
Transcriptome Analysis Reveals That C17 Mycosubtilin Antagonizes Verticillium dahliae by Interfering with Multiple Functional Pathways of Fungi
 , , and
, , and 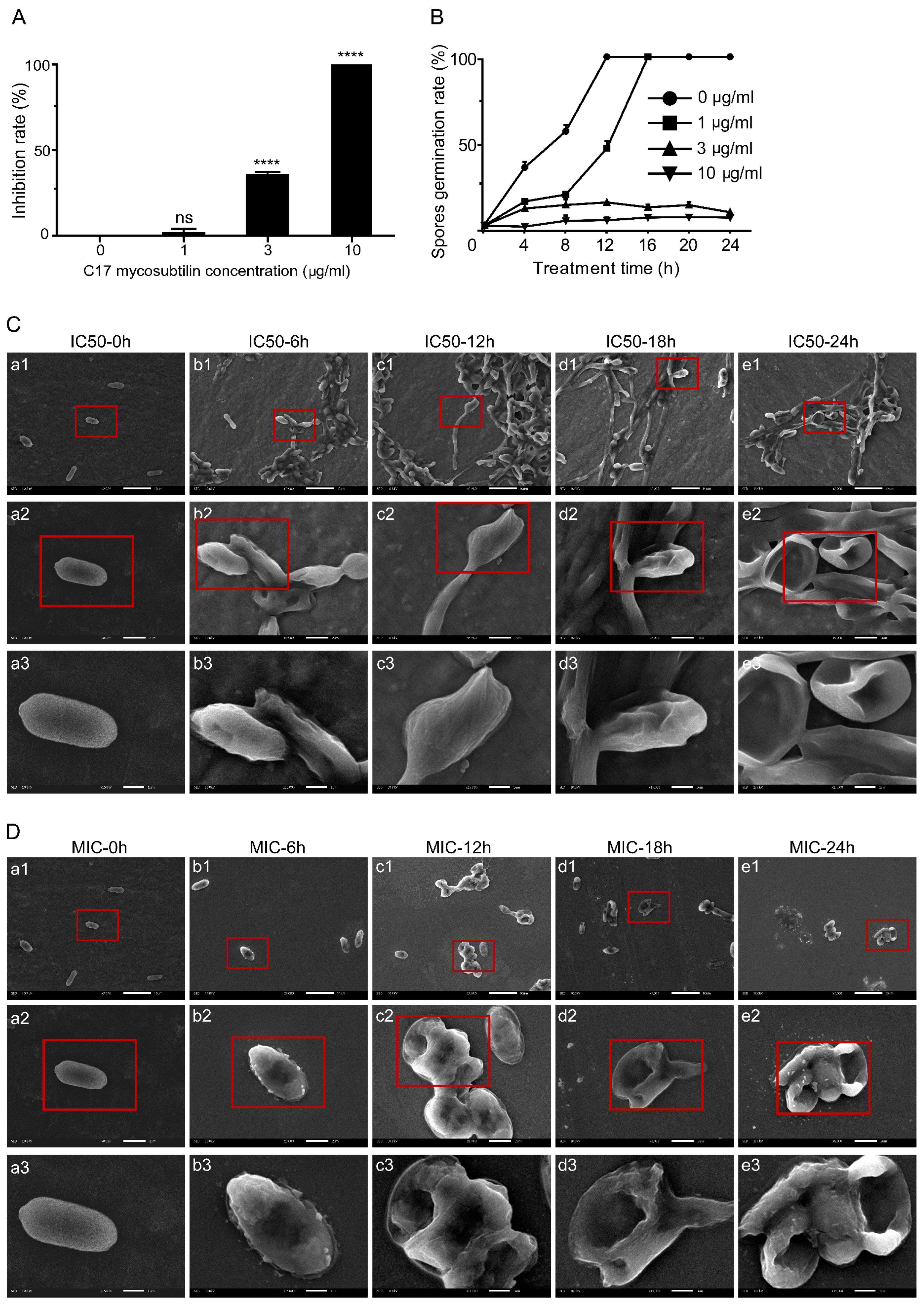{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Purification of C17 Mycosubtilin
2.3. Effects of C17 Mycosubtilin on Growth and Spore Germination of Vd 991
2.4. Observation of Vd 991 Micromorphology by Scanning Electron Microscopy (SEM) and Transmission Electron Microscopy (TEM)
2.5. ANNEXIN V-FITC/PI Staining to Detect the Lethal Effect of C17 Mycosubtilin on Conidia of Vd 991
2.6. RNA Sequencing and Data Analysis
2.7. qRT–PCR
2.8. Statistical Analysis
3. Results and Discussion
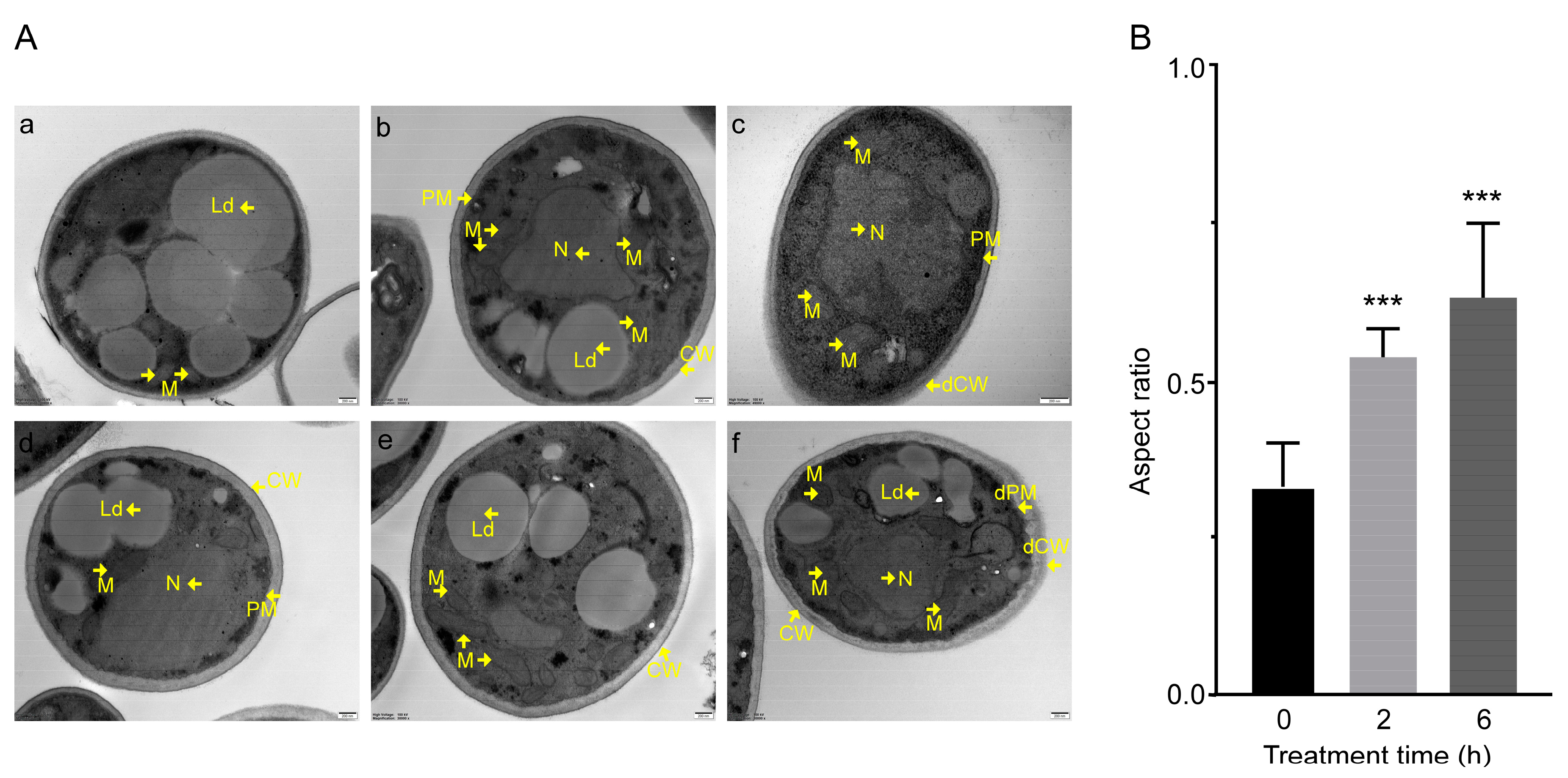
3.1. C17 Mycosubtilin Inhibits the Growth and Conidial Germination of Vd 991 and Affects Its Microscopic Morphology and Organelle Morphological Structure
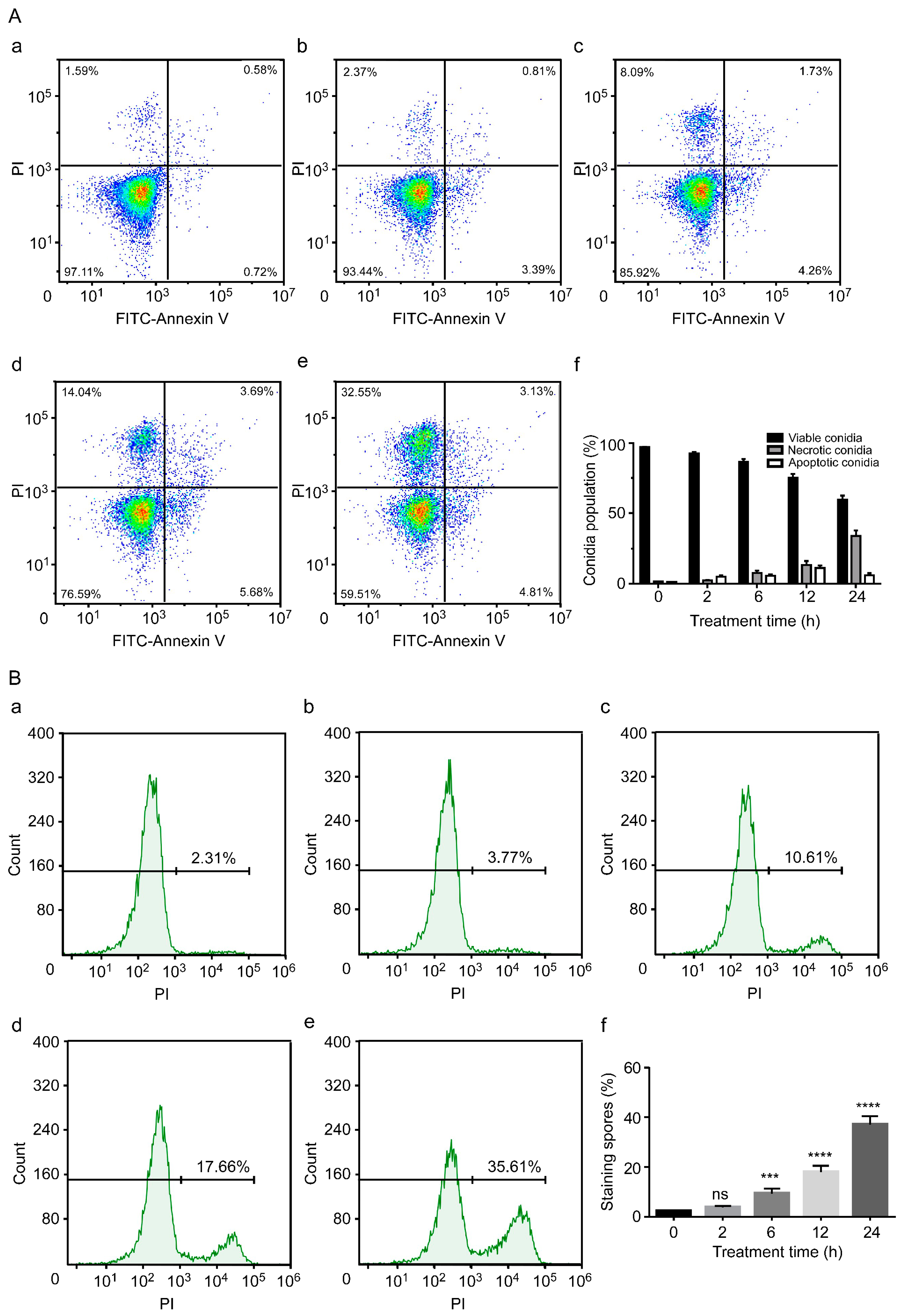
3.2. C17 Mycosubtilin Induces Conidial Necrosis of Vd 991
3.3. Overall Effects of C17 Mycosubtilin on the Gene Transcription of Vd 991
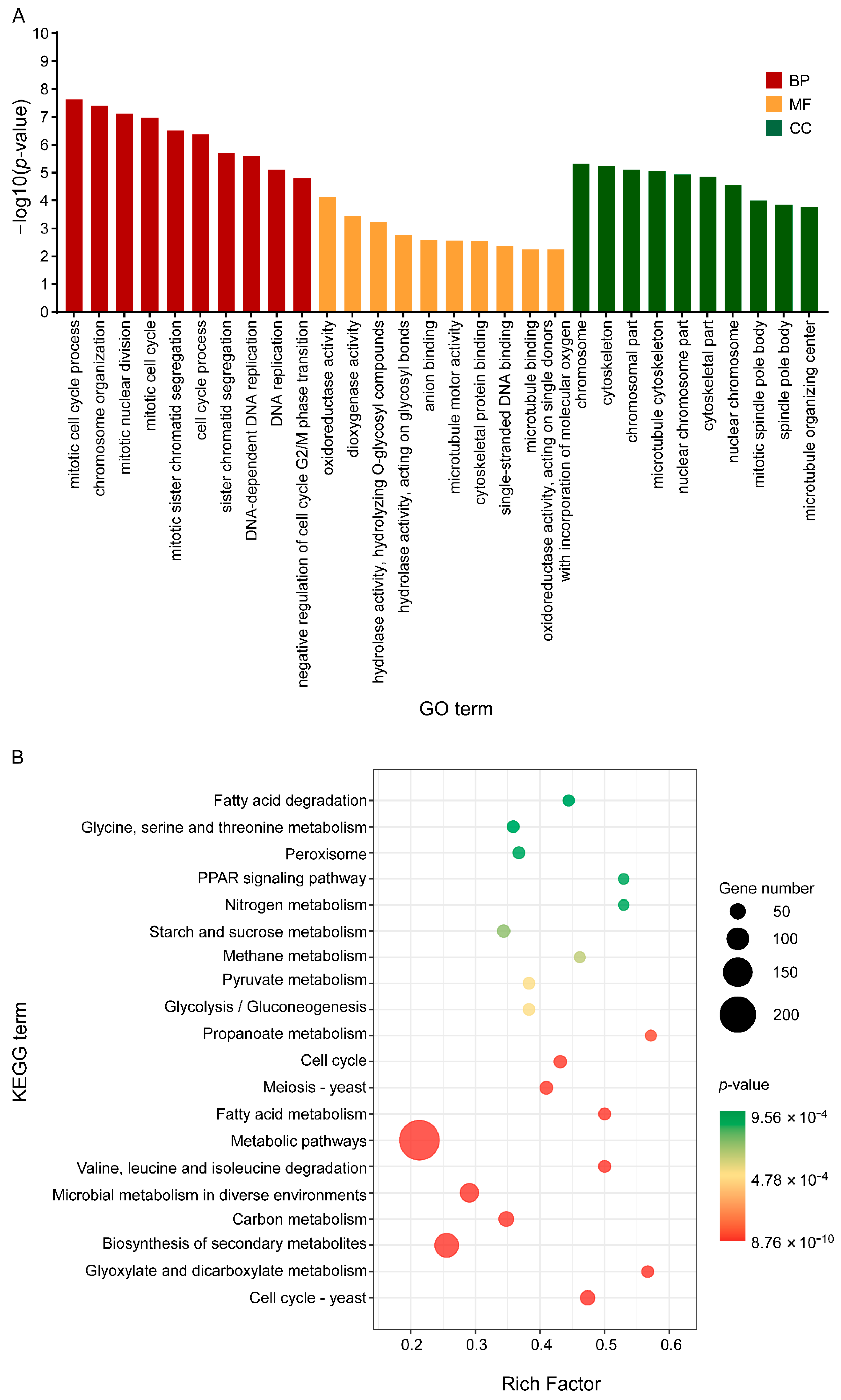
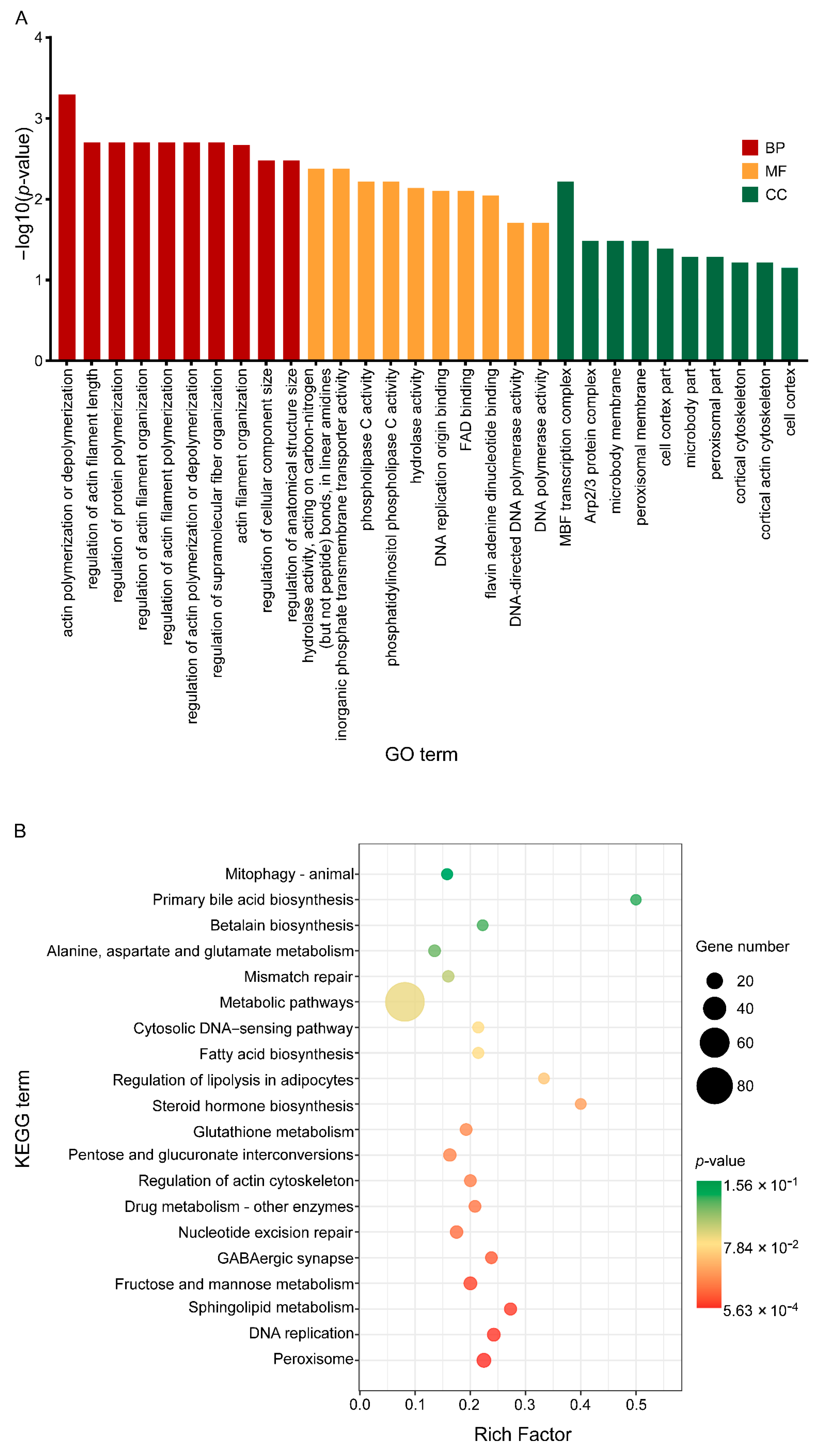
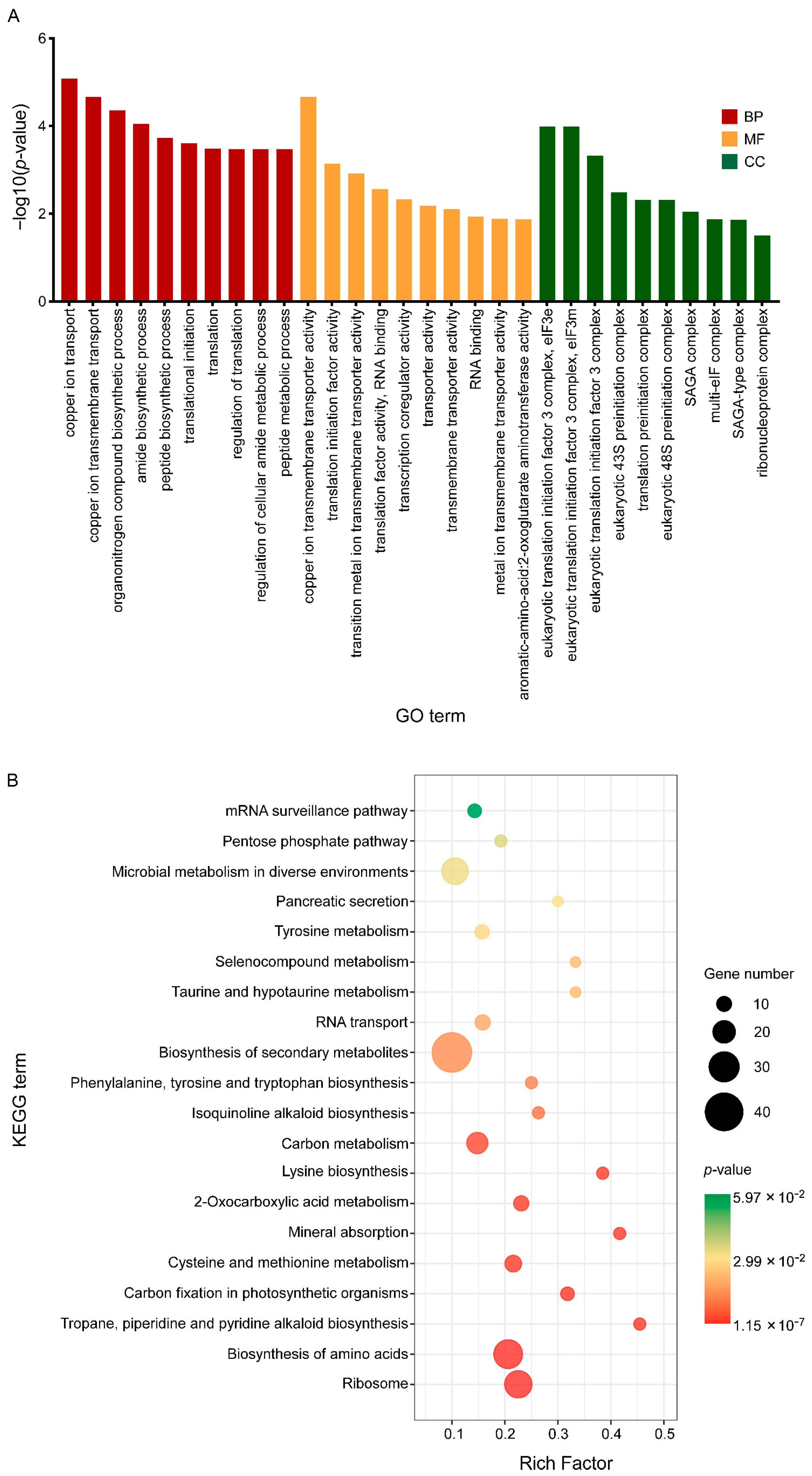
3.4. C17 Mycosubtilin Treatment Interferes with Normal Function of Multiple Functional Units and Metabolic Pathways of Vd
3.5. C17 Mycosubtilin Treatment Disrupts the Structure and Function of Cell Walls and Plasma Membrane of Vd
3.6. C17 Mycosubtilin Treatment Inhibits DNA Replication of Vd and Blocks Its Cell Cycle Process
3.7. C17 Mycosubtilin Treatment Inhibits the Translation Process of Vd
3.8. C17 Mycosubtilin Treatment Affects the Hydrolase Activity of Vd
3.9. C17 Mycosubtilin Treatment Affects the Oxidoreductase System of Vd and Destroys the Process of Material Conversion and Energy Metabolism of Fungi
3.10. C17 Mycosubtilin Treatment Disrupts Energy Metabolism of Vd and Inhibits Energy Production of Fungal Cells
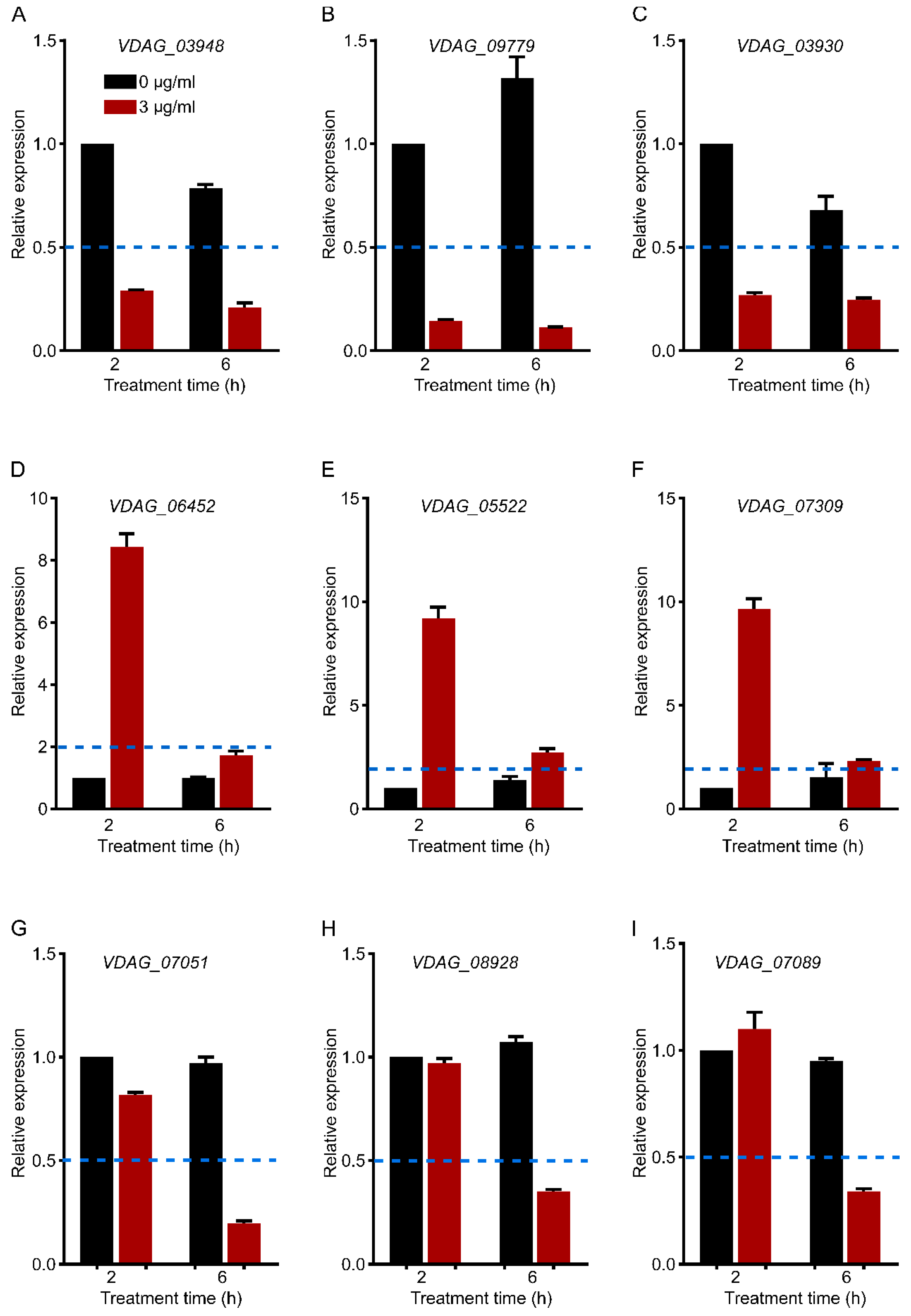
3.11. qRT–PCR Verification of Gene Expression
4. Conclusions
5. Future Recommendations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klosterman, S.J.; Atallah, Z.K.; Vallad, G.E.; Subbarao, K.V. Diversity, pathogenicity, and management of verticillium species. Annu. Rev. Phytopathol. 2009, 47, 39–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Hu, H.L.; Wang, X.N.; Yang, Y.H.; Zhang, C.J.; Zhu, H.Q.; Shi, L.; Tang, C.M.; Zhao, M.W. Dynamic infection of Verticillium dahliae in upland cotton. Plant Biol. 2020, 22, 90–105. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.G.; Subbarao, K.V. Host range specificity in Verticillium dahliae. Phytopathology 1999, 89, 1218–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fradin, E.F.; Thomma, B.P. Physiology and molecular aspects of verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Montes-Osuna, N.; Mercado-Blanco, J. Verticillium wilt of olive and its control: What did we learn during the last decade? Plants 2020, 9, 735. [Google Scholar] [CrossRef]
- Zhang, J.F.; Sanogo, S.; Flynn, R.; Baral, J.B.; Bajaj, S.; Hughs, S.E.; Percy, R.G. Germplasm evaluation and transfer of verticillium wilt resistance from Pima (Gossypium barbadense) to Upland cotton (G. hirsutum). Euphytica 2012, 187, 147–160. [Google Scholar] [CrossRef]
