
A Study on REM Sleep Homeostasis in the Day-Active Tree Shrew (Tupaia belangeri): Cold-Induced Suppression of REM Sleep Is Not Followed by a Rebound
 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Surgery
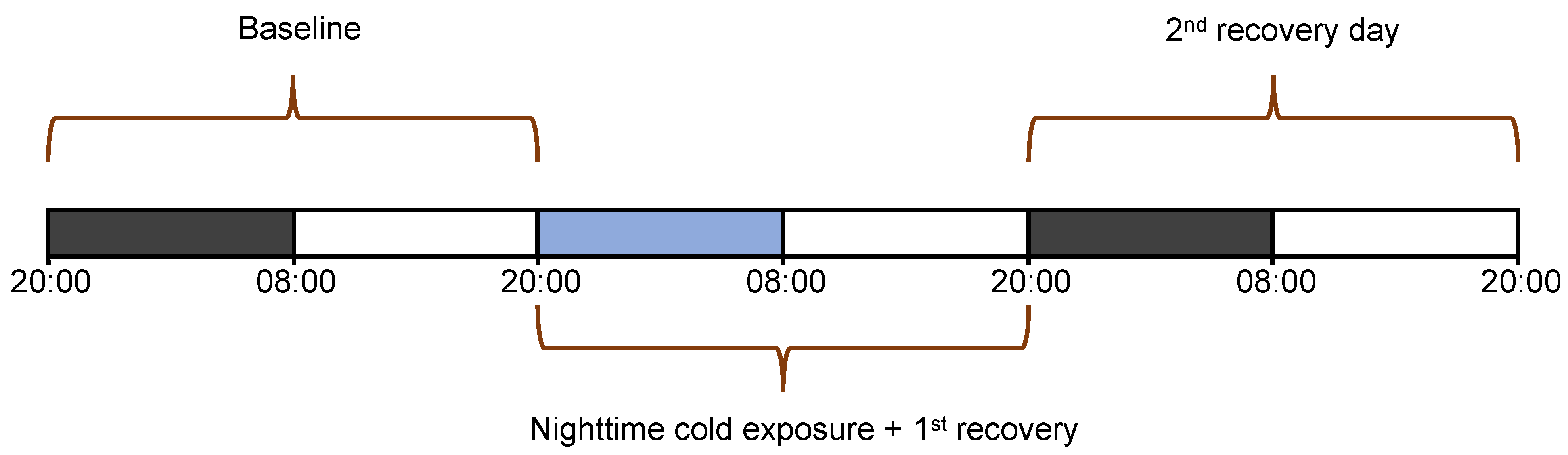
2.3. Experimental Design
2.4. Signal and Data Analyses
2.5. Statistics
3. Results
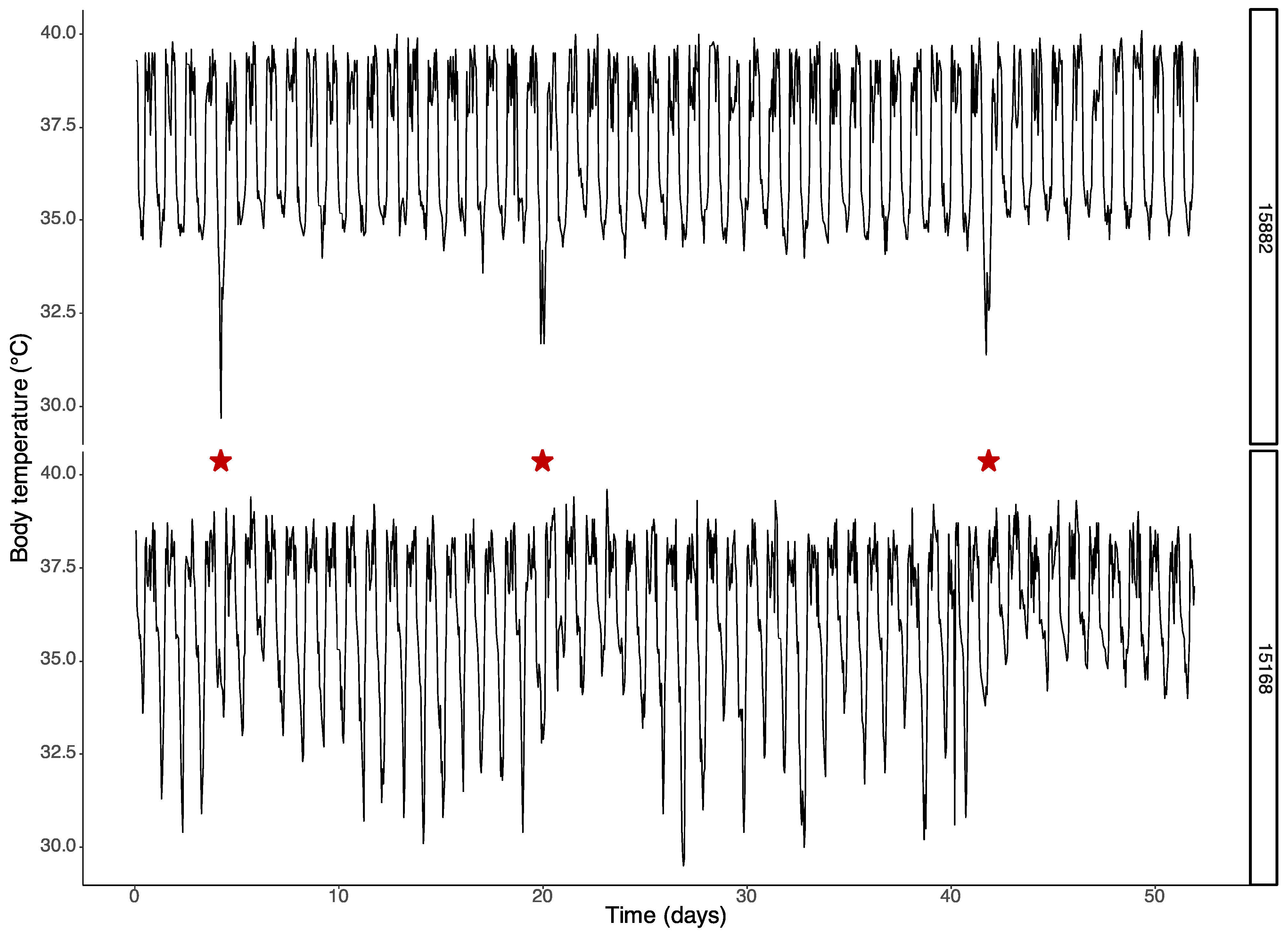
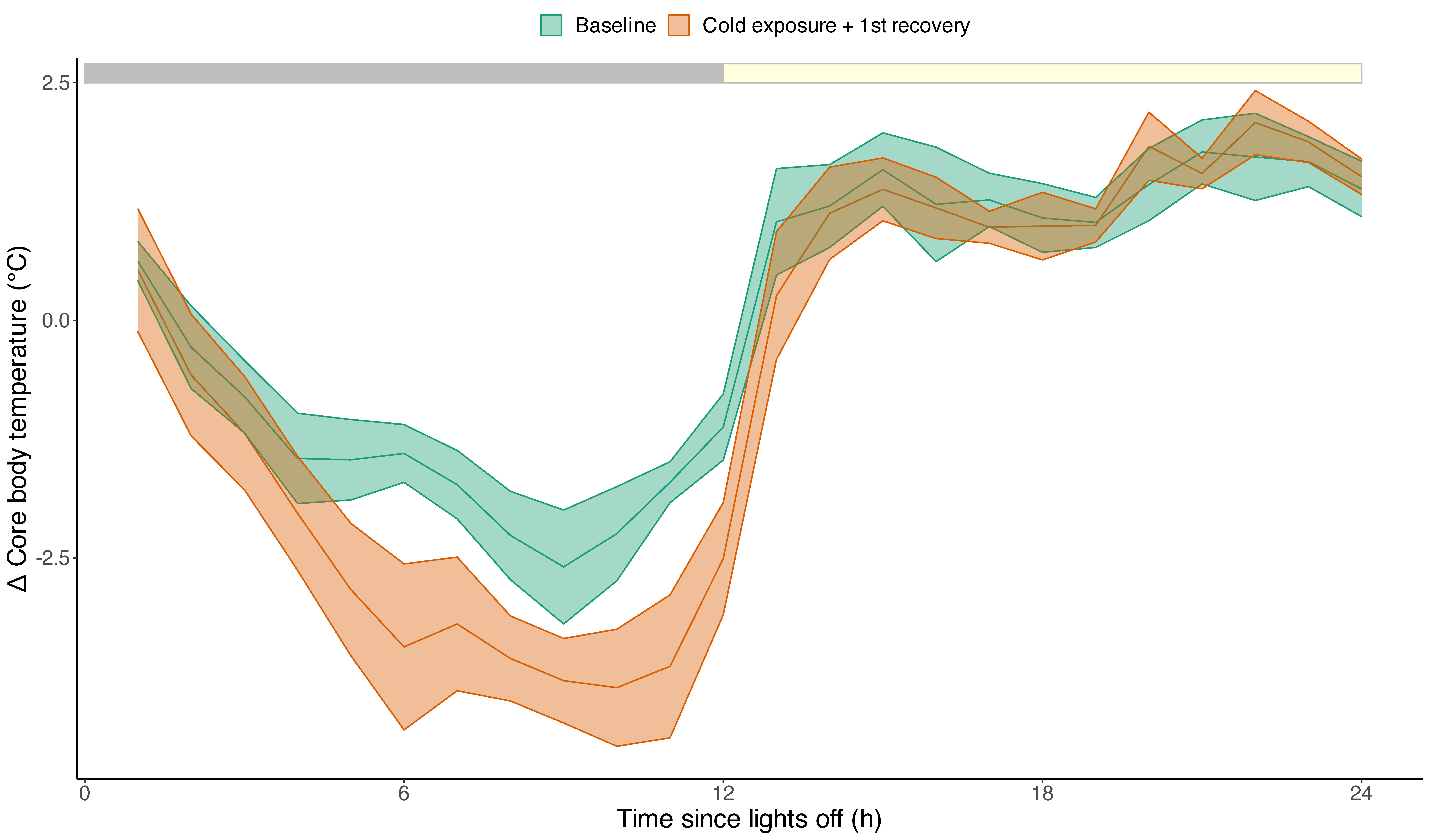
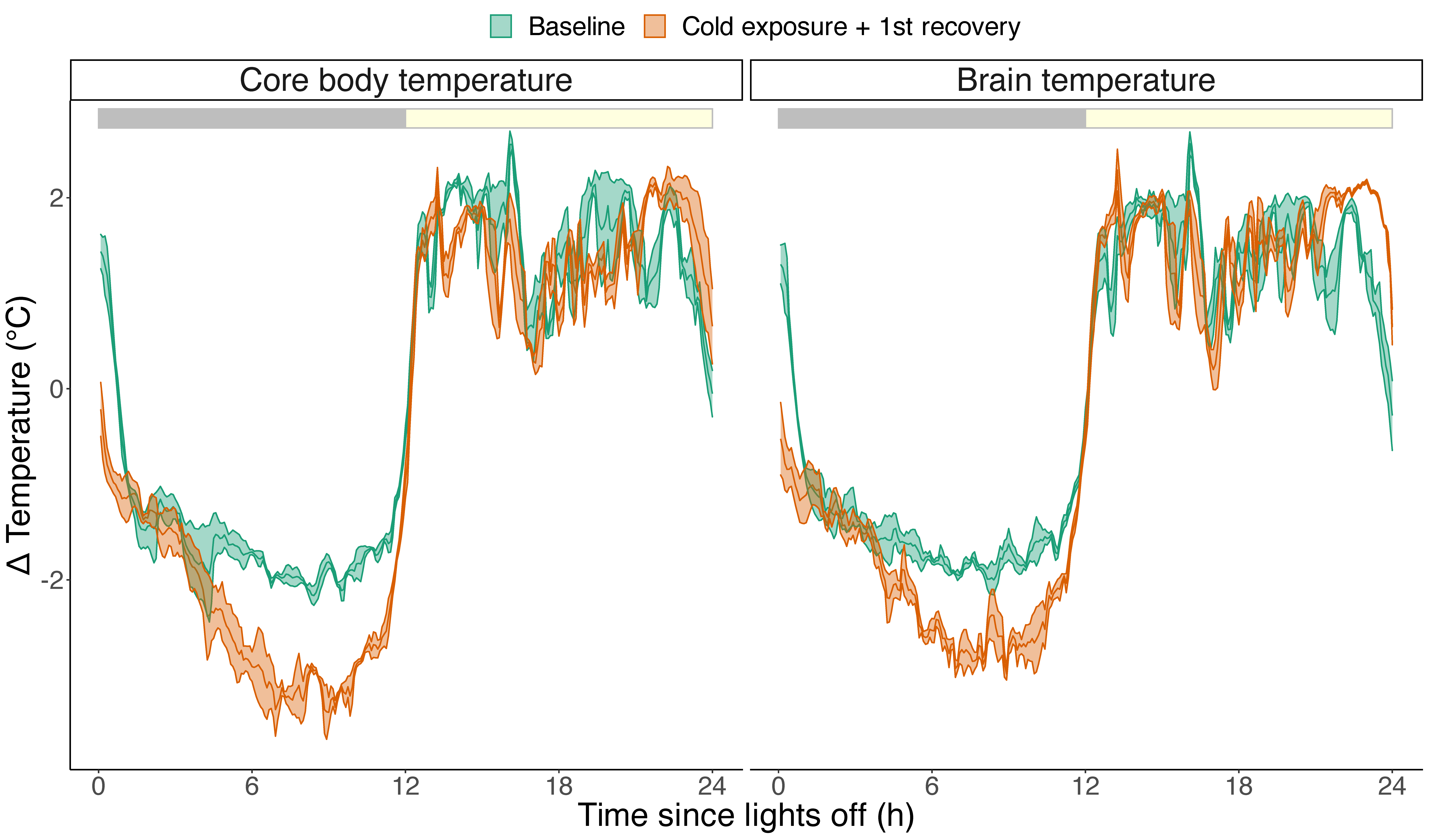
3.1. Effects of Cold Exposure on Body and Cortical Temperature
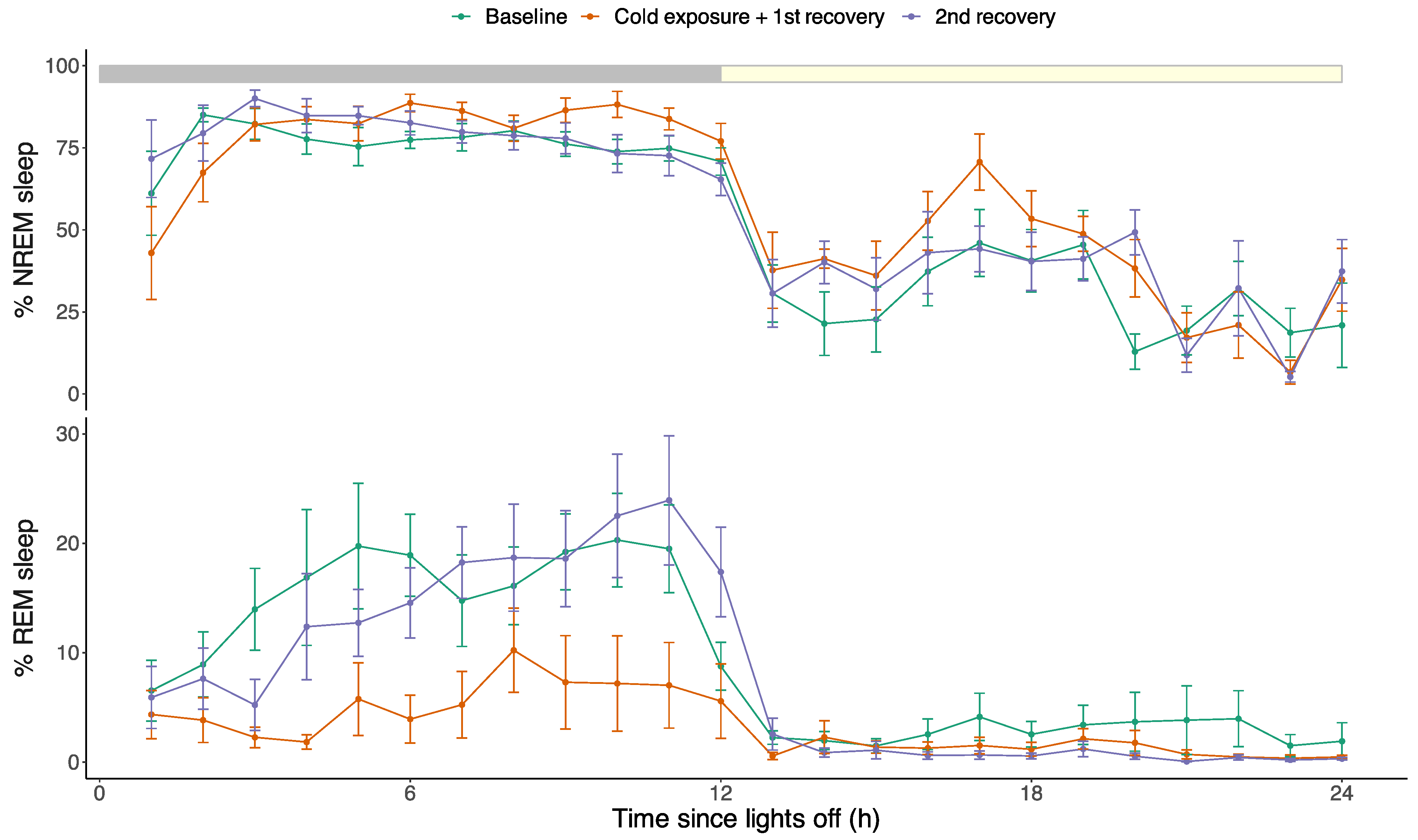
3.2. Effects of Cold Exposure on Sleep Architecture
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campbell, S.S.; Tobler, I. Animal Sleep: A Review of Sleep Duration across Phylogeny. Neurosci. Biobehav. Rev. 1984, 8, 269–300. [Google Scholar] [CrossRef]
- Anafi, R.C.; Kayser, M.S.; Raizen, D.M. Exploring Phylogeny to Find the Function of Sleep. Nat. Rev. Neurosci. 2019, 20, 109–116. [Google Scholar] [CrossRef]
- Joiner, W.J. Unraveling the Evolutionary Determinants of Sleep. Curr. Biol. 2016, 26, R1073–R1087. [Google Scholar] [CrossRef]
