
Effects of Hydrologic Regime Changes on a Taxonomic and Functional Trait Structure of Earthworm Communities in Mountain Wetlands

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
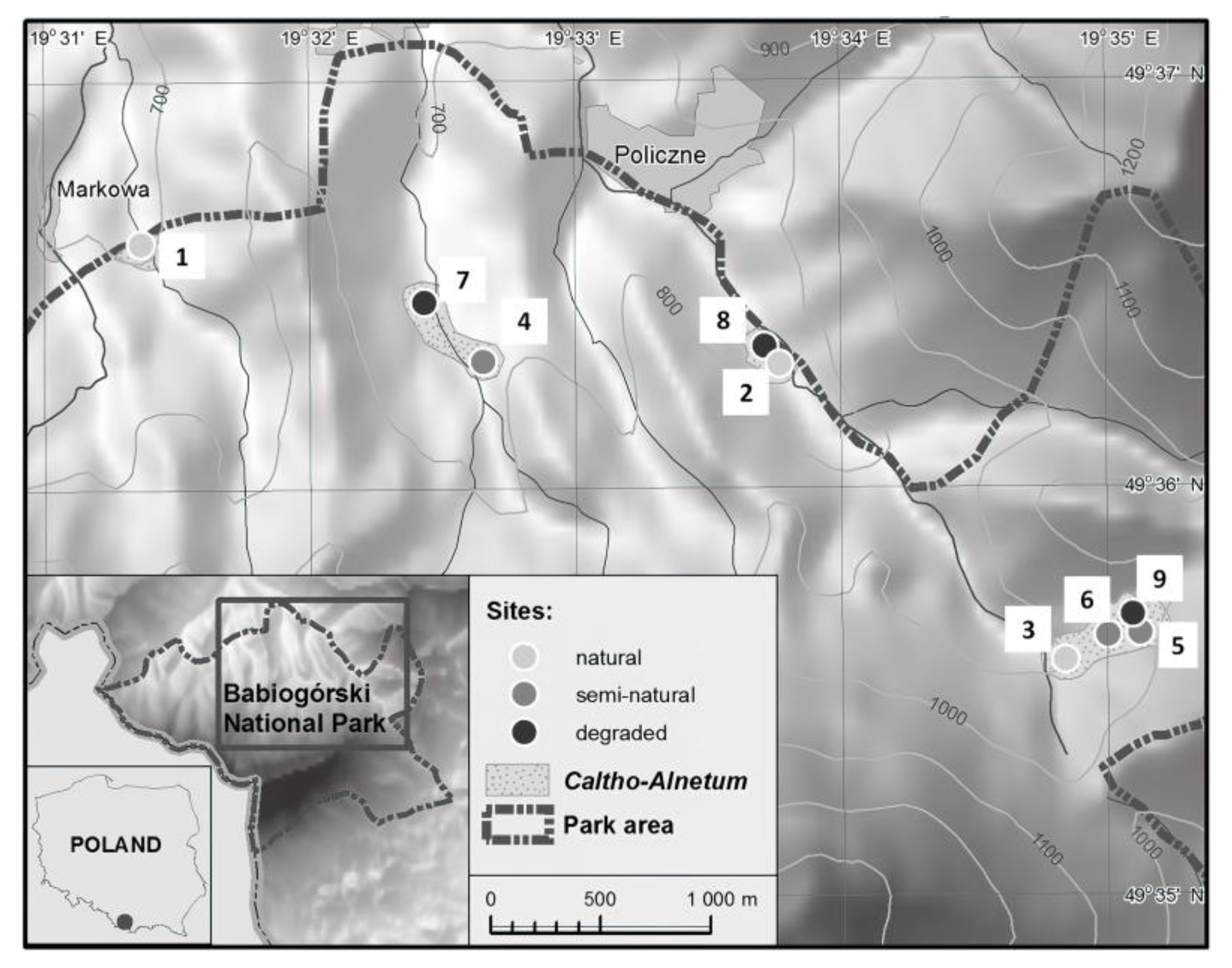
2.1. Study Area
2.2. Sites
2.3. Sampling and Processing
2.4. Functional Traits
2.5. Data Analysis
3. Results
3.1. Soil Properties
3.2. Earthworm Diversity Pattern and Density
3.3. Earthworm Taxonomic Composition Pattern
3.4. Earthworm Trait Composition Pattern
4. Discussion
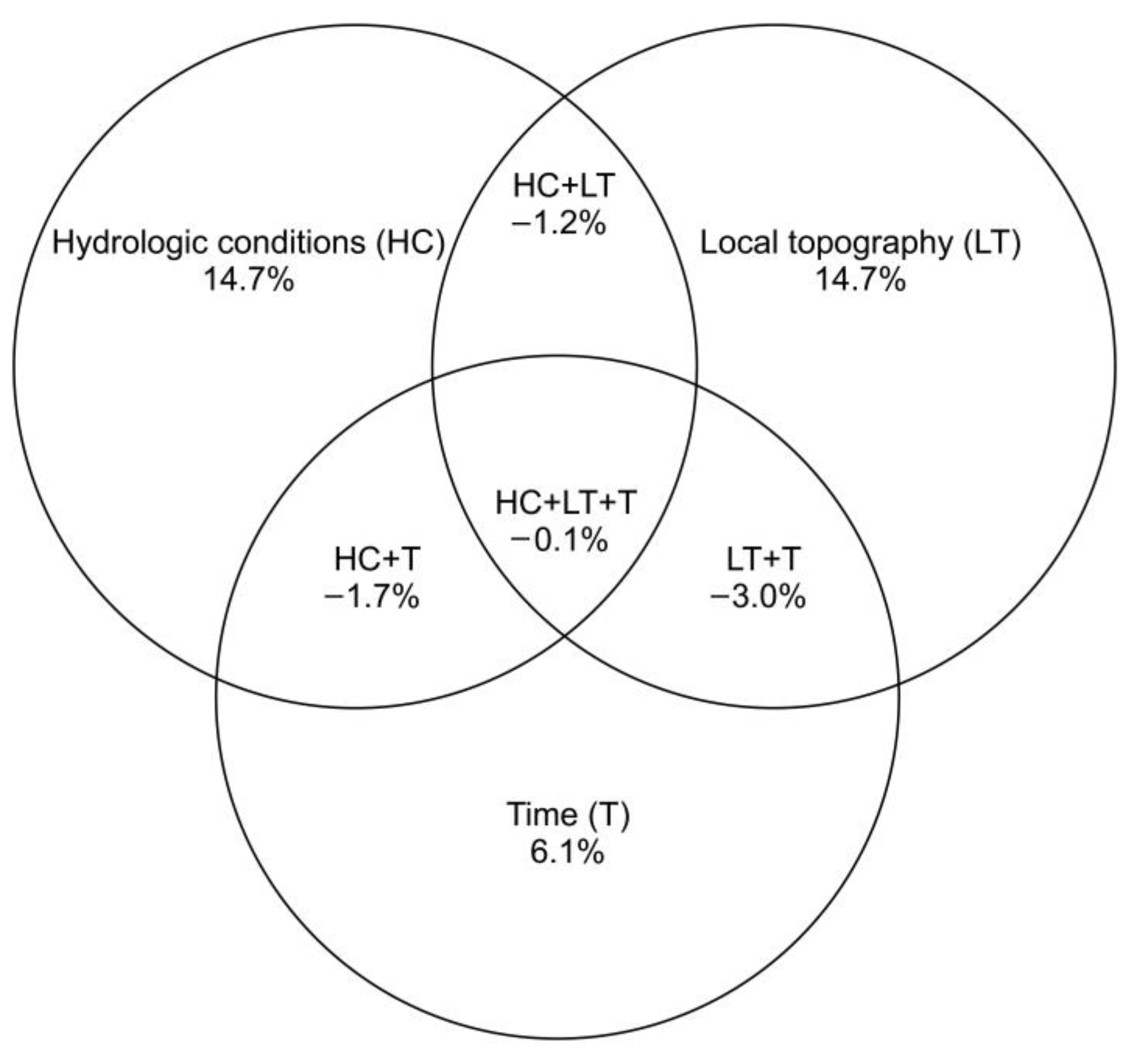
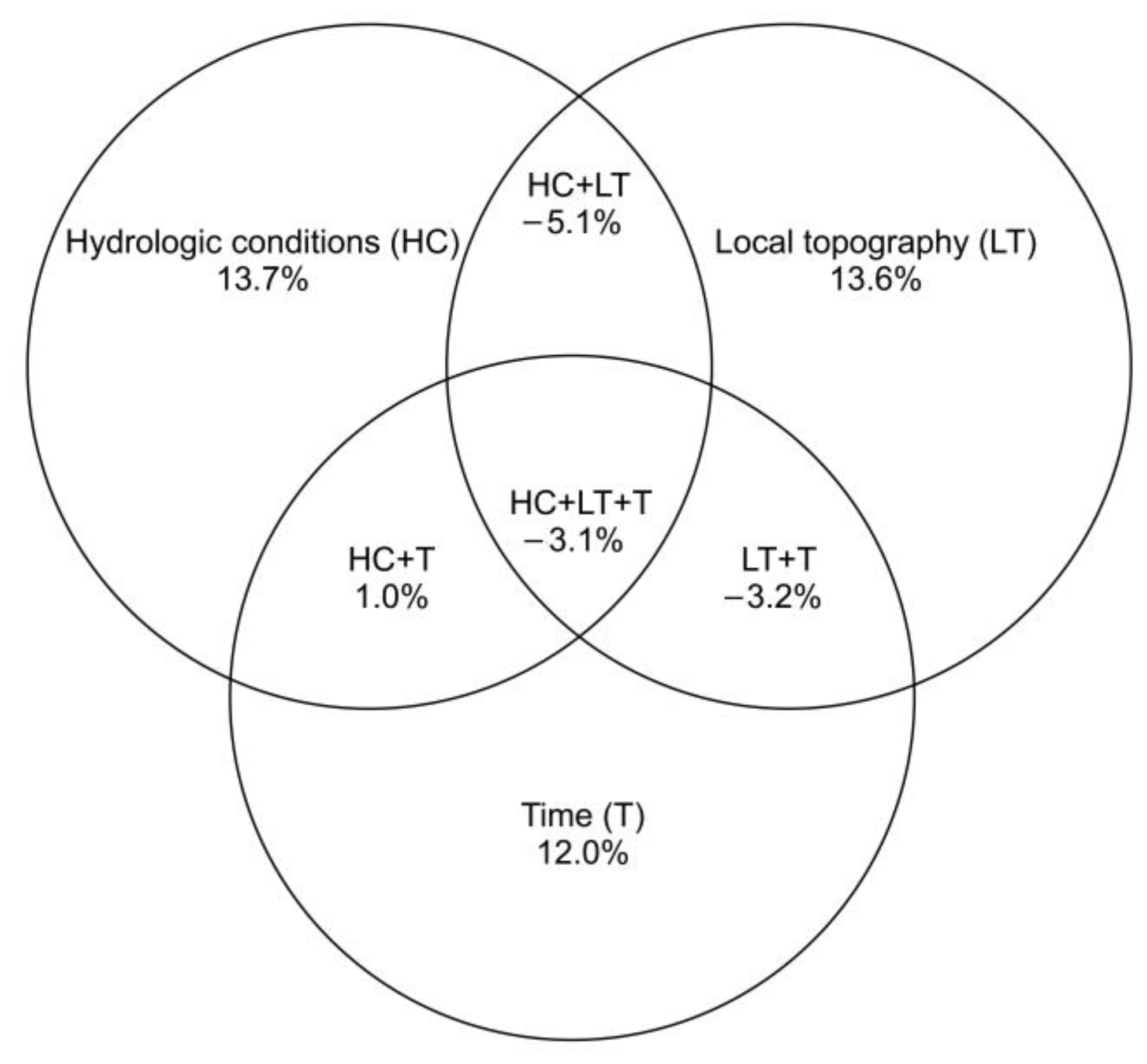
4.1. The Effects of Artificial Drainage (Hydrologic Changes and Local Topography) on Earthworm Community Structure and Composition–Taxonomic Approach
4.2. The Effects of Artificial Drainage (Hydrologic Changes and Local Topography) on the Composition and Distribution Pattern of Earthworm Traits—A Functional Approach
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Picket, S.T.A.; White, P.S. The Ecology of Natural Disturbance and Patch Dynamic; Academic Press: Orlando, FL, USA, 1985; pp. 1–472. [Google Scholar]
- White, P.S.; Jentsch, A. The search for generality in studies of disturbance and ecosystem dynamic. Prog. Bot. 2001, 62, 400–450. [Google Scholar] [CrossRef]
- Grime, J.P. Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences. J. Veg. Sci. 2006, 17, 255–260. [Google Scholar] [CrossRef]
- Menta, C. Soil fauna diversity–function, soil degradation, biological indices, soil restoration. In Biodiversity Conservation and Utilization in a Diverse World; Lameed, G.A., Ed.; InTechOpen: London, UK, 2012; pp. 59–94. [Google Scholar] [CrossRef] [Green Version]