- Deketelaere, S.; Tyvaert, L.; Franca, S.C.; Hofte, M. Desirable traits of a good biocontrol agent against verticillium wilt. Front. Microbiol. 2017, 8, 1186. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Shao, D.; Jiang, C.; Shi, J.; Li, Q.; Huang, Q.; Rajoka, M.S.R.; Yang, H.; Jin, M. Biological activity of lipopeptides from Bacillus. Appl. Microbiol. Biotechnol. 2017, 101, 5951–5960. [Google Scholar] [CrossRef]
- Gan, P.; Jin, D.; Zhao, X.Y.; Gao, Z.Q.; Wang, S.Y.; Du, P.; Qi, G.F. Bacillus-produced surfactin attenuates chronic inflammation in atherosclerotic lesions of ApoE(−/−) mice. Int. Immunopharmacol. 2016, 35, 226–234. [Google Scholar] [CrossRef]
- Duarte, C.; Gudina, E.J.; Lima, C.F.; Rodrigues, L.R. Effects of biosurfactants on the viability and proliferation of human breast cancer cells. Amb. Express 2014, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Jaramillo, L.M.; Aranda, F.J.; Teruel, J.A.; Villegas-Escobar, V.; Ortiz, A. Antimycotic activity of fengycin C biosurfactant and its interaction with phosphatidylcholine model membranes. Colloids Surf. B 2017, 156, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.R.; Park, J.S.; Jung, W.J. Antifungal evaluation of fengycin isoforms isolated from Bacillus amyloliquefaciens PPL against Fusarium oxysporum f. sp. lycopersici. Microb. Pathog. 2020, 149, 104509. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.N.; Lu, J.; Sun, J.; Zhu, X.Y.; Zhou, L.B.; Lu, Z.X.; Lu, Y.J. C16-Fengycin A affect the growth of Candida albicans by destroying its cell wall and accumulating reactive oxygen species. Appl. Microbiol. Biotechnol. 2019, 103, 8963–8975. [Google Scholar] [CrossRef] [PubMed]
- Besson, F.; Peypoux, F.; Michel, G.; Delcambe, L. Characterization of iturin A in antibiotics from various strains of Bacillus subtilis. J. Antibiot. 1976, 29, 1043–1049. [Google Scholar] [CrossRef]
- Park, J.K.; Hasumi, K.; Endo, A. Inhibition of the binding of oxidized low-density-lipoprotein to the macrophages by iturin c-related compounds. J. Antibiot. 1995, 48, 226–232. [Google Scholar] [CrossRef] [Green Version]
- Besson, F.; Michel, G. Isolation and characterization of new Iturins—Iturin D and Iturin E. J. Antibiot. 1987, 40, 437–442. [Google Scholar] [CrossRef] [Green Version]
- Peypoux, F.; Pommier, M.T.; Marion, D.; Ptak, M.; Das, B.C.; Michel, G. Revised structure of mycosubtilin, a peptidolipid antibiotic from Bacillus Subtilis. J. Antibiot. 1986, 39, 636–641. [Google Scholar] [CrossRef] [Green Version]
- Besson, F.; Peypoux, F.; Michel, G.; Delcambe, L. Structure de la bacillomycine L, antibiotique de Bacillus subtilis. Eur. J. Biochem. 1977, 77, 61–67. [Google Scholar] [CrossRef]
- Mhammedi, A.; Peypoux, F.; Besson, F.; Michel, G. Bacillomycin F, a new antibiotic of iturin group: Isolation and characterization. J. Antibiot. 1982, 35, 306–311. [Google Scholar] [CrossRef] [Green Version]
- Peypoux, F.; Besson, F.; Michel, G.; Lenzen, C.; Dierickx, L.; Delcambe, L. Characterization of a new antibiotic of iturin group: Bacillomycin D. J. Antibiot. 1980, 33, 1146–1149. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Wang, N.; Hu, J.; Wang, S. Isolation and characterization of a new iturinic lipopeptide, mojavensin A produced by a marine-derived bacterium Bacillus mojavensis B0621A. J. Antibiot. 2012, 65, 317–322. [Google Scholar] [CrossRef] [Green Version]
