
Anatomical, Histological, and Morphometrical Investigations of the Auditory Ossicles in Chlorocebus aethiops sabaeus from Saint Kitts Island

 ,
,  , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Material of Animal Origin
2.2. Anatomical Image Collection and Processing Technique
2.3. Histological Technique
2.4. Morphometrical Approach
3. Results
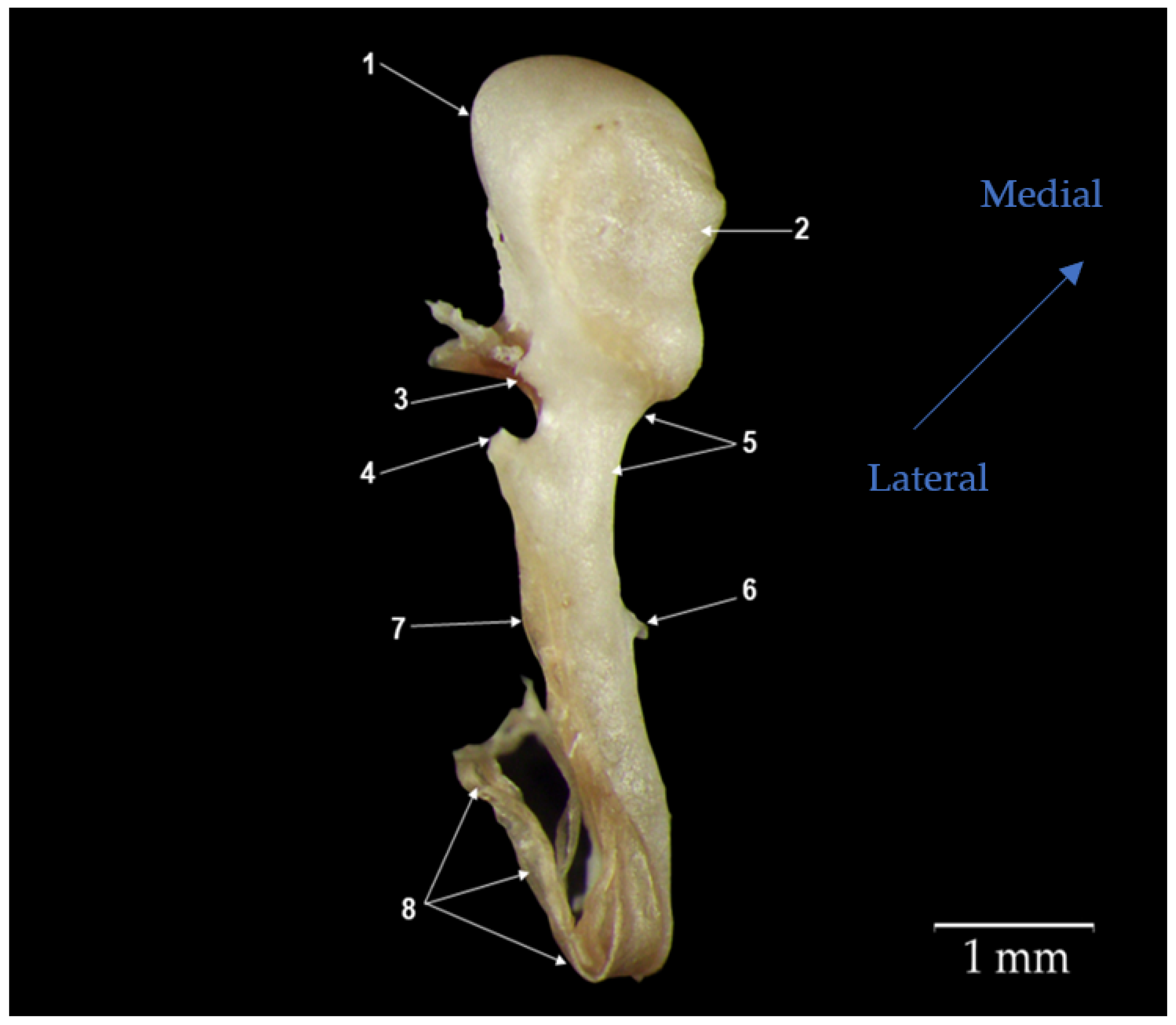
3.1. Malleus
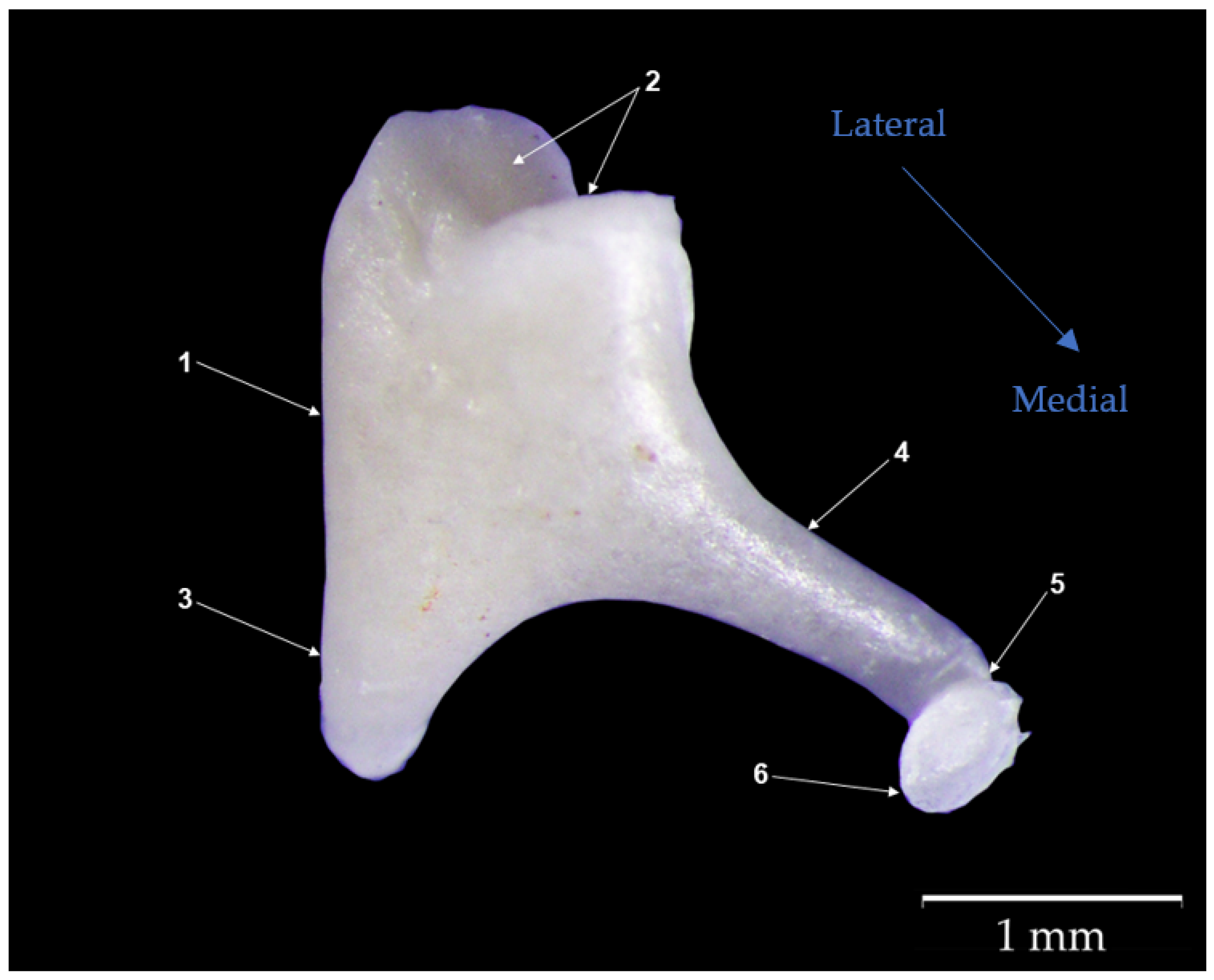
3.2. Incus
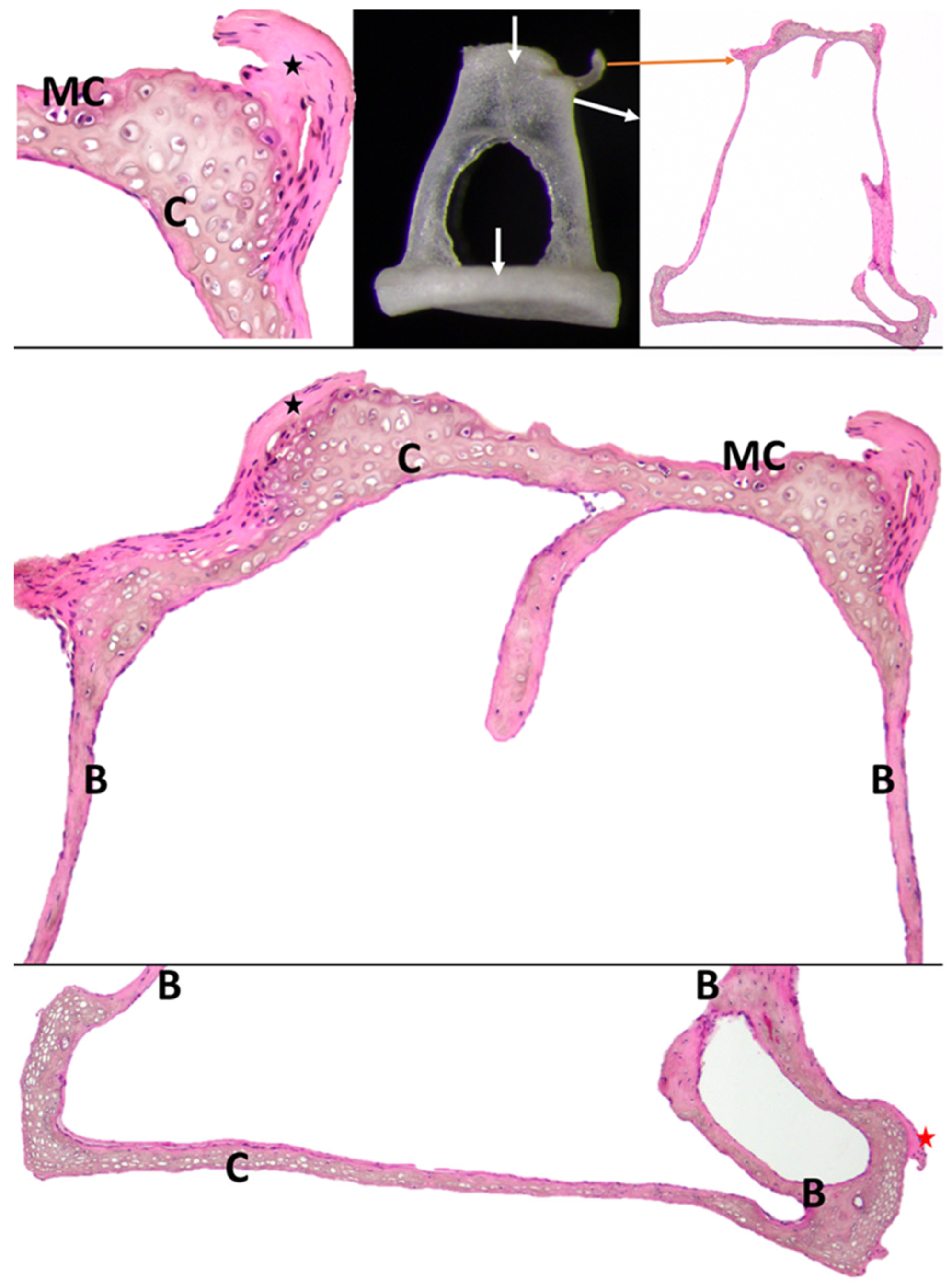
3.3. Stapes
4. Discussion
4.1. Gross Morphological Features
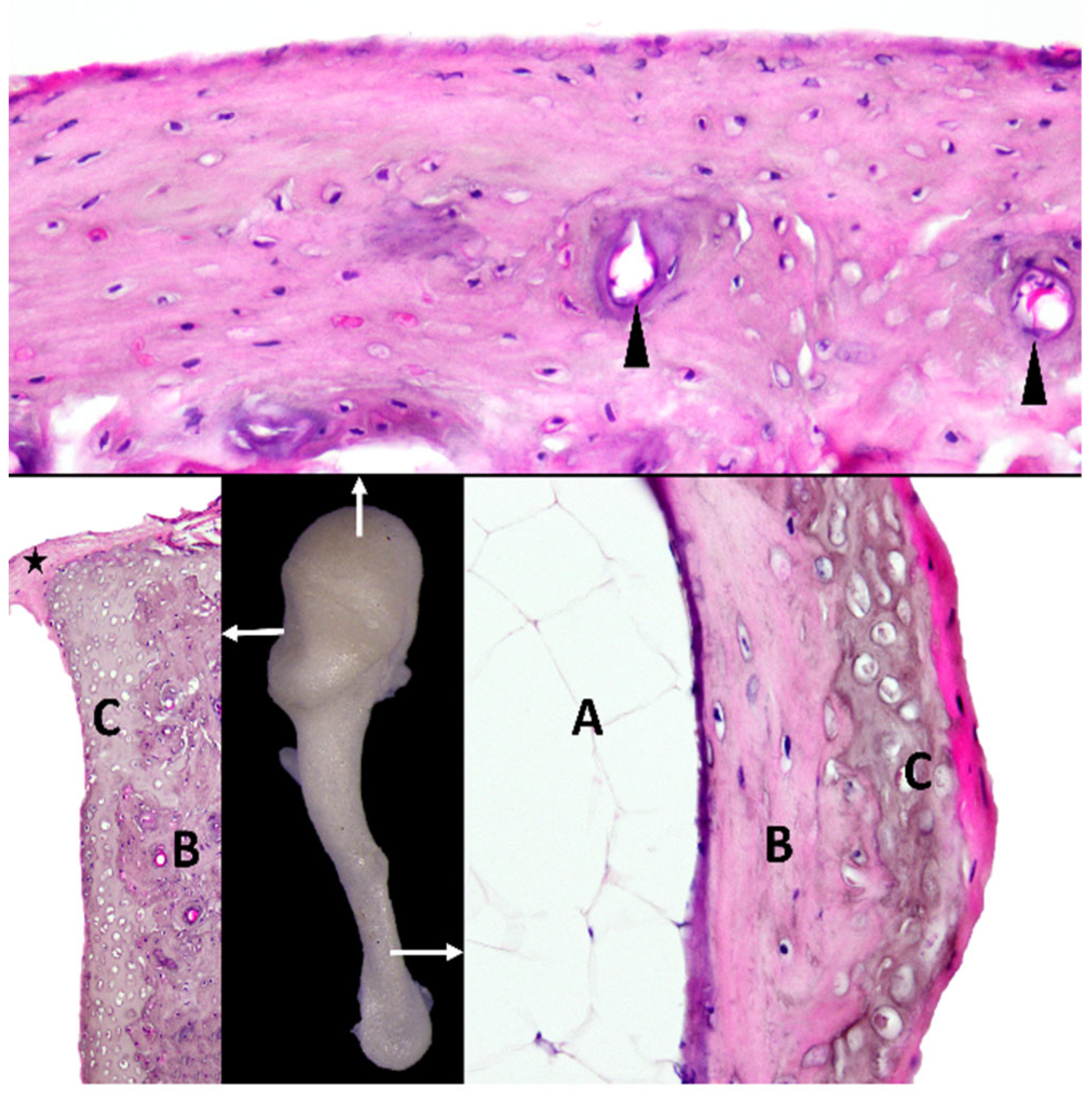
4.2. Micromorphological Features
4.3. Morphometrical Interpretation of Data
4.4. Middle Ear Lever Ratio
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Young, A.N.; du Plessis, W.M.; Rodriguez, D.; Beierschmitt, A. Thoracic Radiographic Anatomy in Vervet Monkeys (Chlorocebus Sabaeus). J. Med. Primatol. 2013, 42, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Ervin, F.; Palmour, R. Primates for 21st Century Biomedicine: The St. Kitts Vervet (Chlorocebus Aethiops, SK). In International Perspectives: The Future of Nonhuman Primate Resources; National Academies Press: Washington, DC, USA, 2003; pp. 49–53. [Google Scholar]
- Amory, J.T.; Du Plessis, W.M.; Beierschmitt, A.; Beeler-Marfisi, J.; Palmour, R.M.; Beths, T. Abdominal Ultrasonography of the Normal St. Kitts Vervet Monkey (Chlorocebus Sabaeus). J. Med. Primatol. 2013, 42, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.R.; Cramer, J.D.; Nisbett, A.; Gray, J.P. A Comparison of Adult Body Size between Captive and Wild Vervet Monkeys (Chlorocebus Aethiops Sabaeus) on the Island of St. Kitts. Primates 2016, 57, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Quam, R.M.; Coleman, M.N.; Martínez, I. Evolution of the Auditory Ossicles in Extant Hominids: Metric Variation in African Apes and Humans. J. Anat. 2014, 225, 167–196. [Google Scholar] [CrossRef]
- Saravanan, C.; Flandre, T.; Hodo, C.L.; Lewis, A.D.; Mecklenburg, L.; Romeike, A.; Turner, O.C.; Yen, H.Y. Research Relevant Conditions and Pathology in Nonhuman Primates. ILAR J. 2020, 61, 139–166. [Google Scholar] [CrossRef]
- Layton, R.C.; Mega, W.; McDonald, J.D.; Brasel, T.L.; Barr, E.B.; Gigliotti, A.P.; Koster, F. Levofloxacin Cures Experimental Pneumonic Plague in African Green Monkeys. PLoS Negl. Trop. Dis. 2011, 5, e959. [Google Scholar] [CrossRef]
- Whitehouse, C.A.; Keirstead, N.; Taylor, J.; Reinhardt, J.L.; Beierschmitt, A. Prevalence of Hypermucoid Klebsiella Pneumoniae among Wildcaught and Captive Vervet Monkeys (Chlorocebus Aethiops Sabaeus) on the Island of St. Kitts. J. Wildl. Dis. 2010, 46, 971–976. [Google Scholar] [CrossRef]
- Hoefer, A.; Boyen, F.; Beierschmitt, A.; Moodley, A.; Roberts, M.C.; Butaye, P. Methicillin-Resistant and Methicillin-Susceptible Staphylococcus from Vervet Monkeys (Chlorocebus Sabaeus) in Saint Kitts. Antibiotics 2021, 10, 290. [Google Scholar] [CrossRef]
- Lerche, N.W.; Yee, J.A.L.; Capuano, S.V.; Flynn, J.L. New Approaches to Tuberculosis Surveillance in Nonhuman Primates. ILAR J. 2008, 49, 170–178. [Google Scholar] [CrossRef]
- Woolsey, C.; Borisevich, V.; Prasad, A.N.; Agans, K.N.; Deer, D.J.; Dobias, N.S.; Heymann, J.C.; Foster, S.L.; Levine, C.B.; Medina, L.; et al. Establishment of an African Green Monkey Model for COVID-19 and Protection against Re-Infection. Nat. Immunol. 2020, 22, 86–98. [Google Scholar] [CrossRef]
- Bjornson-Hooper, Z.B.; Fragiadakis, G.K.; Spitzer, M.H.; Chen, H.; Madhireddy, D.; Hu, K.; Lundsten, K.; McIlwain, D.R.; Nolan, G.P. A Comprehensive Atlas of Immunological Differences between Humans, Mice, and Non-Human Primates. Front. Immunol. 2022, 13, 867015. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.M.; Doyle, W.J.; Gest, T.R. Morphology of the Middle Ear in the Rhesus Monkey (Macaca Mulatta). Acta Anat. 1982, 112, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Doyle, W.J.; Rood, S.R. Anatomy of the Auditory Tube and Related Structures in the Rhesus Monkey (Macaca Mulatta). Cell. Tissues Organs 1979, 105, 209–225. [Google Scholar] [CrossRef] [PubMed]