- Benington, J.H. Sleep Homeostasis and the Function of Sleep. Sleep 2000, 23, 959–966. [Google Scholar] [CrossRef]
- Siegel, J.M. Clues to the Functions of Mammalian Sleep. Nature 2005, 437, 1264–1271. [Google Scholar] [CrossRef]
- Schmidt, M.H. The Energy Allocation Function of Sleep: A Unifying Theory of Sleep, Torpor, and Continuous Wakefulness. Neurosci. Biobehav. Rev. 2014, 47, 122–153. [Google Scholar] [CrossRef]
- Raven, F.; Van der Zee, E.A.; Meerlo, P.; Havekes, R. The Role of Sleep in Regulating Structural Plasticity and Synaptic Strength: Implications for Memory and Cognitive Function. Sleep Med. Rev. 2018, 39, 3–11. [Google Scholar] [CrossRef]
- Hauglund, N.L.; Pavan, C.; Nedergaard, M. Cleaning the Sleeping Brain—The Potential Restorative Function of the Glymphatic System. Curr. Opin. Physiol. 2020, 15, 1–6. [Google Scholar] [CrossRef]
- Deboer, T. Behavioral and Electrophysiological Correlates of Sleep and Sleep Homeostasis. In Current Topics in Behavioral Neurosciences; Springer: Berlin/Heidelberg, Germany, 2013; Volume 25, pp. 1–24. ISBN 978-3-662-46878-4. [Google Scholar]