- Kajimura, Y.; Sugiyama, M.; Kaneda, M. Bacillopeptins, new cyclic lipopeptide antibiotics from Bacillus subtilis Fr 2. J. Antibiot. 1995, 48, 1095–1103. [Google Scholar] [CrossRef]
- Aranda, F.J.; Teruel, J.A.; Ortiz, A. Further aspects on the hemolytic activity of the antibiotic lipopeptide iturin A. BBA Biomembr. 2005, 1713, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.M.; Li, Z.Z.; Shi, Y.H.; Guo, D.; Pang, B.; Chen, X.Q.; Shao, D.Y.; Liu, Y.L.; Shi, J.L. Bacillus subtilis inhibits Aspergillus carbonarius by producing iturin A, which disturbs the transport, energy metabolism, and osmotic pressure of fungal cells as revealed by transcriptomics analysis. Int. J. Food Microbiol. 2020, 330, 108783. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.W.; Guo, X.J.; Qiao, X.L.; Zhang, X.C.; Chen, X.M.; Zhang, D.D. Activity of fengycin and iturin A isolated from Bacillus subtilis Z–14 on Gaeumannomyces graminis var. tritici and soil microbial diversity. Front. Microbiol. 2021, 12, 682437. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.X.; Pang, X.Y.; Liu, H.W.; Lin, F.X.; Lu, F.X.; Bie, X.M.; Lu, Z.X.; Lu, Y.J. Iturin A induces resistance and improves the quality and safety of harvested cherry tomato. Molecules 2021, 26, 6905. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.T.; Kim, S.E.; Lee, W.J.; Fumei, Z.; Cho, M.S.; Moon, J.S.; Oh, H.W.; Park, H.Y.; Kim, S.U. Isolation and characterization of a high iturin yielding Bacillus velezensis UV mutant with improved antifungal activity. PLoS ONE 2020, 15, e0234177. [Google Scholar] [CrossRef]
- Kalai-Grami, L.; Karkouch, I.; Naili, O.; Ben Slimene, I.; Elkahoui, S.; Ben Zekri, R.; Touati, I.; Mnari-Hattab, M.; Hajlaoui, M.R.; Limam, F. Production and identification of iturin A lipopeptide from Bacillus methyltrophicus TEB1 for control of Phoma tracheiphila. J. Basic Microb. 2016, 56, 864–871. [Google Scholar] [CrossRef]
- Zhou, L.F.; Zhao, X.X.; Li, M.X.; Lu, Y.; Ai, C.Y.; Jiang, C.M.; Liu, Y.L.; Pan, Z.L.; Shi, J.L. Antifungal activity of silver nanoparticles synthesized by iturin against Candida albicans in vitro and in vivo. Appl. Microbiol. Biotechnol. 2021, 105, 3759–3770. [Google Scholar] [CrossRef]
- Gong, A.D.; Li, H.P.; Yuan, Q.S.; Song, X.S.; Yao, W.; He, W.J.; Zhang, J.B.; Liao, Y.C. Antagonistic mechanism of iturin A and plipastatin A from Bacillus amyloliquefaciens S76-3 from wheat spikes against Fusarium graminearum. PLoS ONE 2015, 10, e0116871. [Google Scholar] [CrossRef] [Green Version]
- Leclere, V.; Bechet, M.; Adam, A.; Guez, J.S.; Wathelet, B.; Ongena, M.; Thonart, P.; Gancel, F.; Chollet-Imbert, M.; Jacques, P. Mycosubtilin overproduction by Bacillus subtilis BBG100 enhances the organism’s antagonistic and biocontrol activities. Appl. Environ. Microbiol. 2005, 71, 4577–4584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasir, M.N.; Besson, F. Interactions of the antifungal mycosubtilin with ergosterol-containing interfacial monolayers. BBA Biomembr. 2012, 1818, 1302–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasir, M.N.; Besson, F. Specific interactions of mycosubtilin with cholesterol-containing artificial membranes. Langmuir 2011, 27, 10785–10792. [Google Scholar] [CrossRef] [PubMed]
