
A Highly Diverse Olenekian Brachiopod Fauna from the Nanpanjiang Basin, South China, and Its Implications for the Early Triassic Biotic Recovery

Abstract
:Simple Summary
Abstract
1. Introduction
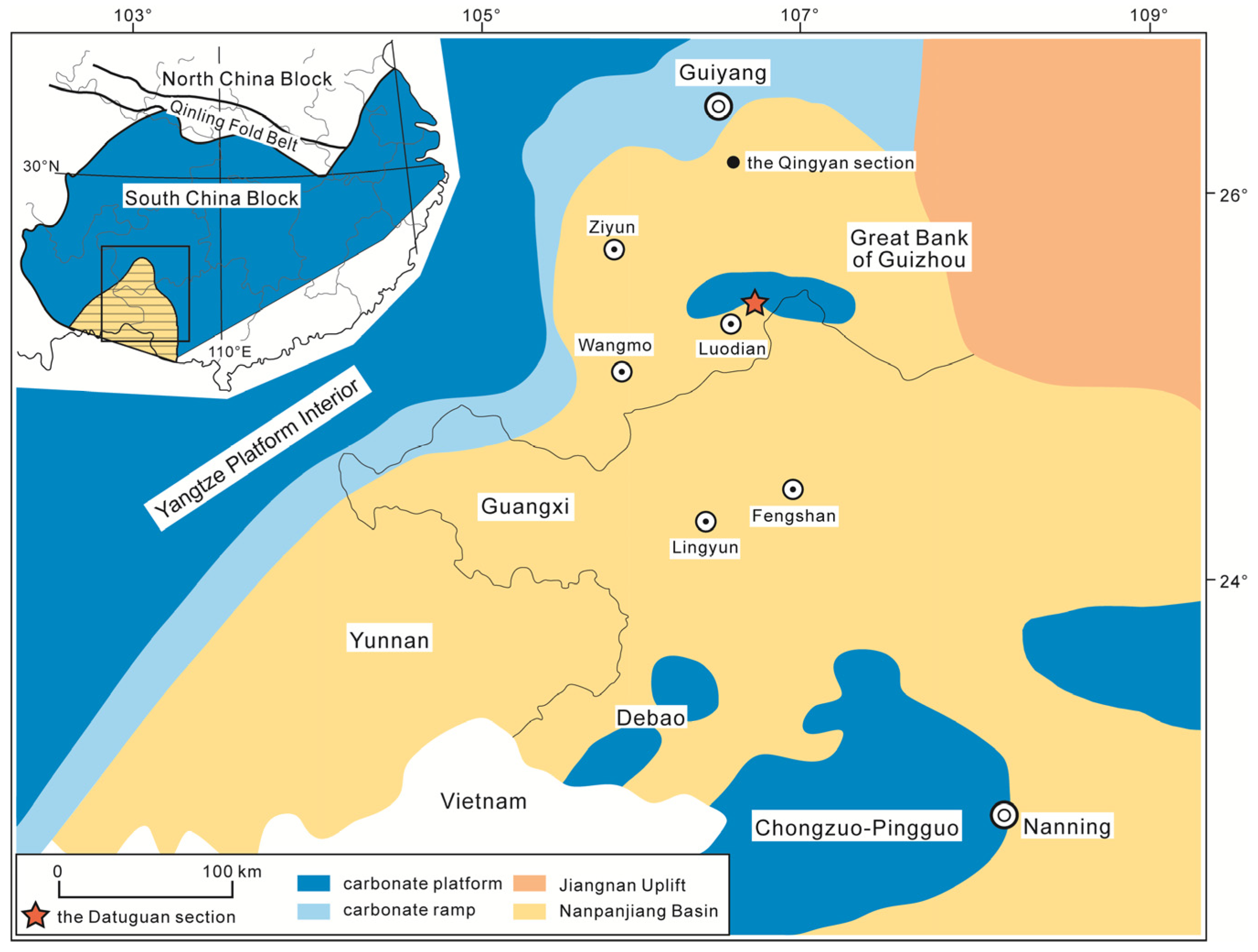
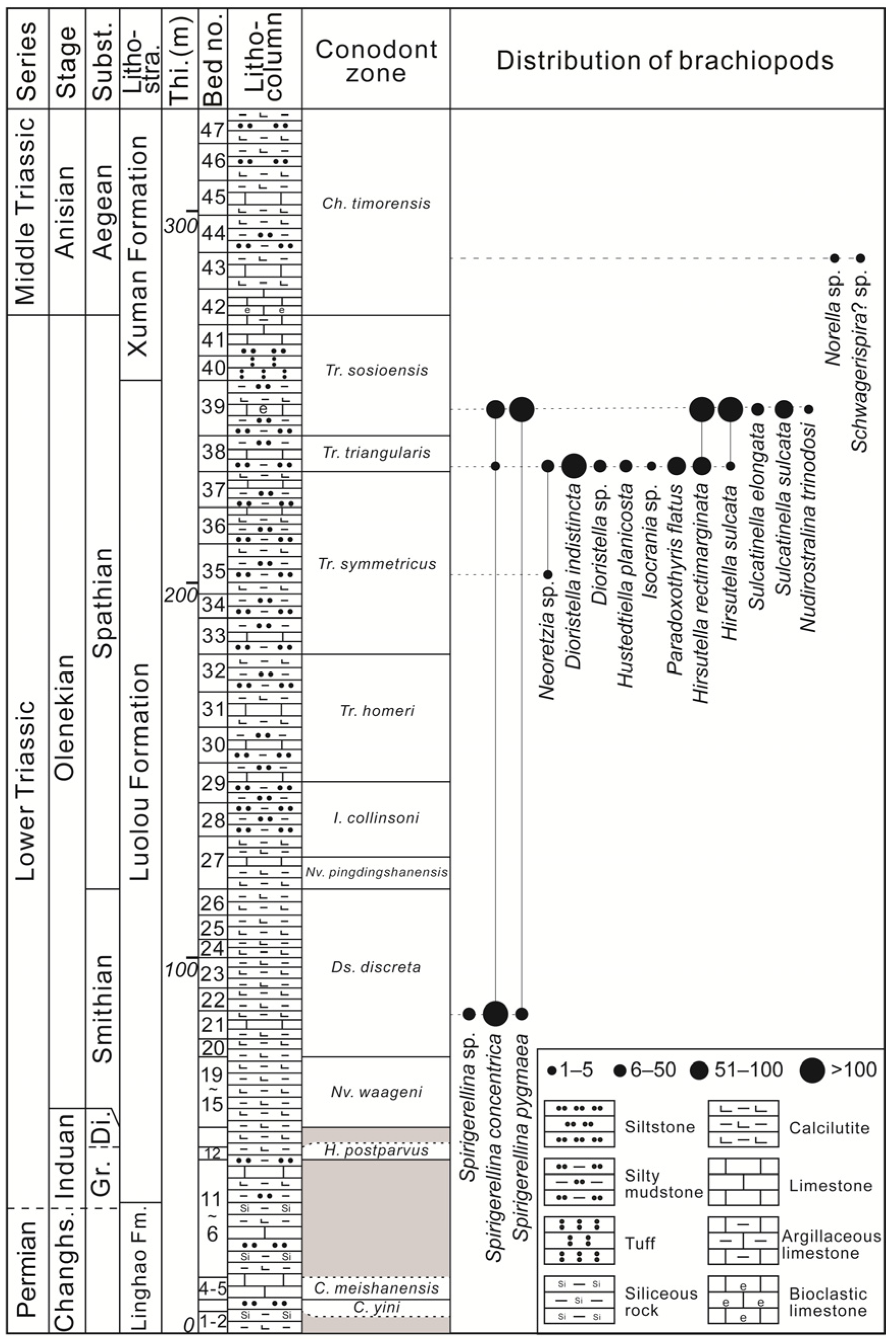
2. Geological Settings and Age
3. Materials and Methods
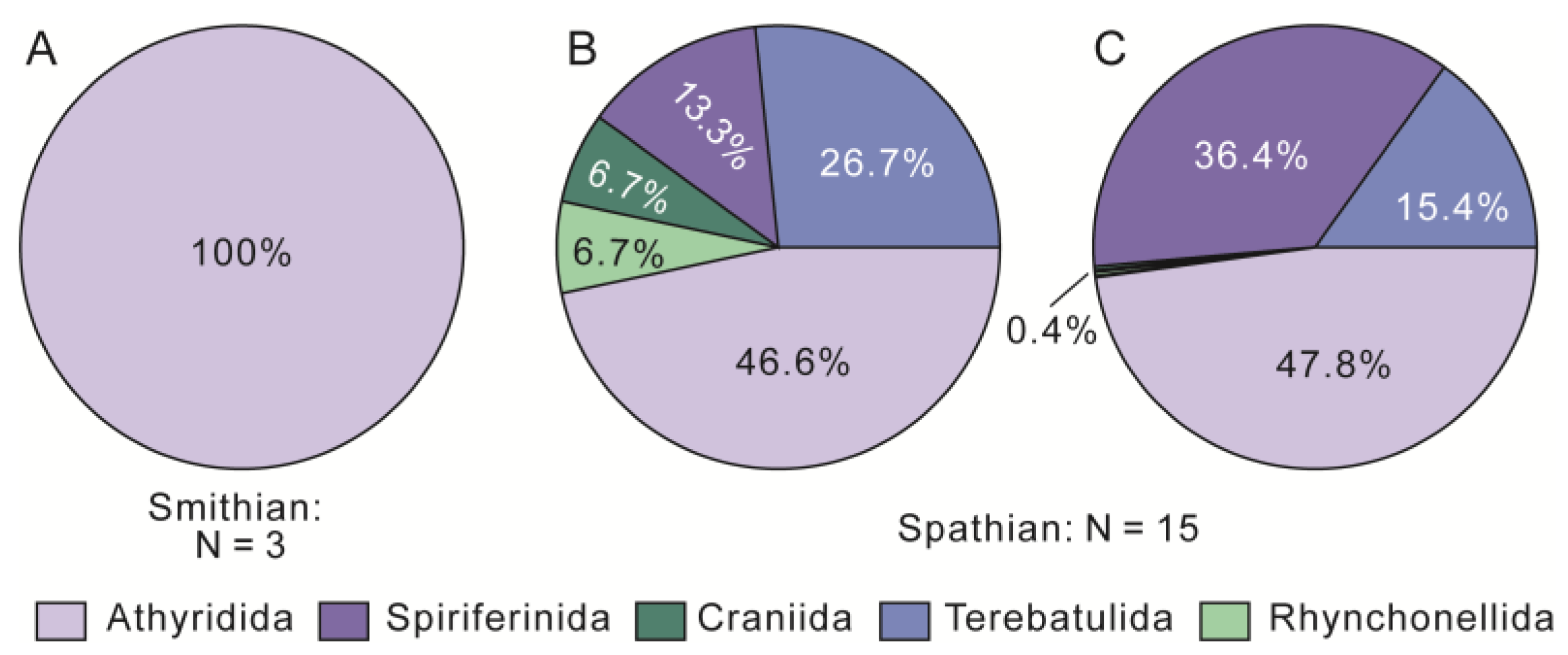
4. Results
5. Systematic Palaeontology
- Order Spiriferinida Ivanova, 1972a [44]
- Suborder Cyrtinidina Carter and Johnson in Carter, et al., 1994 [45]
- Superfamily Suessioidea Waagen, 1883 [46]
- Family Bittnerulidae Schuchert, 1929 [47]
- Subfamily Hirsutellinae Xu and Liu in Yang, et al., 1983 [22]
- Genus Hirsutella Cooper and Muir-Wood, 1951 [48]
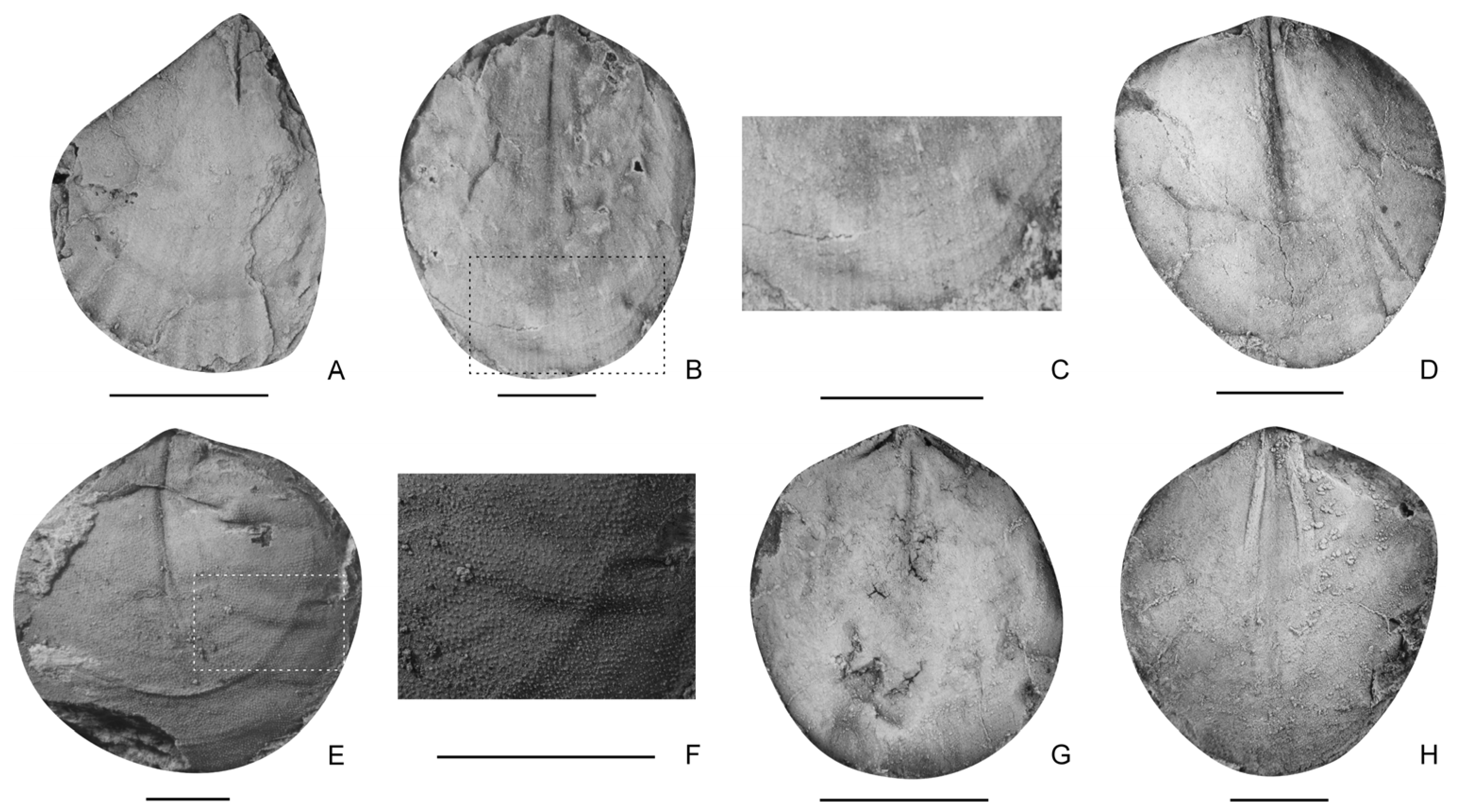
- Hirsutella sulcata sp. nov. (Figure 6F–L)

- Order Terebratulida Waagen, 1883 [46]
- Suborder Terebratulidina Waagen, 1883 [46]
- Superfamily Dielasmatoidea Schuchert, 1913 [53]
- Family Angustothyrididae Dagys, 1972b [54]
- Genus Paradoxothyris Xu, 1978 [55]
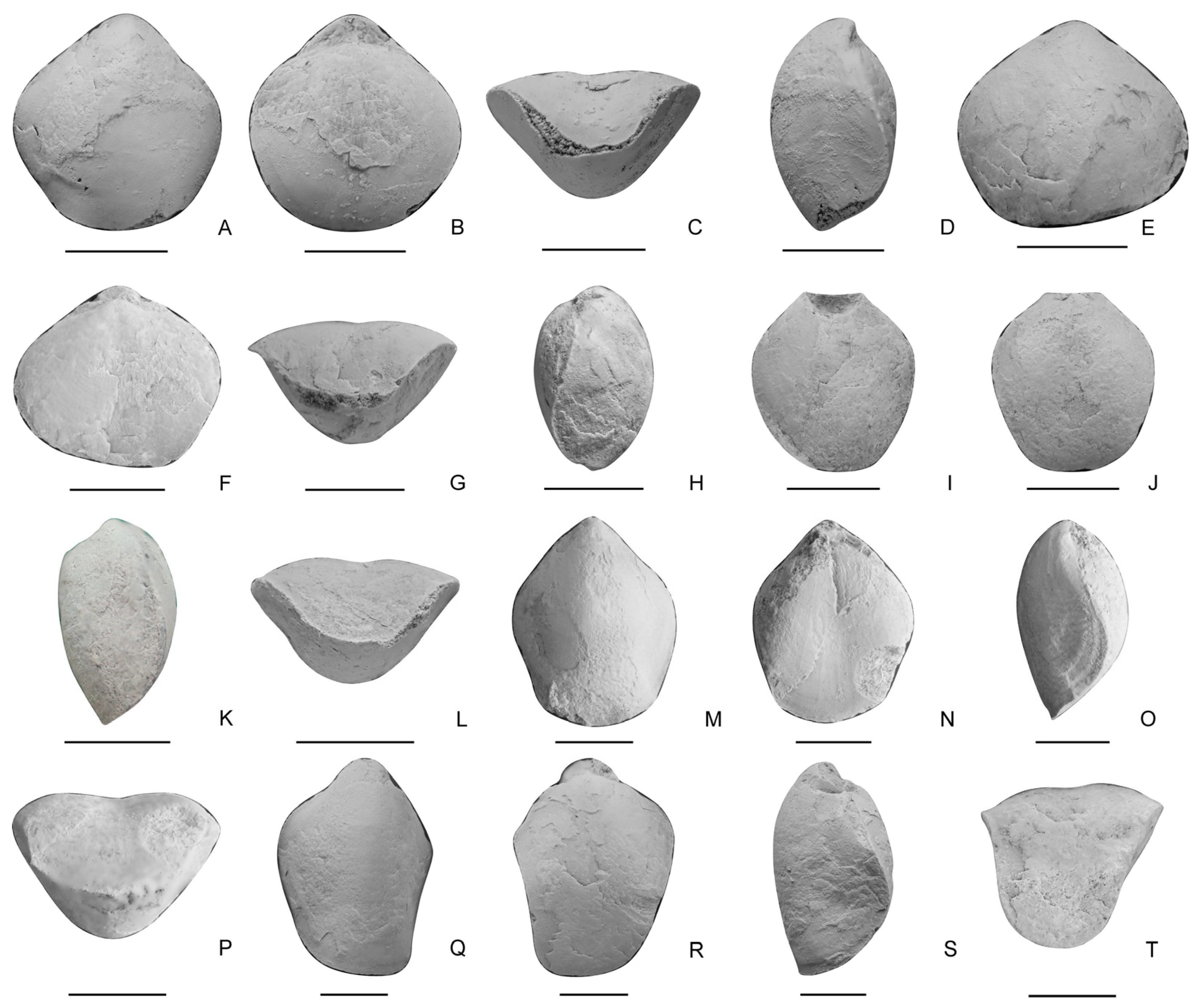
- Paradoxothyris flatus sp. nov. (Figure 7)

- Family Dielasmatidae Schuchert, 1913 [53]
- Subfamily Dielasmatinae Schuchert, 1913 [53]
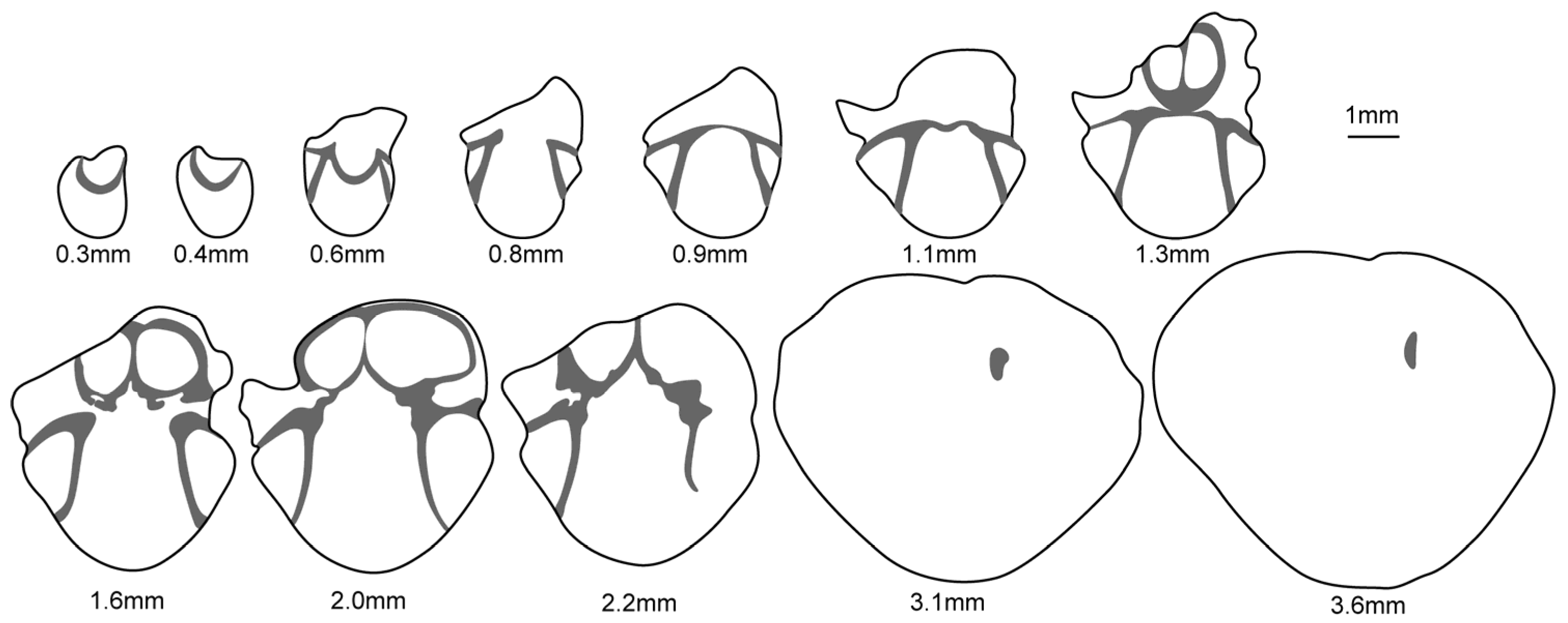
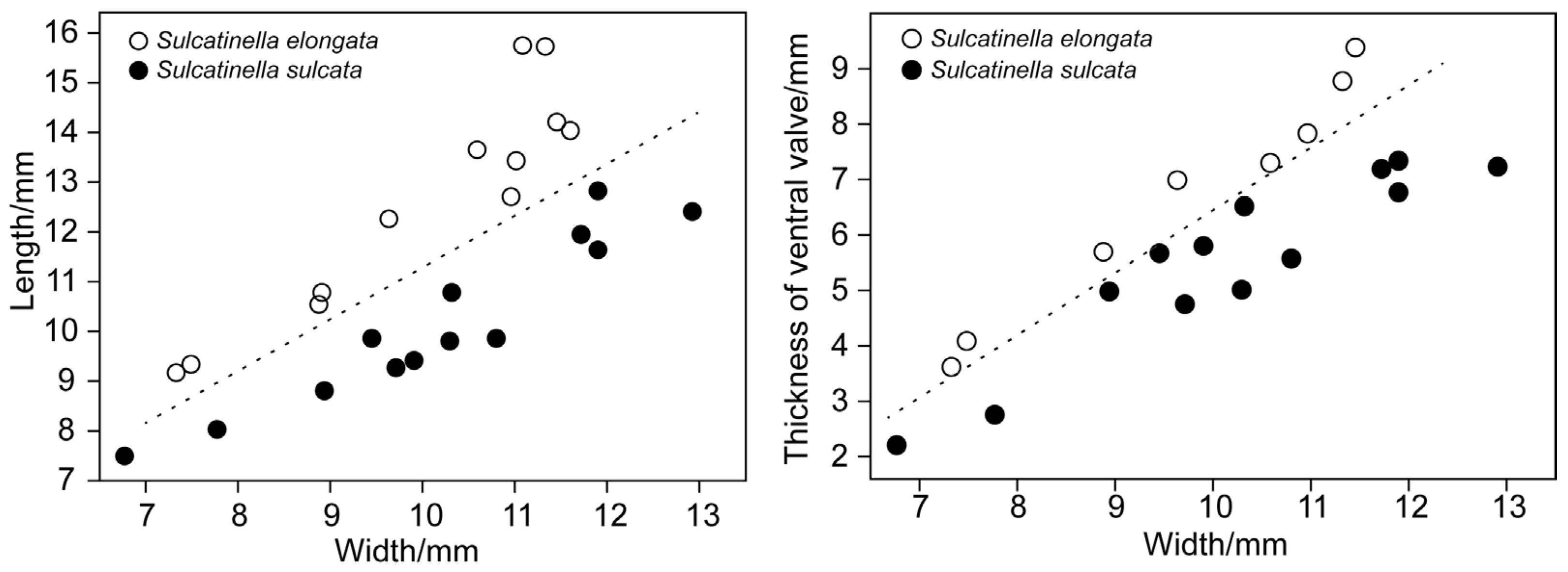
- Genus Sulcatinella Dagys, 1974 [57]



6. Discussion
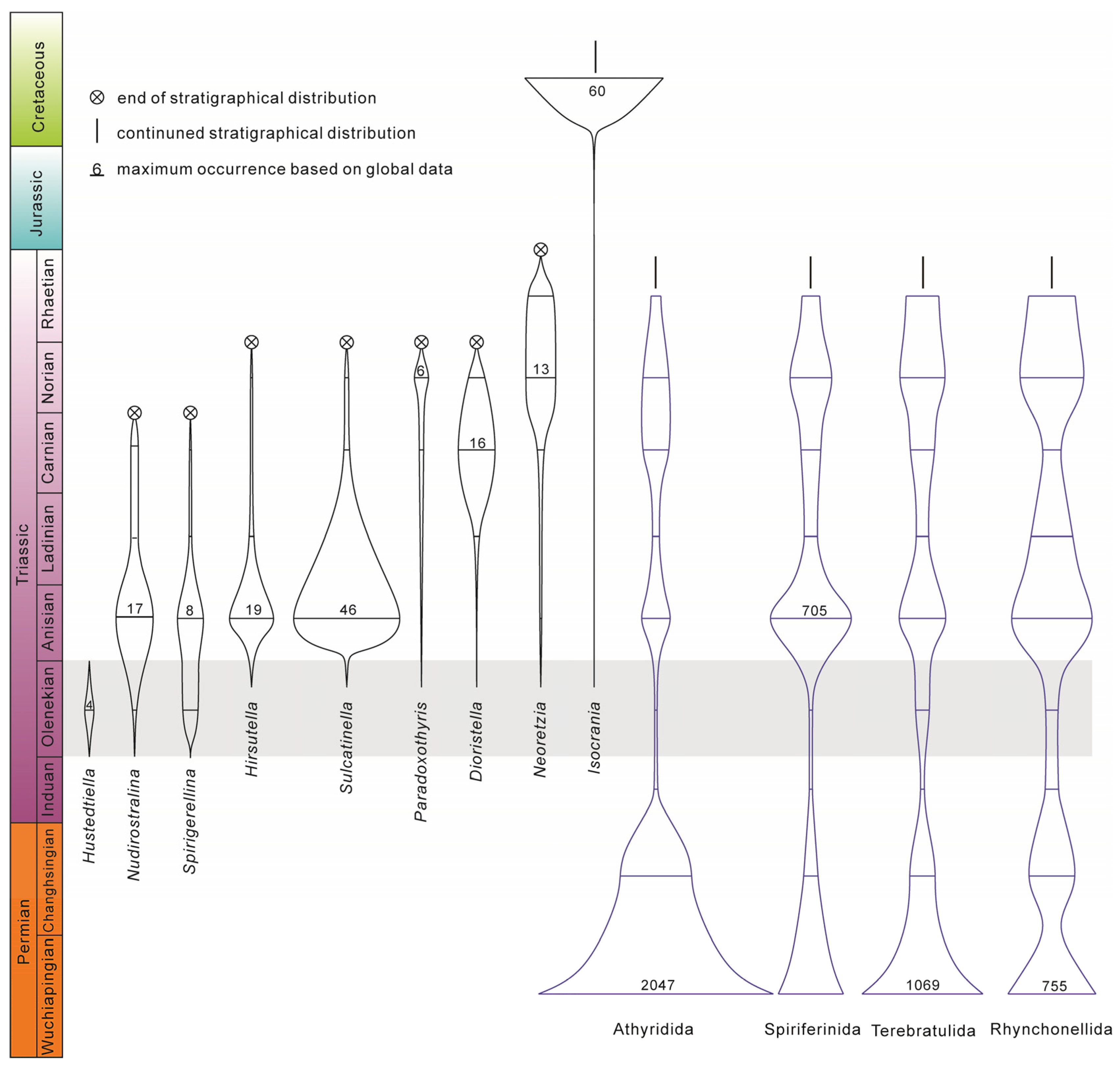
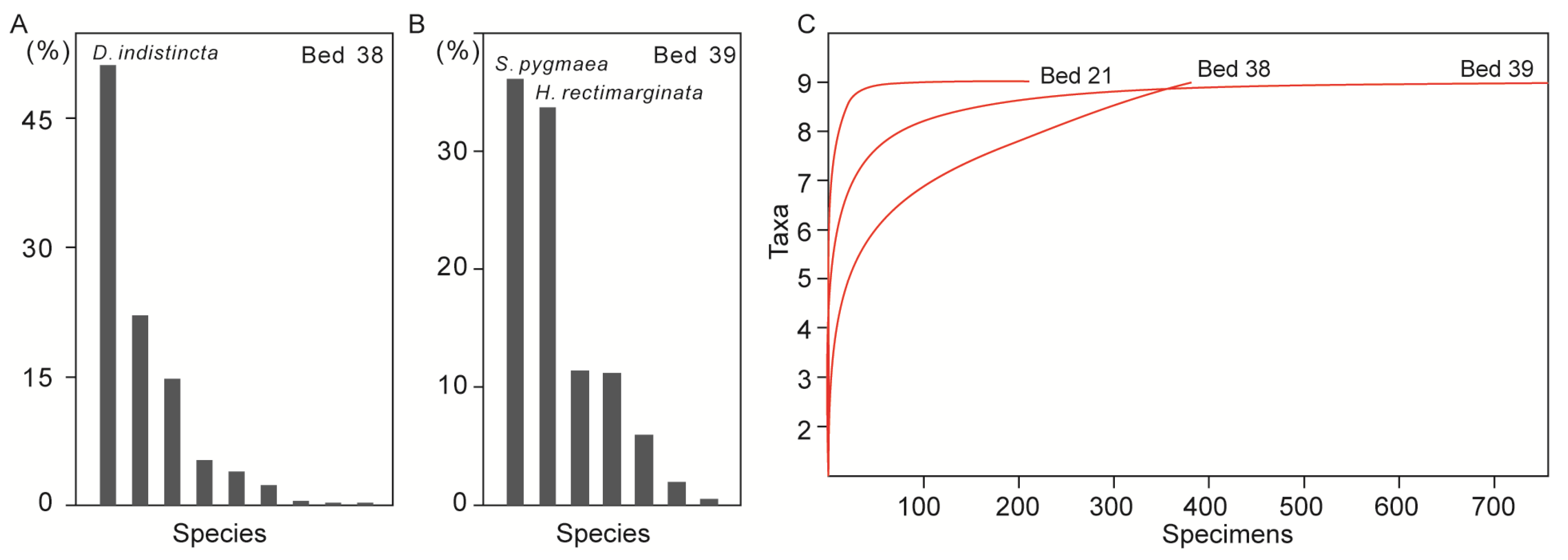
6.1. The Hidden Diversity in the Early Triassic
6.2. Brachiopod Recovery Pattern in the Post-Extinction Interval
7. Conclusions
- A Olenekian brachiopod fauna which is the most diverse one so far, is reported in this study. It contains 14 species in nine genera, among which Hirsutella, Sulcatinella, Paradoxothyris, Dioristella, Neoretzia and Isocrania are found in the Early Triassic for the first time, and three species are newly proposed;
- Brachiopod abundance and diversity data indicated that brachiopod recovery in the studied section happened in the latest Spathian when the environmental condition (deadly temperatures and anoxia) started to ameliorate;
- One of the reasons that brachiopod was widely considered to be in very low abundance in the Early Triassic was the phenomenon of hidden diversity. It could be caused by the decrease of habitat, low taxa abundance and great thickness of strata.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vermeij, G.J. The Mesozoic marine revolution: Evidence from snails, predators and grazers. Paleobiology 1977, 3, 245–258. [Google Scholar] [CrossRef]
- Gould, S.J.; Calloway, C.B. Clams and brachiopods—Ships that pass in the night. Paleobiology 1980, 6, 383–396. [Google Scholar] [CrossRef]