- Owren, M.J.; Hopp, S.L.; Sinnott, J.M.; Petersen, M.R. Absolute Auditory Thresholds in Three Old World Monkey Species (Cercopithecus Aethiops, C. Neglectus, Macaca Fuscata) and Humans (Homo Sapiens). J. Comp. Psychol. 1988, 102, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Masali, M. The Ear Ossicles and the Evolution of the Primate Ear: A Biomechanical Approach. Hum. Evol. 1992, 7, 1–5. [Google Scholar] [CrossRef]
- Coleman, M.N. The Functional Morphology and Evolution of the Primate Auditory System. Ph.D. Thesis, Stony Brook University, New York, NY, USA, 2007. [Google Scholar]
- Coleman, M.N.; Ross, C.F. Primate Auditory Diversity and Its Influence on Hearing Performance. Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2004, 281, 1123–1137. [Google Scholar] [CrossRef] [PubMed]
- Wysocki, J. Topographical Anatomy and Morphometry of the Temporal Bone of the Macaque. Folia Morphol. 2009, 68, 13–22. [Google Scholar]
- Price, T.; Ndiaye, O.; Hammerschmidt, K.; Fischer, J. Limited Geographic Variation in the Acoustic Structure of and Responses to Adult Male Alarm Barks of African Green Monkeys. Behav. Ecol. Sociobiol. 2014, 68, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Stoessel, A.; David, R.; Gunz, P.; Schmidt, T.; Spoor, F.; Hublin, J.J. Morphology and Function of Neandertal and Modern Human Ear Ossicles. Proc. Natl. Acad. Sci. USA 2016, 113, 11489–11494. [Google Scholar] [CrossRef]
- Stoessel, A.; Gunz, P.; David, R.; Spoor, F. Comparative Anatomy of the Middle Ear Ossicles of Extant Hominids—Introducing a Geometric Morphometric Protocol. J. Hum. Evol. 2016, 91, 1–25. [Google Scholar] [CrossRef]
- Almășan, O.; Leucuța, D.C.; Dinu, C.; Buduru, S.; Băciuț, M.; Hedeșiu, M. Petrotympanic Fissure Architecture and Malleus Location in Temporomandibular Joint Disorders. Tomography 2022, 8, 2460–2470. [Google Scholar] [CrossRef]
- de los Ríos, G.Y.L.; Espinosa, A.A.; Laguía, M.S.; Cano, F.G.; Gomariz, F.M.; Fernández, A.L.; Zarzosa, G.R. A Study of the Head during Prenatal and Perinatal Development of Two Fetuses and One Newborn Striped Dolphin (Stenella Coeruleoalba, Meyen 1833) Using Dissections, Sectional Anatomy, CT, and MRI: Anatomical and Functional Implications in Cetaceans and Terrestrial Mammals. Animals 2019, 9, 1139. [Google Scholar] [CrossRef]
- Carter, Y. Monkey Hear: A Morphometric Analysis of the Primate Auditory Ossicles. Master’s Thesis, University of Manitoba, Winnipeg, MB, Canada, 2009. [Google Scholar]
- Hemilä, S.; Nummela, S.; Reuter, T. What Middle Ear Parameters Tell about Impedance Matching and High Frequency Hearing. Hear Res. 1995, 85, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Topsakal, V.; Kachlik, D.; Bahşi, I.; Carlson, M.; Isaacson, B.; Broman, J.; Tubbs, R.S.; Baud, R.; ten Donkelaar, H.J. Relevant Temporal Bone Anatomy for Robotic Cochlear Implantation: An Updated Terminology Combined with Anatomical and Clinical Terms. Transl. Res. Anat. 2021, 25, 100138. [Google Scholar] [CrossRef]
- Arensburg, B.; Harell, M.; Nathan, H. The Human Middle Ear Ossicles: Morphometry, and Taxonomic Implications. J. Hum. Evol. 1981, 10, 199–205. [Google Scholar] [CrossRef]
- Noussios, G.; Chouridis, P.; Kostretzis, L.; Natsis, K. Morphological and Morphometrical Study of the Human Ossicular Chain: A Review of the Literature and a Meta-Analysis of Experience Over 50 Years. J. Clin. Med. Res. 2016, 8, 76. [Google Scholar] [CrossRef]
- Kurtul, I.; Cevik, A.; Bozkurt, E.U.; Dursun, N. A Detailed Subgross Morphometric Study on the Auditory Ossicles of the New Zealand Rabbit. J. Vet. Med. Ser. C Anat. Histol. Embryol. 2003, 32, 249–252. [Google Scholar] [CrossRef]
- Martonos, C.; Damian, A.; Gudea, A.I.; Bud, I.; Stan, G.F. Morphological and Morphometrical Study of the Auditory Ossicles in Chinchilla. J. Vet. Med. Ser. C Anat. Histol. Embryol. 2019, 48, 340–345. [Google Scholar] [CrossRef]