- Deravel, J.; Lemiere, S.; Coutte, F.; Krier, F.; Van Hese, N.; Bechet, M.; Sourdeau, N.; Hofte, M.; Lepretre, A.; Jacques, P. Mycosubtilin and surfactin are efficient, low ecotoxicity molecules for the biocontrol of lettuce downy mildew. Appl. Microbiol. Biotechnol. 2014, 98, 6255–6264. [Google Scholar] [CrossRef]
- Khong, N.G.; Randoux, B.; Tayeh, C.; Coutte, F.; Bourdon, N.; Tisserant, B.; Laruelle, F.; Jacques, P.; Reignault, P. Induction of resistance in wheat against powdery mildew by bacterial cyclic lipopeptides. Commun. Agric. Appl. Biol. Sci. 2012, 77, 39–51. [Google Scholar]
- Mejri, S.; Siah, A.; Coutte, F.; Magnin-Robert, M.; Randoux, B.; Tisserant, B.; Krier, F.; Jacques, P.; Reignault, P.; Halama, P. Biocontrol of the wheat pathogen Zymoseptoria tritici using cyclic lipopeptides from Bacillus subtilis. Environ. Sci. Pollut. Res. Int. 2018, 25, 29822–29833. [Google Scholar] [CrossRef]
- Zhao, J.; Zeng, W.; Li, Y.; Ge, F.; Du, Y.; Yuan, L.; Zhao, Q.; Wang, M.; Xie, H.; Bai, R.; et al. Isolation and identification of antagonistic spore strains of Verticillium dahliae and study on their antibacterial characteristics. J. Beijing Norm. Univ. 2017, 53, 294–300. [Google Scholar]
- Lin, R.; Zhang, Q.; Yin, L.; Zhang, Y.; Yang, Q.; Liu, K.; Wang, Y.; Han, S.; Zhao, H.; Zhao, H. Isolation and characterization of a mycosubtilin homologue antagonizing Verticillium dahliae produced by Bacillus subtilis strain Z15. PLoS ONE 2022, 17, e0269861. [Google Scholar] [CrossRef]
- Cheng, X.X.; Zhao, L.H.; Klosterman, S.J.; Feng, H.J.; Feng, Z.L.; Wei, F.; Shi, Y.Q.; Li, Z.F.; Zhu, H.Q. The endochitinase VDECH from Verticillium dahliae inhibits spore germination and activates plant defense responses. Plant Sci. 2017, 259, 12–23. [Google Scholar] [CrossRef]
- Wang, J.Y.; Cai, Y.; Gou, J.Y.; Mao, Y.B.; Xu, Y.H.; Jiang, W.H.; Chen, X.Y. VdNEP, an elicitor from Verticillium dahliae, induces cotton plant wilting. Appl. Environ. Microbiol. 2004, 70, 4989–4995. [Google Scholar] [CrossRef] [Green Version]
- Raahave, D. Paper disk-agar diffusion assay of penicillin in the presence of streptomycin. Antimicrob. Agents Chemother. 1974, 6, 603–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Zhou, B.J.; Li, G.Y.; Jia, P.S.; Li, H.; Zhao, Y.L.; Zhao, P.; Xia, G.X.; Guo, H.S. A glutamic acid-rich protein identified in Verticillium dahliae from an insertional mutagenesis affects microsclerotial formation and pathogenicity. PLoS ONE 2010, 5, e15319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abuduaini, X.; Aili, A.; Lin, R.; Song, G.; Zhao, H. The lethal effect of Bacillus subtilis Z15 secondary metabolites on Verticillium dahliae. Nat. Prod. Commun. 2021, 16, 1934578X2098672. [Google Scholar]
- Bo, T.; Liu, M.; Zhong, C.; Zhang, Q.; Su, Q.Z.; Tan, Z.L.; Han, P.P.; Jia, S.R. Metabolomic analysis of antimicrobial mechanisms of epsilon-poly-L-lysine on Saccharomyces cerevisiae. J. Agric. Food Chem. 2014, 62, 4454–4465. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Landmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Raithel, S.; Johnson, L.; Galliart, M.; Brown, S.; Shelton, J.; Herndon, N.; Bello, N.M. Inferential considerations for low-count RNA-seq transcripts: A case study on the dominant prairie grass Andropogon gerardii. BMC Genom. 2016, 17, 140. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Lu, X.; Tian, J.; Wen, H.; Jiang, M.; Zhong, S. Microcystin-LR-regulated transcriptome dynamics in ZFL cells. Aquat. Toxicol. 2019, 212, 222–232. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, T.; He, D.; Li, X.Z.; Wu, H.; Liu, W.; Gao, X. Functions of lipopeptides bacillomycin D and fengycin in antagonism of Bacillus amyloliquefaciens C06 towards Monilinia fructicola. J. Mol. Microbiol. Biotechnol. 2011, 20, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.N.; Li, Y.S.; Hu, D.K.; Tian, H.; Jiang, J.Z.; Wang, Y.; Yan, X.J. Synthesis and fungicidal activity of novel 2,5-disubstituted-1,3,4- thiadiazole derivatives containing 5-phenyl-2-furan. Sci. Rep. 2016, 6, 20204. [Google Scholar] [CrossRef] [Green Version]
- Huiskonen, J.T. Image processing for cryogenic transmission electron microscopy of symmetry-mismatched complexes. Biosci. Rep. 2018, 38, BSR20170203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihalache, G.; Balaes, T.; Gostin, I.; Stefan, M.; Coutte, F.; Krier, F. Lipopeptides produced by Bacillus subtilis as new biocontrol products against fusariosis in ornamental plants. Environ. Sci. Pollut. Res. Int. 2017, 25, 29784–29793. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, C.; Liang, J.; Wu, L.; Gao, W.; Jiang, J. Iturin a extracted from Bacillus subtilis WL-2 affects phytophthora infestans via cell structure disruption, oxidative stress, and energy supply dysfunction. Front. Microbiol. 2020, 11, 536083. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.B.; Xu, X.G.; Lei, S.Z.; Shao, D.Y.; Jiang, C.M.; Shi, J.L.; Zhang, Y.W.; Liu, L.; Lei, S.Z.; Sun, H.; et al. Iturin A-like lipopeptides from Bacillus subtilis trigger apoptosis, paraptosis, and autophagy in Caco-2 cells. J. Cell. Physiol. 2019, 234, 6414–6427. [Google Scholar] [CrossRef] [PubMed]
- Li, S.F.; Zhang, S.B.; Lv, Y.Y.; Zhai, H.C.; Hu, Y.S.; Cai, J.P. Transcriptome analysis reveals the underlying mechanism of heptanal against Aspergillus flavus spore germination. Appl. Microbiol. Biotechnol. 2022, 106, 1241–1255. [Google Scholar] [CrossRef]
- Yu, C.J.; Liu, X.; Zhang, X.Y.; Zhang, M.X.; Gu, Y.Y.; Ali, Q.; Mohamed, M.S.R.; Xu, J.H.; Shi, J.R.; Gao, X.W.; et al. Mycosubtilin produced by Bacillus subtilis ATCC6633 inhibits growth and mycotoxin biosynthesis of Fusarium graminearum and Fusarium verticillioides. Toxins 2021, 13, 791. [Google Scholar] [CrossRef]
- Gozalbo, D.; Roig, P.; Villamon Ribate, E.; Gil, M. Candida and candidiasis: The cell wall as a potential molecular target for antifungal therapy. Infect. Disord. Drug Targets 2004, 4, 117–135. [Google Scholar] [CrossRef]
- Hoyer, L.L. The ALS gene family of Candida albicans. Trends Microbiol. 2001, 9, 176–180. [Google Scholar] [CrossRef]
- Hancock, J.F. GPI-anchor synthesis: Ras takes charge. Dev. Cell 2004, 6, 743–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Q.; Yang, Y.; Yuan, Q.; Shi, G.; Wu, L.; Lou, Z.; Huo, R.; Wu, H.; Borriss, R.; Gao, X. Bacillomycin D produced by Bacillus amyloliquefaciens is involved in the antagonistic interaction with the plant-pathogenic fungus Fusarium graminearum. Appl. Environ. Microbiol. 2017, 83, e01075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regenberg, B.; During-Olsen, L.; Kielland-Brandt, M.C.; Holmberg, S. Substrate specificity and gene expression of the amino-acid permeases in Saccharomyces cerevisiae. Curr. Genet. 1999, 36, 317–328. [Google Scholar] [CrossRef]
