
Adaptation of Commensal Escherichia coli in Tomato Fruits: Motility, Stress, Virulence
 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Plant Material
2.2. Tomato Fruits Contamination
2.3. Differential Methylation Analysis
2.3.1. Tomato Contamination and E. coli TOM Isolation
2.3.2. DNA Extraction and Sequencing
2.3.3. qPCR Analysis
2.4. Growth Curves in the Presence of Antibiotics and H2O2
2.5. Motility Test
2.6. Experiments with Human Monocyte-Derived Dendritic Cells
2.6.1. Monocyte-Derived Dendritic Cells Differentiation
2.6.2. Viability and Activation of E. coli-Infected Monocyte-Derived Dendritic Cells
3. Results
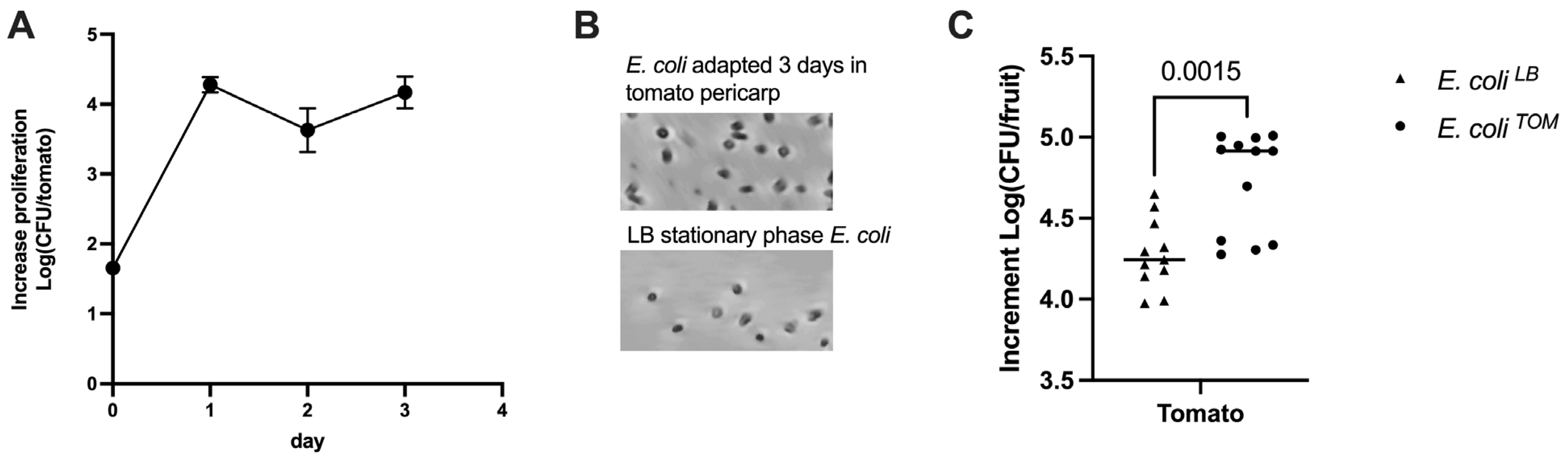
3.1. Pre-Adaptation of E. coli to Tomato Pericarp
3.2. Differential Methylation Analysis and Expression of Genes Involved in Bacterial Adaptation
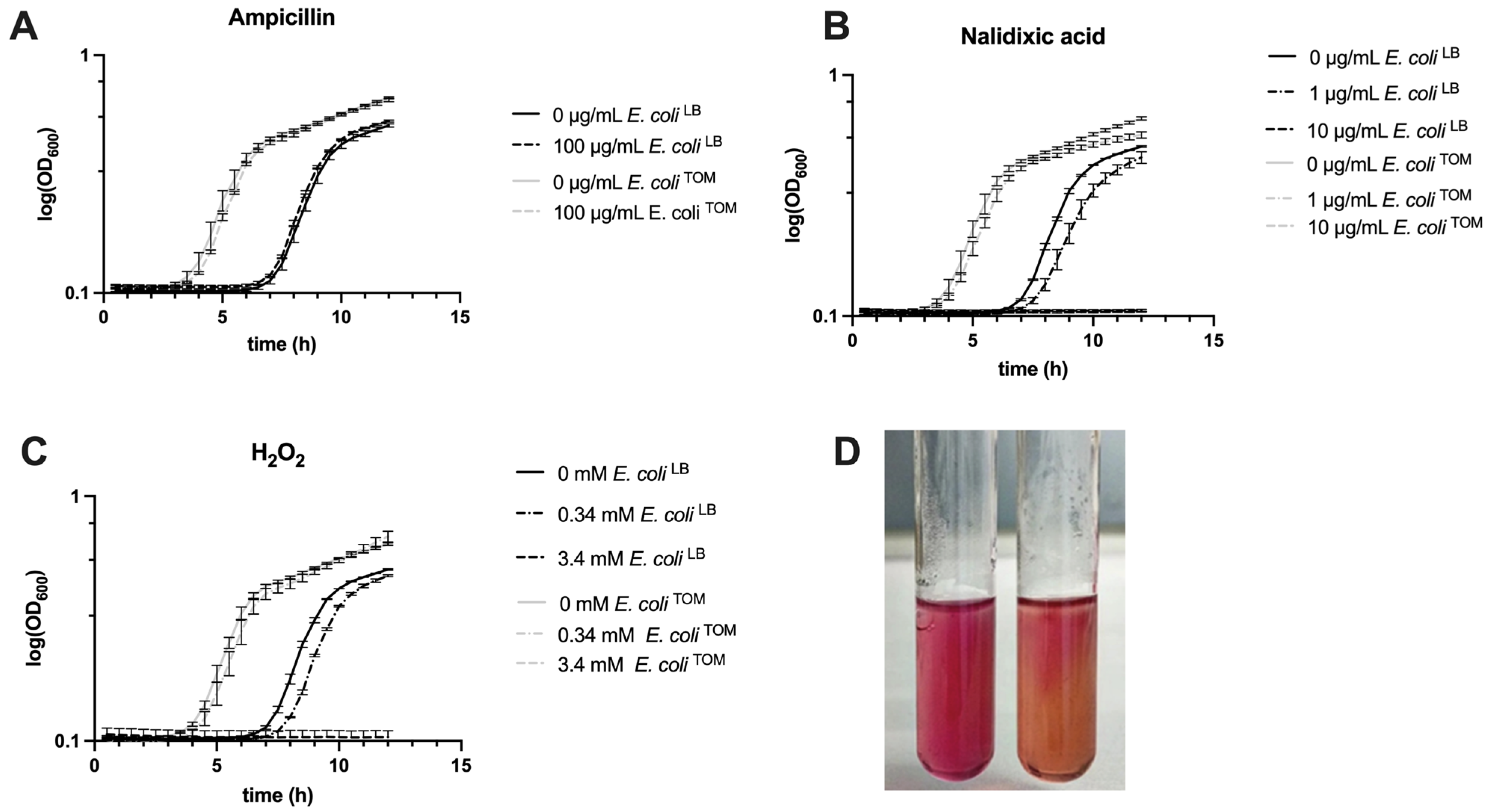
3.3. Effects of Pre-Adaptation on Environmental Stress Resistance and Cell Motility
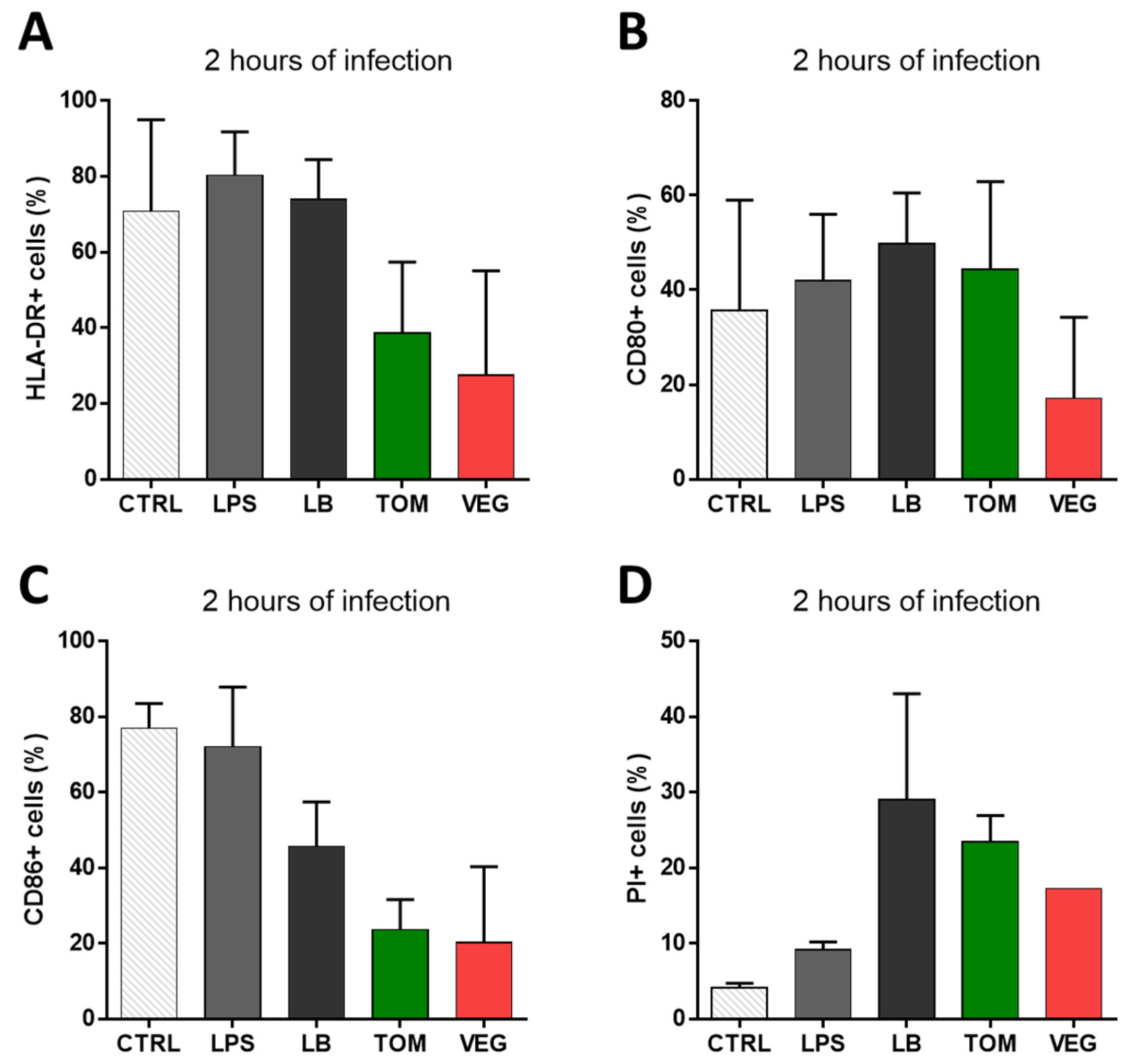
3.4. Effects of Pre-Adaptation on Pathogenicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holden, N.; Pritchard, L.; Toth, I. Colonization outwith the colon: Plants as an alternative environmental reservoir for human pathogenic enterobacteria: Review article. FEMS Microbiol. Rev. 2009, 33, 689–703. [Google Scholar] [CrossRef] [PubMed]
- Conway, T.; Krogfelt, K.A.; Cohen, P.S. The Life of Commensal Escherichia coli in the Mammalian Intestine. EcoSal Plus 2004, 1. [Google Scholar] [CrossRef] [PubMed]
- Bartz, J.A.; Marvasi, M.; Teplitski, M. Salmonella and Tomatoes; Academic Press: Cambridge, MA, USA, 2014; ISBN 9780124046115. [Google Scholar]
- Luna-Guevara, J.J.; Arenas-Hernandez, M.M.P.; Martínez De La Peña, C.; Silva, J.L.; Luna-Guevara, M.L. The Role of Pathogenic E. coli in Fresh Vegetables: Behavior, Contamination Factors, and Preventive Measures. Int. J. Microbiol. 2019, 2019, 2894328. [Google Scholar] [CrossRef] [PubMed]
