
Climate Change and Human Activities, the Significant Dynamic Drivers of Himalayan Goral Distribution (Naemorhedus goral)

 , , ,
, , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
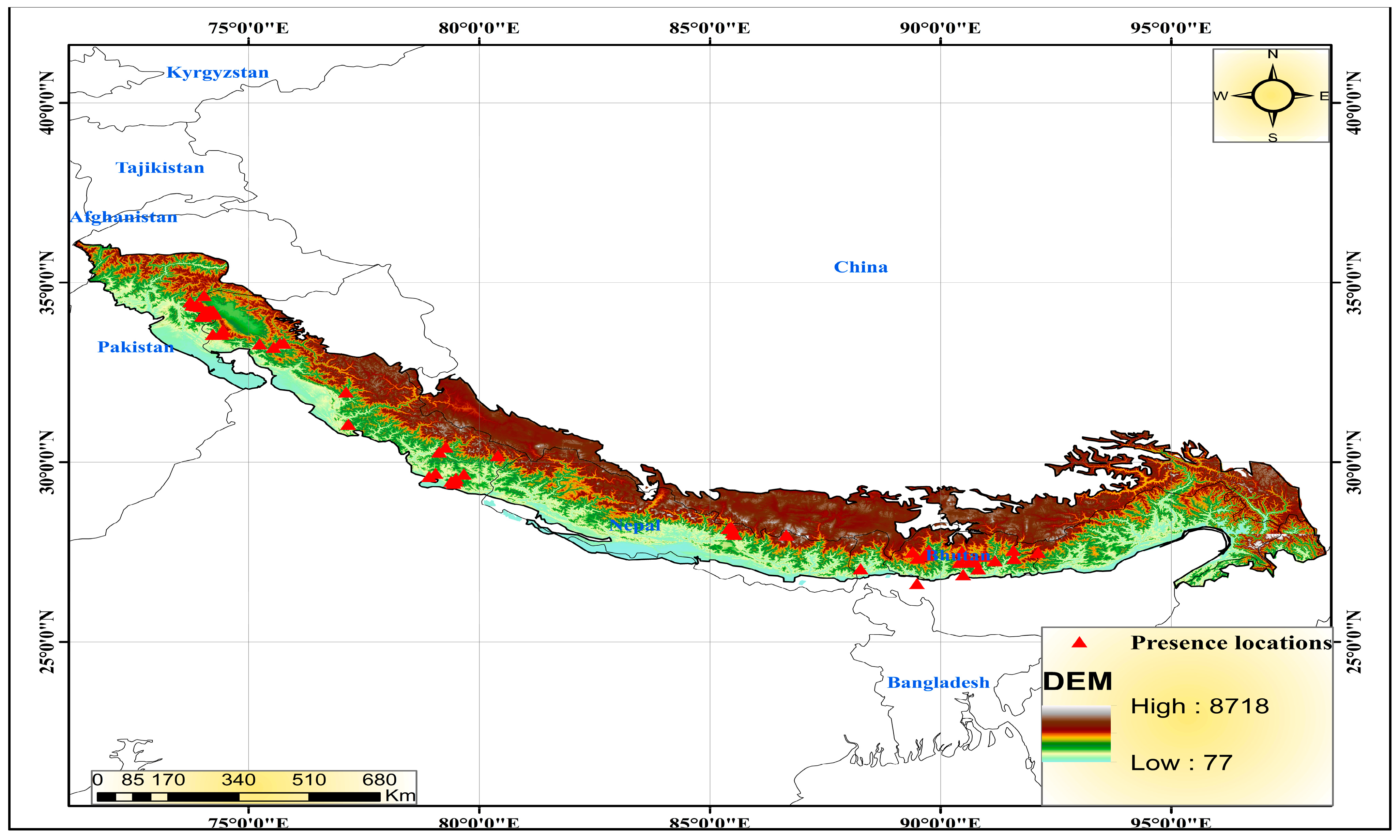
2.1. Study Area
2.2. Species Presence Data
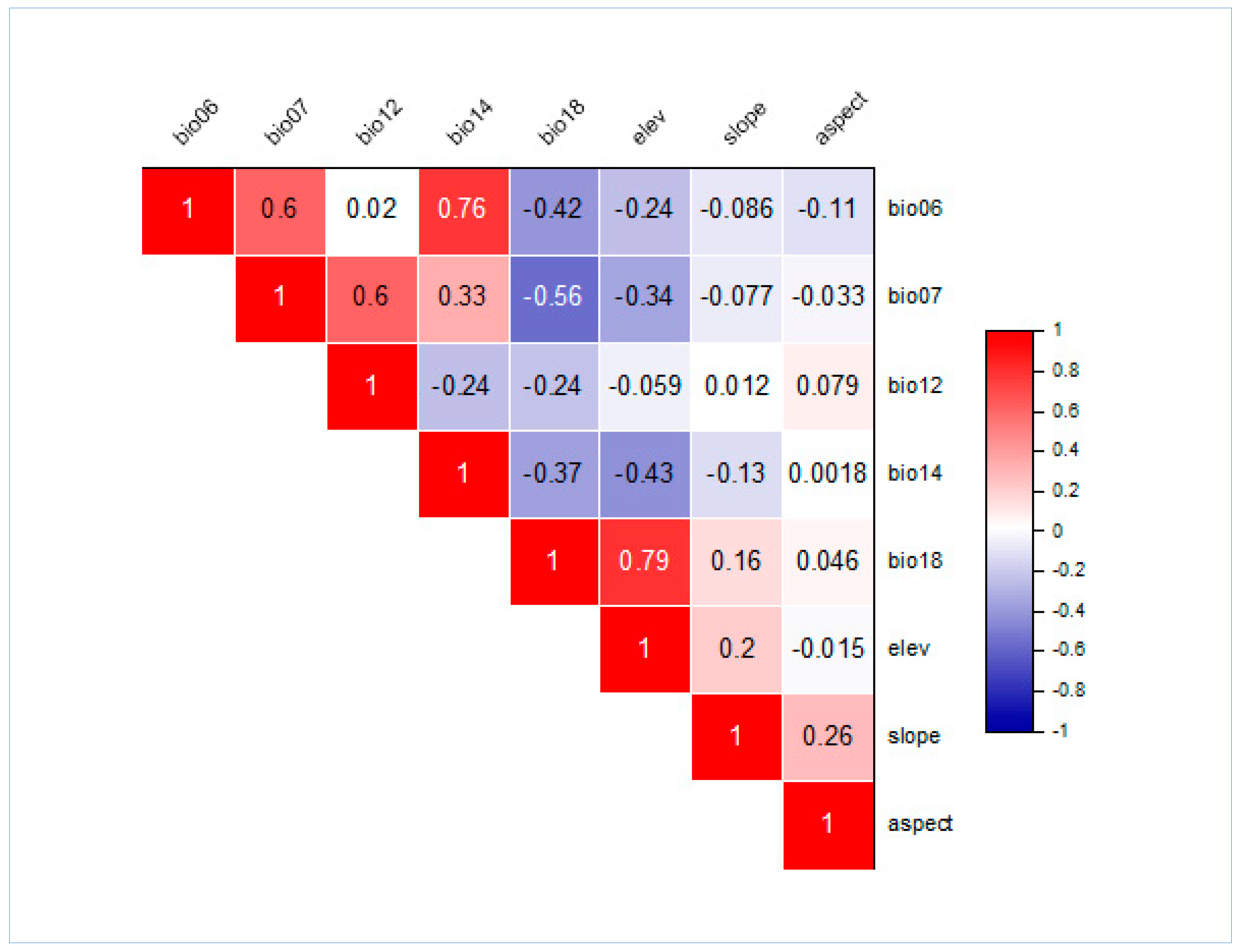
2.3. Environmental Data Collection and Variable Selection
2.4. Preliminary Varaibles Processing