- Tkach, J.M.; Yimit, A.; Lee, A.Y.; Riffle, M.; Costanzo, M.; Jaschob, D.; Hendry, J.A.; Ou, J.; Moffat, J.; Boone, C.; et al. Dissecting DNA damage response pathways by analysing protein localization and abundance changes during DNA replication stress. Nat. Cell Biol. 2012, 14, 966–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzurendova, S.; Zimmermann, B.; Kohler, A.; Reitzel, K.; Nielsen, U.G.; Dupuy-Galet, B.X.; Leivers, S.; Horn, S.J.; Shapaval, V. Calcium affects polyphosphate and lipid accumulation in Mucoromycota fungi. J. Fungi 2021, 7, 300. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Peiter, E. Calcium transport proteins in fungi: The phylogenetic diversity of their relevance for growth, virulence, and stress resistance. Front. Microbiol. 2019, 10, 3100. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Weisenhorn, E.; MacDiarmid, C.W.; Andreini, C.; Bucci, M.; Taggart, J.; Banci, L.; Russell, J.; Coon, J.J.; Eide, D.J. The cellular economy of the Saccharomyces cerevisiae zinc proteome. Metallomics 2018, 10, 1755–1776. [Google Scholar] [CrossRef]
- Ehrensberger, K.M.; Mason, C.; Corkins, M.E.; Anderson, C.; Dutrow, N.; Cairns, B.R.; Dalley, B.; Milash, B.; Bird, A.J. Zinc-dependent regulation of the Adh1 antisense transcript in fission yeast. J. Biol. Chem. 2013, 288, 759–769. [Google Scholar] [CrossRef] [Green Version]
- Maras, B.; Greenblatt, H.M.; Shoham, G.; Spungin-Bialik, A.; Blumberg, S.; Barra, D. Aminopeptidase from Streptomyces griseus: Primary structure and comparison with other zinc-containing aminopeptidases. Eur. J. Biochem. 1996, 236, 843–846. [Google Scholar] [CrossRef]
- Koval, T.; Dohnalek, J. Characteristics and application of S1-P1 nucleases in biotechnology and medicine. Biotechnol. Adv. 2018, 36, 603–612. [Google Scholar] [CrossRef]
- Rae, T.D.; Torres, A.S.; Pufahl, R.A.; O’Halloran, T.V. Mechanism of Cu,Zn-superoxide dismutase activation by the human metallochaperone hCCS. J. Biol. Chem. 2001, 276, 5166–5176. [Google Scholar] [CrossRef] [Green Version]
- Kanteev, M.; Goldfeder, M.; Fishman, A. Structure function correlations in tyrosinases. Protein Sci. 2015, 24, 1360–1369. [Google Scholar] [CrossRef] [Green Version]
- Braunschmid, V.; Fuerst, S.; Perz, V.; Zitzenbacher, S.; Hoyo, J.; Fernandez-Sanchez, C.; Tzanov, T.; Steinkellner, G.; Gruber, K.; Nyanhongo, G.S.; et al. A fungal ascorbate oxidase with unexpected laccase activity. Int. J. Mol. Sci. 2020, 21, 5754. [Google Scholar] [CrossRef] [PubMed]
- Klotz, L.O.; Kroncke, K.D.; Buchczyk, D.P.; Sies, H. Role of copper, zinc, selenium and tellurium in the cellular defense against oxidative and nitrosative stress. J. Nutr. 2003, 133, 1448S–1451S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinett, N.G.; Culbertson, E.M.; Peterson, R.L.; Sanchez, H.; Andes, D.R.; Nett, J.E.; Culotta, V.C. Exploiting the vulnerable active site of a copper-only superoxide dismutase to disrupt fungal pathogenesis. J. Biol. Chem. 2019, 294, 2700–2713. [Google Scholar] [CrossRef] [Green Version]