- Marvasi, M.; Noel, J.T.; George, A.S.; Farias, M.A.; Jenkins, K.T.; Hochmuth, G.; Xu, Y.; Giovanonni, J.J.; Teplitski, M. Ethylene signalling affects susceptibility of tomatoes to Salmonella. Microb. Biotechnol. 2014, 7, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Devleesschauwer, B.; Marvasi, M.; Giurcanu, M.C.; Hochmuth, G.J.; Speybroeck, N.; Havelaar, A.H.; Teplitski, M. High relative humidity pre-harvest reduces post-harvest proliferation of Salmonella in tomatoes. Food Microbiol. 2017, 66, 55–63. [Google Scholar] [CrossRef]
- Kroupitski, Y.; Golberg, D.; Belausov, E.; Pinto, R.; Swartzberg, D.; Granot, D.; Sela, S. Internalization of Salmonella enterica in Leaves Is Induced by Light and Involves Chemotaxis and Penetration through Open Stomata. Appl. Environ. Microbiol. 2009, 75, 6076–6086. [Google Scholar] [CrossRef]
- Lu, S.; Song, J.; Campbell-Palmer, L. A modified chemiluminescence method for hydrogen peroxide determination in apple fruit tissues. Sci. Hortic. 2009, 120, 336–341. [Google Scholar] [CrossRef]
- Nasirizadeh, N.; Shekari, Z.; Nazari, A.; Tabatabaee, M. Fabrication of a novel electrochemical sensor for determination of hydrogen peroxide in different fruit juice samples. J. Food Drug Anal. 2016, 24, 72–82. [Google Scholar] [CrossRef]
- Hyong, W.C.; Young, J.K.; Sung, C.L.; Jeum, K.H.; Byung, K.H. Hydrogen peroxide generation by the pepper extracellular peroxidase CaPO2 activates local and systemic cell death and defense response to bacterial pathogens. Plant Physiol. 2007, 145, 890–904. [Google Scholar] [CrossRef]
- Dublan, M.D.L.A.; Ortiz-Marquez, J.C.F.; Lett, L.; Curatti, L. Plant-adapted Escherichia coli show increased lettuce colonizing ability, resistance to oxidative stress and chemotactic response. PLoS ONE 2014, 9, e110416. [Google Scholar] [CrossRef]