- Fournier, B.; Samaritani, E.; Shrestha, J.; Mitchell, E.A.D.; Le Bayon, R.C. Patterns of earthworm communities and species traits in relation to the perturbation gradient of a restored floodplain. Appl. Soil Ecol. 2012, 59, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Fournier, B.; Gillet, F.; Le Bayon, R.C.; Mitchell, E.A.D.; Moretti, M. Functional response of multitaxa communities to disturbance and stress gradients in a restored floodplain. J. Appl. Ecol. 2015, 52, 1364–1373. [Google Scholar] [CrossRef]
- De Lange, H.J.; Kramer, K.; Faber, J.H. Two approaches using traits to assess ecological resilience: A case study on earthworm communities. Basic Appl. Ecol. 2013, 14, 64–73. [Google Scholar] [CrossRef]
- Sterzyńska, M.; Shrubovych, J.; Kaprus, I. Effect of hydrologic regime and forest age on Collembola in riparian forests. Appl. Soil Ecol. 2014, 75, 199–209. [Google Scholar] [CrossRef]
- Sterzyńska, M.; Tajovsky, K.; Nicia, P. Contrasting responses of millipedes and terrestrial isopods to hydrologic regime changes in forested montane wetlands. Eur. J. Soil Biol. 2015, 68, 33–44. [Google Scholar] [CrossRef]
- Mims, M.C.; Olden, J.D. Life history theory predicts fish assemblage response to hydrologic regime. Ecology 2012, 93, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Poff, N.L. Regional hydrologic response to climate change: An ecological perspective. In Global Climate Change and Freshwater Ecosystems; Firth, P., Fisher, S.G., Eds.; Springer: New York, NY, USA, 1992; pp. 88–115. [Google Scholar] [CrossRef]
- Plum, J. Terrestrial invertebrates in flooded grasslands: A literature review. Wetlands 2005, 25, 721–737. [Google Scholar] [CrossRef]