- Sepkoski, J.J. A factor analytic description of the Phanerozoic marine fossil record. Paleobiology 1981, 7, 36–53. [Google Scholar] [CrossRef]
- Raup, D.M.; Sepkoski, J.J., Jr. Mass extinctions in the marine fossil record. Science 1982, 215, 1501–1503. [Google Scholar] [CrossRef]
- Bambach, R.K.; Knoll, A.H.; Wang, S.C. Origination, extinction, and mass depletions of marine diversity. Paleobiology 2004, 30, 522–542. [Google Scholar] [CrossRef]
- Chen, Z.Q.; Benton, M.J. The timing and pattern of biotic recovery following the end-Permian mass extinction. Nat. Geosci. 2012, 5, 375–383. [Google Scholar] [CrossRef]
- Brayard, A.; Krumenacker, L.J.; Botting, J.P.; Jenks, J.F.; Bylund, K.G.; Fara, E.; Vennin, E.; Olivier, N.; Goudemand, N.; Saucede, T.; et al. Unexpected Early Triassic marine ecosystem and the rise of the modern evolutionary fauna. Sci. Adv. 2017, 3, e1602159. [Google Scholar] [CrossRef]
- Sun, Y.D.; Joachimski, M.M.; Wignall, P.B.; Yan, C.B.; Chen, Y.L.; Jiang, H.S.; Wang, L.N.; Lai, X.L. Lethally hot temperatures during the Early Triassic greenhouse. Science 2012, 338, 366–370. [Google Scholar] [CrossRef]
- Huang, Y.G.; Chen, Z.Q.; Wignall, P.B.; Zhao, L.S. Latest Permian to Middle Triassic redox condition variations in ramp settings, South China: Pyrite framboid evidence. GSA Bull. 2017, 129, 229–243. [Google Scholar] [CrossRef]
- Chen, J.; Chen, Z.Q.; Tong, J.N. Environmental determinants and ecologic selectivity of benthic faunas from nearshore to bathyal zones in the end-Permian mass extinction: Brachiopod evidence from South China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 308, 84–97. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, Y.G.; Rong, J.Y. General conception on the evolutional trends of Brachiopoda. Acta Palaeontol. Sin. 1982, 21, 28–36. (In Chinese) [Google Scholar]
- Carlson, S.J. The evolution of Brachiopoda. Annu. Rev. Earth Planet. Sci. 2016, 44, 409–438. [Google Scholar] [CrossRef]
- Chen, Z.Q.; Kaiho, K.; George, A.D. Survival strategies of brachiopod faunas from the end-Permian mass extinction. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 224, 232–269. [Google Scholar] [CrossRef]
- He, W.H.; Shi, G.R.; Yang, T.L.; Zhang, K.X.; Yue, M.L.; Xiao, Y.F.; Wu, H.T.; Chen, B.; Wu, S.B. Patterns of brachiopod faunal and body-size changes across the Permian–Triassic boundary: Evidence from the Daoduishan section in Meishan area, South China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 448, 72–84. [Google Scholar] [CrossRef]
- Wu, H.T.; He, W.H.; Weldon, E.A. Prelude of benthic community collapse during the end-Permian mass extinction in siliciclastic offshore sub-basin: Brachiopod evidence from South China. Glob. Planet. Chang. 2018, 163, 158–170. [Google Scholar] [CrossRef]