2.5. Model Calibration and Optimization
2.6. Model Evaluation and Preditions Reclassification
3. Results
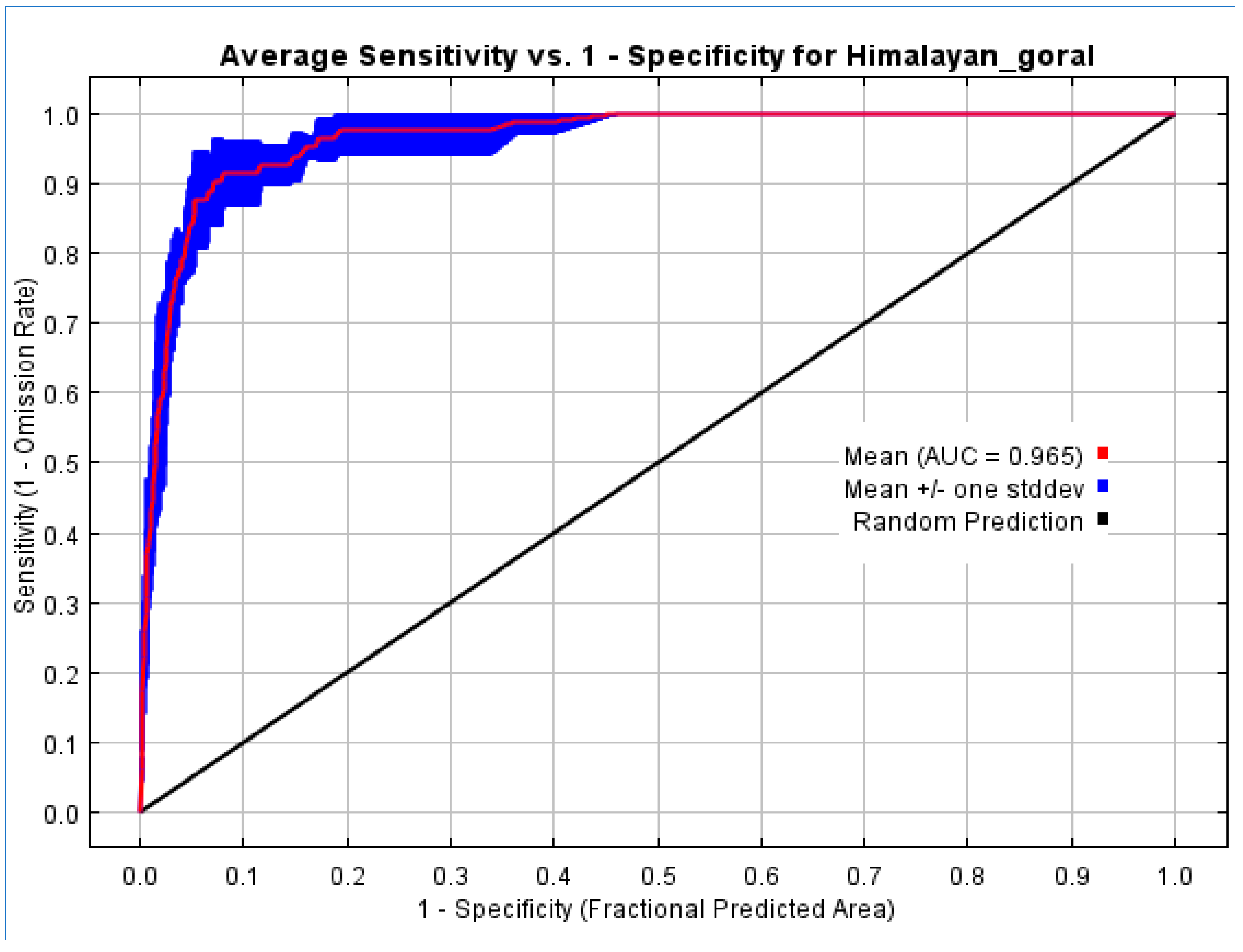
3.1. Model Evaluation
3.2. Environmental Variables Responsible for the Distribution
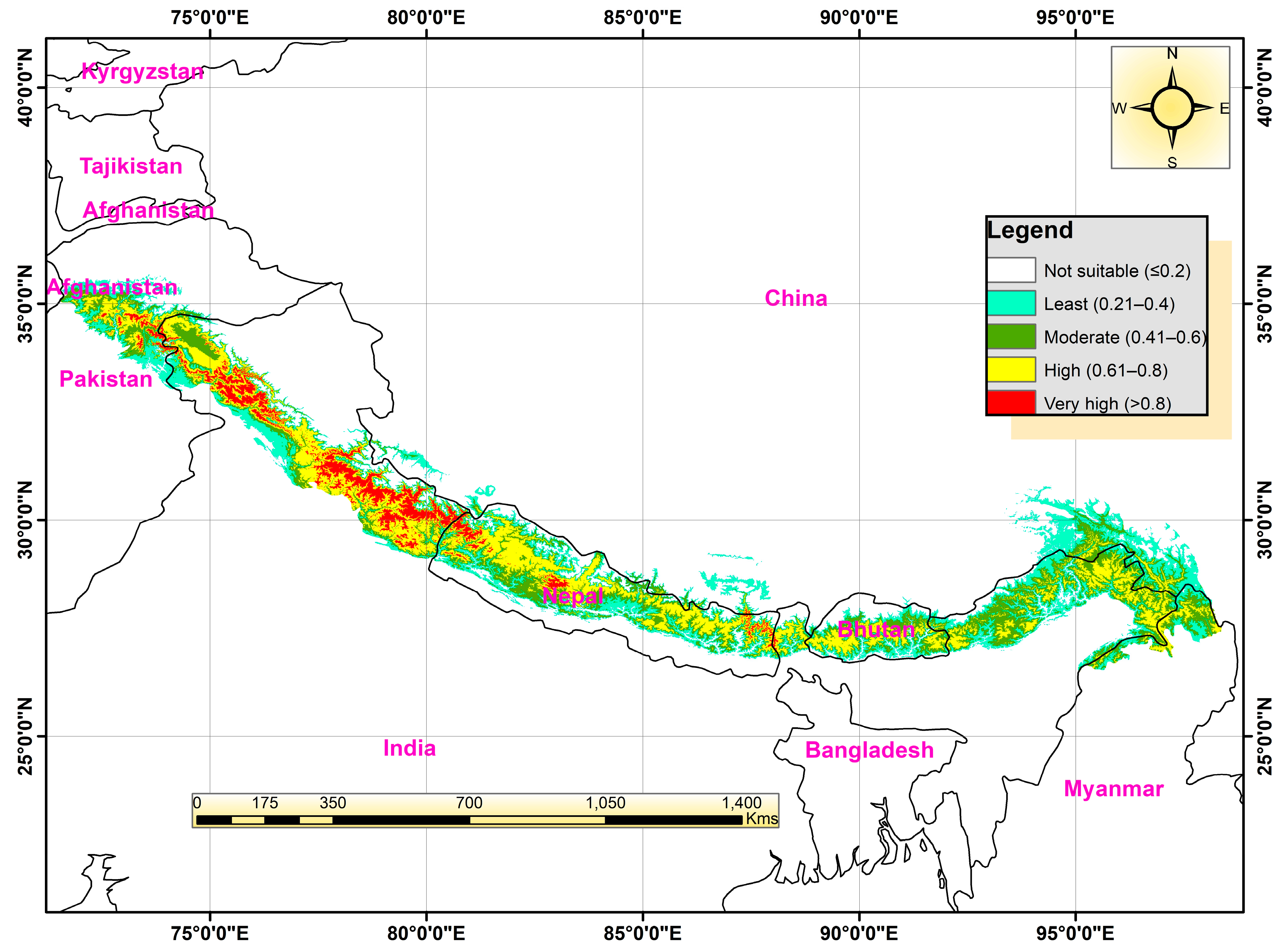
3.3. Current Distribution
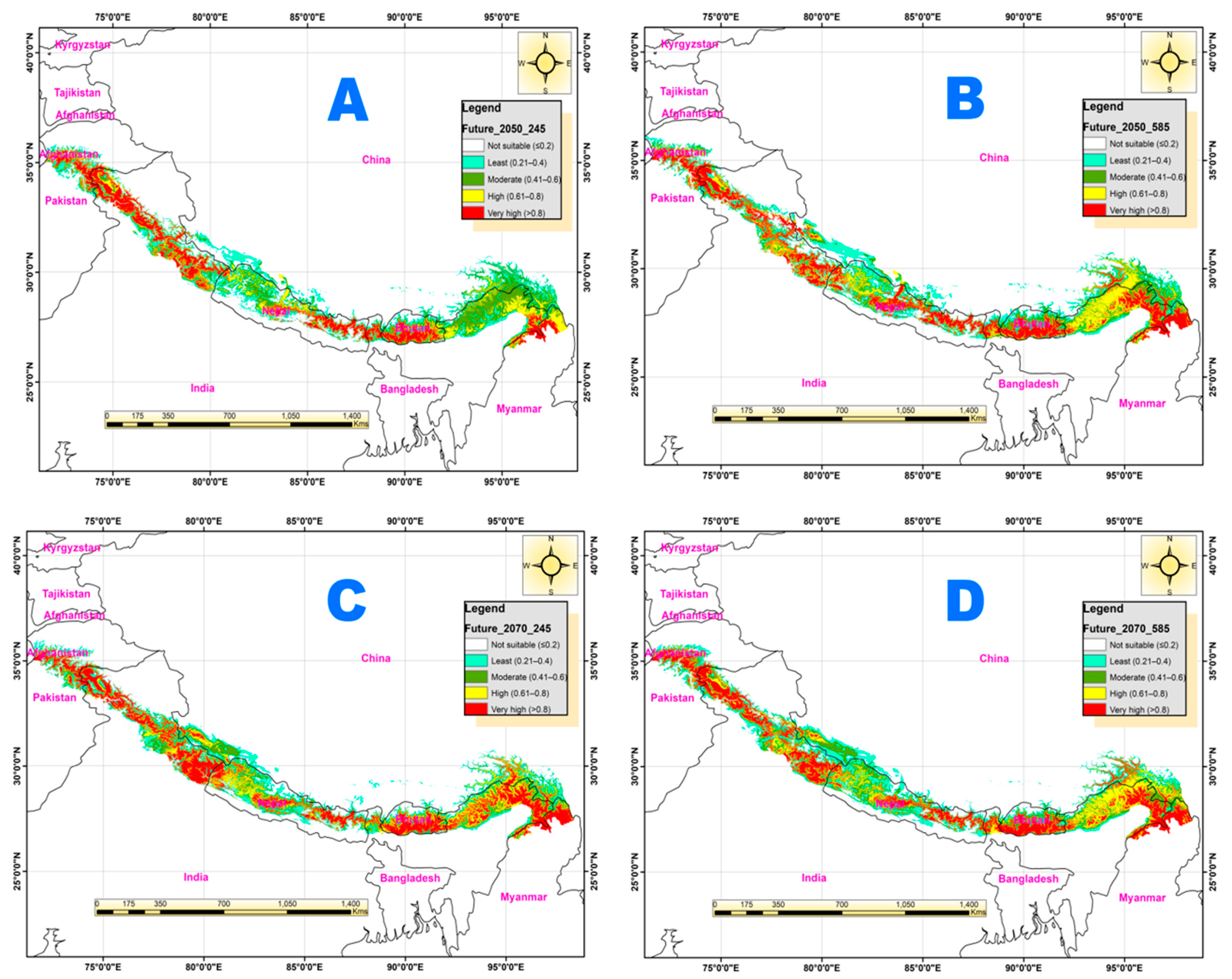
3.4. Potential Habitat Suitability under Future Climate Change Scenarios
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jamwal, P.S.; Di Febbraro, M.; Carranza, M.L.; Savage, M.; Loy, A. Global change on the roof of the world: Vulnerability of Himalayan otter species to land use and climate alterations. Div. Distrib. 2022, 28, 1635–1649. [Google Scholar] [CrossRef]
- Negi, G.C.S.; Rawal, R.S. Himalayan biodiversity in the face of climate change. In Tropical Ecosystems: Structure, Functions and Challenges in the Face of Global Change; Springer: Singapore, 2019; pp. 263–277. [Google Scholar]
- Díaz, S.; Malhi, Y. Biodiversity: Concepts, patterns, trends, and perspectives. Annu. Rev. Env. Resour. 2022, 47, 31–63. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2022-2. Available online: https://www.iucnredlist.org/ (accessed on 10 December 2022).