- Martonos, C.; Gudea, A.; Damian, A.; Lăcătuș, R.; Purdoiu, R.; Cocan, D.; Stan, F.G. Morphological and Morphometrical Aspects of the Auditory Ossicles in Goat (Capra Hircus). J. Vet. Med. Ser. C Anat. Histol. Embryol. 2021, 50, 184–191. [Google Scholar] [CrossRef]
- Matshes, E.W.; Juurlink, B. Human Osteology and Skeletal Radiology: An Atlas and Guide; CRC Press: Boca Raton, FL, USA, 2004; ISBN 9780429233258. [Google Scholar]
- Sarrat, R.; Garcia Guzman, A.; Torres, A. Morphology Variations of Human Ossicula Tympani. Acta Anat. 1988, 131, 146–149. [Google Scholar] [CrossRef]
- Unur, E.; Ülger, H.; Ekinci, N. Morphometrical and Morphological Variations of Middle Ear Ossicles in the Newborn. Erciyes Tip Dergisi 2002, 24, 57–63. [Google Scholar]
- Sodhi, S.; Chaudhary, P.; Arora, N. Histological and Histopathological Study of Incus. J. Clin. Diagn. Res. 2016, 10, AC01–AC03. [Google Scholar] [CrossRef] [PubMed]
- Borin, A.; Covolan, L.; Mello, L.E.; Okada, D.M.; Cruz, O.L.M.; Testa, J.R.G. Estudo Anatômico Do Osso Temporal de Um Primata Não-Humano (Callithrix Sp). Braz. J. Otorhinolaryngol. 2008, 74, 370–373. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.N.; Colbert, M.W. Correlations between Auditory Structures and Hearing Sensitivity in Non-Human Primates. J. Morphol. 2010, 271, 511–532. [Google Scholar] [CrossRef]
- Rosenberger, A.L.; Hershkovitz, P. Living New World Monkeys (Platyrrhini), with an Introduction to Primates. Syst. Zool. 1978, 27, 489. [Google Scholar] [CrossRef]
- Rosowski, J.J. Outer and Middle Ears. In Springer Handbook of Auditory Research Vol 4; Springer: New York, NY, USA, 1994; pp. 172–247. [Google Scholar]
- Fleischer, G. Evolutionary Principles of the Mammalian Middle Ear. Adv. Anat. Embryol. Cell Biol. 1978, 55, 7–50. [Google Scholar] [CrossRef]
- Maier, W.; Tröscher, A.; Ruf, I. The Anterior Process of the Malleus in Extant Lagomorpha (Mammalia). J. Morphol. 2018, 279, 132–146. [Google Scholar] [CrossRef]
- Kamali, Y.; Gholami, S.; Ahrari-Khafi, M.S.; Rasouli, B.; Shayegh, H. The Architecture of the Middle Ear in the Small Indian Mongoose (Herpestes Javanicus). Folia Morphol. 2015, 74, 340–345. [Google Scholar] [CrossRef]
- Mason, M.J.; Farr, M.R.B. Flexibility within the Middle Ears of Vertebrates. J. Laryngol. Otol. 2013, 127, 2–14. [Google Scholar] [CrossRef]
- Casteleyn, C.; Bakker, J.; Breugelmans, S.; Kondova, I.; Saunders, J.; Langermans, J.A.M.; Cornillie, P.; van den Broeck, W.; van Loo, D.; van Hoorebeke, L.; et al. Anatomical Description and Morphometry of the Skeleton of the Common Marmoset (Callithrix Jacchus). Lab. Anim. 2012, 46, 152–163. [Google Scholar] [CrossRef]
- Anwar, K.; Said, M.; Zada, B.; Din, G.; Khan, I.A. Cholesteatoma and the Destruction of Middle Ear and Mastoid Cavities in Ottorhoea. Pak. J. Med. Health Sci. 2012, 6, 51–54. [Google Scholar]
- Mathur, N.N.; Kakar, P.; Singh, T.; Sawhney, K.L. Ossicular Pathology in Unsafe Chronic Suppurative Otitis Media. Indian J. Otolaryngol. 1991, 43, 9–12. [Google Scholar] [CrossRef]
- Salvinelli, F.; Trivelli, M.; Greco, F.; Linthicum, J. Chronic Otitis Media: Histopathological Changes: A Post Mortem Study on Temporal Bones. Eur. Rev. Med. Pharmacol. Sci. 1999, 3, 175–178. [Google Scholar] [PubMed]
- Mason, M.J. Structure and Function of the Mammalian Middle Ear. II: Inferring Function from Structure. J. Anat. 2016, 228, 300–312. [Google Scholar] [CrossRef]
- Mason, M.J. Internally Coupled Ears in Living Mammals. Biol. Cybern. 2016, 110, 345–358. [Google Scholar] [CrossRef]
- Martins, L.L.; Almeida-Silva, I.; Rossato, M.; Murashima, A.A.B.; Hyppolito, M.A.; Machado, M.R.F. Macroscopic Description of the External and Middle Ear of Paca (Cuniculus Paca Linnaeus, 1766). Pesqui. Vet. Bras. 2015, 35, 583–589. [Google Scholar] [CrossRef]
- Argyle, E.C.; Mason, M.J. Middle Ear Structures of Octodon Degus (Rodentia: Octodontidae), in Comparison with Those of Subterranean Caviomorphs. J. Mammal. 2008, 89, 1447–1455. [Google Scholar] [CrossRef]