- Peng, Y.Q.; Shi, G.R. New Early Triassic Lingulidae (Brachiopoda) genera and species from South China. Alcheringa 2008, 32, 149–170. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, G.R.; Wu, H.T.; Yang, T.L.; He, W.H.; Yuan, A.Y.; Lei, Y. Community replacement, ecological shift and early warning signals prior to the end-Permian mass extinction: A case study from a nearshore clastic-shelf section in South China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 487, 118–135. [Google Scholar] [CrossRef]
- Chen, Z.Q.; Shi, G.R.; Kaiho, K. A new genus of rhynchonellid brachiopod from the Lower Triassic of South China and implications for timing the recovery of Brachiopoda after the end-Permian mass extinction. Palaeontology 2002, 45, 149–164. [Google Scholar] [CrossRef]
- Wang, F.Y.; Chen, J.; Dai, X.; Song, H.J. A new Dienerian (Early Triassic) brachiopod fauna from South China and implications for biotic recovery after the Permian–Triassic extinction. Pap. Palaeontol. 2017, 3, 425–439. [Google Scholar] [CrossRef]
- Girty, G.H. Descriptions of new species of Carboniferous and Triassic fossils (from southeastern Idaho). United States Geol. Surv. Prof. Pap. 1927, 152, 411–446. [Google Scholar]
- Perry, D.G.; Chatterton, B.D.E. Late Early Triassic brachiopod and conodont fauna, Thaynes Formation, southeastern Idaho. J. Paleontol. 1979, 53, 307–319. [Google Scholar]
- Yang, Z.Y.; Yin, H.F.; Xu, G.R.; Wu, S.B.; He, Y.L.; Liu, G.C.; Yin, J.R. Triassic of the South Qilian MTS; Geological Publishing House: Beijing, China, 1983; p. 224, (In Chinese with English Abstract). [Google Scholar]
- Popov, A.M.; Zakharov, Y.D. Olenekian brachiopods from the Kamenushka River Basin, South Primorye: New data on the brachiopod recovery after the end-Permian mass extinction. Paleontol. J. 2017, 51, 735–745. [Google Scholar] [CrossRef]
- Grădinaru, E.; Gaetani, M. Upper Spathian to Bithynian (Lower to Middle Triassic) brachiopods from North Dobrogea (Romania). Riv. Ital. Paleontol. Stratigr. 2019, 125, 91–123. [Google Scholar]
- Wang, F.Y.; Chen, J.; Dai, X.; Song, H.J. A new Early Triassic brachiopod fauna from southern Tibet, China: Implications on brachiopod recovery and the late Smithian extinction in southern Tethys. J. Paleontol. 2022, 96, 1–32. [Google Scholar] [CrossRef]
- Feng, Z.Z.; Yang, Y.Q.; Jin, Z.K.; Li, S.W.; Bao, Z.D. Lithofacies Paleogeography of Permian of South China; Petroleum University Press: Dongying, China, 1997; p. 242, (In Chinese with English Abstract). [Google Scholar]
- Zhang, L.J.; Buatois, L.A.; Mángano, M.G.; Qi, Y.A.; Tai, C. Early Triassic trace fossils from South China marginal-marine settings: Implications for biotic recovery following the end-Permian mass extinction. GSA Bull. 2019, 131, 235–251. [Google Scholar] [CrossRef]
