
Abnormal Characterization and Distribution of Circulating Regulatory T Cells in Patients with Chronic Spinal Cord Injury According to the Period of Evolution
 , , , , , , and
, , , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Patients and Methods
2.1. Study Design
2.2. Sample Collection and Inclusion/Exclusion Criteria
2.3. Isolation of Peripheral Blood Mononuclear Cells and Immunophenotype Studies
2.4. Statistical Analysis
3. Results
3.1. Patients Demographics
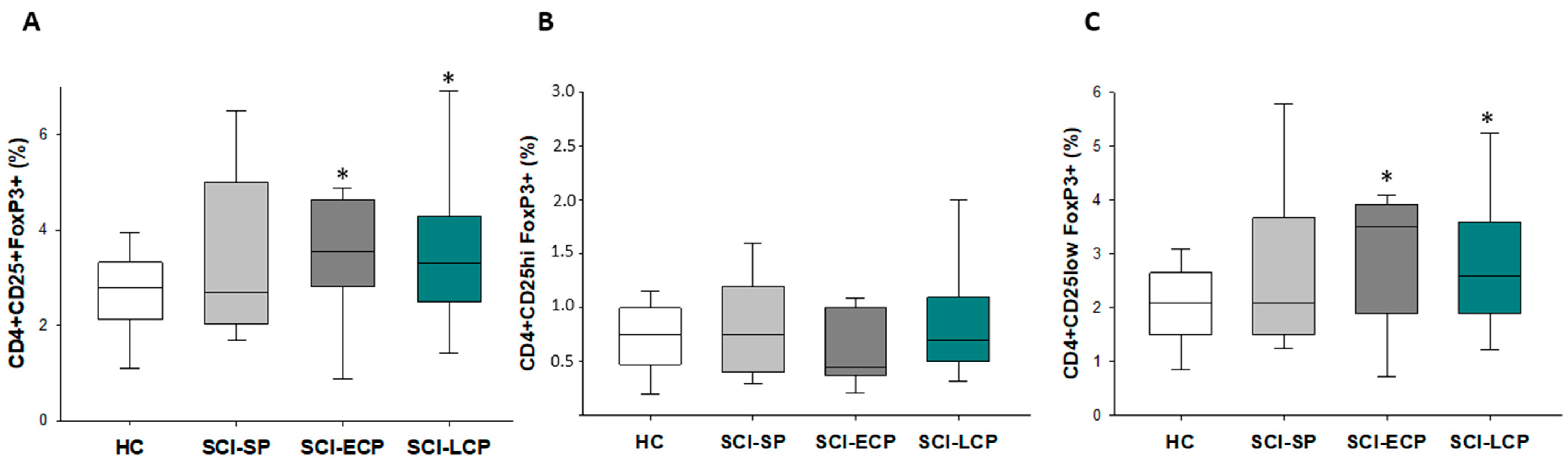
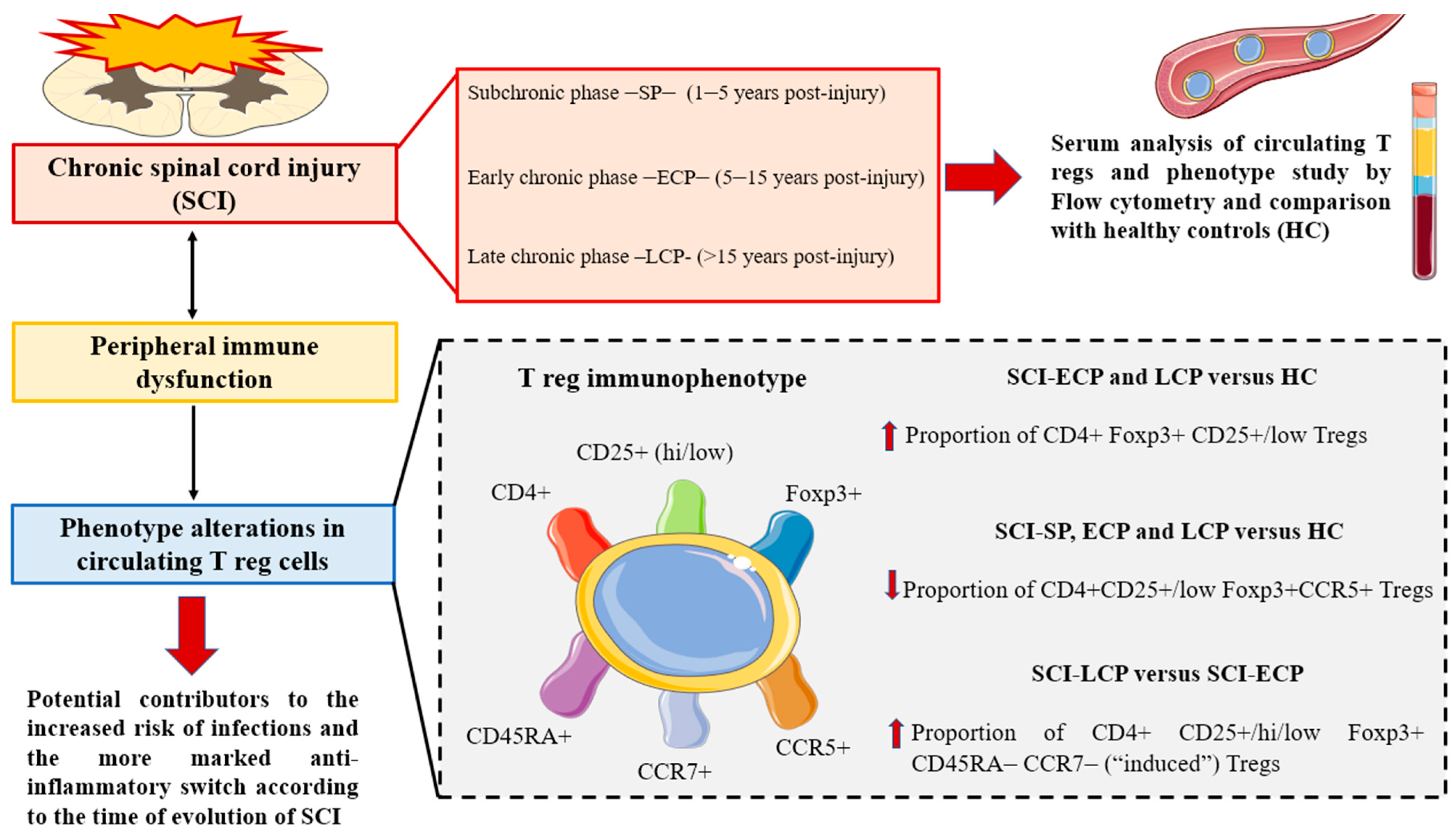
3.2. Patients with a Long Period of Chronic Spinal Cord Injury Present Significant Differences in Circulating CD4+ Foxp3+ CD25+/CD25low Regulatory T Cells Populations
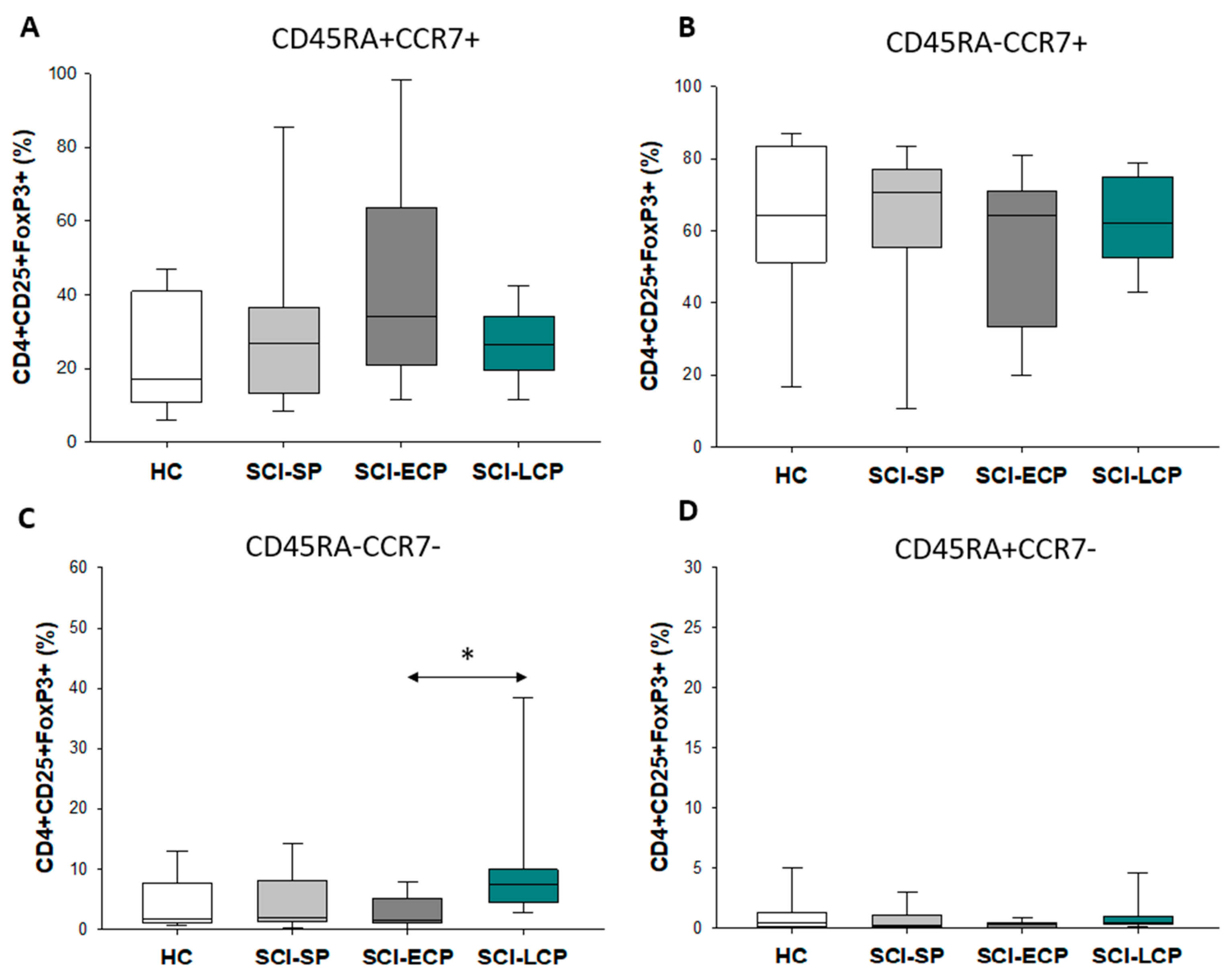
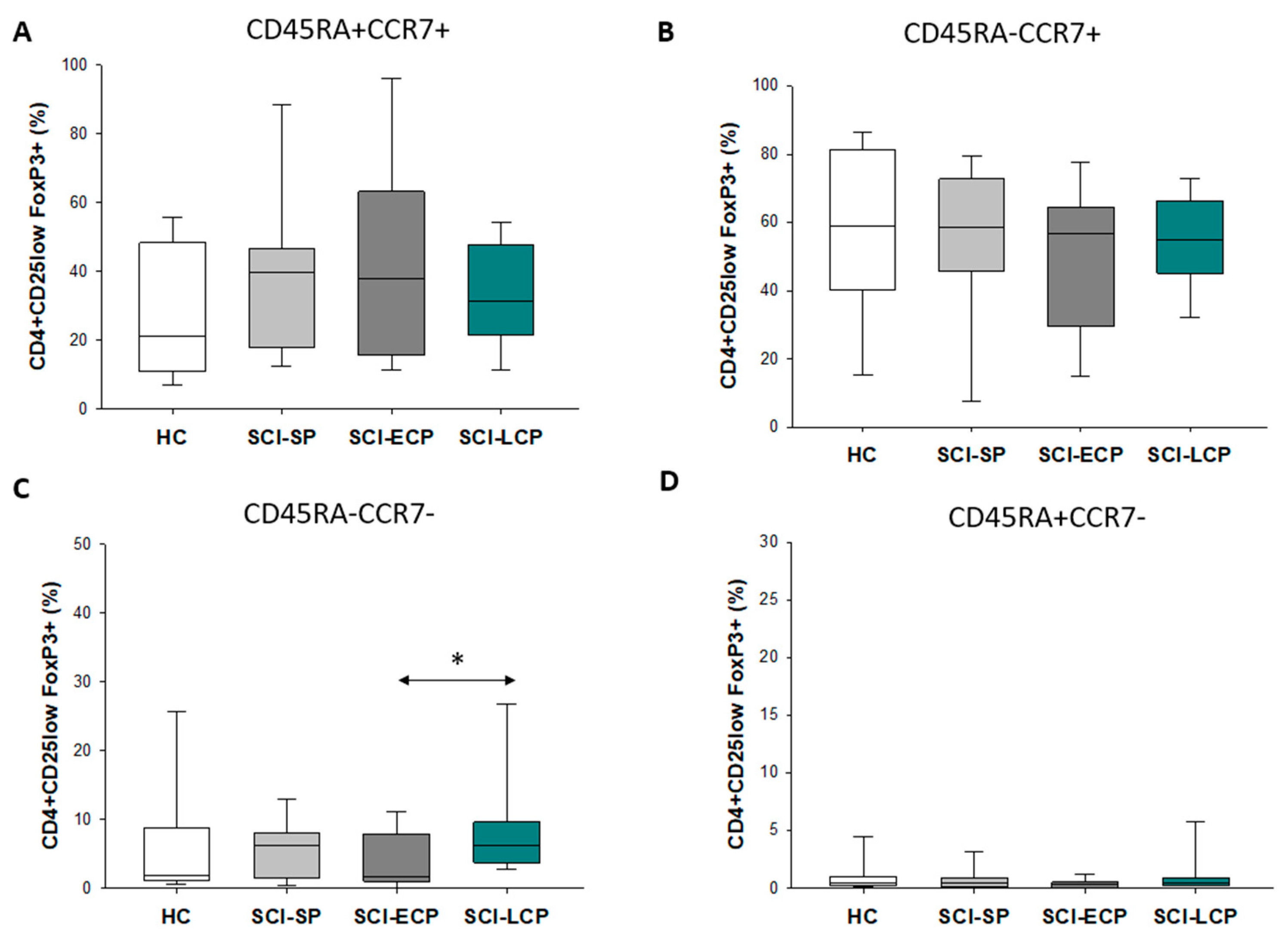
3.3. Patients with a Long Duration of Chronic Spinal Cord Injury Present a Significant Increase in Circulating CD4+ CD25+ Foxp3+ Regulatory T Cell Populations Negative for CD45RA and CCR7 Expression
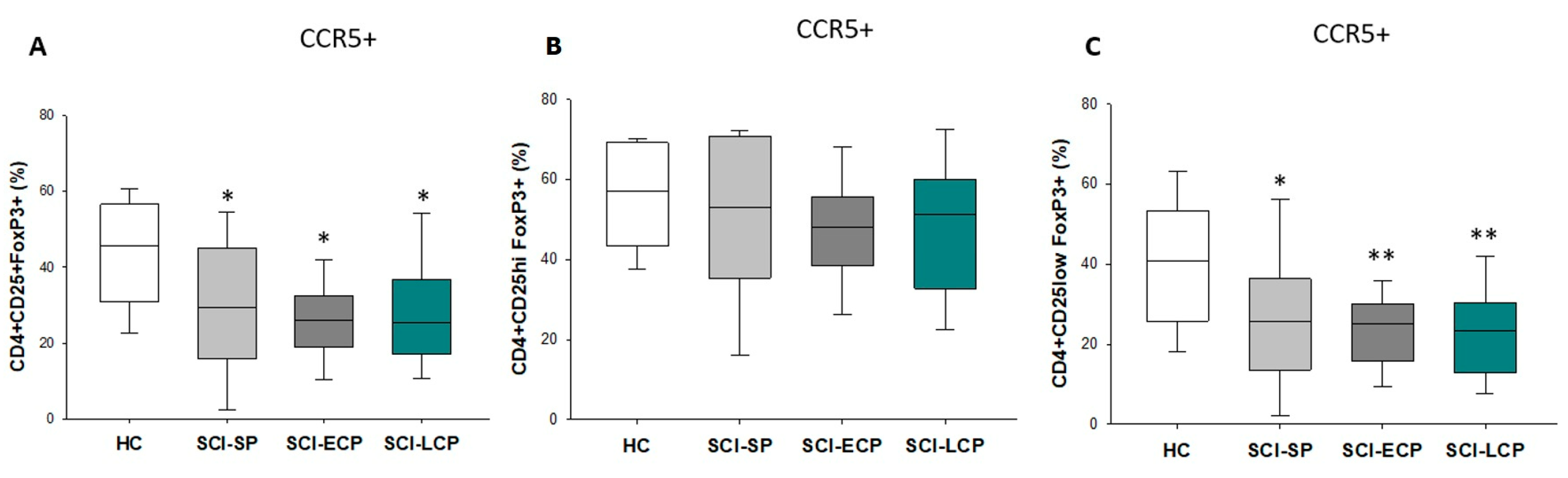
3.4. Patients with a Long Period of Chronic Spinal Cord Injury Present Statistically Significant Differences in Circulating CD4+ CD25+ Foxp3+ Regulatory T Cells Populations Positive for CCR5 Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ali, Z.S.; Whitmore, R.G. Spinal Cord Injuries. In Surgical Intensive Care Medicine, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 181–193. [Google Scholar] [CrossRef]
- Silva, N.A.; Sousa, N.; Reis, R.L.; Salgado, A.J. From Basics to Clinical: A Comprehensive Review on Spinal Cord Injury. Prog. Neurobiol. 2014, 114, 25–57. [Google Scholar] [CrossRef] [PubMed]