- Amin, S.; Tucker, A.S. Joint Formation in the Middle Ear: Lessons from the Mouse and Guinea Pig. Dev. Dyn. 2006, 235, 1326–1333. [Google Scholar] [CrossRef]
- Guan, M.; Zhang, J.; Jia, Y.; Cao, X.; Lou, X.; Li, Y.; Gao, X. Middle Ear Structure and Transcanal Approach Appropriate for Middle Ear Surgery in Rabbits. Exp. Ther. Med. 2019, 17, 1248–1255. [Google Scholar] [CrossRef]
- Carrasco, M.L.; Maass, O.J.C.; Dentone, S.L.; Miranda, G.G.; Kukuljan, P.M. Estudio Morfológico Del Oído Medio e Interno de La Chinchilla Laniger. Rev. De Otorrinolaringol. Cirugía De Cabeza Cuello 2008, 68, 263–274. [Google Scholar] [CrossRef]
- Wang, X.; Gan, R.Z. 3D Finite Element Model of the Chinchilla Ear for Characterizing Middle Ear Functions. Biomech. Model Mechanobiol. 2016, 15, 1263–1277. [Google Scholar] [CrossRef] [PubMed]
- Anthwal, N.; Joshi, L.; Tucker, A.S. Evolution of the Mammalian Middle Ear and Jaw: Adaptations and Novel Structures. J. Anat. 2013, 222, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Beșoluk, K.; Orhun Dayan, M.; Eken, E.; Turgut, N.; Aydogdu, S. Macroanatomic Structure and Morphometric Analysis of Middle Ear in Ostrich (Struthio Camelus). Arch. Vet. Sci. Med. 2019, 2, 8–16. [Google Scholar] [CrossRef]
- Muyshondt, P.G.G.; Claes, R.; Aerts, P.; Dirckx, J.J.J. Sound Attenuation in the Ear of Domestic Chickens (Gallus Gallus Domesticus) as a Result of Beak Opening. R. Soc. Open Sci. 2017, 4, 171286. [Google Scholar] [CrossRef] [PubMed]
- Mills, R. Applied Comparative Anatomy of the Avian Middle Ear. J. R. Soc. Med. 1994, 87, 155–156. [Google Scholar] [CrossRef]
- Takechi, M.; Kuratani, S. History of Studies on Mammalian Middle Ear Evolution: A Comparative Morphological and Developmental Biology Perspective. J. Exp. Zool. B Mol. Dev. Evol. 2010, 314B, 417–433. [Google Scholar] [CrossRef]
- Anthwal, N.; Thompson, H. The Development of the Mammalian Outer and Middle Ear. J. Anat. 2016, 228, 217–232. [Google Scholar] [CrossRef]
- Simaei, N.; Soltanalinejad, F.; Najafi, G.; Shalizar Jalali, A. Anatomical and Morphometrical Study of Middle Ear Ossicles in 2 to 3-Month-Old Makouei Sheep Fetuses. Vet. Res. Forum 2017, 8, 237–241. [Google Scholar]
- Wadhwa, S.; Kaul, J.M.; Agarwal, A.K. Morphometric Study of Stapes and Its Clinical Implications. J. Anat. Soc. India 2005, 54, 1–9. [Google Scholar]
- Qvist, M.; Grøntved, A.M. Auditory Ossicles in Archaeological Skeletal Material from Medieval Denmark. In Acta Oto-Laryngologica, Supplement; Taylor and Francis Ltd.: Abingdon, UK, 2000. [Google Scholar]
- Anson, B.; Bast, T.H. The Development of the Otic Capsule in the Region of the Vestibular Aqueduct. Q. Bull. Northwestern Univ. Med. Sch. 1951, 25, 96–107. [Google Scholar] [CrossRef]
- Anson, B.J.; Harper, D.G.; Hanson, J.R. LIV Vascular Anatomy of the Auditory Ossicles and Petrous Part of the Temporal Bone in Man. Ann. Otol. Rhinol. Laryngol. 1962, 71, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, A.A.S.; Rossato, M.; de Oliveira, J.A.A.; Hyppolito, M.A. Conhecimento Da Anatomia Da Orelha de Cobaias e Ratos e Sua Aplicação Na Pesquisa Otológica Básica. Braz. J. Otorhinolaryngol. 2009, 75, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Mohammadpour, A.A. Morphology and Morphometrical Study of Hamster Middle Ear Bones. Iran J. Vet. Res. 2011, 12, 121–126. [Google Scholar] [CrossRef]
- Rodríguez-Vázquez, J.F. Development of the Stapes and Associated Structures in Human Embryos. J. Anat. 2005, 207, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, G. Endoscopic Anatomy of the Pediatric Middle Ear. Otolaryngol. Head Neck Surg. 2014, 150, 6–15. [Google Scholar] [CrossRef]
- Potu, B.K.; Nayak, S.R.; Jaradat, A.A. Morphometrics of the Stapedius Muscle: A Systematic Review of Cadaveric Studies. Eur. J. Anat. 2021, 25, 117–124. [Google Scholar]
- Hitier, M.; Zhang, M.; Labrousse, M.; Barbier, C.; Patron, V.; Moreau, S. Persistent Stapedial Arteries in Human: From Phylogeny to Surgical Consequences. Surg. Radiol. Anat. 2013, 35, 883–891. [Google Scholar] [CrossRef]