- Cruz, C.; Santulli-Sanzo, G.; Ceballos, G. Global patterns of raptor distribution and protected areas optimal selection to reduce the extinction crises. Proc. Natl. Acad. Sci. USA 2021, 118, e2018203118. [Google Scholar] [CrossRef] [PubMed]
- Caro, T.; Rowe, Z.; Berger, J.; Wholey, P.; Dobson, A. An inconvenient misconception: Climate change is not the principal driver of biodiversity loss. Conserv. Lett 2022, 15, e12868. [Google Scholar] [CrossRef]
- Manes, S.; Costello, M.J.; Beckett, H.; Debnath, A.; Devenish-Nelson, E.; Grey, K.A.; Jenkins, R.; Khan, T.M.; Kiessling, W.; Krause, C.; et al. Endemism increases species’ climate change risk in areas of global biodiversity importance. Biol. Conserv. 2021, 257, 109070. [Google Scholar] [CrossRef]
- Seddon, N.; Smith, A.; Smith, P.; Key, I.; Chausson, A.; Girardin, C.; House, J.; Srivastava, S.; Turner, B. Getting the message right on nature-based solutions to climate change. Glob. Change Biol. Bioenergy 2021, 27, 1518–1546. [Google Scholar] [CrossRef]
- Zhang, Y.; Tariq, A.; Hughes, A.C.; Hong, D.; Wei, F.; Sun, H.; Sardans, J.; Peñuelas, J.; Perry, G.; Qiao, J.; et al. Challenges and solutions to biodiversity conservation in arid lands. Sci. Total Environ. 2022, 857, 159695. [Google Scholar] [CrossRef]
- Frans, V.F.; Augé, A.A.; Fyfe, J.; Zhang, Y.; McNally, N.; Edelhoff, H.; Balkenhol, N.; Engler, J.O. Integrated SDM database: Enhancing the relevance and utility of species distribution models in conservation management. Methods Ecol. Evol. 2022, 13, 243–261. [Google Scholar] [CrossRef]
- Hernandez, J.O.; Buot Jr, I.E.; Park, B.B. Prioritizing choices in the conservation of flora and fauna: Research trends and methodological approaches. Land 2022, 11, 1645. [Google Scholar] [CrossRef]
- Curd, A.; Chevalier, M.; Vasquez, M.; Boyé, A.; Firth, L.B.; Marzloff, M.P.; Bricheno, L.M.; Burrows, M.T.; Bush, L.E.; Cordier, C.; et al. Applying landscape metrics to species distribution model predictions to characterize internal range structure and associated changes. Glob. Change Biol. Bioenergy 2023, 29, 631–647. [Google Scholar] [CrossRef]
- Arenas-Castro, S.; Regos, A.; Martins, I.; Honrado, J.; Alonso, J. Effects of input data sources on species distribution model predictions across species with different distributional ranges. J. Biogeogr. 2022, 49, 1299–1312. [Google Scholar] [CrossRef]
- Arshad, F.; Waheed, M.; Fatima, K.; Harun, N.; Iqbal, M.; Fatima, K.; Umbreen, S. Predicting the suitable current and future potential distribution of the native endangered tree Tecomella undulata (Sm.) Seem. in Pakistan. Sustainability 2022, 14, 7215. [Google Scholar] [CrossRef]
- Dhyani, S.; Singh, S.; Kadaverugu, R.; Pujari, P.; Verma, P. Habitat suitability modelling and nature-based solutions: An efficient combination to realise the targets of Bonn challenge and SDGs in South Asia. In Nature-Based Solutions for Resilient Ecosystems and Societies; Springer: Berlin/Heidelberg, Germany, 2020; pp. 347–364. [Google Scholar]
- Fedriani, J.M.; Ayllón, D.; Wiegand, T.; Grimm, V. Intertwined effects of defaunation, increased tree mortality and density compensation on seed dispersal. Ecography 2020, 43, 1352–1363. [Google Scholar] [CrossRef]
- Duckworth, J.W.; MacKinnon, J. Naemorhedus goral . In The IUCN Red List of Threatened Species 2008; IUCN: Gland, Switzerland, 2008. [Google Scholar]
- Abbas, F.; Ruckstuhl, K.E.; Mian, A.; Akhtar, T.; Rooney, T.P. Distribution, population size, and structure of Himalayan grey goral Naemorhedus goral bedfordi (Cetartiodactyla: Bovidae) in Pakistan. Mammalia 2012, 76, 143–147. [Google Scholar]
- Sathyakumar, S. Species of the greater Himalaya. In ENVIS Bulletin: Wildlife and Protected Areas; Sathyakumar, S., Ed.; Wildlife Institute of India: Dehradun, India, 2002; pp. 44–49. [Google Scholar]
- Cooke, S.J. Biotelemetry and biologging in endangered species research and animal conservation: Relevance to regional, national, and IUCN Red List threat assessments. Endanger. Species Res. 2008, 4, 165–185. [Google Scholar] [CrossRef]
- Suraprasit, K.; Jaeger, J.J.; Shoocongdej, R.; Chaimanee, Y.; Wattanapituksakul, A.; Bocherens, H. Long-term isotope evidence on the diet and habitat breadth of Pleistocene to Holocene caprines in Thailand: Implications for the extirpation and conservation of Himalayan gorals. Front. Ecol. Evol. 2020, 8, 67. [Google Scholar] [CrossRef]
