler, C. Morningness–eveningness and satisfaction with life. Soc. Indic. Res. 2008, 86, 297–302. [Google Scholar] [CrossRef]
- Horne, J.A.; Ostberg, O. A self-assessment questionnaire to determine morningness-eveningness in human circadian rhythms. Int. J. Chronobiol. 1976, 4, 97–110. [Google Scholar]
- Ehret, C.F. The sense of time: Evidence for its molecular basis in the eukaryotic gene-action system. Adv. Biol. Med. Phys. 1974, 15, 47–77. [Google Scholar]
- Samis, H.V. (Ed.) Aging and Biological Rhythms (Advances in Experimental Medicine and Biology), 15th ed.; Springer: New York, NY, USA, 1978; ISBN 9781475744620. [Google Scholar]
- Pittendrigh, C.S. Temporal organization: Reflections of a Darwinian clock-watcher. Annu. Rev. Physiol. 1993, 55, 17–54. [Google Scholar] [CrossRef] [PubMed]
- Menaker, M. The search for principles of physiological organization in vertebrate circadian systems. In Vertebrate Circadian Systems; Proceedings in Life Sciences; Aschoff, J., Daan, S., Groos, G.A., Eds.; Springer: Berlin/Heidelberg, Germany, 1982; pp. 1–12. ISBN 9783642686511. [Google Scholar]
- Menaker, M.; Moreira, L.F.; Tosini, G. Evolution of circadian organization in vertebrates. Braz. J. Med. Biol. Res. 1997, 30, 305–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Merrow, M. The circadian clock and human health. Curr. Biol. CB 2016, 26, R432–R443. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.S.P.; Chang, A.-M.; Shulman, J.M.; Raj, T.; Chibnik, L.B.; Cain, S.W.; Rothamel, K.; Benoist, C.; Myers, A.J.; Czeisler, C.A.; et al. A common polymorphism near PER1 and the timing of human behavioral rhythms. Ann. Neurol. 2012, 72, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; Smits, M.; Spence, W.; Srinivasan, V.; Cardinali, D.P.; Lowe, A.D.; Kayumov, L. Dim light melatonin onset (DLMO): A tool for the analysis of circadian phase in human sleep and chronobiological disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2007, 31, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Kuehnle, T.; Juda, M.; Kantermann, T.; Allebrandt, K.; Gordijn, M.; Merrow, M. Epidemiology of the human circadian clock. Sleep Med. Rev. 2007, 11, 429–438. [Google Scholar] [CrossRef] [Green Version]
- Terman, J.S.; Terman, M.; Lo, E.S.; Cooper, T.B. Circadian time of morning light administration and therapeutic response in winter depression. Arch. Gen. Psychiatry Res. 2001, 58, 69–75. [Google Scholar] [CrossRef]
- Juda, M.; Vetter, C.; Roenneberg, T. The Munich chronotype questionnaire for shift-workers (MCTQShift). J. Biol. Rhythms 2013, 28, 130–140. [Google Scholar] [CrossRef]
- Ghotbi, N.; Pilz, L.K.; Winnebeck, E.; Vetter, C.; Zerbini, G.; Lenssen, D.; Frighetto, G.; Salamanca, M.; Costa, R.; Montagnese, S.; et al. The µMCTQ—An ultra-short version of the Munich ChronoType Questionnaire. in press.