- Varma, A.K.; Das, A.; Wallace, G.; Barry, J.; Vertegel, A.A.; Ray, S.K.; Banik, N.L. Spinal Cord Injury: A Review of Current Therapy, Future Treatments, and Basic Science Frontiers. Neurochem. Res. 2013, 38, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Jazayeri, S.B.; Beygi, S.; Shokraneh, F.; Hagen, E.M.; Rahimi-Movaghar, V. Incidence of Traumatic Spinal Cord Injury Worldwide: A Systematic Review. Eur. Spine J. 2015, 24, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Oyinbo, C.A. Secondary Injury Mechanisms in Traumatic Spinal Cord Injury: A Nugget of This Multiply Cascade. Acta Neurobiol. Exp. 2011, 71, 281–299. [Google Scholar]
- Sun, X.; Jones, Z.B.; Chen, X.-M.; Zhou, L.; So, K.-F.; Ren, Y. Multiple Organ Dysfunction and Systemic Inflammation after Spinal Cord Injury: A Complex Relationship. J. Neuroinflamm. 2016, 13, 1–11. [Google Scholar] [CrossRef]
- Jeffries, M.A.; Tom, V.J. Peripheral Immune Dysfunction: A Problem of Central Importance after Spinal Cord Injury. Biology 2021, 10, 928. [Google Scholar] [CrossRef]
- Kasinathan, N.; Vanathi, M.; Subrahmanyam, V.; Rao, J. A Review on Response of Immune System in Spinal Cord Injury and Therapeutic Agents Useful in Treatment. Curr. Pharm. Biotechnol. 2015, 16, 26–34. [Google Scholar] [CrossRef]
- Schwab, J.M.; Zhang, Y.; Kopp, M.A.; Brommer, B.; Popovich, P.G. The Paradox of Chronic Neuroinflammation, Systemic Immune Suppression and Autoimmunity after Traumatic Chronic Spinal Cord Injury. Exp. Neurol. 2014, 258, 121–129. [Google Scholar] [CrossRef]