- Padmini, P.; Rao, B.N. Morphological Variations in Human Fetal Ear Ossicles—A Study. Int. J. Anat. Res. 2013, 1, 40–42. [Google Scholar]
- Rolvien, T.; Schmidt, F.N.; Milovanovic, P.; Jähn, K.; Riedel, C.; Butscheidt, S.; Püschel, K.; Jeschke, A.; Amling, M.; Busse, B. Early Bone Tissue Aging in Human Auditory Ossicles Is Accompanied by Excessive Hypermineralization, Osteocyte Death and Micropetrosis. Sci. Rep. 2018, 8, 1920. [Google Scholar] [CrossRef]
- Farahani, R.M.; Nooranipour, M. Anatomy and Anthropometry of Human Stapes. Am. J. Otolaryngol. 2008, 29, 42–47. [Google Scholar] [CrossRef]
- Rosowski, J.J. Comparative Middle Ear Structure and Function in Vertebrates. In The Middle Ear; Springer Handbook of Auditory Research; Puria, S., Fay, R., Popper, A., Eds.; Springer: New York, NY, USA, 2013; pp. 31–65. [Google Scholar] [CrossRef]
- Koyabu, D.; Hosojima, M.; Endo, H. Into the Dark: Patterns of Middle Ear Adaptations in Subterranean Eulipotyphlan Mammals. R Soc. Open Sci. 2017, 4, 170608. [Google Scholar] [CrossRef] [PubMed]
- Heffner, R.S. Primate Hearing from a Mammalian Perspective. Anat. Rec. A Discov. Mol. Cell Evol. Biol. 2004, 281A, 1111–1122. [Google Scholar] [CrossRef] [PubMed]
- Kojima, S. Comparison of Auditory Functions in the Chimpanzee and Human. Folia Primatol. 1990, 55, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Price, T.; Fischer, J. Meaning Attribution in the West African Green Monkey: Influence of Call Type and Context. Anim. Cogn. 2014, 17, 277. [Google Scholar] [CrossRef]
- Stebbins, W.C.; Pearson, R.D.; Moody, D.B. Hearing in the Monkey (Macaca): Absolute and Differential Sensitivity. J. Acoust. Soc. Am. 2005, 47, 67. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| X-axis | Midpoint of the minimum neck width—the most noticeable point along the top of the head |
| Y-axis | Most inferior point of the short process and the manubrium |
| 1. Total length | Tip of the manubrium to the top of the head |
| 2. Manubrium length | Tip of the short process to the tip of the manubrium following X-axis |
| 3. Manubrium M-L thickness | M-L thickness of the manubrium at mid manubrium length, perpendicular to X-axis |
| 4. Manubrium arc depth | Maximum depth of the curvature of the arc of the manubrium, following X-axis |
| 5. Corpus length | Tip of the head to the lower border of the manubrium following X-axis |
| 6. Neck width | Anterior and posterior borders of the neck |
| 7. S-L head width | Maximum distance between 2 parallel lines marking the widest points of the margin of the head, taken following the X-axis |
| 8. Angle between axes | X-Y angle |
| Manubrium/length index | (Manubrium length/total length) × 100 |
| Manubrium robusticity index | (Manubrium ML thickness/corpus length) × 100 |
| Manubrium/corpus index | (Manubrium length/corpus length) × 100 |
| Corpus/length index | (Corpus length/total length) × 100 |
| X-axis | Line that joins the most salient point along the anterior portion of the superior border of the body |
| Y-axis | Line that joins the tip of the long process to the most salient point along the superior border of the body |
| Z-axis | Line joining the tip of the long process to the most external point along the margin of the anterior facet |
| 9. Short process length | Maximum distance from the tip of the short process to the most salient point along the anterior portion of the superior border of the body, following X-axis |
| 10. Long process length | Maximum distance from the tip of the long process to the most salient point along the superior border of the body |
| 11. Functional length of the long process | Perpendicular distance from the Z-axis (rotational axis) to the tip of the long process |
| 12. Arc depth of the long process | Maximum depth of the arc along the long process measured from the plane defined by the lateral outmost point along the tip of the long process |
| 13. Articular facet height | Max height of the articular facet with the bone oriented along the rotational axis |
