
Immunologic, Anti-Inflammatory, and Anti-Muscle Damage Profile of Supplemented Vitamin D3 in Healthy Adults on Strenuous Endurance Exercise
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
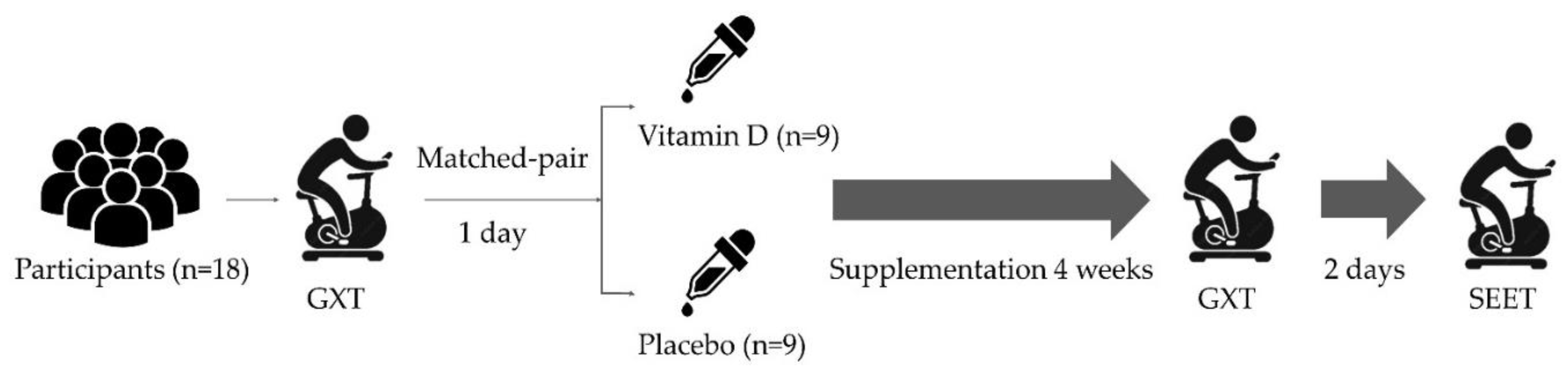
2.2. Experimental Design and Procedure
2.3. Vitamin D3 Supplementation
2.4. GXT Protocol
2.5. Strenuous Endurance Exercise Test (SEET)
2.6. Biochemical Variables
2.7. Statistical Analysis
3. Result
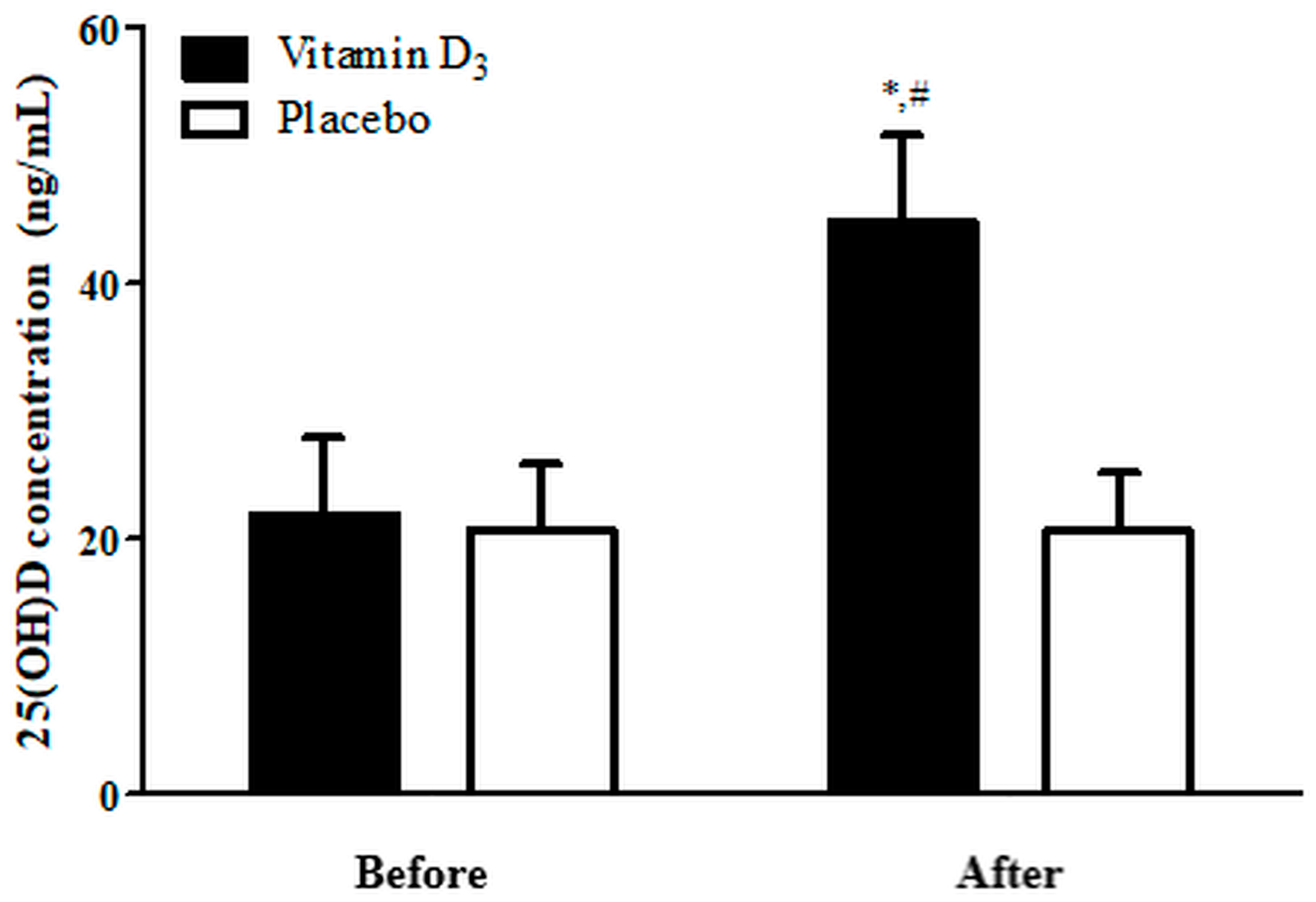
3.1. Blood 25(OH)D Levels
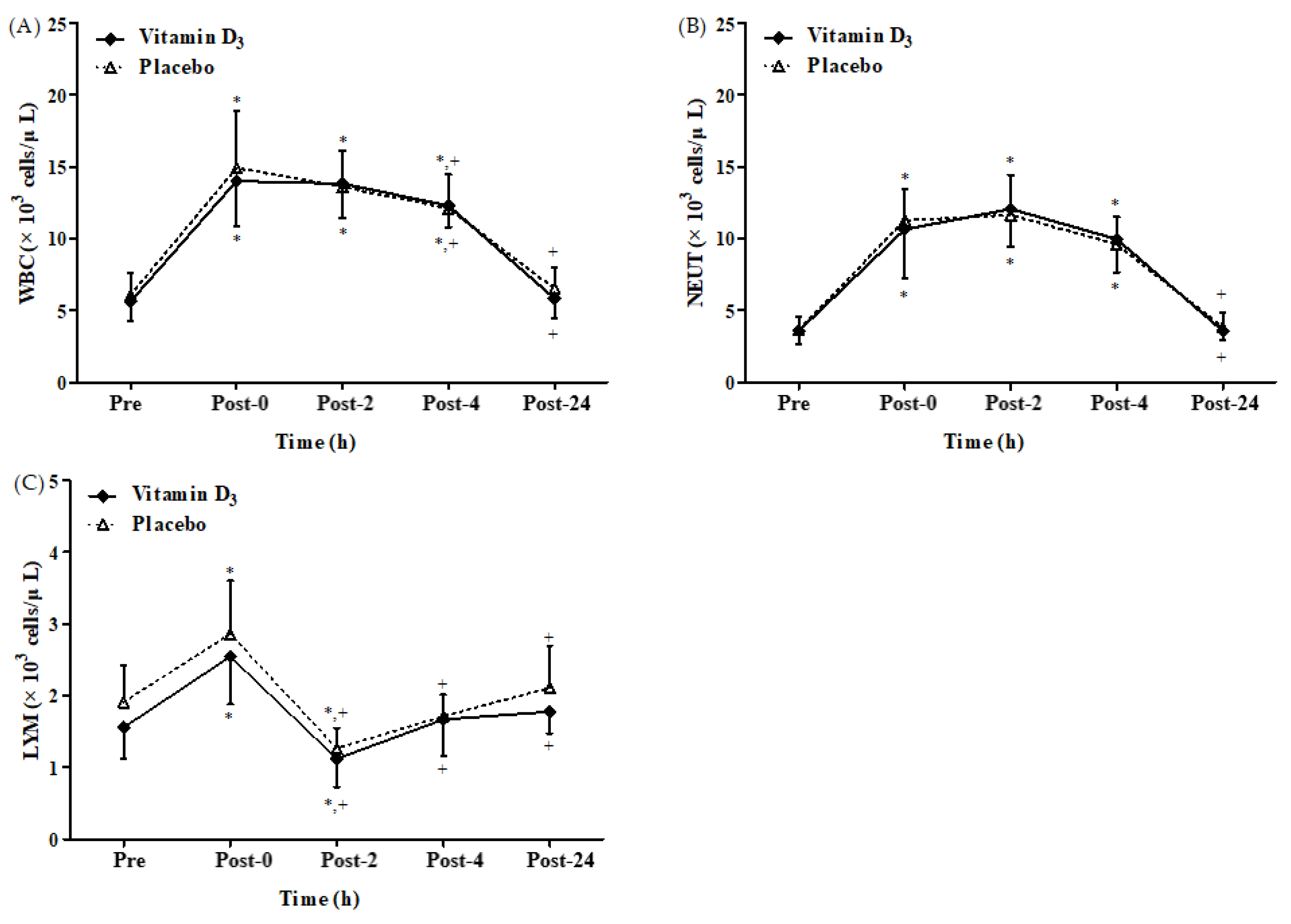
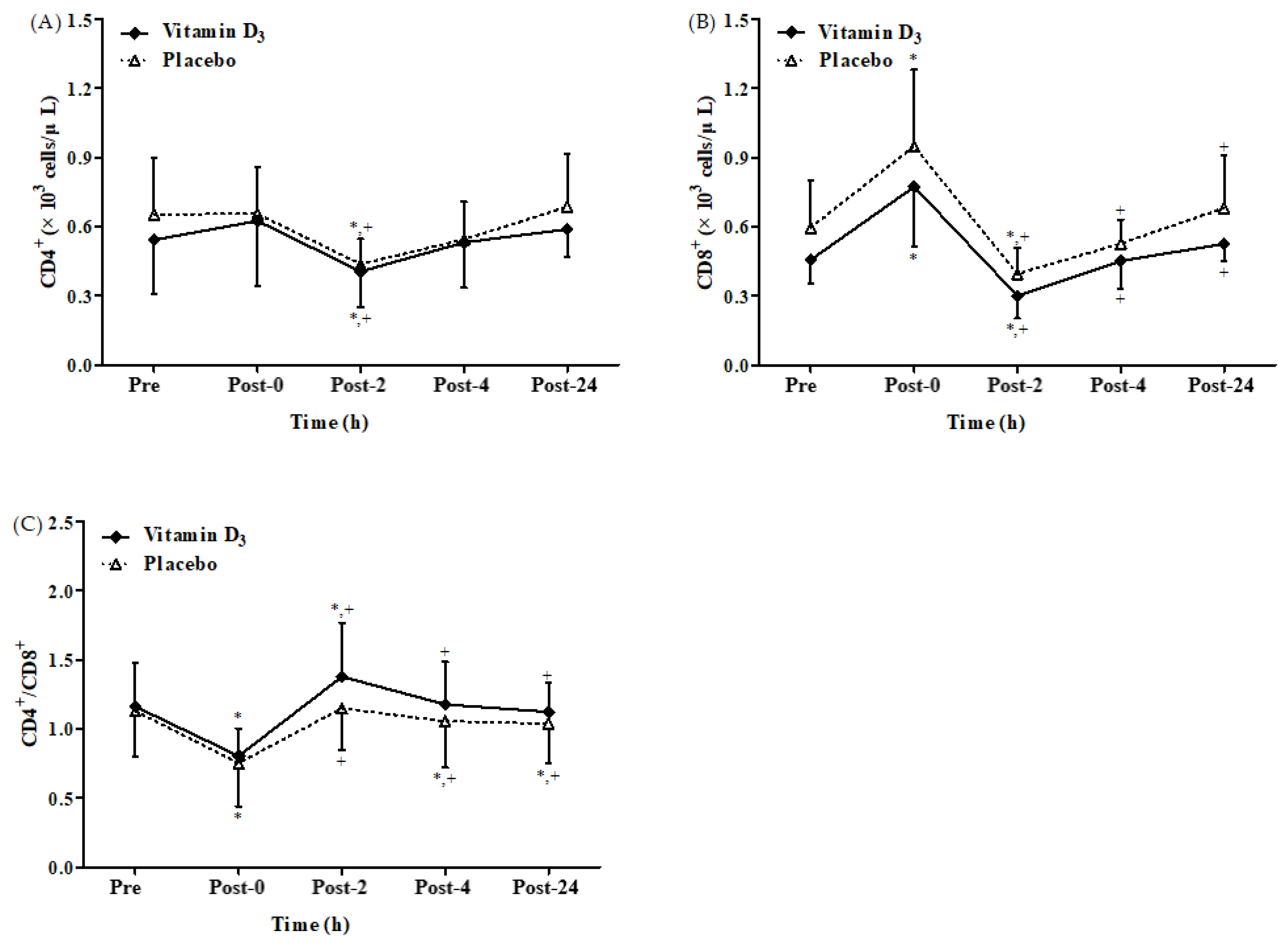
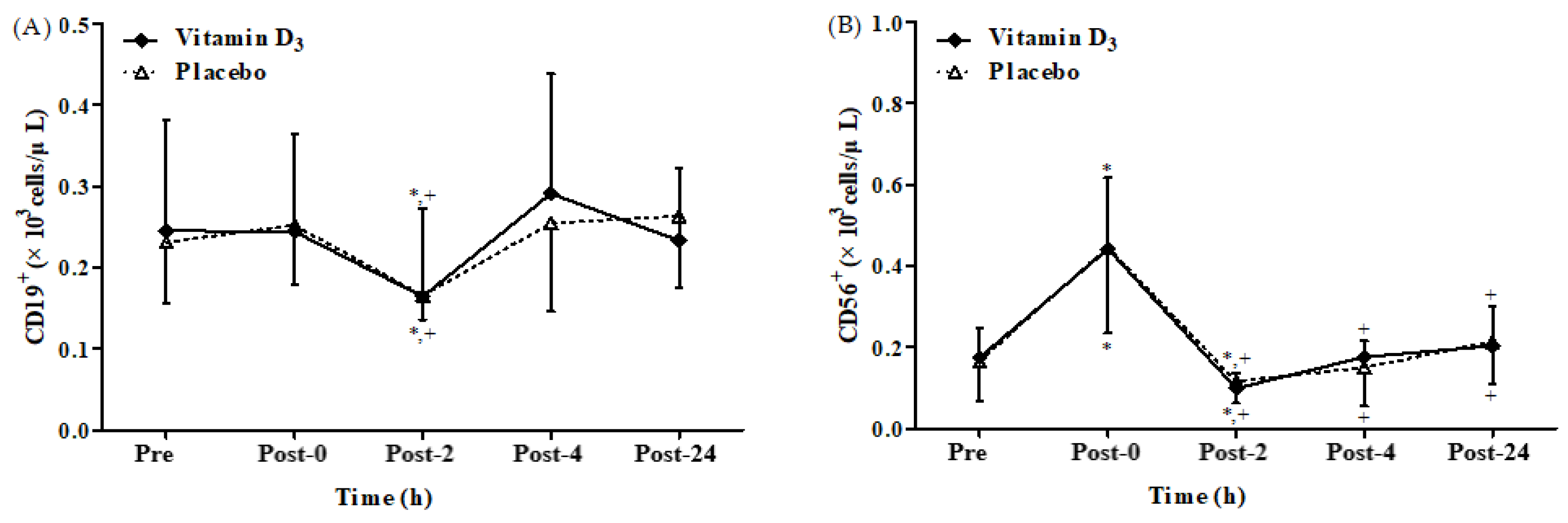
3.2. Effects of Vitamin D3 Supplementation on Immune Response
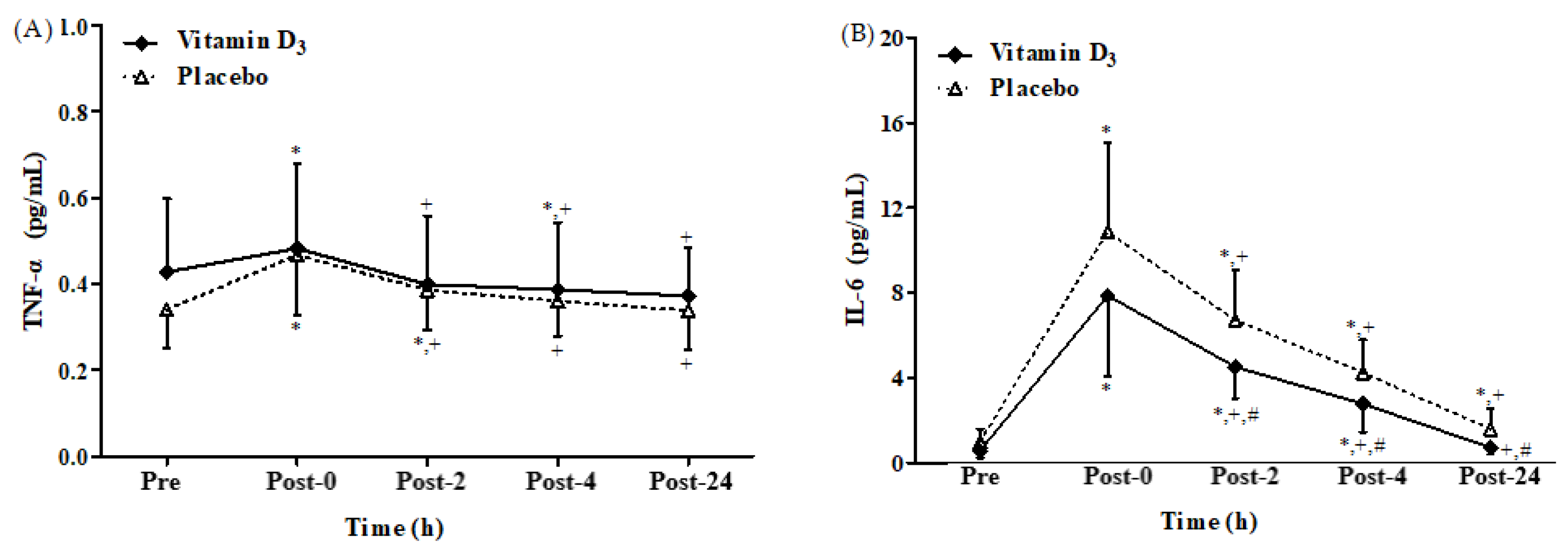
3.3. Effects of Vitamin D3 Supplementation on Inflammatory Cytokines
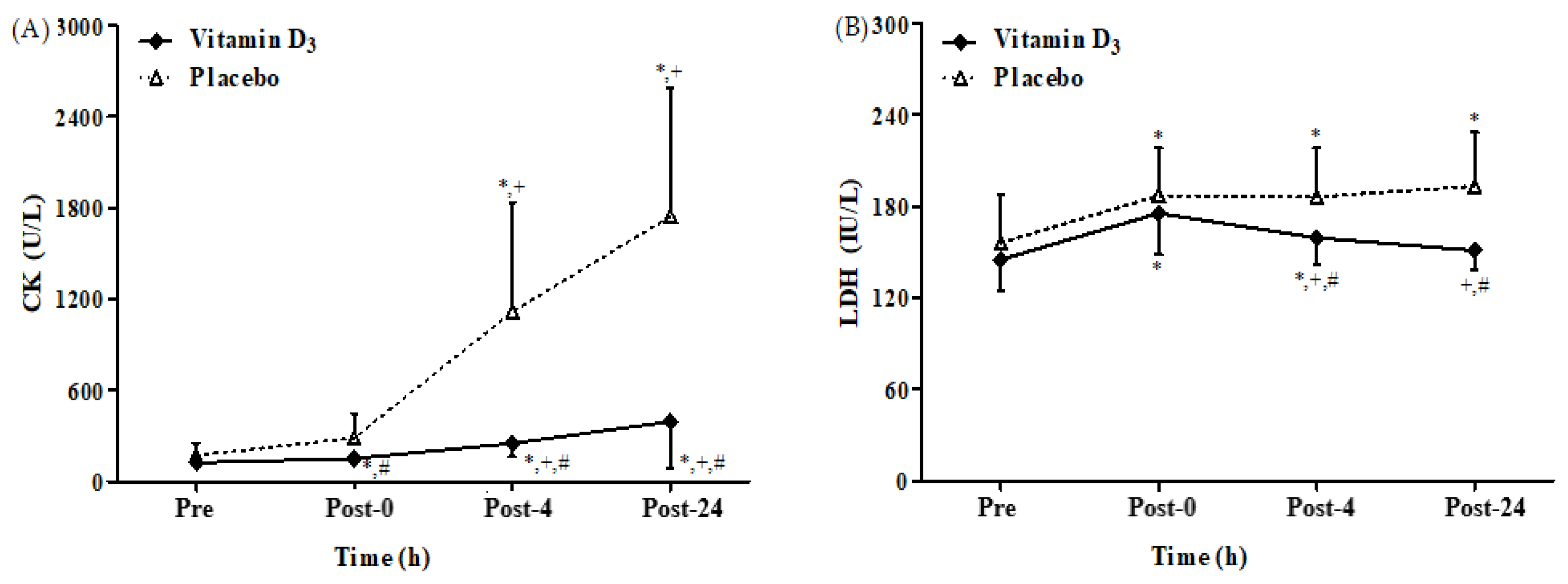
3.4. Effects of Vitamin D3 Supplementation on Muscle Damage
3.5. Impact of Vitamin D3 Supplementation on Aerobic Capacity
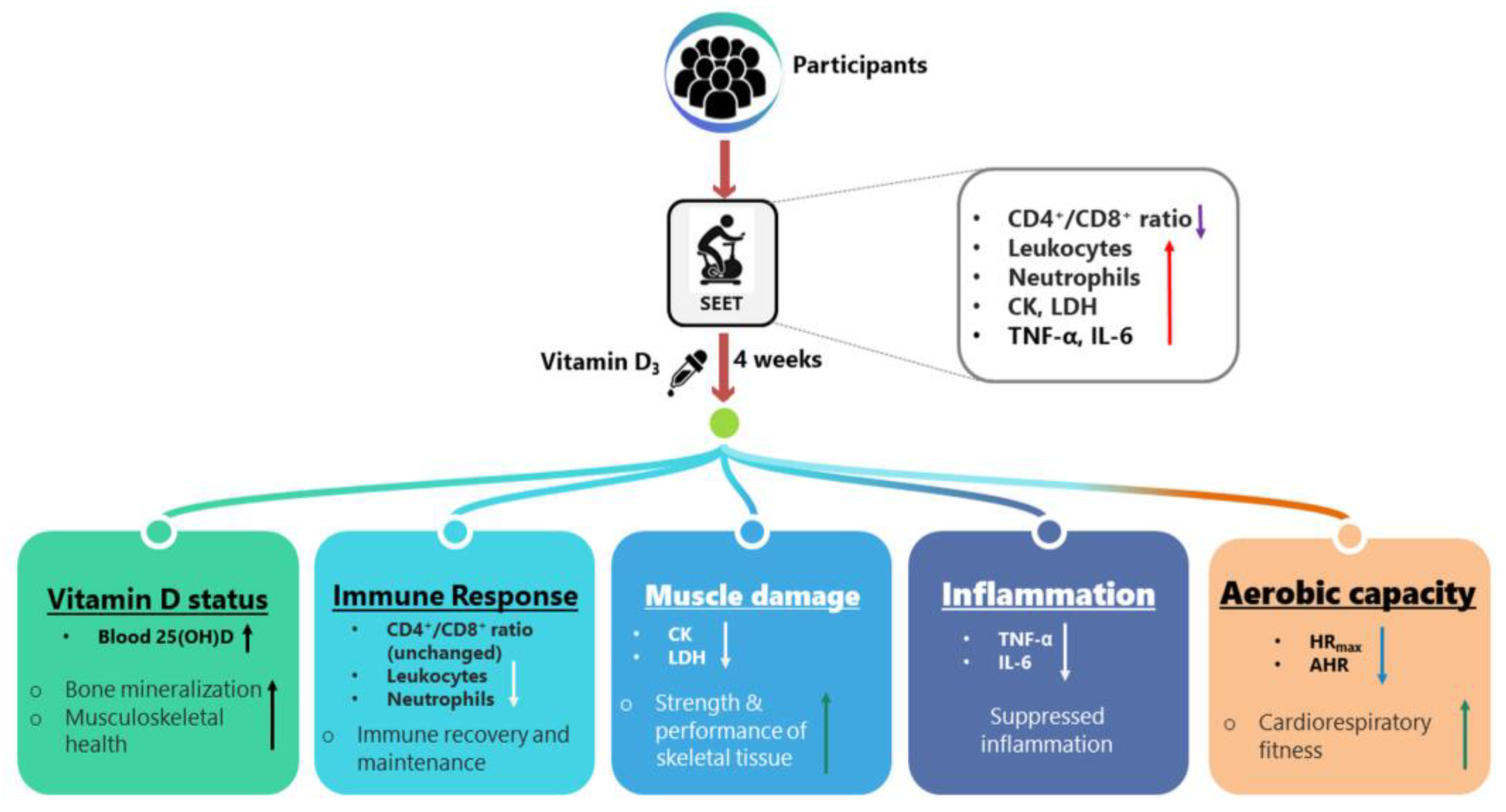
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cerqueira, É.; Marinho, D.A.; Neiva, H.P.; Lourenço, O. Inflammatory Effects of High and Moderate Intensity Exercise—A Systematic Review. Front. Physiol. 2020, 10, 1550. [Google Scholar] [CrossRef]
- Peake, J.M.; Neubauer, O.; Della Gatta, P.A.; Nosaka, K. Muscle damage and inflammation during recovery from exercise. J Appl. Physiol. 2017, 122, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.M.; Neubauer, O.; Walsh, N.P.; Simpson, R.J. Recovery of the immune system after exercise. J. Appl. Physiol. 2017, 122, 1077–1087. [Google Scholar] [CrossRef] [PubMed]
- Walsh, N.P.; Oliver, S.J. Exercise, immune function and respiratory infection: An update on the influence of training and environmental stress. Immunol. Cell Biol. 2016, 94, 132–139. [Google Scholar] [CrossRef]
- Kakanis, M.; Peake, J.; Hooper, S.; Gray, B.; Marshall-Gradisnik, S. The open window of susceptibility to infection after acute exercise in healthy young male elite athletes. J. Sci. Med. Sport 2010, 13, e85–e86. [Google Scholar] [CrossRef]
- Ferrer, M.D.; Tauler, P.; Sureda, A.; Tur, J.A.; Pons, A. Antioxidant regulatory mechanisms in neutrophils and lymphocytes after intense exercise. J. Sport. Sci. 2009, 27, 49–58. [Google Scholar] [CrossRef]
- Kalina, T.; Fišer, K.; Pérez-Andrés, M.; Kuzílková, D.; Cuenca, M.; Bartol, S.J.W.; Blanco, E.; Engel, P.; van Zelm, M.C. CD maps—Dynamic profiling of CD1–CD100 surface expression on human leukocyte and lymphocyte subsets. Front. Immunol. 2019, 10, 2434. [Google Scholar] [CrossRef]
- Garrido-Rodríguez, V.; Herrero-Fernández, I.; Castro, M.J.; Castillo, A.; Rosado-Sánchez, I.; Galvá, M.I.; Ramos, R.; Olivas-Martínez, I.; Bulnes-Ramos, Á.; Cañizares, J.; et al. Immunological features beyond CD4/CD8 ratio values in older individuals. Aging 2021, 13, 13443–13459. [Google Scholar] [CrossRef]
- Pedersen BK, H.-G.L. Exercise and the immune system: Regulation, integration, and adaptation. Physiol Rev. 2000, 80, 1055–1081. [Google Scholar] [CrossRef]
- Mooren, F.C.; Lechtermann, A.; Fromme, A.; Thorwesten, L.; Völker, K. Alterations in intracellular calcium signaling of lymphocytes after exhaustive exercise. Med. Sci. Sport. Exerc. 2001, 33, 242–248. [Google Scholar] [CrossRef]