- Jones, C.R.; Campbell, S.S.; Zone, S.E.; Cooper, F.; DeSano, A.; Murphy, P.J.; Jones, B.; Czajkowski, L.; Ptacek, L.J. Familial advanced sleep-phase syndrome: A short-period circadian rhythm variant in humans. Nat. Med. 1999, 5, 1062–1065. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.E.; Tyrrell, J.; Wood, A.R.; Beaumont, R.N.; Ruth, K.S.; Tuke, M.A.; Yaghootkar, H.; Hu, Y.; Teder-Laving, M.; Hayward, C.; et al. Genome-wide association analyses in 128,266 individuals identifies new morningness and sleep duration loci. PLoS Genet. 2016, 12, e1006125. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Padiath, Q.S.; Shapiro, R.E.; Jones, C.R.; Wu, S.C.; Saigoh, N.; Saigoh, K.; Ptáček, L.J.; Fu, Y.-H. Functional consequences of a CKIδ mutation causing familial advanced sleep phase syndrome. Nature 2005, 434, 640. [Google Scholar] [CrossRef] [PubMed]
- Beale, A.D.; Pedrazzoli, M.; da Silva, B.G.B.; Beijamini, F.; Duarte, N.E.; Egan, K.J.; Knutson, K.L.; von Schantz, M.; Roden, L.C. Comparison between an African town and a neighbouring village shows delayed, but not decreased, sleep during the early stages of urbanisation. Sci. Rep. 2017, 7, 5697. [Google Scholar] [CrossRef]
- Moreno, C.R.C.; Vasconcelos, S.; Marqueze, E.C.; Lowden, A.; Middleton, B.; Fischer, F.M.; Louzada, F.M.; Skene, D.J. Sleep patterns in Amazon rubber tappers with and without electric light at home. Sci. Rep. 2015, 5, 14074. [Google Scholar] [CrossRef]
- Pilz, L.K.; Levandovski, R.; Oliveira, M.A.B.; Hidalgo, M.P.; Roenneberg, T. Sleep and light exposure across different levels of urbanisation in Brazilian communities. Sci. Rep. 2018, 8, 11389. [Google Scholar] [CrossRef]
- De la Iglesia, H.O.; Fernández-Duque, E.; Golombek, D.A.; Lanza, N.; Duffy, J.F.; Czeisler, C.A.; Valeggia, C.R. Access to electric light is associated with shorter sleep duration in a traditionally hunter-gatherer community. J. Biol. Rhythms 2015, 30, 342–350. [Google Scholar] [CrossRef]
- Borisenkov, M.F.; Perminova, E.V.; Kosova, A.L. Chronotype, sleep length, and school achievement of 11-to 23-year-old students in northern European Russia. Chronobiol. Int. 2010, 27, 1259–1270. [Google Scholar] [CrossRef]
- Carvalho, F.G.; Hidalgo, M.P.; Levandovski, R. Differences in circadian patterns between rural and urban populations: An epidemiological study in countryside. Chronobiol. Int. 2014, 31, 442–449. [Google Scholar] [CrossRef]
- Roenneberg, T.; Kumar, C.J.; Merrow, M. The human circadian clock entrains to sun time. Curr. Biol. CB 2007, 17, R44–R45. [Google Scholar] [CrossRef] [Green Version]
- Haraszti, R.Á.; Ella, K.; Gyöngyösi, N.; Roenneberg, T.; Káldi, K. Social jetlag negatively correlates with academic performance in undergraduates. Chronobiol. Int. 2014, 31, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Allebrandt, K.V.; Teder-Laving, M.; Kantermann, T.; Peters, A.; Campbell, H.; Rudan, I.; Wilson, J.F.; Metspalu, A.; Roenneberg, T. Chronotype and sleep duration: The influence of season of assessment. Chronobiol. Int. 2014, 31, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Borisenkov, M.F. The pattern of entrainment of the human sleep-wake rhythm by the natural photoperiod in the north. Chronobiol. Int. 2011, 28, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Kantermann, T.; Juda, M.; Merrow, M.; Roenneberg, T. The human circadian clock’s seasonal adjustment is disrupted by daylight saving time. Curr. Biol. 2007, 17, 1996–2000. [Google Scholar] [CrossRef] [PubMed]