| 14. Angle between the axes | Angle formed by the X- and Y-axes |
| 15. Interprocess length | Maximum distance between the most salient points along the superior margin of the short process and the tip of the long process |
| 16. Interprocess arc depth | Maximum depth of the curvature between the short and long process tips |
| Incudal index | 9/10 × 100 |
| Long process index | 11/10 × 100 |
| Relative articular facet height | 13/10 × 100 |
| X-axis | Line joining the antero-superior corner of the footplate and the tip of the head |
| Y-axis | Line joining the posterior superior corner of the footplate and the tip of the head |
| Z-axis | Line joining the most inferior points along the footplate margin anteriorly and posteriorly |
| 19. Total height of the stapes | Maximum height from the lower margin of the footplate to the tip of the head perpendicular to the Z-axis |
| 20. Head height | Minimum distance between the superior margin of the obturator foramen and the top of the head, measured perpendicular to the Z-axis |
| 21. Obturator foramen height | Maximum height of the obturator foramen measured perpendicular to the Z-axis |
| 22. Obturator foramen width | Maximum width of the obturator foramen measured parallel to the Z-axis |
| 23. Maximum width of the crura | Maximum width across the anterior and posterior crura, measured on the external aspect and parallel to the Z-axis |
| 24. Posterior crus length | Maximum distance from the posterior superior corner of the footplate to the tip of the head, following Y-axis |
| 25. Posterior crus arc depth | Maximum depth of the curvature of the posterior crus measured parallel to the Y-axis |
| 26. Anterior crus length | Maximum distance from the antero-superior corner of the footplate to the tip of the head following X-axis |
| 27. Anterior crus arc depth | Maximum depth of the curvature of the anterior crus measured parallel to the X-axis |
| 28. Angle A | Angle between the anterior and posterior crura or between the X- and Y-axes |
| 29. Angle B | Angle between the anterior crus and the footplate or between the X- and Z-axes |
| 30. Angle C | Angle between the posterior crus and the footplate between Y- and Z-axes |
| 31. Footplate length | Maximum length of the footplate |
| 32. Footplate width | Maximum width of the footplate |
| 33. Footplate area | Measured area of the footplate |
| Stapedial index | 31/19 × 100 |
| Relative head height | 20/19 × 100 |
| Obturator foramen index | 21/22 × 100 |
| Footplate index | 31/32 × 100 |
| Crural index | 36/24 × 100 |
| Specimen | Measurement Number [5] | Mean Values—Right Side | Mean Values—Left Side | Mean Values |
|---|---|---|---|---|
| M1 | 9 | 2.475 mm | 2.475 mm | |
| 10 | 3.23 mm | 3.23 mm | ||
| 11 | 2.18 mm | 2.18 mm | ||
| 12 | 2.56 mm | 2.56 mm | ||
| 13 | 1.025 mm | 1.025 mm | ||
| 14 | 56.43° | 56.43° | ||
| 15 | 2.445 mm | 2.445 mm | ||
| 16 | 0.82 mm | 0.82 mm | ||
| M2 | 9 | 2.68 mm | 2.51 mm | 2.57 mm |
| 10 | 3.37 mm | 3.24 mm | 3.28 mm | |
| 11 | 2.21 mm | 2.16 mm | 2.18 mm | |
| 12 | 2.56 mm | 2.44 mm | 2.48 mm | |
| 13 | 1.18 mm | 1.14 mm | 1.15 mm | |
| 14 | 49.8° | 52.54° | 51.6° | |
| 15 | 2.55 mm | 2.44 mm | 2.47 mm | |
| 16 | 0.8 mm | 0.68 mm | 0.75 mm | |
| M3 | 9 | 2.47 mm | 2.39 mm | 2.40 mm |
| 10 | 2.95 mm | 3.014 mm | 3.0 mm | |
| 11 | 2.13 mm | 2.06 mm | 2.07 mm | |
| 12 | 2.38 mm | 2.38 mm | 2.38 mm | |
| 13 | 1.12 mm | 1.18 mm | 1.17 mm | |
| 14 | 46.1° | 56.8° | 55.08° | |
| 15 | 2.09 mm | 2.42 mm | 2.36 mm | |
| 16 | 0.78 mm | 0.70 mm | 0.71 mm | |
| M4 | 9 | 2.36 mm | 2.59 mm | 2.42 mm |
| 10 | 3.18 mm | 3.175 mm | 3.18 mm | |
| 11 | 2.21 mm | 2.05 mm | 2.17 mm | |
| 12 | 2.57 mm | 2.445 mm | 2.54 mm | |
| 13 | 1.31 mm | 1.19 mm | 1.28 mm | |
| 14 | 52.53° | 54.49° | 53.02° | |
| 15 | 2.38 mm | 2.43 mm | 2.39 mm | |
| 16 | 0.69 mm | 0.68 mm | 0.69 mm | |
| M5 | 9 | 2.72 mm | 2.56 mm | 2.61 mm |
| 10 | 3.45 mm | 3.36 mm | 3.39 mm | |
| 11 | 2.23 mm | 2.20 mm | 2.21 mm | |