- Shek, P.N.; Sabiston, B.; Buguet, A.; Radomski, M. Strenuous exercise and immunological changes: A multiple-time-point analysis of leukocyte subsets, CD4/CD8 ratio, immunoglobulin production and NK cell response. Int. J. Sport. Med. 1995, 16, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Inflammation 2010: New adventures of an old flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef]
- Markworth, J.F.; Maddipati, K.R.; Cameron-Smith, D. Emerging roles of pro-resolving lipid mediators in immunological and adaptive responses to exercise-induced muscle injury. Exerc. Immunol. Rev. 2016, 22, 110–134. [Google Scholar] [PubMed]
- Fischer, C.P. Interleukin-6 in acute exercise and training: What is the biological relevance? Exerc Immunol Rev 2006, 12, 6–33. [Google Scholar]
- Suzuki, K. Cytokine response to exercise and its modulation. Antioxidants 2018, 7, 17. [Google Scholar] [CrossRef]
- Usui, T.; Yoshikawa, T.; Ueda, S.-Y.; Katsura, Y.; Orita, K.; Fujimoto, S. Effects of acute prolonged strenuous exercise on the salivary stress markers and inflammatory cytokines. Jpn. J. Phys. Fit. Sport. Med. 2011, 60, 295–304. [Google Scholar] [CrossRef]
- Sachdev, S.; Davies, K.J. Production, detection, and adaptive responses to free radicals in exercise. Free Radic. Biol. Med. 2008, 44, 215–223. [Google Scholar] [CrossRef]
- Nie, J.; Tong, T.K.; George, K.; Fu, F.H.; Lin, H.; Shi, Q. Resting and post-exercise serum biomarkers of cardiac and skeletal muscle damage in adolescent runners. Scand. J. Med. Sci. Sport. 2011, 21, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Kennel, K.A.; Drake, M.T.; Hurley, D.L. Vitamin D deficiency in adults: When to test and how to treat. Mayo Clin. Proc. 2010, 85, 752–758. [Google Scholar] [CrossRef]
- Wiciński, M.; Adamkiewicz, D.; Adamkiewicz, M.; Śniegocki, M.; Podhorecka, M.; Szychta, P.; Malinowski, B. Impact of vitamin D on physical efficiency and exercise performance—A review. Nutrients 2019, 11, 2826. [Google Scholar] [CrossRef]
- Maughan, R.J.; Burke, L.M.; Dvorak, J.; Larson-Meyer, D.E.; Peeling, P.; Phillips, S.M.; Rawson, E.S.; Walsh, N.P.; Garthe, I.; Geyer, H.; et al. IOC consensus statement: Dietary supplements and the high-performance athlete. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 104–125. [Google Scholar] [CrossRef] [PubMed]
- Farrokhyar, F.; Tabasinejad, R.; Dao, D.; Peterson, D.; Ayeni, O.R.; Hadioonzadeh, R.; Bhandari, M. Prevalence of vitamin D inadequacy in athletes: A systematic-review and meta-analysis. Sport. Med. 2015, 45, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.; Maseeh, A. Vitamin D: The “sunshine” vitamin. J. Pharm. Pharm. 2012, 3, 118–126. [Google Scholar] [CrossRef]
- Barcal, J.N.; Thomas, J.T.; Hollis, B.W.; Austin, K.J.; Alexander, B.M.; Larson-Meyer, D.E. Vitamin D and weight cycling: Impact on injury, illness, and inflammation in collegiate wrestlers. Nutrients 2016, 8, 775. [Google Scholar] [CrossRef] [PubMed]