- Carpenter, R.S.; Marbourg, J.M.; Brennan, F.H.; Mifflin, K.A.; Hall, J.C.E.; Jiang, R.R.; Mo, X.M.; Karunasiri, M.; Burke, M.H.; Dorrance, A.M.; et al. Spinal Cord Injury Causes Chronic Bone Marrow Failure. Nat. Commun. 2020, 11, 3702. [Google Scholar] [CrossRef]
- Brommer, B.; Engel, O.; Kopp, M.A.; Watzlawick, R.; Müller, S.; Prüss, H.; Chen, Y.; DeVivo, M.J.; Finkenstaedt, F.W.; Dirnagl, U.; et al. Spinal Cord Injury-Induced Immune Deficiency Syndrome Enhances Infection Susceptibility Dependent on Lesion Level. Brain 2016, 139, 692–707. [Google Scholar] [CrossRef]
- Bigford, G.E.; Garshick, E. Systemic Inflammation after Spinal Cord Injury: A Review of Biological Evidence, Related Health Risks, and Potential Therapies. Curr. Opin. Pharmacol. 2022, 67, 102303. [Google Scholar] [CrossRef] [PubMed]
- Pavlicek, D.; Krebs, J.; Capossela, S.; Bertolo, A.; Engelhardt, B.; Pannek, J.; Stoyanov, J. Immunosenescence in Persons with Spinal Cord Injury in Relation to Urinary Tract Infections—A Cross-Sectional Study. Immun. Ageing 2017, 14, 22. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.R.N.; Druschel, C.; Curt, A.; Fehlings, M.G. Traumatic Spinal Cord Injury. Nat. Rev. Dis. Prim. 2017, 3, 17018. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, D.D.; Hoffman, J.M.; Kirshblum, S.; McKinley, W. Etiology and Incidence of Rehospitalization after Traumatic Spinal Cord Injury: A Multicenter Analysis. Arch. Phys. Med. Rehabil. 2004, 85, 1757–1763. [Google Scholar] [CrossRef]