- Chatterjee, S.; Goswami, A.; Scotese, C.R. The longest voyage: Tectonic, magmatic, and paleoclimatic evolution of the Indian plate during its northward flight from Gondwana to Asia. Gondwana Res. 2013, 23, 238–267. [Google Scholar] [CrossRef]
- Yin, A.; Harrison, T.M. Geologic evolution of the Himalayan-Tibetan orogen. Annu. Rev. Earth Planet. Sci. 2000, 28, 211–280. [Google Scholar] [CrossRef]
- Hassan, T.; Hamid, M.; Wani, S.A.; Malik, A.H.; Waza, S.A.; Khuroo, A.A. Substantial shifts in flowering phenology of Sternbergia vernalis in the Himalaya: Supplementing decadal field records with historical and experimental evidences. Sci. Total Environ. 2021, 795, 148811. [Google Scholar] [CrossRef]
- Gao, X.; Liu, J.; Huang, Z. The impact of climate change on the distribution of rare and endangered tree Firmiana kwangsiensis using the Maxent modeling. Ecol. Evol. 2022, 12, e9165. [Google Scholar] [CrossRef]
- Bosso, L.; Di Febbraro, M.; Cristinzio, G.; Zoina, A.; Russo, D. Shedding light on the effects of climate change on the potential distribution of Xylella fastidiosa in the Mediterranean Basin. Biol. Invasions 2016, 18, 1759–1768. [Google Scholar] [CrossRef]
- Khattak, R.H.; Teng, L.; Ahmad, S.; Bari, F.; Rehman, E.U.; Shah, A.A.; Liu, Z. In pursuit of new spaces for threatened mammals: Assessing habitat suitability for Kashmir Markhor (Capra falconeri cashmeriensis) in the Hindukush Range. Sustainability 2022, 14, 1544. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high-resolution interpolated climate surfaces for global land areas. Int. J. Climatol 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef]
- Taylor, K.E.; Stouffer, R.J.; Meehl, G.A. An overview of CMIP5 and the experiment design. Bull. Am. Meteorol. Soc. 2012, 93, 485–498. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Bai, D.-F.; Chen, P.-J.; Atzeni, L.; Cering, L.; Li, Q.; Shi, K. Assessment of habitat Suitability of the Snow Leopard (Panthera uncia) in Qomolangma National Nature Reserve Based on MaxEnt Modeling. Zool. Res. 2018, 39, 373. [Google Scholar]
- Fourcade, Y.; Engler, J.O.; Rödder, D.; Secondi, J. Mapping species distributions with MAXENT using a geographically biased sample of presence data: A Performance Assessment of Methods for Correcting Sampling Bias. PLoS ONE 2014, 9, e97122. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Summers, D.M.; Bryan, B.A.; Crossman, N.D.; Meyer, W.S. Species vulnerability to climate change: Impacts on spatial conservation priorities and species representation. Glob. Change Biol. 2012, 18, 2335–2348. [Google Scholar] [CrossRef]
- Elith, J.H.; Graham, C.P.; Anderson, R.; Dudík, M.; Ferrier, S.; Guisan, A.J.; Hijmans, R.; Huettmann, F.R.; Leathwick, J.; Lehmann, A. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef]
- Deb, J.C.; Forbes, G.; MacLean, D.A. Modelling the spatial distribution of selected North American woodland mammals under future climate scenarios. Mamm. Rev. 2020, 50, 440–452. [Google Scholar] [CrossRef]
- Yang, X.; Kushwaha, S.; Saran, S.; Xu, J.; Roy, P. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Vilà, M.; Gassó, N.; Thuiller, W.; Pino, J. Potential distribution range of invasive plant species in Spain. NeoBiota 2012, 12, 25–40. [Google Scholar] [CrossRef]
- Zhao, D.; He, H.; Wang, W.; Wang, L.; Du, H.; Liu, K.; Zong, S. Predicting wetland distribution changes under climate change and human activities in a Mid- and High-Latitude Region. Sustainability 2018, 10, 863. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Le Lay, G. Habitat Suitability Modelling and Niche Theory. J. Appl. Ecol. 2008, 45, 1372–1381. [Google Scholar] [CrossRef]
- Smeraldo, S.; Di Febbraro, M.; Ćirović, D.; Bosso, L.; Trbojević, I.; Russo, D. Species distribution models as a tool to predict range expansion after reintroduction: A case study on Eurasian beavers (Castor fiber). J. Nat. Conserv. 2017, 37, 12–20. [Google Scholar] [CrossRef]
- Wogan, G.O. Life history traits and niche instability impact accuracy and temporal transferability for historically calibrated distribution models of North American birds. PLoS ONE 2016, 11, e0151024. [Google Scholar] [CrossRef]