- Yin, H.F.; Yochelson, E.L. Middle Triassic Gastropoda from Qingyan, Guizhou Province, China: 1. Pleurotomariacea and Murchisoniacea. J. Paleontol. 1983, 57, 162–187. [Google Scholar]
- Yin, H.F.; Yochelson, E.L. Middle Triassic Gastropoda from Qingyan, Guizhou Province, China: 2. Trochacea and Neritacea. J. Paleontol. 1983, 57, 515–538. [Google Scholar]
- Yin, H.F.; Yochelson, E.L. Middle Triassic Gastropoda from Qingyan, Guizhou Province, China: 3. Euomphalacea and Loxonematacea. J. Paleontol. 1983, 57, 1098–1127. [Google Scholar]
- Guo, Z.; Chen, Z.Q.; Harper, D.A.T. The Anisian (Middle Triassic) brachiopod fauna from Qingyan, Guizhou, south-western China. J. Syst. Palaeontol. 2020, 18, 647–701. [Google Scholar] [CrossRef]
- Brühwiler, T.; Brayard, A.; Bucher, H.; Kuang, G.D. Griesbachian and Dienerian (Early Triassic) ammonoid faunas from north-western Guangxi and southern Guizhou (South China). Palaeontology 2008, 51, 1151–1180. [Google Scholar] [CrossRef]
- Dai, X.; Song, H.J.; Brayard, A.; Ware, D. A new Griesbachian–Dienerian (Induan, Early Triassic) ammonoid fauna from Gujiao, South China. J. Paleontol. 2019, 93, 48–71. [Google Scholar] [CrossRef]
- Qiu, X.C.; Tian, L.; Wu, K.; Benton, M.J.; Sun, D.Y.; Yang, H.; Tong, J.N. Diverse earliest Triassic ostracod fauna of the non-microbialite-bearing shallow marine carbonate of the Yangou section, South China. Lethaia 2019, 52, 583–596. [Google Scholar] [CrossRef]
- Song, T.; Tong, J.N.; Tian, L.; Chu, D.L.; Huang, Y.F. Taxonomic and ecological variations of Permian–Triassic transitional bivalve communities from the littoral clastic facies in southwestern China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 519, 108–123. [Google Scholar] [CrossRef]
- Liu, X.K.; Song, H.J.; Bond, D.P.G.; Tong, J.N.; Benton, M.J. Migration controls extinction and survival patterns of foraminifers during the Permian–Triassic crisis in South China. Earth-Sci. Rev. 2020, 209, 103329. [Google Scholar] [CrossRef]
- Lehrmann, D.J.; Stepchinski, L.; Altiner, D.; Orchard, M.J.; Montgomery, P.; Enos, P.; Ellwood, B.B.; Bowring, S.A.; Ramezani, J.; Wang, H.M.; et al. An integrated biostratigraphy (conodont and foraminifers) and chronostratigraphy (paleomagnetic reversals, magnetic susceptibility, elemental chemistry, carbon isotopes and geochronology) for the Permian–Upper Triassic strata of Guandao section, Nanpanjiang Basin, South China. J. Asian Earth Sci. 2015, 108, 117–135. [Google Scholar]
- Chen, A.F.; Zhang, Y.; Golding, M.L.; Wu, H.T.; Liu, J.Q. Upper Changhsingian to lower Anisian conodont biostratigraphy of the Datuguan section, Nanpanjiang Basin, South China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2023, 616, 111470. [Google Scholar] [CrossRef]