- Huang, W.; Vodovotz, Y.; Kusturiss, M.B.; Barclay, D.; Greenwald, K.; Boninger, M.L.; Coen, P.M.; Brienza, D.; Sowa, G. Identification of Distinct Monocyte Phenotypes and Correlation with Circulating Cytokine Profiles in Acute Response to Spinal Cord Injury: A Pilot Study. PM R 2014, 6, 332. [Google Scholar] [CrossRef]
- Gao, T.Y.; Huang, F.F.; Xie, Y.Y.; Wang, W.Q.; Wang, L.D.; Mu, D.; Cui, Y.; Wang, B. Dynamic Changes in the Systemic Immune Responses of Spinal Cord Injury Model Mice. Neural Regen. Res. 2021, 16, 382–387. [Google Scholar] [CrossRef]
- Diaz, D.; Lopez-Dolado, E.; Haro, S.; Monserrat, J.; Martinez-Alonso, C.; Balomeros, D.; Albillos, A.; Alvarez-Mon, M. Systemic Inflammation and the Breakdown of Intestinal Homeostasis Are Key Events in Chronic Spinal Cord Injury Patients. Int. J. Mol. Sci. 2021, 22, 744. [Google Scholar] [CrossRef]
- Sauls, R.S.; McCausland, C.; Taylor, B.N. Histology, T-Cell Lymphocyte. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Carpenter, R.S.; Jiang, R.R.; Brennan, F.H.; Hall, J.C.E.; Gottipati, M.K.; Niewiesk, S.; Popovich, P.G. Human Immune Cells Infiltrate the Spinal Cord and Impair Recovery after Spinal Cord Injury in Humanized Mice. Sci. Rep. 2019, 9, 1910. [Google Scholar] [CrossRef]
- Vignali, D.A.A.; Collison, L.W.; Workman, C.J. How Regulatory T Cells Work. Nat. Rev. Immunol. 2008, 8, 523–532. [Google Scholar] [CrossRef]
- Staats, J. Immunophenotyping of Human Regulatory T Cells. Methods Mol. Biol. 2019, 2032, 141–177. [Google Scholar] [CrossRef]