- Fischer, D.; Lombardi, D.A.; Marucci-Wellman, H.; Roenneberg, T. Chronotypes in the US–influence of age and sex. PLoS ONE 2017, 12, e0178782. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Kuehnle, T.; Pramstaller, P.P.; Ricken, J.; Havel, M.; Guth, A.; Merrow, M. A marker for the end of adolescence. Curr. Biol. 2004, 14, R1038–R1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roenneberg, T.; Keller, L.K.; Fischer, D.; Matera, J.L.; Vetter, C.; Winnebeck, E.C. Human activity and rest in situ. Methods Enzymol. 2015, 552, 257–283. [Google Scholar]
- Crowley, S.J.; Cain, S.W.; Burns, A.C.; Acebo, C.; Carskadon, M.A. Increased sensitivity of the circadian system to light in early/mid-puberty. J. Clin. Endocrinol. Metab. 2015, 100, 4067–4073. [Google Scholar] [CrossRef]
- Hagenauer, M.H.; Lee, T.M. The neuroendocrine control of the circadian system: Adolescent chronotype. Front. Neuroendocrinol. 2012, 33, 211–229. [Google Scholar] [CrossRef] [Green Version]
- Samson, D.R.; Crittenden, A.N.; Mabulla, I.A.; Mabulla, A.Z.P.; Nunn, C.L. Chronotype variation drives night-time sentinel-like behaviour in hunter-gatherers. Proc. Biol. Sci. 2017, 284, 20170967. [Google Scholar] [CrossRef]
- Kantermann, T.; Sung, H.; Burgess, H.J. Comparing the morningness-eveningness questionnaire and Munich chronotype questionnaire to the dim light melatonin onset. J. Biol. Rhythms 2015, 30, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.; Hida, A.; Aritake, S.; Higuchi, S.; Enomoto, M.; Kato, M.; Vetter, C.; Roenneberg, T.; Mishima, K. Validity of the Japanese version of the Munich chronotype questionnaire. Chronobiol. Int. 2014, 31, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Facer-Childs, E.R.; Campos, B.M.; Middleton, B.; Skene, D.J.; Bagshaw, A.P. Circadian phenotype impacts the brain’s resting-state functional connectivity, attentional performance, and sleepiness. Sleep 2019, 42, zsz033. [Google Scholar] [CrossRef] [PubMed]
- Santisteban, J.A.; Brown, T.G.; Gruber, R. Association between the Munich chronotype questionnaire and wrist actigraphy. Sleep Disord. 2018, 2018, 5646848. [Google Scholar] [CrossRef] [PubMed]
- Kühnle, T. Quantitative Analysis of Human Chronotypes. Ph.D. Thesis, Ludwig-Maximilians-Universität München, München, Germany, 2006. [Google Scholar]
- Keijzer, H.; Smits, M.G.; Duffy, J.F.; Curfs, L.M.G. Why the dim light melatonin onset (DLMO) should be measured before treatment of patients with circadian rhythm sleep disorders. Sleep Med. Rev. 2014, 18, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Braun, R.; Kath, W.L.; Iwanaszko, M.; Kula-Eversole, E.; Abbott, S.M.; Reid, K.J.; Zee, P.C.; Allada, R. Universal method for robust detection of circadian state from gene expression. Proc. Natl. Acad. Sci. USA 2018, 115, E9247–E9256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laing, E.E.; Möller-Levet, C.S.; Poh, N.; Santhi, N.; Archer, S.N.; Dijk, D.-J. Blood transcriptome based biomarkers for human circadian phase. eLife 2017, 6, e20214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittenbrink, N.; Ananthasubramaniam, B.; Münch, M.; Koller, B.; Maier, B.; Weschke, C.; Bes, F.; de Zeeuw, J.; Nowozin, C.; Wahnschaffe, A.; et al. High-accuracy determination of internal circadian time from a single blood sample. J. Clin. Investig. 2018, 128, 3826–3839. [Google Scholar] [CrossRef]