- Domíguez-Vega, H.; Monroy-Vilchis, O.; Balderas-Valdivia, C.J.; Gienger, C.M.; Ariano-Sánchez, D. Predicting the potential distribution of the beaded lizard and identification of priority areas for conservation. J. Nat. Conserv. 2012, 20, 247–253. [Google Scholar] [CrossRef]
- Ahmad, S.; Yang, L.; Khan, T.U.; Wanghe, K.; Li, M.; Luan, X. Using an ensemble modelling approach to predict the potential distribution of Himalayan gray goral (Naemorhedus goral bedfordi) in Pakistan. Glob. Ecol. Conserv. 2020, 21, e00845. [Google Scholar] [CrossRef]
- Thapa, A.; Thapa, S.; Poudel, B.S. Habitat and distribution of Goral (Naemorhedus goral) in Mahabharat Goral Wildlife Conservation Area, Nawalparasi, Western Nepal. Nepal J. For. 2011, 14, 1. [Google Scholar]
- Cavallini, P. Status of goral (Naemorhaedus goral) in Himachal Pradesh, India. Caprinae News 1990, 5, 4–6. [Google Scholar]
- Acharya, D.; Aryal, B.; Joshi, S. Locally declared Goral Conservation Area. Conserv. Watch-Nepal. News 2008, 1, 1. [Google Scholar]
- Quintero, I.; Wiens, J.J. Rates of projected climate change dramatically exceed past rates of climatic niche evolution among vertebrate species. Ecol. Lett. 2013, 16, 1095–1103. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution: Int. J. Org. Evol. 2008, 62, 2868–2883. [Google Scholar] [CrossRef]
- Muñoz-Mendoza, C.; D’Elía, G.; Panzera, A.; Villalobos-Leiva, A.; Sites, J.W., Jr.; Victoriano, P.F. Geography and past climate changes have shaped the evolution of a widespread lizard from the Chilean hotspot. Mol. Phylogenet. Evol. 2017, 116, 157–171. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef]
- Thuiller, W.; Broennimann, O.; Hughes, G.; Alkemade, J.R.M.; Midgley, G.F.; Corsi, F. Vulnerability of African mammals to anthropogenic climate change under conservative land transformation assumptions. Glob. Change Biol. 2006, 12, 424–440. [Google Scholar] [CrossRef]
- Levinsky, I.; Skov, F.; Svenning, J.C.; Rahbek, C. Potential impacts of climate change on the distributions and diversity patterns of European mammals. Biodiv. Conserv. 2007, 16, 3803–3816. [Google Scholar] [CrossRef]
- Ogawa-Onishi, Y.; Berry, P.M.; Tanaka, N. Assessing the potential impacts of climate change and their conservation implications in Japan: A case study of conifers. Biol. Conserv. 2010, 143, 1728–1736. [Google Scholar] [CrossRef]
- Khanum, R.; Mumtaz, A.S.; Kumar, S. Predicting impacts of climate change on medicinal asclepiads of Pakistan using Maxent modeling. Acta Oecologica 2013, 49, 23–31. [Google Scholar] [CrossRef]
- Abolmaali, S.M.R.; Tarkesh, M.; Bashari, H. MaxEnt modeling for predicting suitable habitats and identifying the effects of climate change on a threatened species, Daphne mucronata, in central Iran. Ecol. Inf. 2018, 43, 116–123. [Google Scholar] [CrossRef]
- Kadmon, R.; Farber, O.; Danin, A. Effect of roadside bias on the accuracy of predictive maps produced by bioclimatic models. Ecol. Appl. 2004, 14, 401–413. [Google Scholar] [CrossRef]
- Segurado, P.; Araujo, M.B.; Kunin, W.E. Consequences of spatial autocorrelation for nichebased models. J. Appl. Ecol. 2006, 43, 433–444. [Google Scholar] [CrossRef]
- Khan, A.M.; Li, Q.; Saqib, Z.; Khan, N.; Habib, T.; Khalid, N.; Majeed, M.; Tariq, A. MaxEnt Modelling and Impact of Climate Change on Habitat Suitability Variations of Economically Important Chilgoza Pine (Pinus gerardiana Wall.) in South Asia. Forests 2022, 13, 715. [Google Scholar] [CrossRef]
- Ali, F.; Khan, N.; Khan, A.M.; Ali, K.; Abbas, F. Species distribution modelling of Monotheca buxifolia (Falc.) A. DC.: Present distribution and impacts of potential climate change. Heliyon 2023, 9, e13417. [Google Scholar] [CrossRef]
- Kabir, M.; Hameed, S.; Ali, H.; Bosso, L.; Din, J.U.; Bischof, R.; Redpath, S.; Nawaz, M.A. Habitat suitability and movement corridors of grey wolf (Canis lupus) in Northern Pakistan. PLoS ONE 2017, 12, e0187027. [Google Scholar] [CrossRef]
- Haq, S.M.; Yaqoob, U.; Calixto, E.S.; Kumar, M.; Rahman, I.U.; Hashem, A.; Abd_Allah, E.F.; Alakeel, M.A.; Alqarawi, A.A.; Abdalla, M.; et al. Long-term impact of transhumance pastoralism and associated disturbances in high-altitude forests of Indian Western Himalaya. Sustainability 2021, 13, 12497. [Google Scholar] [CrossRef]