- Hornero, R.A.; Betts, G.J.; Sawitzki, B.; Vogt, K.; Harden, P.N.; Wood, K.J. CD45RA Distinguishes CD4+ CD25+ CD127−/Low TSDR Demethylated Regulatory T Cell Subpopulations With Differential Stability and Susceptibility to Tacrolimus-Mediated Inhibition of Suppression. Transplantation 2017, 101, 302. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.A.; Meingassner, J.G.; Lipp, M.; Moore, H.D.; Rot, A. CCR7 Is Required for the in Vivo Function of CD4+ CD25+ Regulatory T Cells. J. Exp. Med. 2007, 204, 735–745. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, C.E.; Gasparoto, T.H.; Pinheiro, C.R.; Amôr, N.G.; Nogueira, M.R.S.; Kaneno, R.; Garlet, G.P.; Lara, V.S.; Silva, J.S.; Cavassani, K.A.; et al. CCR5-Dependent Homing of T Regulatory Cells to the Tumor Microenvironment Contributes to Skin Squamous Cell Carcinoma Development. Mol. Cancer Ther. 2017, 16, 2871–2880. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Wang, N.; Vasanthakumar, A.; Zhang, Y.; Chopin, M.; Nutt, S.L.; Kallies, A.; Lew, A.M. CCR2 Enhances CD25 Expression by FoxP3+ Regulatory T Cells and Regulates Their Abundance Independently of Chemotaxis and CCR2+ Myeloid Cells. Cell. Mol. Immunol. 2020, 17, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.X.; Beck, K.D.; Anderson, A.J. Quantitative Assessment of Immune Cells in the Injured Spinal Cord Tissue by Flow Cytometry: A Novel Use for a Cell Purification Method. J. Vis. Exp. 2011, 50, e2698. [Google Scholar] [CrossRef]