- Dubnov-Raz, G.; Hemilä, H.; Cohen, A.H.; Rinat, B.; Choleva, L.; Constantini, N.W. Vitamin D supplementation and upper respiratory tract infections in adolescent swimmers: A randomized controlled trial. Pediatr. Exerc. Sci. 2015, 27, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, B.; Whiteley, R.; Farooq, A.; Chalabi, H. Vitamin D concentration in 342 professional football players and association with lower limb isokinetic function. J. Sci. Med. Sport 2014, 17, 139–143. [Google Scholar] [CrossRef]
- Chen, S.; Sims, G.P.; Chen, X.X.; Gu, Y.Y.; Chen, S.; Lipsky, P.E. Modulatory effects of 1,25-dihydroxyvitamin D3 on human B cell differentiation. J. Immunol. 2007, 179, 1634–1647. [Google Scholar] [CrossRef]
- Boonstra, A.; Barrat, F.J.; Crain, C.; Heath, V.L.; Savelkoul, H.F.J.; O’Garra, A. 1α,25-Dihydroxyvitamin D3 Has a Direct Effect on Naive CD4+ T Cells to Enhance the Development of Th2 Cells1. J. Immunol. 2001, 167, 4974–4980. [Google Scholar] [CrossRef]
- Daniel, C.; Sartory, N.A.; Zahn, N.; Radeke, H.H.; Stein, J.M. Immune modulatory treatment of trinitrobenzene sulfonic acid colitis with calcitriol is associated with a change of a T helper (Th) 1/Th17 to a Th2 and regulatory T cell profile. J. Pharm. Exp. 2008, 324, 23–33. [Google Scholar] [CrossRef]
- Barrat, F.J.; Cua, D.J.; Boonstra, A.; Richards, D.F.; Crain, C.; Savelkoul, H.F.; de Waal-Malefyt, R.; Coffman, R.L.; Hawrylowicz, C.M.; O’Garra, A. In vitro generation of interleukin 10-producing regulatory CD4(+) T cells is induced by immunosuppressive drugs and inhibited by T helper type 1 (Th1)- and Th2-inducing cytokines. J. Exp. Med. 2002, 195, 603–616. [Google Scholar] [CrossRef] [PubMed]
- Gorman, S.; Kuritzky, L.A.; Judge, M.A.; Dixon, K.M.; McGlade, J.P.; Mason, R.S.; Finlay-Jones, J.J.; Hart, P.H. Topically Applied 1,25-Dihydroxyvitamin D3 Enhances the Suppressive Activity of CD4+CD25+ Cells in the Draining Lymph Nodes1. J. Immunol. 2007, 179, 6273–6283. [Google Scholar] [CrossRef]
- Aranow, C. Vitamin D and the immune system. J. Investig. Med. 2011, 59, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Almerighi, C.; Sinistro, A.; Cavazza, A.; Ciaprini, C.; Rocchi, G.; Bergamini, A. 1α,25-Dihydroxyvitamin D3 inhibits CD40L-induced pro-inflammatory and immunomodulatory activity in Human Monocytes. Cytokine 2009, 45, 190–197. [Google Scholar] [CrossRef] [PubMed]
- He, C.S.; Fraser, W.D.; Tang, J.; Brown, K.; Renwick, S.; Rudland-Thomas, J.; Teah, J.; Tanqueray, E.; Gleeson, M. The effect of 14 weeks of vitamin D3 supplementation on antimicrobial peptides and proteins in athletes. J. Sport. Sci. 2016, 34, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.C.; Seo, M.W.; Lee, S.; Jung, S.W.; Song, J.K. Correcting vitamin D insufficiency improves some but not all aspects of physical performance during winter training in taekwondo athletes. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 635–643. [Google Scholar] [CrossRef]
- Zebrowska, A.; Sadowska-Krepa, E.; Stanula, A.; Waskiewicz, Z.; Lakomy, O.; Bezuglov, E.; Nikolaidis, P.T.; Rosemann, T.; Knechtle, B. The effect of vitamin D supplementation on serum total 25(OH) levels and biochemical markers of skeletal muscles in runners. J. Int. Soc. Sport. Nutr. 2020, 17, 18. [Google Scholar] [CrossRef]