- Zavada, A.; Gordijn, M.C.M.; Beersma, D.G.M.; Daan, S.; Roenneberg, T. Comparison of the Munich chronotype questionnaire with the Horne-Ostberg’s morningness-eveningness score. Chronobiol. Int. 2005, 22, 267–278. [Google Scholar] [CrossRef]
- Da Silva Sinha, V. Event-based time in three indigenous Amazonian and Xinguan cultures and languages. Front. Psychol. 2019, 10, 454. [Google Scholar] [CrossRef]
- Sinha, C.; Sinha, V.D.S.; Zinken, J.; Sampaio, W. When time is not space: The social and linguistic construction of time intervals and temporal event relations in an Amazonian culture. Lang. Cogn. 2011, 3, 137–169. [Google Scholar] [CrossRef]
- Borbely, A.A. A two process model of sleep regulation. Hum. Neurobiol. 1982, 1, 195–204. [Google Scholar] [PubMed]
- Steyer, R.; Ferring, D.; Schmitt, M.J. States and traits in psychological assessment. Eur. J. Psychol. Assess. 1992, 8, 79–98. [Google Scholar]
- Gehrman, P.R.; Ghorai, A.; Goodman, M.; McCluskey, R.; Barilla, H.; Almasy, L.; Roenneberg, T.; Bucan, M. Twin-based heritability of actimetry traits. Genes Brain Behav. 2019, 18, e12569. [Google Scholar] [CrossRef] [PubMed]
- Facer-Childs, E.R.; Middleton, B.; Skene, D.J.; Bagshaw, A.P. Resetting the late timing of “night owls” has a positive impact on mental health and performance. Sleep Med. 2019, 60, 236–247. [Google Scholar] [CrossRef]
- Nag, C.; Pradhan, R.K. Impact of lifestyle on circadian orientation and sleep behaviour. Sleep Biol. Rhythms 2012, 10, 94–99. [Google Scholar] [CrossRef]
- Von Schantz, M.; Taporoski, T.P.; Horimoto, A.R.V.R.; Duarte, N.E.; Vallada, H.; Krieger, J.E.; Pedrazzoli, M.; Negrão, A.B.; Pereira, A.C. Distribution and heritability of diurnal preference (chronotype) in a rural Brazilian family-based cohort, the Baependi study. Sci. Rep. 2015, 5, 9214. [Google Scholar] [CrossRef] [Green Version]
- Zerbini, G.; Kantermann, T.; Merrow, M. Strategies to decrease social jetlag: Reducing evening blue light advances sleep and melatonin. Eur. J. Neurosci. 2018. [Google Scholar] [CrossRef]
- Anothaisintawee, T.; Lertrattananon, D.; Thamakaison, S.; Knutson, K.L.; Thakkinstian, A.; Reutrakul, S. Later chronotype is associated with higher hemoglobin A1c in prediabetes patients. Chronobiol. Int. 2017, 34, 393–402. [Google Scholar] [CrossRef]
- Randler, C.; Haun, J.; Schaal, S. Assessing the influence of sleep-wake variables on body mass index (BMI) in adolescents. Eur. J. Psychol. 2013, 9, 339–347. [Google Scholar] [CrossRef]
- Antypa, N.; Vogelzangs, N.; Meesters, Y.; Schoevers, R.; Penninx, B.W.J.H. Chronotype associations with depression and anxiety disorders in a large cohort study. Depress. Anxiety 2016, 33, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Levandovski, R.; Dantas, G.; Fernandes, L.C.; Caumo, W.; Torres, I.; Roenneberg, T.; Hidalgo, M.P.L.; Allebrandt, K.V. Depression scores associate with chronotype and social jetlag in a rural population. Chronobiol. Int. 2011, 28, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Pilz, L.K.; Keller, L.K.; Lenssen, D.; Roenneberg, T. Time to rethink sleep quality: PSQI scores reflect sleep quality on workdays. Sleep 2018, 41, zsy029. [Google Scholar] [CrossRef] [PubMed]