- Tobler, I.; Borbély, A.A. Sleep EEG in the Rat as a Function of Prior Waking. Electroencephalogr. Clin. Neurophysiol. 1986, 64, 74–76. [Google Scholar] [CrossRef]
- Dijk, D.J.; Beersma, D.G.M.; Daan, S. EEG Power Density during Nap Sleep: Reflection of an Hourglass Measuring the Duration of Prior Wakefulness. J. Biol. Rhythm. 1987, 2, 207–219. [Google Scholar] [CrossRef]
- Huber, R.; Deboer, T.; Tobler, I. Effects of Sleep Deprivation on Sleep and Sleep EEG in Three Mouse Strains: Empirical Data and Simulations. Brain. Res. 2000, 857, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Borbély, A.A.; Neuhaus, H.U. Sleep-Deprivation: Effects on Sleep and EEG in the Rat. J. Comp. Physiol. 1979, 133, 71–87. [Google Scholar] [CrossRef]
- Dement, W. The Effect of Dream Deprivation. Science 1960, 131, 1705–1707. [Google Scholar] [CrossRef] [PubMed]
- Borbély, A.A.; Tobler, I.; Hanagasioglu, M. Effect of Sleep Deprivation on Sleep and EEG Power Spectra in the Rat. Behav. Brain Res. 1984, 14, 171–182. [Google Scholar] [CrossRef]