| 12 | 2.43 mm | 2.46 mm | 2.45 mm | |
| 13 | 1.35 mm | 1.28 mm | 1.30 mm | |
| 14 | 55.13° | 53.56° | 54.08° | |
| 15 | 2.78 mm | 2.54 mm | 2.62 mm | |
| 16 | 0.59 mm | 0.65 mm | 0.63 mm | |
| M6 | 9 | 2.61 mm | 2.61 mm | |
| 10 | 3.37 mm | 3.37 mm | ||
| 11 | 2.31 mm | 2.31 mm | ||
| 12 | 2.61 mm | 2.61 mm | ||
| 13 | 1.54 mm | 1.54 mm | ||
| 14 | 58.45° | 58.45° | ||
| 15 | 2.65 mm | 2.65 mm | ||
| 16 | 0.74 mm | 0.74 mm |
| Specimen | Measurement Number [5] | Mean Values—Right Side | Mean Values—Left Side | Mean Values |
|---|---|---|---|---|
| M2 | 19 | 1.54 mm | 1.54 mm | |
| 20 | 0.61 mm | 0.61 mm | ||
| 21 | 0.93 mm | 0.93 mm | ||
| 22 | 0.74 mm | 0.74 mm | ||
| 23 | 1.31 mm | 1.31 mm | ||
| 24 | 1.77 mm | 1.77 mm | ||
| 25 | 0.22 mm | 0.22 mm | ||
| 26 | 1.82 mm | 1.82 mm | ||
| 27 | 0.22 mm | 0.22 mm | ||
| 28 | 59.05 | 59.05 | ||
| 29 | 61.22 | 61.22 | ||
| 30 | 56.3 | 56.3 | ||
| 31 | 1.81 mm | 1.81 mm | ||
| 32 | 0.722 mm | 0.722 mm | ||
| 33 | 0.904 mm2 | 0.904 mm2 | ||
| M3 | 19 | 1.54 mm | 1.54 mm | |
| 20 | 0.61 mm | 0.61 mm | ||
| 21 | 0.93 mm | 0.93 mm | ||
| 22 | 0.74 mm | 0.74 mm | ||
| 23 | 1.31 mm | 1.31 mm | ||
| 24 | 1.77 mm | 1.77 mm | ||
| 25 | 0.22 mm | 0.22 mm | ||
| 26 | 1.82 mm | 1.82 mm | ||
| 27 | 0.22 mm | 0.22 mm | ||
| 28 | 59.05 | 59.05 | ||
| 29 | 61.22 | 61.22 | ||
| 30 | 56.3 | 56.3 | ||
| 31 | 1.81 mm | 1.81 mm | ||
| 32 | 0.722 mm | 0.722 mm | ||
| 33 | 0.904 mm2 | 0.904 mm2 | ||
| M4 | 19 | 1.65 mm | 1.65 mm | |
| 20 | 0.64 mm | 0.64 mm | ||
| 21 | 1.01 mm | 1.01 mm | ||
| 22 | 0.806 mm | 0.806 mm | ||
| 23 | 1.35 mm | 1.35 mm | ||
| 24 | 1.83 mm | 1.83 mm | ||
| 25 | 0.18 mm | 0.18 mm | ||
| 26 | 1.876 mm | 1.876 mm | ||
| 27 | 0.176 mm | 0.176 mm | ||
| 28 | 58.93 | 58.93 | ||
| 29 | 62.26 | 62.26 | ||
| 30 | 60.36 | 60.36 | ||
| 31 | 1.853 mm | 1.853 mm | ||
| 32 | - | - | ||
| 33 | ||||
| M5 | 19 | 1.65 mm | 1.65 mm | |
| 20 | 0.673 mm | 0.673 mm | ||
| 21 | 0.976 mm | 0.976 mm | ||
| 22 | 0.793 mm | 0.793 mm | ||
| 23 | 1.253 mm | 1.253 mm | ||
| 24 | 1.856 mm | 1.856 mm | ||
| 25 | 0.213 mm | 0.213 mm | ||
| 26 | 1.863 mm | 1.863 mm | ||
| 27 | 0.193 mm | 0.193 mm | ||
| 28 | 53.96 | 53.96 | ||
| 29 | 63.83 | 63.83 | ||
| 30 | 58.96 | 58.96 | ||
| 31 | 1.653 mm | 1.653 mm | ||
| 32 | 0.813 mm | 0.813 mm | ||
| 33 | 0.974 mm2 | 0.974 mm2 | ||
| M6 | 19 | 1.595 mm | 1.68 mm | 1.658 mm |
| 20 | 0.685 mm | 0.765 mm | 0.738 mm | |
| 21 | 0.91 mm | 0.925 mm | 0.92 mm | |
| 22 | 0.79 mm | 0.79 mm | 0.79 mm | |
| 23 | 1.36 mm | 1.29 mm | 1.315 mm | |
| 24 | 1.845 mm | 1.92 mm | 1.896 mm | |
| 25 | 0.26 mm | 0.227 mm | 0.238 mm | |
| 26 | 1.975 mm | 2.045 mm | 2.02 mm | |
| 27 | 0.235 mm | 0.245 mm | 0.241 mm | |
| 28 | 55.3 | 51.46 | 52.74 | |
| 29 | 61.9 | 62.1 | 62.11 | |
| 30 | 62.69 | 64.6 | 63.96 | |
| 31 | 1.669 mm | 1.684 mm | 1.678 mm | |
| 32 | 0.83 mm | 0.813 mm | 0.82 mm | |
| 33 | 1.03 mm2 | 0.975 mm2 | 0.997 mm2 |
| Specimen | Measurement Number | Mean Values—Right Side | Mean Values—Left Side | Mean Values |
|---|---|---|---|---|
| M1 | 1 | 4.73 mm | 4.86 mm | 4.79 mm |
| 2 | ||||
| 3 | 0.39 mm | 0.43 mm | 0.41 mm | |
| 4 | ||||
| 5 | ||||
| 6 | 0.55 mm | 0.54 mm | 0.54 mm | |
| 7 | 1.63 mm | 1.60 mm | 1.60 mm | |
| 8 | - | - | - | |
| M2 | 1 | 4.84 mm | 4.76 mm | 4.81 mm |
| 2 | 3.02 mm | 2.96 mm | 3.01 mm | |
| 3 | 0.35 mm | 0.39 mm | 0.36 mm | |
| 4 | 0.26 mm | 0.26 mm | 0.26 mm | |
| 5 | 3.38 mm | 3.10 mm | 3.35 mm | |
| 6 | 0.58 mm | 0.55 mm | 0.57 mm | |
| 7 | 1.26 mm | 1.30 mm | 1.27 mm | |
| 8 | 149.7° | 149.7° | 149.7° | |
| M3 | 1 | 4.66 mm | 4.64 mm | 4.65 mm |
| 2 | ||||
| 3 | 0.40 mm | 0.43 mm | 0.41 mm | |
| 4 | ||||
| 5 | ||||
| 6 | 0.57 mm | 0.56 mm | 0.56 mm | |
| 7 | 1.34 mm | 1.40 mm | 1.37 mm | |
| 8 | ||||
| M4 | 1 | 4.94 mm | 4.94 mm | 4.94 mm |
| 2 | 2.86 mm | 2.89 mm | 2.88 mm | |
| 3 | 0.40 mm | 0.40 mm | 0.40 mm | |
| 4 | 0.27 mm | 0.29 mm | 0.28 mm | |
| 5 | 3.01 mm | 3.09 mm | 3.05 mm | |
| 6 | 0.58 mm | 0.56 mm | 0.57 mm | |