- Haq, S.M.; Calixto, E.S.; Yaqoob, U.; Ahmed, R.; Mahmoud, A.H.; Bussmann, R.W.; Mohammed, O.B.; Ahmad, K.; Abbasi, A.M. Traditional usage of wild fauna among the local inhabitants of Ladakh, Trans-Himalayan Region. Animals 2020, 10, 2317. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.U.; Adair, E.C.; Cardinale, B.J.; Byrnes, J.E.K.; Hungate, B.A.; Matulich, K.L.; Gonzalez, A.; Duffy, J.E.; Gamfeldt, L.; Connor, M.I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 2012, 486, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Haq, S.M.; Yaqoob, U.; Majeed, M.; Amjad, M.S.; Hassan, M.; Ahmad, R.; Waheed, M.; Bussmann, R.W.; Soares Calixto, E.; Proćków, J.; et al. Quantitative ethnoveterinary study on plant resource utilization by indigenous communities in high-altitude regions. Front. Vet. Sci. 2022, 9, 944046. [Google Scholar] [CrossRef]
- Kanagaraj, R.; Araujo, M.B.; Barman, R.; Davidar, P.; De, R.; Digal, D.K.; Gopi, G.V.; Johnsingh, A.J.T.; Kakati, K.; Kramer-Schadt, S.; et al. Predicting range shifts of Asian elephants under global change. Div. Distrib. 2019, 25, 822–838. [Google Scholar] [CrossRef]
- Waheed, M.; Arshad, F.; Majeed, M.; Haq, S.M.; Aziz, R.; Bussmann, R.W.; Ali, K.; Subhan, F.; Jones, D.A.; Zaitouny, A. Potential distribution of a noxious weed (Solanum viarum Du-nal), current status, and future invasion risk based on MaxEnt modeling. Geol. Ecol. Landsc. 2023, 1–16. [Google Scholar] [CrossRef]
- Yamaura, Y.; Higa, M.; Senzaki, M.; Koizumi, I. Can charismatic megafauna be surrogate species for biodiversity conservation? Mechanisms and a test using citizen data and a hierarchical community model. In Biodiversity Conservation Using Umbrella Species: Blakiston’s Fish Owl and the Red-Crowned Crane; Nakamura, F., Ed.; Springer: Singapore, 2018; pp. 151–179. [Google Scholar]
- Haq, S.M.; Hassan, M.; Jan, H.A.; Al-Ghamdi, A.A.; Ahmad, K.; Abbasi, A.M. Traditions for Future Cross-National Food Security—Food and Foraging Practices among Different Native Communities in the Western Himalayas. Biology 2022, 11, 455. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Code | Percent Contribution |
|---|---|---|
| Annual Precipitation | bio12 | 38.2 |
| Elevation | elev | 32.2 |
| Precipitation of Driest Month | bio14 | 11 |
| Aspect | spect | 8.2 |
| Min. Temperature of Coldest Month | bio06 | 4.6 |
| Slope | slope | 2.5 |
| Precipitation of Warmest Quarter | bio18 | 2.1 |
| Temperature Annual Range | bio07 | 1.3 |
| Climate Change Scenario | Predicted Probability of Occurrence of Himalayan Goral under Considered Habitat Suitability Classes | |||||
|---|---|---|---|---|---|---|
| Not Suitable | Least | Moderate | High | Very High | Total Suitable Land Area (km2) | |
| (p ≤ 0.2) | (p 0.21–0.4) | (p 0.41–0.6) | (p 0.61–0.8) | (p ≥ 0.81) | ||
| Current climate | 515,799 | 25,200 | 24,113 | 23,595 | 6294 | 79,201 |
| SSPs_245_2050 | 512,854 | 21,129 | 24,781 | 15,775 | 20,461 | 82,146 |
| Rate of change (%) | −0.6 | −17.6 | 2.7 | −40.3 | 117.9 | 3.7 |
| SSPs_585_2050 | 505,989 | 21,810 | 19,076 | 20,171 | 27,953 | 89,011 |
| Rate of change (%) | −1.9 | −14.4 | −23.4 | −15.7 | 149.1 | 11.7 |
| SSPs_245_2070 | 508,792 | 19,319 | 17,182 | 19,642 | 30,065 | 86,208 |
| Rate of change (%) | −1.4 | −26.6 | −33.9 | −18.3 | 156.4 | 8.5 |
| SSPs_585_2070 | 504,857 | 21,034 | 19,829 | 21,006 | 28,275 | 90,143 |
| Rate of change (%) | −2.14 | −18.07 | −19.56 | −11.62 | 150.24 | 12.94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haq, S.M.; Waheed, M.; Ahmad, R.; Bussmann, R.W.; Arshad, F.; Khan, A.M.; Casini, R.; Alataway, A.; Dewidar, A.Z.; Elansary, H.O. Climate Change and Human Activities, the Significant Dynamic Drivers of Himalayan Goral Distribution (Naemorhedus goral). Biology 2023, 12, 610. https://doi.org/10.3390/biology12040610
Haq SM, Waheed M, Ahmad R, Bussmann RW, Arshad F, Khan AM, Casini R, Alataway A, Dewidar AZ, Elansary HO. Climate Change and Human Activities, the Significant Dynamic Drivers of Himalayan Goral Distribution (Naemorhedus goral). Biology. 2023; 12(4):610. https://doi.org/10.3390/biology12040610
Chicago/Turabian StyleHaq, Shiekh Marifatul, Muhammad Waheed, Riyaz Ahmad, Rainer W. Bussmann, Fahim Arshad, Arshad Mahmood Khan, Ryan Casini, Abed Alataway, Ahmed Z. Dewidar, and Hosam O. Elansary. 2023. "Climate Change and Human Activities, the Significant Dynamic Drivers of Himalayan Goral Distribution (Naemorhedus goral)" Biology 12, no. 4: 610. https://doi.org/10.3390/biology12040610