- Landstorfer, R.; Simon, S.; Schober, S.; Keim, D.; Scherer, S.; Neuhaus, K. Comparison of strand-specific transcriptomes of enterohemorrhagic Escherichia coli O157: H7 EDL933 (EHEC) under eleven different environmental conditions including radish sprouts and cattle feces. BMC Genom. 2014, 15, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Jacob, C.; Velásquez, A.C.; Josh, N.A.; Settles, M.; He, S.Y.; Melotto, M. Dual transcriptomic analysis reveals metabolic changes associated with differential persistence of human pathogenic bacteria in leaves of Arabidopsis and lettuce. G3 Genes Genomes Genet. 2021, 11, jkab331. [Google Scholar] [CrossRef] [PubMed]
- Beaulaurier, J.; Schadt, E.E.; Fang, G. Deciphering bacterial epigenomes using modern sequencing technologies. Nat. Rev. Genet. 2018, 20, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Romero, M.A.; Casadesús, J. The bacterial epigenome. Nat. Rev. Microbiol. 2019, 18, 7–20. [Google Scholar] [CrossRef]
- Rand, A.C.; Jain, M.; Eizenga, J.M.; Musselman-Brown, A.; Olsen, H.E.; Akeson, M.; Paten, B. Mapping DNA methylation with high-throughput nanopore sequencing. Nat. Methods 2017, 14, 411–413. [Google Scholar] [CrossRef]
- Simpson, J.T.; Workman, R.E.; Zuzarte, P.C.; David, M.; Dursi, L.J.; Timp, W. Detecting DNA cytosine methylation using nanopore sequencing. Nat. Methods 2017, 14, 407–410. [Google Scholar] [CrossRef]
- Kittana, H.; Gomes-Neto, J.C.; Heck, K.; Geis, A.L.; Segura Muñoz, R.R.; Cody, L.A.; Schmaltz, R.J.; Bindels, L.B.; Sinha, R.; Hostetter, J.M.; et al. Commensal Escherichia coli strains can promote intestinal inflammation via differential interleukin-6 production. Front. Immunol. 2018, 9, 2318. [Google Scholar] [CrossRef]
- Kelly, D.; Campbell, J.I.; King, T.P.; Grant, G.; Jansson, E.A.; Coutts, A.G.P.; Pettersson, S.; Conway, S. Commensal anaerobic gut bacteria attenuate inflammation by regulating nuclear-cytoplasmic shutting of PPAR-$γ$ and ReIA. Nat. Immunol. 2004, 5, 104–112. [Google Scholar] [CrossRef]