- Hammer; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Kaesler, R.L. Treatise on Invertebrate Paleontology. Part H. Brachiopoda Vols 2 & 3. Linguliformea, Craniiforemea, & Rhynchonelliformea (Part); Geological Society of America & University of Kansas: Lawrence, KS, USA, 2000; p. 919. [Google Scholar]
- Kaesler, R.L. Treatise on Invertebrate Paleontology. Part H. Brachiopoda Vol. 4. Rhynchonelliformea (Part) (Revised); Geological Society of America & University of Kansas: Lawrence, KS, USA, 2002; pp. 921–1688. [Google Scholar]
- Kaesler, R.L. Treatise on Invertebrate Paleontology. Part H. Brachiopoda Vol. 5. Rhynchonelliformea (Part) (Revised); Geological Society of America & University of Kansas: Lawrence, KS, USA, 2006; pp. 1689–2320. [Google Scholar]
- Selden, P.A. Treatise on Invertebrate Paleontology. Part H. Brachiopoda Revised Vol. 6; Geological Society of America & University of Kansas: Lawrence, KS, USA, 2007; pp. 2321–3226. [Google Scholar]
- Ivanova, E.A. Osnovnye zakonomernosti evoliutsii spiriferid (Brachiopoda) (Main features of spiriferid evolution (Brachiopoda)). Paleontol. Zhurnal 1972, 3, 28–42. [Google Scholar]
- Carter, J.L.; Johnson, J.G.; Rémy, G.; Hou, H.F. A revised classification of the spiriferid brachiopods. Ann. Carnegie Mus. 1994, 63, 327–374. [Google Scholar] [CrossRef]
- Waagen, W.H. Salt Range fossils: Productus limestone fossils, Part 4 (1–5): Brachiopoda. Palaeontol. Indica 1883, 13, 329–770. [Google Scholar]
- Schuchert, C. Classification of brachiopod genera, fossil and recent. 10–15. In Fossilium Catalogus; Pompeckj, J.F., Ed.; W. Junk: Berlin, Germany, 1929; Volume 1. [Google Scholar]
- Cooper, G.A.; Muir-wood, H.M. Brachiopod homonyms. J. Wash. Acad. Sci. 1951, 41, 195–196. [Google Scholar]
- Yang, Z.Y.; Ding, P.Z.; Yin, H.F.; Zhang, S.X.; Fan, J.S. The brachiopod fauna of Carboniferous, Permian and Triassic in the Qilianshan region. In Monograph on the Geology of the Qilian Mountains; Institute of Geology and Palaeontology, Chinese Academy of Sciences, Institute of Geology, Chinese Academy of Sciences, Beijing College of Geology, Eds.; Science Press: Beijing, China, 1962; Volume 4, pp. 1–134. (In Chinese) [Google Scholar]
- von Alberti, F.A. Überblick Über die Trias, Mit Berücksichtigung Ihres Vorkommens in Den Alpen; J.G. Cottaschen Buchhandlung: Stuttgart, Germany, 1864; Volume 8, p. 7. [Google Scholar]
- Sun, D.L.; Shi, Q. Late Paleozoic and Triassic brachiopods from the east of the Qinghai-Xizang Plateau. In Stratigraphy and Paleontology of Western Sichuan and Eastern Xizang China Part 3; Regional Geological Survey Team of Sichuan Geological Bureau, Nanjing Institute of Geology and Paleontology, Eds.; Sichuan Science and Technology Press: Chengdu, China, 1985; pp. 186–212. (In Chinese) [Google Scholar]