| 7 | 1.43 mm | 1.36 mm | 1.39 mm | |
| 8 | 145.1° | 150.7° | 148.12° | |
| M5 | 1 | 4.84 mm | 4.93 mm | 4.88 mm |
| 2 | 3.14 mm | 3.14 mm | 3.14 mm | |
| 3 | 0.38 mm | 0.45 mm | 0.41 mm | |
| 4 | 0.30 mm | 0.26 mm | 0.28 mm | |
| 5 | 3.05 mm | 2.91 mm | 2.99 mm | |
| 6 | 0.55 mm | 0.55 mm | 0.55 mm | |
| 7 | 1.33 mm | 1.35 mm | 1.34 mm | |
| 8 | 152.73° | 145.3 | 149.56° | |
| M6 | 1 | 4.99 mm | 5.07 mm | 5.03 mm |
| 2 | 3.15 mm | 3.15 mm | ||
| 3 | 0.40 mm | 0.39 mm | 0.39 mm | |
| 4 | 0.18 mm | 0.18 mm | ||
| 5 | 3.79 mm | 3.79 mm | ||
| 6 | 0.55 mm | 0.54 mm | 0.55 mm | |
| 7 | 1.43 mm | 1.52 mm | 1.48 mm | |
| 8 | - | 154.13° | 154.13° |
| Measurement No. | Right from Left | Male vs. Female |
|---|---|---|
| 1 | t = −1.032; p = 0.151901 The result is not significant at p < 0.05 Mean value for right = 4.85 (n = 81); mean value for left = 4.55 (n = 65) | t= −2.0770; p = 0.039581 The result is significant at p < 0.05 Mean value for females = 4.81; mean value for males = 4.88 |
| 3 | t = −3.10126; p = 0.001161 The result is significant at p < 0.05 Mean value = 0.38 (n = 80); mean value for left = 0.41 (n = 65) | t= 2.077; p = 0.00196 The result is significant at p < 0.05 Mean value for males = 0.4 (n = 113); mean value for females = 0.37 (n = 32) |
| 6 | t = 1.45139; p = 0.074431 The result is not significant at p < 0.05 Mean value for right = 0.57 (n = 81); mean value for left = 0.55 (n = 64) | t = −0.57247; p = 0.283953 The result is not significant at p < 0.05 Mean value for males = 0.56 (n = 112); mean value for females = 0.57 (n = 33) |
| 7 | t = −1.279; p = 0.101485 The result is not significant at p < 0.05 Mean value for right = 1.37 (n = 80); mean value for left = 1.39 (n = 65) | t = 2.15017; p = 0.016612 The result is significant at p < 0.05 Mean value for males = 1.39 (n = 113); mean value for females = 1.34 (n = 32) |
| Index | Value (Average/Variation) |
|---|---|
| Incudal index | 77.76 (69.7–87.16) |
| Long process index | 67.30 (61.3–86.9) |
| Relative articular facet height | 38.66 (30.69–50.33) |
| Malleus Index | Value (Average/Variation) |
|---|---|
| Manubrium/length index | 62.36 (57.9–72.4) |
| Manubrium robusticity index | 12.19 (6.2–17.02) |
| Manubrium/corpus index | 96.38 (77.88–118.32) |
| Corpus/length index | 64.0 (53.5–87.7) |
| Index | Value (Average/Min and Max) |
|---|---|
| Stapedial index | 108.67 (100–117) |
| Relative head height | 41.48 (37.03–48.2) |
| Obturator foramen index | 124.13 (106.4–138.23) |
| Footplate index | 214.29 (176.7–270) |
| Crural index | 102.16 (89.28–11.95) |
| Specimen Number | Functional Length of Malleus | Functional Length of Long Process | Lever Ratio |
|---|---|---|---|
| M1 | No available data | 2.18 (n = 2) | |
| M2 | 3.01 (n = 12) | 2.18 (n = 9) | 1.38 |
| M3 | No available data | 2.07 (n = 6) | |
| M4 | 2.88 (n = 16) | 2.17 (n = 8) | 1.32 |
| M5 | 3.14 (n = 14) | 2.21 (n = 9) | 1.42 |
| M6 | 3.15 (n = 3) | 2.31 (n = 4) | 1.36 |
| Average value | 3.045 | 2.31 | 1.3730 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martonos, C.O.; Gudea, A.I.; Ratiu, I.A.; Stan, F.G.; Bolfă, P.; Little, W.B.; Dezdrobitu, C.C. Anatomical, Histological, and Morphometrical Investigations of the Auditory Ossicles in Chlorocebus aethiops sabaeus from Saint Kitts Island. Biology 2023, 12, 631. https://doi.org/10.3390/biology12040631
Martonos CO, Gudea AI, Ratiu IA, Stan FG, Bolfă P, Little WB, Dezdrobitu CC. Anatomical, Histological, and Morphometrical Investigations of the Auditory Ossicles in Chlorocebus aethiops sabaeus from Saint Kitts Island. Biology. 2023; 12(4):631. https://doi.org/10.3390/biology12040631
Chicago/Turabian StyleMartonos, Cristian Olimpiu, Alexandru Ion Gudea, Ioana A. Ratiu, Florin Gheorghe Stan, Pompei Bolfă, William Brady Little, and Cristian Constantin Dezdrobitu. 2023. "Anatomical, Histological, and Morphometrical Investigations of the Auditory Ossicles in Chlorocebus aethiops sabaeus from Saint Kitts Island" Biology 12, no. 4: 631. https://doi.org/10.3390/biology12040631