- Borbély, A.A.; Daan, S.; Wirz-Justice, A.; Deboer, T. The two-process model of sleep regulation: A reappraisal. J. Sleep Res. 2016, 25, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, J.; Reilly, T.; Atkinson, G.; Edwards, B. Jet lag: Trends and coping strategies. Lancet Lond. Engl. 2007, 369, 1117–1129. [Google Scholar] [CrossRef]
- Stokkan, K.-A.; Yamazaki, S.; Tei, H.; Sakaki, Y.; Menaker, M. Entrainment of the circadian clock in the liver by feeding. Science 2001, 291, 490–493. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, S.; Numano, R.; Abe, M.; Hida, A.; Takahashi, R.; Ueda, M.; Block, G.D.; Sakaki, Y.; Menaker, M.; Tei, H. Resetting central and peripheral circadian oscillators in transgenic rats. Science 2000, 288, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Vetter, C. Circadian disruption: What do we actually mean? Eur. J. Neurosci. 2018. [Google Scholar] [CrossRef]
- De Zeeuw, J.; Wisniewski, S.; Papakonstantinou, A.; Bes, F.; Wahnschaffe, A.; Zaleska, M.; Kunz, D.; Münch, M. The alerting effect of the wake maintenance zone during 40 hours of sleep deprivation. Sci. Rep. 2018, 8, 11012. [Google Scholar] [CrossRef]
- Silva, C.M.; Mota, M.C.; Miranda, M.T.; Paim, S.L.; Waterhouse, J.; Crispim, C.A. Chronotype, social jetlag and sleep debt are associated with dietary intake among Brazilian undergraduate students. Chronobiol. Int. 2016, 33, 740–748. [Google Scholar] [CrossRef]
- Randler, C.; Vollmer, C.; Kalb, N.; Itzek-Greulich, H. Breakpoints of time in bed, midpoint of sleep, and social jetlag from infancy to early adulthood. Sleep Med. 2019, 57, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Roenneberg, T.; Allebrandt, K.V.; Merrow, M.; Vetter, C. Social jetlag and obesity. Curr. Biol. CB 2012, 22, 939–943. [Google Scholar] [CrossRef] [PubMed]
- Almoosawi, S.; Palla, L.; Walshe, I.; Vingeliene, S.; Ellis, J.G. Long sleep duration and social jetlag are associated inversely with a healthy dietary pattern in adults: Results from the UK national diet and nutrition survey rolling programme Y1–4. Nutrients 2018, 10, 1131. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Morales, J.F.; Escribano, C. Social jetlag, academic achievement and cognitive performance: Understanding gender/sex differences. Chronobiol. Int. 2015, 32, 822–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randler, C.; Vollmer, C. Aggression in young adults—A matter of short sleep and social jetlag? Psychol. Rep. 2013, 113, 754–765. [Google Scholar] [CrossRef] [PubMed]
- Alves, M.S.; Andrade, R.Z.; Silva, G.C.; Mota, M.C.; Resende, S.G.; Teixeira, K.R.; Gonçalves, B.F.; Crispim, C.A. Social jetlag among night workers is negatively associated with the frequency of moderate or vigorous physical activity and with energy expenditure related to physical activity. J. Biol. Rhythms 2017, 32, 83–93. [Google Scholar] [CrossRef]
- Koopman, A.D.M.; Rauh, S.P.; van ‘t Riet, E.; Groeneveld, L.; van der Heijden, A.A.; Elders, P.J.; Dekker, J.M.; Nijpels, G.; Beulens, J.W.; Rutters, F. The association between social jetlag, the metabolic syndrome, and type 2 diabetes mellitus in the general population: The new Hoorn study. J. Biol. Rhythms 2017, 32, 359–368. [Google Scholar] [CrossRef]
- Malone, S.K.; Zemel, B.; Compher, C.; Souders, M.; Chittams, J.; Thompson, A.L.; Pack, A.; Lipman, T.H. Social jetlag, chronotype, and body mass index in 14 to 17 year old adolescents. Chronobiol. Int. 2016, 33, 1255–1266. [Google Scholar] [CrossRef]