- Yang, Z.Y.; Xu, G.R. Triassic Brachiopods of Central Guizhou Province, China; China Industry Publishing House: Beijing, China, 1966; p. 151, (In Chinese with English Summary). [Google Scholar]
- Schuchert, C. Class 2. Brachiopoda. Text-Book Palaeontol. 1913, 1, 355–420. [Google Scholar]
- DAGYS, A.S. Postembrional’noe razvitie brakhidiia pozdnepaleozoiskikh i rannemezozoiskikh Terebratulida. Akad. Nauk. SSSR Sib. Otd. Inst. Geol. I Geofiz. Tr. 1972, 112, 22–58. [Google Scholar]
- XU, Q.J. Brachiopoda, Mesozoic. 267–314. In Paleontological Atlas of Southwest China, Sichuan Province, Volume 2, Carboniferous–Mesozoic; Southwest Geological Science and Technology Research Institute, Ed.; Geological Publishing House: Beijing, China, 1978. (In Chinese) [Google Scholar]
- Jin, Y.G.; Ye, S.L.; Xu, H.K.; Sun, D.L. Brachiopoda. 60–217. In Paleontological Atlas of North-Western China, Qinghai Province; Institute of Geological Science of Qinghai Province and Nanjing Institute of Geology and Palaeonotology, Chinese Academy of Sciences, Eds.; Geological Publishing House: Beijing, China, 1979; Volume 1. (In Chinese) [Google Scholar]
- Dagys, A.S. Triasovye Brakhiopody (Morfologiia, Sistema, Filogeniia, Stratigraficheskoe Znachenie I Biogeografiia) [Triassic Brachiopods (Morphology, Classification, Phylogeny, Stratigraphical Significance and Biogeography)]; Sibirskoe Otdelenie Izdatel’stvo “Nauka.”: Novosibirsk, Russia, 1974; Volume 214, pp. 1–49. [Google Scholar]
- Pietsch, C.; Ritterbush, K.A.; Thompson, J.R.; Petsios, E.; Bottjer, D.J. Evolutionary models in the Early Triassic marine realm. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 513, 65–85. [Google Scholar] [CrossRef]
- Wignall, P.B.; Bond, D.P.G.; Sun, Y.D.; Grasby, S.E.; Beauchamp, B.; Joachimski, M.M.; Blomeier, D.P.G. Ultra-shallow-marine anoxia in an Early Triassic shallow-marine clastic ramp (Spitsbergen) and the suppression of benthic radiation. Geol. Mag. 2016, 153, 316–331. [Google Scholar] [CrossRef]
- Chen, J.; Shen, S.Z.; Zhang, Y.C.; Angiolini, L.; Gorgij, M.N.; Crippa, G.; Wang, W.; Zhang, H.; Yuan, D.X.; Li, X.H.; et al. Abrupt warming in the latest Permian detected using high-resolution in situ oxygen isotopes of conodont apatite from Abadeh, central Iran. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 560, 109973. [Google Scholar] [CrossRef]
- Beatty, T.W.; Zonneveld, J.P.; Henderson, C.M. Anomalously diverse Early Triassic ichnofossil assemblages in northwest Pangaea: A case for a shallow-marine habitable zone. Geology 2008, 36, 771–774. [Google Scholar] [CrossRef]