- Chapman, T.A.; Wu, X.-Y.; Barchia, I.; Bettelheim, K.A.; Driesen, S.; Trott, D.; Wilson, M.; Chin, J.J.C. Comparison of virulence gene profiles of Escherichia coli strains isolated from healthy and diarrheic swine. Appl. Environ. Microbiol. 2006, 72, 4782–4795. [Google Scholar] [CrossRef]
- Semenzato, G.; Alonso-Vásquez, T.; Duca, S.D.; Vassallo, A.; Riccardi, C.; Zaccaroni, M.; Mucci, N.; Padula, A.; Emiliani, G.; Piccionello, A.P.; et al. Genomic Analysis of Endophytic Bacillus-Related Strains Isolated from the Medicinal Plant Origanum vulgare L. Revealed the Presence of Metabolic Pathways Involved in the Biosynthesis of Bioactive Compounds. Microorganisms 2022, 10, 919. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-Z.; Chang, M.-X.; Yang, L.; Liu, Y.-Y.; Chen, P.-X.; Jiang, H.-X. Upregulation of AcrEF in quinolone resistance development in Escherichia coli when AcrAB-TolC function is impaired. Microb. Drug Resist. 2018, 24, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Lehti, T.A.; Bauchart, P.; Dobrindt, U.; Korhonen, T.K.; Westerlund-Wikström, B. The fimbriae activator MatA switches off motility in Escherichia coli by repression of the flagellar master operon flhDC. Microbiology 2012, 158, 1444–1455. [Google Scholar] [CrossRef]
- Riber, L.; Hansen, L.H. Epigenetic Memories: The Hidden Drivers of Bacterial Persistence? Trends Microbiol. 2021, 29, 190–194. [Google Scholar] [CrossRef]
- Orozco-Cárdenas, M.L.; Narváez-Vásquez, J.; Ryan, C.A. Hydrogen Peroxide Acts as a Second Messenger for the Induction of Defense Genes in Tomato Plants in Response to Wounding, Systemin, and Methyl Jasmonate. Plant Cell 2001, 13, 179–191. [Google Scholar] [CrossRef]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive Oxygen Species in Plant Signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef]