- Plum, J.; Filser, J. Floods and drought: Response of earthworms and potworms (Oligochaeta: Lumbricidae, Enchytraeidae) to hydrological extremes in wet grassland. Pedobiologia 2005, 49, 443–453. [Google Scholar] [CrossRef]
- Marx, M.T.; Guhamnn, P.; Decker, P. Adaptations and Predispositions of Different Middle European Arthropod Taxa (Collembola, Araneae, Chilopoda, Diplopoda) to Flooding and Drought Conditions. Animals 2012, 2, 564–590. [Google Scholar] [CrossRef] [Green Version]
- Berga, M.; Székely, A.J.; Langenheder, S. 2012. Effects of disturbance intensity and frequency on bacterial community composition and function. PLoS ONE 2012, 7, e36959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional. Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Diaz, S.; Purvis, A.; Cornelissen, J.H.C.; Mace, G.M.; Donoghue, M.J.; Ewers, R.M.; Jordano, P.; Pearse, W.D. Functional traits, the phylogeny of function and ecosystem service vulnerability. Ecol. Evol. 2013, 3, 2958–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandewalle, M.; de Bello, F.; Berg, M.P.; Bolger, T.; Dolédec, S.; Dubs, F.; Feld, C.K.; Harrington, R.; Harrison, P.A.; Lavorel, S.; et al. Functional traits as indicators of biodiversity response to land use changes across ecosystems and organisms. Biodivers. Conserv. 2010, 19, 2921–2947. [Google Scholar] [CrossRef] [Green Version]
- Mouillot, D.; Mason, N.W.H.; Wilson, J.B. Is the abundance of species determined by their functional traits? A new method with a test using plant communities. Oecologia 2007, 152, 729–737. [Google Scholar] [CrossRef]