- Raposo, C.; Graubardt, N.; Cohen, M.; Eitan, C.; London, A.; Berkutzki, T.; Schwartz, M. CNS Repair Requires Both Effector and Regulatory T Cells with Distinct Temporal and Spatial Profiles. J. Neurosci. 2014, 34, 10141–10155. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xiong, W.; Wan, B.; Kong, G.; Wang, S.; Wang, Y.; Fan, J. Role of Peripheral Immune Cells in Spinal Cord Injury. Cell Mol. Life Sci. 2023, 80, 2. [Google Scholar] [CrossRef]
- Walsh, J.T.; Zheng, J.; Smirnov, I.; Lorenz, U.; Tung, K.; Kipnis, J. Regulatory T Cells in Central Nervous System Injury: A Double-Edged Sword. J. Immunol. 2014, 193, 5013. [Google Scholar] [CrossRef]
- Monahan, R.; Stein, A.; Gibbs, K.; Bank, M.; Bloom, O. Circulating T Cell Subsets Are Altered in Individuals with Chronic Spinal Cord Injury. Immunol. Res. 2015, 63, 3–10. [Google Scholar] [CrossRef]
- Fraussen, J.; Beckers, L.; van Laake-Geelen, C.C.M.; Depreitere, B.; Deckers, J.; Cornips, E.M.J.; Peuskens, D.; Somers, V. Altered Circulating Immune Cell Distribution in Traumatic Spinal Cord Injury Patients in Relation to Clinical Parameters. Front. Immunol. 2022, 13, 3296. [Google Scholar] [CrossRef]
- Roberts, T.T.; Leonard, G.R.; Cepela, D.J. Classifications In Brief: American Spinal Injury Association (ASIA) Impairment Scale. Clin. Orthop. Relat. Res. 2017, 475, 1499. [Google Scholar] [CrossRef] [PubMed]
- Bauman, W.A.; Wecht, J.M.; Biering-Sorensen, F. International Spinal Cord Injury Endocrine and Metabolic Extended Data Set. Spinal Cord 2017, 55, 466–477. [Google Scholar] [CrossRef]