- Scott, R.A.; Thilmony, R.; Harden, L.A.; Zhou, Y.; Brandl, M.T. Escherichia coli O157:H7 Converts Plant-Derived Choline to Glycine Betaine for Osmoprotection during Pre- and Post-harvest Colonization of Injured Lettuce Leaves. Front. Microbiol. 2017, 8, 2436. [Google Scholar] [CrossRef]
- de Moraes, M.H.; Desai, P.; Porwollik, S.; Canals, R.; Perez, D.R.; Chu, W.; McClelland, M.; Teplitski, M. Salmonella Persistence in Tomatoes Requires a Distinct Set of Metabolic Functions Identified by Transposon Insertion Sequencing. Appl. Environ. Microbiol. 2017, 83, e03028-16. [Google Scholar] [CrossRef]
- Prusky, D.B.; Wilson, R.A. Does increased nutritional carbon availability in fruit and foliar hosts contribute to modulation of pathogen colonization? Postharvest Biol. Technol. 2018, 145, 27–32. [Google Scholar] [CrossRef]
- Ferelli, A.M.C.; Bolten, S.; Szczesny, B.; Micallef, S.A. Salmonella enterica elicits and is restricted by nitric oxide and reactive oxygen species on tomato. Front. Microbiol. 2020, 1, 391. [Google Scholar] [CrossRef] [PubMed]
- Marvasi, M.; George, A.S.; Giurcanu, M.; Hochmuth, G.J.; Noel, J.T.; Gause, E.; Teplitski, M. Effects of nitrogen and potassium fertilization on the susceptibility of tomatoes to post-harvest proliferation of Salmonella enterica. Food Microbiol. 2014, 43, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Marvasi, M.; George, A.S.; Giurcanu, M.C.; Hochmuth, G.J.; Noel, J.T.; Teplitski, M. Effect of the irrigation regime on the susceptibility of pepper and tomato to post-harvest proliferation of Salmonella enterica. Food Microbiol. 2015, 46, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Lenzi, A.; Marvasi, M.; Baldi, A. Agronomic practices to limit pre- and post-harvest contamination and proliferation of human pathogenic Enterobacteriaceae in vegetable produce. Food Control 2021, 119, 107486. [Google Scholar] [CrossRef]
- Marvasi, M. Potential use and perspectives of nitric oxide donors in agriculture. J. Sci. Food Agric. 2017, 97, 1065–1072. [Google Scholar] [CrossRef]