- Schatzman, S.S.; Peterson, R.L.; Teka, M.; He, B.; Cabelli, D.E.; Cormack, B.P.; Culotta, V.C. Copper-only superoxide dismutase enzymes and iron starvation stress in Candida fungal pathogens. J. Biol. Chem. 2020, 295, 570–583. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.P.; Dutta, A. DNA replication in eukaryotic cells. Annu. Rev. Biochem. 2002, 71, 333–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgers, P.M.J. Eukaryotic DNA polymerase δ. In Encyclopedia of Biological Chemistry, 2nd ed.; Lennarz, W.J., Lane, M.D., Eds.; Academic Press: Waltham, MA, USA, 2013; Volume 3, pp. 245–248. [Google Scholar]
- Ernsting, B.R.; Dixon, J.E. The PPS1 gene of Saccharomyces cerevisiae codes for a dual specificity protein phosphatase with a role in the DNA synthesis phase of the cell cycle. J. Biol. Chem. 1997, 272, 9332–9343. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Yan, L.; Xu, X.; Jiang, C.; Shi, J.; Zhang, Y.; Liu, L.; Lei, S.; Shao, D.; Huang, Q. Potential of Bacillus subtilis lipopeptides in anti-cancer I: Induction of apoptosis and paraptosis and inhibition of autophagy in K562 cells. AMB Express 2018, 8, 78. [Google Scholar] [CrossRef]
- Dey, G.; Bharti, R.; Dhanarajan, G.; Das, S.; Dey, K.K.; Kumar, B.N.; Sen, R.; Mandal, M. Marine lipopeptide iturin A inhibits Akt mediated GSK3beta and FoxO3a signaling and triggers apoptosis in breast cancer. Sci. Rep. 2015, 5, 10316. [Google Scholar] [CrossRef] [Green Version]
- Mitterer, V.; Murat, G.; Rety, S.; Blaud, M.; Delbos, L.; Stanborough, T.; Bergler, H.; Leulliot, N.; Kressler, D.; Pertschy, B. Sequential domain assembly of ribosomal protein S3 drives 40S subunit maturation. Nat. Commun. 2016, 7, 10336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, M.; Kong, C.J.; Yoshida, H.; Nakamura, T.; Wada, A.; Shimoda, C.; Kaneda, Y. Ribosomal proteins S0 and S21 are involved in the stability of 18S rRNA in fission yeast, Schizosaccharomyces pombe. Biochem. Biophys. Res. Commun. 2003, 311, 942–947. [Google Scholar] [CrossRef] [PubMed]
- van Beekvelt, C.A.; de Graaff-Vincent, M.; Faber, A.W.; van’t Riet, J.; Venema, J.; Raue, H.A. All three functional domains of the large ribosomal subunit protein L25 are required for both early and late pre-rRNA processing steps in Saccharomyces cerevisiae. Nucleic Acids Res. 2001, 29, 5001–5008. [Google Scholar] [CrossRef] [PubMed]
- Gamalinda, M.; Woolford, J.L., Jr. Deletion of L4 domains reveals insights into the importance of ribosomal protein extensions in eukaryotic ribosome assembly. RNA 2014, 20, 1725–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Ryu, J.H.; Han, S.J.; Choi, K.H.; Nam, K.B.; Jang, I.H.; Lemaitre, B.; Brey, P.T.; Lee, W.J. Gram-negative bacteria-binding protein, a pattern recognition receptor for lipopolysaccharide and beta-1,3-glucan that mediates the signaling for the induction of innate immune genes in Drosophila melanogaster cells. J. Biol. Chem. 2000, 275, 32721–32727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Esquer, F.; Rodriguez-Pena, J.M.; Diaz, G.; Rodriguez, E.; Briza, P.; Nombela, C.; Arroyo, J. CRR1, a gene encoding a putative transglycosidase, is required for proper spore wall assembly in Saccharomyces cerevisiae. Microbiology 2004, 150, 3269–3280. [Google Scholar] [CrossRef] [Green Version]
- Arvind, K.; Naraian, R.B. Differential Expression of the Microbial β-1,4-xylanase, and β-1,4-endoglucanase Genes. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; Volume 3, pp. 95–111. [Google Scholar]