- Mouillot, D.; Graham, N.A.J.; Villeger, S.; Mason, N.W.H.; Bellwood, D.R. A functional approach reveals community responses to disturbances. Trends Ecol. Evol. 2013, 28, 167–177. [Google Scholar] [CrossRef]
- Castro-Diez, P. Functional traits analyses: Scaling-up from species to community level. Plant Soil 2012, 357, 9–12. [Google Scholar] [CrossRef]
- Lebrija-Trejos, E.; Pérez-Garcia, A.; Maeve, J.A.; Bongers, F.; Poorter, L. Functional traits and environmental filtering drive community assembly in a species-rich tropical system. Ecology 2010, 91, 386–398. [Google Scholar] [CrossRef] [Green Version]
- Sterk, M.; Gort, G.; Klimkowska, A.; van Ruijven, J.; van Teeffelen, A.J.A.; Wamelink, G.W.W. Asses ecosystem resilience: Linking response and effect traits to environmental variability. Ecol. Indic. 2013, 30, 21–27. [Google Scholar] [CrossRef]
- Laughlin, D.C. Applying trait-base models to archive functional targets for theory-driven ecological restoration. Ecol. Lett. 2014, 17, 771–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadotte, M.W.; Arnillas, C.A.; Livingstone, S.W.; Yasui, S.M.L. Predicting communities from functional traits. Trends Ecol. Evol. 2015, 30, 510–511. [Google Scholar] [CrossRef] [PubMed]
- Suding, K.N.; Lavorel, S.; Chapin, F.S.; Cornelissen, H.C.; Diaz, S.; Garnier, E.; Golderg, D.; Hooer, D.U.; Jackson, S.T.; Navas, M.L. Scaling environmental change through the community-level: A trait-based response-and-effect framework for plants. Glob. Chang. Biol. 2008, 145, 1125–1140. [Google Scholar] [CrossRef] [Green Version]