- Marvasi, M.; Durie, I.A.; McLamore, E.S.; Vanegas, D.C.; Chaturvedi, P. Salmonella enterica biofilm-mediated dispersal by nitric oxide donors in association with cellulose nanocrystal hydrogels. AMB Express 2015, 5, 1–9. [Google Scholar] [CrossRef]
- Henriquez, T.; Lenzi, A.; Baldi, A.; Marvasi, M. Frontiers in Plant Breeding: Perspectives for the Selection of Vegetables Less Susceptible to Enteric Pathogens. Front. Microbiol. 2020, 11, 1087. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Function | Methylation 1 in E. coli TOM | Relative Expression (E. coli TOM vs. E. coli LB) |
|---|---|---|---|
| narG | Respiratory nitrate reductase 1 alpha chain | + | −13.9 |
| papA_1 | Pap fimbrial major pilin protein | − | −11.5 |
| fimH | Type 1 fimbrin D-mannose specific adhesin | − | −36.1 |
| atoBCD | Metabolism of Acetyl-CoA | + | −2.52 |
| acrF | Multidrug export protein AcrF | − | +1.6 |
| murE-ftsI-ftsL | Peptidoglycan | + | −1.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vassallo, A.; Amoriello, R.; Guri, P.; Casbarra, L.; Ramazzotti, M.; Zaccaroni, M.; Ballerini, C.; Cavalieri, D.; Marvasi, M. Adaptation of Commensal Escherichia coli in Tomato Fruits: Motility, Stress, Virulence. Biology 2023, 12, 633. https://doi.org/10.3390/biology12040633
Vassallo A, Amoriello R, Guri P, Casbarra L, Ramazzotti M, Zaccaroni M, Ballerini C, Cavalieri D, Marvasi M. Adaptation of Commensal Escherichia coli in Tomato Fruits: Motility, Stress, Virulence. Biology. 2023; 12(4):633. https://doi.org/10.3390/biology12040633
Chicago/Turabian StyleVassallo, Alberto, Roberta Amoriello, Prandvera Guri, Lorenzo Casbarra, Matteo Ramazzotti, Marco Zaccaroni, Clara Ballerini, Duccio Cavalieri, and Massimiliano Marvasi. 2023. "Adaptation of Commensal Escherichia coli in Tomato Fruits: Motility, Stress, Virulence" Biology 12, no. 4: 633. https://doi.org/10.3390/biology12040633