- Cartwright, R.D.; Monroe, L.J.; Palmer, C. Individual Differences in Response to REM Deprivation. Arch. Gen. Psychiatry 1967, 16, 297–303. [Google Scholar] [CrossRef]
- Endo, T.; Roth, C.; Landolt, H.P.; Werth, E.; Aeschbach, D.; Achermann, P.; Borbély, A.A. Selective REM Sleep Deprivation in Humans: Effects on Sleep and Sleep EEG. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1998, 274, R1186–R1194. [Google Scholar] [CrossRef]
- Coolen, A.; Hoffmann, K.; Barf, R.P.; Fuchs, E.; Meerlo, P. Telemetric Study of Sleep Architecture and Sleep Homeostasis in the Day-Active Tree Shrew Tupaia Belangeri. Sleep 2012, 35, 879–888. [Google Scholar] [CrossRef]
- Borbely, A.A. A Two Process Model of Sleep Regulation. Hum. Neurobiol. 1982, 1, 195–204. [Google Scholar]
- Benington, J.H.; Heller, H.C. Does the Function of REM Sleep Concern Non-REM Sleep or Waking? Prog. Neurobiol. 1994, 44, 433–449. [Google Scholar] [CrossRef]
- Fuchs, E.; Corbach-Söhle, S. Tree Shrews. In The UFAW Handbook on the Care and Management of Laboratory and Other Research Animals; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2010; pp. 262–275. ISBN 9781444318777. [Google Scholar]