- Kasprowicz, K.; Ratkowski, W.; Wolyniec, W.; Kaczmarczyk, M.; Witek, K.; Zmijewski, P.; Renke, M.; Jastrzebski, Z.; Rosemann, T.; Nikolaidis, P.T.; et al. The effect of vitamin D3 supplementation on hepcidin, iron, and IL-6 responses after a 100 km ultra-marathon. Int. J. Environ. Res. Public Health 2020, 17, 2962. [Google Scholar] [CrossRef]
- Pilch, W.; Kita, B.; Piotrowska, A.; Tota, Ł.; Maciejczyk, M.; Czerwińska-Ledwig, O.; Sadowska- Krepa, E.; Kita, S.; Pałka, T. The effect of vitamin D supplementation on the muscle damage after eccentric exercise in young men: A randomized, control trial. J. Int. Soc. Sport. Nutr. 2020, 17, 53. [Google Scholar] [CrossRef]
- Shanely, R.A.; Nieman, D.C.; Knab, A.M.; Gillitt, N.D.; Meaney, M.P.; Jin, F.; Sha, W.; Cialdella-Kam, L. Influence of vitamin D mushroom powder supplementation on exercise-induced muscle damage in vitamin D insufficient high school athletes. J. Sport. Sci. 2014, 32, 670–679. [Google Scholar] [CrossRef]
- Jastrzebska, M.; Kaczmarczyk, M.; Michalczyk, M.; Radziminski, L.; Stepien, P.; Jastrzebska, J.; Wakuluk, D.; Suarez, A.D.; Lopez Sanchez, G.F.; Cieszczyk, P.; et al. Can supplementation of vitamin D improve aerobic capacity in well trained youth soccer players? J. Hum. Kinet. 2018, 61, 63–72. [Google Scholar] [CrossRef]
- Kendall, K.L.; Smith, A.E.; Graef, J.L.; Walter, A.A.; Moon, J.R.; Lockwood, C.M.; Beck, T.W.; Cramer, J.T.; Stout, J.R. Validity of electromyographic fatigue threshold as a noninvasive method for tracking changes in ventilatory threshold in college-aged men. J. Strength Cond. Res. 2010, 24, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Borg, G. Perceived exertion as an indicator of somatic stress. Scand. J. Rehabil. Med. Suppl. 1970, 2, 92–98. [Google Scholar]
- Ward, A.; Ebbeling, C.; Ahlquist, L. Indirect methods for estimation of aerobic power. In Physiological Assessment of Human Fitness; Maud, P.J., Foster, C., Eds.; Human Kinetics: Champaign, IL, USA, 1995; pp. 37–56. [Google Scholar]
- Lim, S.; Lee, K.; Woo, H.Y.; Park, H.; Kwon, M.J. Evaluation of Cobas 8000 analyzer series module e801 analytical performance. Ann. Clin. Lab. Sci. 2019, 49, 372–379. [Google Scholar] [PubMed]
- Cohen, J. A power primer. Psychol. Bull 1992, 112, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Meekins, M.E.; Oberhelman, S.S.; Lee, B.R.; Gardner, B.M.; Cha, S.S.; Singh, R.J.; Pettifor, J.M.; Fischer, P.R.; Thacher, T. Pharmacokinetics of daily versus monthly vitamin D3 supplementation in non-lactating women. Eur. J. Clin. Nutr. 2014, 68, 632–634. [Google Scholar] [CrossRef]
- Oberhelman, S.S.; Meekins, M.E.; Fischer, P.R.; Lee, B.R.; Singh, R.J.; Cha, S.S.; Gardner, B.M.; Pettifor, J.M.; Croghan, I.T.; Thacher, T.D. Maternal vitamin D supplementation to improve the vitamin D status of breast-fed infants: A randomized controlled trial. Mayo Clin. Proc. 2013, 88, 1378–1387. [Google Scholar] [CrossRef]
- Rhind, S.G.; Gannon, G.A.; Suzui, M.; Shephard, R.J.; Shek, P.N. Indomethacin inhibits circulating PGE2 and reverses postexercise suppression of natural killer cell activity. Am. J. Physiol. 1999, 276, R1496–R1505. [Google Scholar] [CrossRef]
- Turner, J.E.; Bosch, J.A.; Aldred, S. Measurement of exercise-induced oxidative stress in lymphocytes. Biochem. Soc. Trans. 2011, 39, 1299–1304. [Google Scholar] [CrossRef]