- Ahrazem, O.; Gomez-Gomez, L.; Rodrigo, M.J.; Avalos, J.; Limon, M.C. Carotenoid cleavage oxygenases from microbes and photosynthetic organisms: Features and functions. Int. J. Mol. Sci. 2016, 17, 1781. [Google Scholar] [CrossRef] [Green Version]
- Walter, M.H.; Floss, D.S.; Hans, J.; Fester, T.; Strack, D. Apocarotenoid biosynthesis in arbuscular mycorrhizal roots: Contributions from methylerythritol phosphate pathway isogenes and tools for its manipulation. Phytochemistry 2007, 68, 130–138. [Google Scholar] [CrossRef]
- Hayaishi, O. Oxygenases. In Encyclopedia of Biological Chemistry, 2nd ed.; Lennarz, W.J., Lane, M.D., Eds.; Academic Press: Waltham, MA, USA, 2013; Volume 3, pp. 371–374. [Google Scholar]
- Li, D.D.; Chai, D.; Huang, X.W.; Guan, S.X.; Du, J.; Zhang, H.Y.; Sun, Y.; Jiang, Y.Y. Potent In Vitro Synergism of Fluconazole and Osthole against Fluconazole-Resistant Candida albicans. Antimicrob. Agents Chemother. 2017, 61, e00436-17. [Google Scholar] [CrossRef] [Green Version]
- Gottwald, E.M.; Duss, M.; Bugarski, M.; Haenni, D.; Schuh, C.D.; Landau, E.M.; Hall, A.M. The targeted anti-oxidant MitoQ causes mitochondrial swelling and depolarization in kidney tissue. Physiol. Rep. 2018, 6, e13667. [Google Scholar] [CrossRef]
- Cheong, H.; Yorimitsu, T.; Reggiori, F.; Legakis, J.E.; Wang, C.W.; Klionsky, D.J. Atg17 regulates the magnitude of the autophagic response. Mol. Biol. Cell. 2005, 16, 3438–3453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Dong, C.; Shang, Q.; Han, Y.; Li, P. New insights into membrane-active action in plasma membrane of fungal hyphae by the lipopeptide antibiotic bacillomycin L. BBA Biomembr. 2013, 1828, 2230–2237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fickers, P.; Leclere, V.; Guez, J.S.; Bechet, M.; Coucheney, F.; Joris, B.; Jacques, P. Temperature dependence of mycosubtilin homologue production in Bacillus subtilis ATCC6633. Res. Microbiol. 2008, 159, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Peypoux, F.; Besson, F.; Michel, G.; Delcambe, L. Preparation and antibacterial activity upon Micrococcus luteus of derivatives of iturin A, mycosubtilin and bacillomycin L, antibiotics from Bacillus subtilis. J. Antibiot. 1979, 32, 136–140. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Lin, R.; Yang, J.; Zhao, J.; Li, H.; Liu, K.; Xue, X.; Zhao, H.; Han, S.; Zhao, H. Transcriptome Analysis Reveals That C17 Mycosubtilin Antagonizes Verticillium dahliae by Interfering with Multiple Functional Pathways of Fungi. Biology 2023, 12, 513. https://doi.org/10.3390/biology12040513
Zhang Q, Lin R, Yang J, Zhao J, Li H, Liu K, Xue X, Zhao H, Han S, Zhao H. Transcriptome Analysis Reveals That C17 Mycosubtilin Antagonizes Verticillium dahliae by Interfering with Multiple Functional Pathways of Fungi. Biology. 2023; 12(4):513. https://doi.org/10.3390/biology12040513
Chicago/Turabian StyleZhang, Qi, Rongrong Lin, Jun Yang, Jingjing Zhao, Haoran Li, Kai Liu, Xiuhua Xue, Huixin Zhao, Shengcheng Han, and Heping Zhao. 2023. "Transcriptome Analysis Reveals That C17 Mycosubtilin Antagonizes Verticillium dahliae by Interfering with Multiple Functional Pathways of Fungi" Biology 12, no. 4: 513. https://doi.org/10.3390/biology12040513