- Janečka, J.E.; Miller, W.; Pringle, T.H.; Wiens, F.; Zitzmann, A.; Helgen, K.M.; Springer, M.S.; Murphy, W.J. Molecular and Genomic Data Identify the Closest Living Relative of Primates. Science 2007, 318, 792–794. [Google Scholar] [CrossRef]
- Kriegs, J.O.; Churakov, G.; Jurka, J.; Brosius, J.; Schmitz, J. Evolutionary History of 7SL RNA-Derived SINEs in Supraprimates. Trends Genet. 2007, 23, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Dimanico, M.M.; Klaassen, A.L.; Wang, J.; Kaeser, M.; Harvey, M.; Rasch, B.; Rainer, G. Aspects of Tree Shrew Consolidated Sleep Structure Resemble Human Sleep. Commun. Biol. 2021, 4, 722. [Google Scholar] [CrossRef] [PubMed]
- Amici, R.; Zamboni, G.; Perez, E.; Jones, C.A.; Parmeggiani, P.L. The Influence of a Heavy Thermal Load on REM Sleep in the Rat. Brain Res. 1998, 781, 252–258. [Google Scholar] [CrossRef]
- Cerri, M.; Ocampo-Garces, A.; Amici, R.; Baracchi, F.; Capitani, P.; Jones, C.A.; Luppi, M.; Perez, E.; Parmeggiani, P.L.; Zamboni, G. Cold Exposure and Sleep in the Rat: Effects on Sleep Architecture and the Electroencephalogram. Sleep 2005, 28, 694–705. [Google Scholar] [CrossRef] [PubMed]
- Amici, R.; Cerri, M.; Ocampo-Garcés, A.; Baracchi, F.; Dentico, D.; Jones, C.A.; Luppi, M.; Perez, E.; Parmeggiani, P.L.; Zamboni, G. Cold Exposure and Sleep in the Rat: REM Sleep Homeostasis and Body Size. Sleep 2008, 31, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Parmeggiani, P.L.; Rabini, C. Sleep and Environmental Temperature. Arch. Ital. Biol. 1970, 108, 369–387. [Google Scholar]
- Allocca, G.; Ma, S.; Martelli, D.; Cerri, M.; Del Vecchio, F.; Bastianini, S.; Zoccoli, G.; Amici, R.; Morairty, S.R.; Aulsebrook, A.E.; et al. Validation of ‘Somnivore’, a Machine Learning Algorithm for Automated Scoring and Analysis of Polysomnography Data. Front. Neurosci. 2019, 13, 207. [Google Scholar] [CrossRef]
- R Development Core Team 3.0.1. A Language and Environment for Statistical Computing. In R Foundation for Statistical Computing; Vienna, Austria, 2013; Volume 2, Available online: https://www.R-project.org (accessed on 28 March 2023).