- Wang, J.-S.; Huang, Y.-H. Effects of exercise intensity on lymphocyte apoptosis induced by oxidative stress in men. Eur. J. Appl. Physiol. 2005, 95, 290–297. [Google Scholar] [CrossRef]
- Hewison, M.; Freeman, L.; Hughes, S.V.; Evans, K.N.; Bland, R.; Eliopoulos, A.G.; Kilby, M.D.; Moss, P.A.H.; Chakraverty, R. Differential Regulation of Vitamin D Receptor and Its Ligand in Human Monocyte-Derived Dendritic Cells1. J. Immunol. 2003, 170, 5382–5390. [Google Scholar] [CrossRef]
- Enioutina, E.Y.; Bareyan, D.; Daynes, R.A. TLR-induced local metabolism of vitamin D3 plays an important role in the diversification of adaptive immune responses. J. Immunol. 2009, 182, 4296–4305. [Google Scholar] [CrossRef]
- Di Rosa, M.; Malaguarnera, M.; Nicoletti, F.; Malaguarnera, L. Vitamin D3: A helpful immuno-modulator. Immunology 2011, 134, 123–139. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.-C.; Huang, S.-C. Active vitamin D3 attenuates the severity of Salmonella colitis in mice by orchestrating innate immunity. Immun. Inflamm. Dis. 2021, 9, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Colotta, F.; Jansson, B.; Bonelli, F. Modulation of inflammatory and immune responses by vitamin D. J. Autoimmun. 2017, 85, 78–97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Leung, D.Y.M.; Richers, B.N.; Liu, Y.; Remigio, L.K.; Riches, D.W.; Goleva, E. Vitamin D Inhibits Monocyte/Macrophage Proinflammatory Cytokine Production by Targeting MAPK Phosphatase-1. J. Immunol. 2012, 188, 2127–2135. [Google Scholar] [CrossRef]
- Kim, T.-H.; Lee, B.; Kwon, E.; Choi, S.J.; Lee, Y.H.; Song, G.G.; Sohn, J.; Ji, J.D. Regulation of TREM-1 expression by 1,25-dihydroxyvitamin D3 in human monocytes/macrophages. Immunol. Lett. 2013, 154, 80–85. [Google Scholar] [CrossRef]
- Bouchon, A.; Dietrich, J.; Colonna, M. Cutting Edge: Inflammatory Responses Can Be Triggered by TREM-1, a Novel Receptor Expressed on Neutrophils and Monocytes1. J. Immunol. 2000, 164, 4991–4995. [Google Scholar] [CrossRef]
- Zughaier, S.M.; Lubberts, E.; Bener, A. Editorial: Immune-Modulatory Effects of Vitamin D. Front. Immunol. 2020, 11, 596611. [Google Scholar] [CrossRef]
- White, J.H. Vitamin D metabolism and signaling in the immune system. Rev. Endocr. Metab. Disord. 2012, 13, 21–29. [Google Scholar] [CrossRef]
- Gombart, A.F.; Borregaard, N.; Koeffler, H.P. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is strongly up-regulated in myeloid cells by 1,25-dihydroxyvitamin D3. FASEB J. 2005, 19, 1067–1077. [Google Scholar] [CrossRef]
- Wang, T.-T.; Nestel, F.P.; Bourdeau, V.r.; Nagai, Y.; Wang, Q.; Liao, J.; Tavera-Mendoza, L.; Lin, R.; Hanrahan, J.W.; Mader, S.; et al. Cutting Edge: 1,25-Dihydroxyvitamin D3 Is a Direct Inducer of Antimicrobial Peptide Gene Expression1. J. Immunol. 2004, 173, 2909–2912. [Google Scholar] [CrossRef]
- Edfeldt, K.; Liu, P.T.; Chun, R.; Fabri, M.; Schenk, M.; Wheelwright, M.; Keegan, C.; Krutzik, S.R.; Adams, J.S.; Hewison, M.; et al. T-cell cytokines differentially control human monocyte antimicrobial responses by regulating vitamin D metabolism. Proc. Natl. Acad. Sci. USA 2010, 107, 22593–22598. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Lal, G. The molecular mechanism of natural killer cells function and its importance in cancer immunotherapy. Front. Immunol. 2017, 8, 1124. [Google Scholar] [CrossRef] [PubMed]