- Parsons, M.J.; Moffitt, T.E.; Gregory, A.M.; Goldman-Mellor, S.; Nolan, P.M.; Poulton, R.; Caspi, A. Social jetlag, obesity and metabolic disorder: Investigation in a cohort study. Int. J. Obes. 2015, 39, 842–848. [Google Scholar] [CrossRef]
- Rutters, F.; Lemmens, S.G.; Adam, T.C.; Bremmer, M.A.; Elders, P.J.; Nijpels, G.; Dekker, J.M. Is social jetlag associated with an adverse endocrine, behavioral, and cardiovascular risk profile? J. Biol. Rhythms 2014, 29, 377–383. [Google Scholar] [CrossRef]
- Wong, P.M.; Hasler, B.P.; Kamarck, T.W.; Muldoon, M.F.; Manuck, S.B. Social jetlag, chronotype, and cardiometabolic risk. J. Clin. Endocrinol. Metab. 2015, 100, 4612–4620. [Google Scholar] [CrossRef] [PubMed]
- Borisenkov, M.F.; Petrova, N.B.; Timonin, V.D.; Fradkova, L.I.; Kolomeichuk, S.N.; Kosova, A.L.; Kasyanova, O.N. Sleep characteristics, chronotype and winter depression in 10-20-year-olds in northern European Russia. J. Sleep Res. 2015, 24, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Polugrudov, A.S.; Panev, A.S.; Smirnov, V.V.; Paderin, N.M.; Borisenkov, M.F.; Popov, S.V. Wrist temperature and cortisol awakening response in humans with social jetlag in the North. Chronobiol. Int. 2016, 33, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Schimitt, R.; Levandovski, R.; Hidalgo, M.P.L. Relations between social rhythm, sleep phase, and minor psychiatric symptoms in healthy workers. Biol. Rhythm Res. 2013, 44, 403–409. [Google Scholar] [CrossRef]
- Sheaves, B.; Porcheret, K.; Tsanas, A.; Espie, C.A.; Foster, R.G.; Freeman, D.; Harrison, P.J.; Wulff, K.; Goodwin, G.M. Insomnia, nightmares, and chronotype as markers of risk for severe mental illness: Results from a student population. Sleep 2016, 39, 173–181. [Google Scholar] [CrossRef] [PubMed]
- De Souza, C.M.; Hidalgo, M.P.L. Midpoint of sleep on school days is associated with depression among adolescents. Chronobiol. Int. 2014, 31, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Knapen, S.E.; Riemersma-van der Lek, R.F.; Antypa, N.; Meesters, Y.; Penninx, B.W.J.H.; Schoevers, R.A. Social jetlag and depression status: Results obtained from the Netherlands study of depression and anxiety. Chronobiol. Int. 2018, 35, 1–7. [Google Scholar] [CrossRef]
- Burgess, H.J.; Eastman, C.I. A late wake time phase delays the human dim light melatonin rhythm. Neurosci. Lett. 2006, 395, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Crowley, S.J.; Carskadon, M.A. Modifications to weekend recovery sleep delay circadian phase in older adolescents. Chronobiol. Int. 2010, 27, 1469–1492. [Google Scholar] [CrossRef]
- Stothard, E.R.; McHill, A.W.; Depner, C.M.; Birks, B.R.; Moehlman, T.M.; Ritchie, H.K.; Guzzetti, J.R.; Chinoy, E.D.; LeBourgeois, M.K.; Axelsson, J.; et al. Circadian entrainment to the natural light-dark cycle across seasons and the weekend. Curr. Biol. 2017, 27, 508–513. [Google Scholar] [CrossRef]
- Vondrasova-Jelinkova, D.; Hajek, I.; Illnerova, H. Adjustment of the human melatonin and cortisol rhythms to shortening of the natural summer photoperiod. Brain Res 1999, 816, 249–253. [Google Scholar] [CrossRef]
- Zerbini, G. Conflicted Clocks: Social Jetlag, Entrainment and the Role of Chronotype: From Physiology to Academic Performance; From Students to Working Adults. Ph.D. Thesis, University of Groningen, Groningen, The Netherlands, 2017. [Google Scholar]