- Song, H.J.; Wignall, P.B.; Chu, D.L.; Tong, J.N.; Sun, Y.D.; Song, H.Y.; He, W.H.; Tian, L. Anoxia/high temperature double whammy during the Permian–Triassic marine crisis and its aftermath. Sci. Rep. 2014, 4, 4231. [Google Scholar] [CrossRef]
- Foster, W.J.; Danise, S.; Sedlacek, A.; Price, G.D.; Hips, K.; Twitchett, R.J. Environmental controls on the post-Permian recovery of benthic, tropical marine ecosystems in western Palaeotethys (Aggtelek Karst, Hungary). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 440, 374–394. [Google Scholar] [CrossRef]
- He, W.H.; Zhang, K.X.; Chen, Z.Q.; Yan, J.X.; Yang, T.L.; Zhang, Y.; Gu, S.Z.; Wu, S.B. A new genus Liaous of early Anisian Stage (Middle Triassic) brachiopods from southwestern China: Systematics, reassessment of classification of the Spiriferinioidea, community paleoecology, and paleoenvironmental implications. J. Paleontol. 2015, 89, 966–979. [Google Scholar] [CrossRef]
- Hautmann, M.; Bagherpour, B.; Brosse, M.; Frisk, A.; Hofmann, R.; Baud, A.; Nützel, A.; Goudemand, N.; Bucher, H. Competition in slow motion: The unusual case of benthic marine communities in the wake of the end-Permian mass extinction. Palaeontology 2015, 58, 871–901. [Google Scholar] [CrossRef]
- Feng, X.Q.; Chen, Z.Q.; Benton, M.J.; Su, C.M.; Bottjer, D.J.; Cribb, A.T.; Li, Z.H.; Zhao, L.S.; Zhu, G.Y.; Huang, Y.G.; et al. Resilience of infaunal ecosystems during the Early Triassic greenhouse Earth. Sci. Adv. 2022, 8, eabo0597. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.T.; Zhang, Y.; Sun, Y.L. A brachiopod fauna from latest Permian to Induan of northern Guizhou, South China and its evolutionary pattern. Geol. J. 2021, 56, 6189–6198. [Google Scholar] [CrossRef]
- Hautmann, M.; Smith, A.B.; Mcgowan, A.J.; Bucher, H. Bivalves from the Olenekian (Early Triassic) of south-western Utah: Systematics and evolutionary significance. J. Syst. Palaeontol. 2013, 11, 263–293. [Google Scholar] [CrossRef]
- Foster, W.J.; Lehrmann, D.J.; Yu, M.; Ji, L.; Martindale, R.C. Persistent environmental stress delayed the recovery of marine communities in the aftermath of the latest Permian mass extinction. Paleoceanogr. Paleoclimatol. 2018, 33, 338–353. [Google Scholar] [CrossRef]
- Woods, A.D.; Alms, P.D.; Monarrez, P.M.; Mata, S. The interaction of recovery and environmental conditions: An analysis of the outer shelf edge of western North America during the Early Triassic. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 513, 52–64. [Google Scholar] [CrossRef]