- Shaw, D.M.; Merien, F.; Braakhuis, A.; Dulson, D. T-cells and their cytokine production: The anti-inflammatory and immunosuppressive effects of strenuous exercise. Cytokine 2018, 104, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.P.; Turner, J.E. Debunking the myth of exercise-induced immune suppression: Redefining the impact of exercise on immunological health across the lifespan. Front. Immunol. 2018, 9, 648. [Google Scholar] [CrossRef] [PubMed]
- Shephard, R.J.; Rhind, S.; Shek, P.N. Exercise and training: Influences on cytotoxicity, interleukin-1, interleukin-2 and receptor structures. Int. J. Sport. Med. 1994, 15, S154–S166. [Google Scholar] [CrossRef] [PubMed]
- Wikby, A.; Maxson, P.; Olsson, J.; Johansson, B.; Ferguson, F.G. Changes in CD8 and CD4 lymphocyte subsets, T cell proliferation responses and non-survival in the very old: The Swedish longitudinal OCTO-immune study. Mech. Ageing Dev. 1998, 102, 187–198. [Google Scholar] [CrossRef]
- Villaggio, B.; Soldano, S.; Cutolo, M. 1, 25-dihydroxyvitamin D3 downregulates aromatase expression and inflammatory cytokines in human macrophages. Clin. Exp. Rheumatol. 2012, 30, 934–938. [Google Scholar]
- Guo, J.; Ma, Z.; Ma, Q.; Wu, Z.; Fan, P.; Zhou, X.; Chen, L.; Zhou, S.; Goltzman, D.; Miao, D. 1, 25(OH) 2D3 inhibits hepatocellular carcinoma development through reducing secretion of inflammatory cytokines from immunocytes. Curr. Med. Chem. 2013, 20, 4131–4141. [Google Scholar] [CrossRef]
- Gysemans, C.A.; Cardozo, A.K.; Callewaert, H.; Giulietti, A.; Hulshagen, L.; Bouillon, R.; Eizirik, D.c.L.; Mathieu, C. 1,25-Dihydroxyvitamin D3 Modulates Expression of Chemokines and Cytokines in Pancreatic Islets: Implications for Prevention of Diabetes in Nonobese Diabetic Mice. Endocrinology 2005, 146, 1956–1964. [Google Scholar] [CrossRef]
- Choi, M.; Park, H.; Cho, S.; Lee, M. Vitamin D3 supplementation modulates inflammatory responses from the muscle damage induced by high-intensity exercise in SD rats. Cytokine 2013, 63, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, A.E.; Flynn, M.G.; Pinkston, C.; Markofski, M.M.; Jiang, Y.; Donkin, S.S.; Teegarden, D. Vitamin D supplementation during exercise training does not alter inflammatory biomarkers in overweight and obese subjects. Eur. J. Appl. Physiol. 2012, 112, 3045–3052. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.M.; Pedersen, B.K. The anti-inflammatory effect of exercise. J. Appl. Physiol. 2005, 98, 1154–1162. [Google Scholar] [CrossRef]
- Dumas, S.N.; Guo, C.A.; Kim, J.K.; Friedline, R.H.; Ntambi, J.M. Interleukin-6 derived from cutaneous deficiency of stearoyl-CoA desaturase- 1 may mediate metabolic organ crosstalk among skin, adipose tissue and liver. Biochem. Biophys. Res. Commun. 2019, 508, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Roca, E.; Nescolarde, L.; Lupón, J.; Barallat, J.; Januzzi, J.L.; Liu, P.; Cruz Pastor, M.; Bayes-Genis, A. The Dynamics of Cardiovascular Biomarkers in non-Elite Marathon Runners. J. Cardiovasc. Transl. Res. 2017, 10, 206–208. [Google Scholar] [CrossRef]