- Biering-Sørensen, F.; Charlifue, S.; DeVivo, M.; Noonan, V.; Post, M.; Stripling, T.; Wing, P. International Spinal Cord Injury Data Sets. Spinal Cord 2006, 44, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Mon, M.A.; Gomez-Lahoz, A.M.; Orozco, A.; Lahera, G.; Sosa-Reina, M.D.; Diaz, D.; Albillos, A.; Quintero, J.; Molero, P.; Monserrat, J.; et al. Blunted Expansion of Regulatory T Lymphocytes Is Associated With Increased Bacterial Translocation in Patients With Major Depressive Disorder. Front. Psychiatry 2021, 11, 1530. [Google Scholar] [CrossRef]
- Tang, X.; Li, Q.; Huang, T.; Zhang, H.; Chen, X.; Ling, J.; Yang, Y. Regenerative Role of T Cells in Nerve Repair and Functional Recovery. Front. Immunol. 2022, 13, 3389. [Google Scholar] [CrossRef]
- Xu, L.; Ye, X.; Wang, Q.; Xu, B.; Zhong, J.; Chen, Y.; Wang, L. T-cell Infiltration, Contribution and Regulation in the Central Nervous System Post-traumatic Injury. Cell Prolif. 2021, 54, e13092. [Google Scholar] [CrossRef]
- Walsh, J.T.; Kipnis, J. Regulatory T Cells in CNS Injury: The Simple, the Complex, and the Confused. Trends Mol. Med. 2011, 17, 541. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, P.; Eder, R.; Boeld, T.J.; Doser, K.; Piseshka, B.; Andreesen, R.; Edinger, M. Only the CD45RA+ Subpopulation of CD4+CD25high T Cells Gives Rise to Homogeneous Regulatory T-Cell Lines upon in Vitro Expansion. Blood 2006, 108, 4260–4267. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wu, C.; Xu, G.; Liu, Y.; Chen, J.; Zhang, J.; Hong, H.; Ji, C.; Cui, Z. CCR7-Mediated T Follicular Helper Cell Differentiation Is Associated with the Pathogenesis and Immune Microenvironment of Spinal Cord Injury-Induced Immune Deficiency Syndrome. Front. Neurosci. 2022, 16, 1019406. [Google Scholar] [CrossRef]