- Kozur, H.; Krainer, K.; Mostler, H. Neospathodus sosioensis n. sp.; a new conodont species from the late Olenekian (uppermost Scythian) of western Sicily, Italy. In Proceedings of the Fifth Radiolarian Symposium; Special Volume; News of Osaka Micropaleontologists: Osaka, Japan, 1997; Volume 10, pp. 109–113. [Google Scholar]
- Henderson, C.M.; Golding, M.L.; Orchard, M.J. Conodont sequence biostratigraphy of the Lower Triassic Montney Formation. Bull. Can. Pet. Geol. 2018, 66, 7–22. [Google Scholar]
- Sun, Y.D.; Richoz, S.; Krystyn, L.; Grasby, S.E.; Chen, Y.L.; Banerjee, D.; Joachimski, M.M. Integrated bio-chemostratigraphy of Lower and Middle Triassic marine successions at Spiti in the Induan Himalaya: Implications for the Early Triassic nutrient crisis. Glob. Planet. Chang. 2021, 196, 103363. [Google Scholar] [CrossRef]
- Pietsch, C.; Petsios, E.; Bottjer, D.J. Sudden and extreme hyperthermals, low-oxygen, and sediment influx drove community phase shifts following the end-Permian mass extinction. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 451, 183–196. [Google Scholar] [CrossRef]
- Takahashi, S.; Hori, R.S.; Yamakita, S.; Aita, Y.; Takemura, A.; Ikehara, M.; Xiong, Y.J.; Poulton, S.W.; Wignall, P.B.; Itai, T.; et al. Progressive development of ocean anoxia in the end-Permian pelagic Panthalassa. Glob. Planet. Chang. 2021, 207, 103650. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Genus/ Species | Order | Age | Reference |
|---|---|---|---|---|
| Primorye, Russia | 6/6 | Rhynchonellida, Terebratulida, Spiriferinida, Athyridida | Olenekian | [23] |
| Qilian Area, China | 4/10 | Rhynchonellida, Terebratulida, Athyridida | Olenekian | [22] |
| Tibet, China | 3/3 | Rhynchonellida, Terebratulida, Athyridida | Smithian | [25] |
| Idaho, USA | 4/5 | Rhynchonellida, Terebratulida, Spiriferinida | Spathian | [20,21] |
| Dobrogea, Romania | 3/3 | Rhynchonellida | Spathian | [24] |
| Guizhou, China | 9/14 | Rhynchonellida, Terebratulida, Spiriferinida, Athyridida, Craniida | Olenekian | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Zhang, Y.; Chen, A.; Stubbs, T.L. A Highly Diverse Olenekian Brachiopod Fauna from the Nanpanjiang Basin, South China, and Its Implications for the Early Triassic Biotic Recovery. Biology 2023, 12, 622. https://doi.org/10.3390/biology12040622
Wu H, Zhang Y, Chen A, Stubbs TL. A Highly Diverse Olenekian Brachiopod Fauna from the Nanpanjiang Basin, South China, and Its Implications for the Early Triassic Biotic Recovery. Biology. 2023; 12(4):622. https://doi.org/10.3390/biology12040622
Chicago/Turabian StyleWu, Huiting, Yang Zhang, Anfeng Chen, and Thomas L. Stubbs. 2023. "A Highly Diverse Olenekian Brachiopod Fauna from the Nanpanjiang Basin, South China, and Its Implications for the Early Triassic Biotic Recovery" Biology 12, no. 4: 622. https://doi.org/10.3390/biology12040622