- Tokinoya, K.; Ishikura, K.; Ra, S.-G.; Ebina, K.; Miyakawa, S.; Ohmori, H. Relationship between early-onset muscle soreness and indirect muscle damage markers and their dynamics after a full marathon. J. Exerc. Sci. Fit. 2020, 18, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Bernat-Adell, M.D.; Collado-Boira, E.J.; Moles-Julio, P.; Panizo-González, N.; Martínez-Navarro, I.; Hernando-Fuster, B.; Hernando-Domingo, C. Recovery of Inflammation, Cardiac, and Muscle Damage Biomarkers After Running a Marathon. J. Strength Cond. Res. 2021, 35, 626–632. [Google Scholar] [CrossRef]
- Heavens, K.R.; Szivak, T.K.; Hooper, D.R.; Dunn-Lewis, C.; Comstock, B.A.; Flanagan, S.D.; Looney, D.P.; Kupchak, B.R.; Maresh, C.M.; Volek, J.S.; et al. The effects of high intensity short rest resistance exercise on muscle damage markers in men and women. J. Strength Cond. Res. 2014, 28, 1041–1049. [Google Scholar] [CrossRef]
- Ogan, D.; Pritchett, K. Vitamin D and the athlete: Risks, recommendations, and benefits. Nutrients 2013, 5, 1856–1868. [Google Scholar] [CrossRef]
- Ainbinder, A.; Boncompagni, S.; Protasi, F.; Dirksen, R.T. Role of Mitofusin-2 in mitochondrial localization and calcium uptake in skeletal muscle. Cell Calcium 2015, 57, 14–24. [Google Scholar] [CrossRef]
- Von Hurst, P.R.; Beck, K.L. Vitamin D and skeletal muscle function in athletes. Curr Opin Clin Nutr Metab Care 2014, 17, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Buitrago, C.; Pardo, V.G.; Boland, R. Role of VDR in 1α,25-dihydroxyvitamin D3-dependent non-genomic activation of MAPKs, Src and Akt in skeletal muscle cells. J. Steroid. Biochem. Mol. Biol. 2013, 136, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Mendes, M.M.; Botelho, P.B.; Ribeiro, H. Vitamin D and musculoskeletal health: Outstanding aspects to be considered in the light of current evidence. Endocr. Connect 2022, 11, e210596. [Google Scholar] [CrossRef] [PubMed]
- Owens, D.J.; Fraser, W.D.; Close, G.L. Vitamin D and the athlete: Emerging insights. Eur. J. Sport Sci. 2015, 15, 73–84. [Google Scholar] [CrossRef]
- Bello, H.J.; Caballero-García, A.; Pérez-Valdecantos, D.; Roche, E.; Noriega, D.C.; Córdova-Martínez, A. Effects of Vitamin D in Post-Exercise Muscle Recovery. A Systematic Review and Meta-Analysis. Nutrients 2021, 13, 4013. [Google Scholar] [CrossRef]
- Johnson, Q.R.; Goatcher, J.D.; Diehl, C.; Lockie, R.G.; Orr, R.M.; Alvar, B.; Smith, D.B.; Dawes, J.J. Heart rate responses during simulated fire ground scenarios among full-time firefighters. Int. J. Exerc. Sci. 2020, 13, 374–382. [Google Scholar]
- Dimkpa, U.; Chidi, E.; Unaeze, B.; Besong, E.; Umahi, O.; Enemuo, C.; Okafor, E.; Okeke, M. A comparative study of cardio-metabolic responses to exercise between untrained non-athletic young Nigerian adults and trained soccer players. Comp. Exerc. Physiol. 2020, 16, 217–224. [Google Scholar] [CrossRef]
- Whyte, G.P.; George, K.; Shave, R.; Middleton, N.; Nevill, A.M. Training induced changes in maximum heart rate. Int. J. Sport. Med. 2008, 29, 129–133. [Google Scholar] [CrossRef]