- Li, F.; Cheng, B.; Cheng, J.; Wang, D.; Li, H.; He, X. CCR5 Blockade Promotes M2 Macrophage Activation and Improves Locomotor Recovery after Spinal Cord Injury in Mice. Inflammation 2015, 38, 126–133. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Lahoz, A.M.; Girón, S.H.; Sanz, J.M.; Fraile-Martínez, O.; Garcia-Montero, C.; Jiménez, D.J.; de Leon-Oliva, D.; Ortega, M.A.; Atienza-Perez, M.; Diaz, D.; et al. Abnormal Characterization and Distribution of Circulating Regulatory T Cells in Patients with Chronic Spinal Cord Injury According to the Period of Evolution. Biology 2023, 12, 617. https://doi.org/10.3390/biology12040617
Gómez-Lahoz AM, Girón SH, Sanz JM, Fraile-Martínez O, Garcia-Montero C, Jiménez DJ, de Leon-Oliva D, Ortega MA, Atienza-Perez M, Diaz D, et al. Abnormal Characterization and Distribution of Circulating Regulatory T Cells in Patients with Chronic Spinal Cord Injury According to the Period of Evolution. Biology. 2023; 12(4):617. https://doi.org/10.3390/biology12040617
Chicago/Turabian StyleGómez-Lahoz, Ana M., Sergio Haro Girón, Jorge Monserrat Sanz, Oscar Fraile-Martínez, Cielo Garcia-Montero, Diego J. Jiménez, Diego de Leon-Oliva, Miguel A. Ortega, Mar Atienza-Perez, David Diaz, and et al. 2023. "Abnormal Characterization and Distribution of Circulating Regulatory T Cells in Patients with Chronic Spinal Cord Injury According to the Period of Evolution" Biology 12, no. 4: 617. https://doi.org/10.3390/biology12040617