- Lenth, R.V. Least-Squares Means: The R Package Lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef]
- Franken, P.; Tobler, I.; Borbély, A.A. Effects of 12-h Sleep Deprivation and of 12-h Cold Exposure on Sleep Regulation and Cortical Temperature in the Rat. Physiol. Behav. 1993, 54, 885–894. [Google Scholar] [CrossRef]
- Lyamin, O.I.; Kosenko, P.O.; Korneva, S.M.; Vyssotski, A.L.; Mukhametov, L.M.; Siegel, J.M. Fur Seals Suppress REM Sleep for Very Long Periods without Subsequent Rebound. Curr. Biol. 2018, 28, 2000–2005.e2. [Google Scholar] [CrossRef]
- Parmeggiani, P.L.; Cianci, T.; Calasso, M.; Zamboni, G.; Perez, E. Quantitative Analysis of Short Term Deprivation and Recovery of Desynchronized Sleep in Cats. Electroencephalogr. Clin. Neurophysiol. 1980, 50, 293–300. [Google Scholar] [CrossRef]
- Franken, P. Long-Term vs. Short-Term Processes Regulating REM Sleep. J. Sleep Res. 2002, 11, 17–28. [Google Scholar] [CrossRef]
- Benca, R.M.; Bergmann, B.M.; Leung, C.; Nummy, D.; Rechtschaffen, A. Rat Strain Differences in Response to Dark Pulse Triggering of Paradoxical Sleep. Physiol. Behav. 1991, 49, 83–87. [Google Scholar] [CrossRef]
- Leung, C.; Bergmann, B.M.; Rechtschaffen, A.; Benca, R.M. Heritability of Dark Pulse Triggering of Paradoxical Sleep in Rats. Physiol. Behav. 1992, 52, 127–131. [Google Scholar] [CrossRef]
- Rampin, C.; Cespuglio, R.; Chastrette, N.; Jouvet, M. Immobilisation Stress Induces a Paradoxical Sleep Rebound in Rat. Neurosci. Lett. 1991, 126, 113–118. [Google Scholar] [CrossRef]
- Meerlo, P.; Easton, A.; Bergmann, B.M.; Turek, F.W. Restraint Increases Prolactin and REM Sleep in C57BL/6J Mice but Not in BALB/CJ Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, R846–R854. [Google Scholar] [CrossRef]
- Smith, C.; Lapp, L. Prolonged Increases in Both PS and Number of REMs Following a Shuttle Avoidance Task. Physiol. Behav. 1986, 36, 1053–1057. [Google Scholar] [CrossRef]
- Smith, C.; Young, J.; Young, W. Prolonged Increases in Paradoxical Sleep during and after Avoidance-Task Acquisition. Sleep 1980, 3, 67–81. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Hasselt, S.J.; Epifani, L.; Zantinge, D.; Vitkute, K.; Kas, M.J.H.; Allocca, G.; Meerlo, P. A Study on REM Sleep Homeostasis in the Day-Active Tree Shrew (Tupaia belangeri): Cold-Induced Suppression of REM Sleep Is Not Followed by a Rebound. Biology 2023, 12, 614. https://doi.org/10.3390/biology12040614
van Hasselt SJ, Epifani L, Zantinge D, Vitkute K, Kas MJH, Allocca G, Meerlo P. A Study on REM Sleep Homeostasis in the Day-Active Tree Shrew (Tupaia belangeri): Cold-Induced Suppression of REM Sleep Is Not Followed by a Rebound. Biology. 2023; 12(4):614. https://doi.org/10.3390/biology12040614
Chicago/Turabian Stylevan Hasselt, Sjoerd J., Luisa Epifani, Danique Zantinge, Kornelija Vitkute, Martien J. H. Kas, Giancarlo Allocca, and Peter Meerlo. 2023. "A Study on REM Sleep Homeostasis in the Day-Active Tree Shrew (Tupaia belangeri): Cold-Induced Suppression of REM Sleep Is Not Followed by a Rebound" Biology 12, no. 4: 614. https://doi.org/10.3390/biology12040614