- Tishkoff, D.X.; Nibbelink, K.A.; Holmberg, K.H.; Dandu, L.; Simpson, R.U. Functional Vitamin D Receptor (VDR) in the T-Tubules of Cardiac Myocytes: VDR Knockout Cardiomyocyte Contractility. Endocrinology 2008, 149, 558–564. [Google Scholar] [CrossRef]
- Reihmane, D.; Dela, F. Interleukin-6: Possible biological roles during exercise. Eur. J. Sport Sci. 2014, 14, 242–250. [Google Scholar] [CrossRef]
- Skalska, M.; Nikolaidis, P.T.; Knechtle, B.; Rosemann, T.J.; Radzimiński, Ł.; Jastrzębska, J.; Kaczmarczyk, M.; Myśliwiec, A.; Dragos, P.; López-Sánchez, G.F. Vitamin D supplementation and physical activity of young soccer players during high-intensity training. Nutrients 2019, 11, 349. [Google Scholar] [CrossRef] [PubMed]
- Fairbairn, K.A.; Ceelen, I.J.M.; Skeaff, C.M.; Cameron, C.M.; Perry, T.L. Vitamin D3 supplementation does not improve sprint performance in professional rugby players: A randomized, placebo-controlled, double-blind intervention study. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kujach, S.; Lyzwinski, D.; Chroboczek, M.; Bialowas, D.; Antosiewicz, J.; Laskowski, R. The effect of vitamin D3 supplementation on physical capacity among active college-aged males. Nutrients 2020, 12, 1936. [Google Scholar] [CrossRef] [PubMed]
- Mielgo-Ayuso, J.; Calleja-González, J.; Urdampilleta, A.; León-Guereño, P.; Córdova, A.; Caballero-García, A.; Fernandez-Lázaro, D. Effects of vitamin D supplementation on haematological values and muscle recovery in elite male traditional rowers. Nutrients 2018, 10, 1968. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Vitamin D3 Group | Placebo Group | p-Value |
|---|---|---|---|
| Age (years) | 21.9 ± 1.4 | 22.1 ± 2.0 | 0.784 |
| Body height (cm) | 173.1 ± 6.3 | 173.3 ± 6.8 | 0.944 |
| Body weight (kg) | 64.2 ± 9.1 | 69.1 ± 8.2 | 0.248 |
| Groups | Exhaustion (min) | HRmax (beats/min) | AHR (beats/min) | VO2max-1 (mL/kg/min) | VO2max-2 (mL/kg/min) |
|---|---|---|---|---|---|
| Vitamin D3 | 121.48 ± 1.59 | 176.00 ± 9.06 # | 156.89 ± 6.37 # | 51.11 ± 5.93 | 47.89 ± 8.81 |
| Placebo | 121.15 ± 0.23 | 186.22 ± 7.73 | 167.33 ± 10.31 | 47.56 ± 5.29 | 44.78 ± 6.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.-C.; Weng, P.-W.; Chen, S.-C.; Liu, T.-H.; Huang, H.-W.; Huang, C.-T.; Yang, C.-T.; Mishra, V.K.; Yang, M.-T. Immunologic, Anti-Inflammatory, and Anti-Muscle Damage Profile of Supplemented Vitamin D3 in Healthy Adults on Strenuous Endurance Exercise. Biology 2023, 12, 657. https://doi.org/10.3390/biology12050657
Liu M-C, Weng P-W, Chen S-C, Liu T-H, Huang H-W, Huang C-T, Yang C-T, Mishra VK, Yang M-T. Immunologic, Anti-Inflammatory, and Anti-Muscle Damage Profile of Supplemented Vitamin D3 in Healthy Adults on Strenuous Endurance Exercise. Biology. 2023; 12(5):657. https://doi.org/10.3390/biology12050657
Chicago/Turabian StyleLiu, Ming-Che, Pei-Wei Weng, Sheng-Chang Chen, Ting-Hao Liu, Hsiang-Wei Huang, Chang-Ti Huang, Cheng-Tse Yang, Viraj Krishna Mishra, and Ming-Ta Yang. 2023. "Immunologic, Anti-Inflammatory, and Anti-Muscle Damage Profile of Supplemented Vitamin D3 in Healthy Adults on Strenuous Endurance Exercise" Biology 12, no. 5: 657. https://doi.org/10.3390/biology12050657