- Hadlow, N.; Brown, S.; Wardrop, R.; Conradie, J.; Henley, D. Where in the world? Latitude, longitude and season contribute to the complex co-ordinates determining cortisol levels. Clin. Endocrinol. (Oxf.) 2018, 89, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Randler, C. Differences in sleep and circadian preference between Eastern and Western German adolescents. Chronobiol. Int. 2008, 25, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, K.S. Social jet lag: Sleep-corrected formula. Chronobiol. Int. 2017, 34, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Depner, C.M.; Melanson, E.L.; Eckel, R.H.; Snell-Bergeon, J.K.; Perreault, L.; Bergman, B.C.; Higgins, J.A.; Guerin, M.K.; Stothard, E.R.; Morton, S.J.; et al. Ad libitum weekend recovery sleep fails to prevent metabolic dysregulation during a repeating pattern of insufficient sleep and weekend recovery sleep. Curr. Biol. 2019, 29, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Åkerstedt, T.; Ghilotti, F.; Grotta, A.; Zhao, H.; Adami, H.-O.; Trolle-Lagerros, Y.; Bellocco, R. Sleep duration and mortality—Does weekend sleep matter? J. Sleep Res. 2019, 28, e12712. [Google Scholar] [CrossRef]
- Beauvalet, J.C.; Quiles, C.L.; de Oliveira, M.A.B.; Ilgenfritz, C.A.V.; Hidalgo, M.P.L.; Tonon, A.C. Social jetlag in health and behavioral research: A systematic review. ChronoPhysiol. Ther. 2017, 7, 19–31. [Google Scholar] [CrossRef]
- Gill, S.; Panda, S. A smartphone app reveals erratic diurnal eating patterns in humans that can be modulated for health benefits. Cell Metab. 2015, 22, 789–798. [Google Scholar] [CrossRef]
- Zerbini, G.; Merrow, M. Time to learn: How chronotype impacts education. PsyCh J. 2017, 6, 263–276. [Google Scholar] [CrossRef] [Green Version]
- Lévi, F. Circadian chronotherapy for human cancers. Lancet Oncol. 2001, 2, 307–315. [Google Scholar] [CrossRef]
- Peeples, L. Medicine’s secret ingredient—It’s in the timing. Nature 2018, 556, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Selfridge, J.M.; Gotoh, T.; Schiffhauer, S.; Liu, J.; Stauffer, P.E.; Li, A.; Capelluto, D.G.S.; Finkielstein, C.V. Chronotherapy: Intuitive, sound, founded…but not broadly applied. Drugs 2016, 76, 1507–1521. [Google Scholar] [CrossRef]
- Vetter, C.; Fischer, D.; Matera, J.L.; Roenneberg, T. Aligning work and circadian time in shift workers improves sleep and reduces circadian disruption. Curr. Biol. CB 2015, 25, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Van der Vinne, V.; Zerbini, G.; Siersema, A.; Pieper, A.; Merrow, M.; Hut, R.A.; Roenneberg, T.; Kantermann, T. Timing of examinations affects school performance differently in early and late chronotypes. J. Biol. Rhythms 2015, 30, 53–60. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roenneberg, T.; Pilz, L.K.; Zerbini, G.; Winnebeck, E.C. Chronotype and Social Jetlag: A (Self-) Critical Review. Biology 2019, 8, 54. https://doi.org/10.3390/biology8030054
Roenneberg T, Pilz LK, Zerbini G, Winnebeck EC. Chronotype and Social Jetlag: A (Self-) Critical Review. Biology. 2019; 8(3):54. https://doi.org/10.3390/biology8030054
Chicago/Turabian StyleRoenneberg, Till, Luísa K. Pilz, Giulia Zerbini, and Eva C. Winnebeck. 2019. "Chronotype and Social Jetlag: A (Self-) Critical Review" Biology 8, no. 3: 54. https://doi.org/10.3390/biology8030054