- Pey, B.; Nahmani, J.; Auclerc, A.; Capowiez, Y.; Cluzeau, D.; Cortet, J.; Decaëns, T.; Deharveng, L.; Dubis, F.; Joimel, S.; et al. Current use of and future needs for soil invertebrate functional traits in community ecology. Basic Appl. Ecol. 2014, 15, 194–206. [Google Scholar] [CrossRef] [Green Version]
- Makkonen, M.; Berg, M.P.; van Hal, J.R.; Callaghan, T.V.; Press, M.C.; Aerts, R. Traits explain the response of a sub-arctic Collembola community to climate manipulation. Soil Biol. Biochem. 2011, 43, 377–384. [Google Scholar] [CrossRef]
- Malmström, A. Life-history traits predict recovery patterns in Collembola species after fire: A 10 year study. Appl. Soil Ecol. 2012, 56, 35–42. [Google Scholar] [CrossRef]
- Hedde, M.; van Oort, F.; Lamy, I. Functional traits of soil invertebrates as indicators for exposure to soil disturbance. Environ. Pollut. 2012, 164, 59–65. [Google Scholar] [CrossRef]
- Andriuzzi, W.S.; Schmidt, O.; Brussard, L.; Faber, J.H.; Bolger, T. Earthworm functional traits and interspecific interaction affect plant nitrogen acquisition and primary production. Appl. Soil Ecol. 2016, 104, 148–156. [Google Scholar] [CrossRef]
- Southwood, T.R.E. Habitat, the template for ecological strategies. J. Anim. Ecol. 1977, 46, 337–365. [Google Scholar] [CrossRef]
- Southwood, T.R.E. Tactics, strategies and templates. Oikos 1988, 52, 3–18. [Google Scholar] [CrossRef]
- Keane, R. Disturbance regimes and the historical range of variation in terrestrial ecosystems. In Encyclopaedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: Orlando, FL, USA, 2013; pp. 568–581. [Google Scholar] [CrossRef]
- Walker, L.R. Ecosystem of Disturbed Ground, 1st ed.; Elsevier: Amsterdam, The Netherlands, 1999; pp. 1–868. [Google Scholar]
- Bernhart-Römermann, M.; Gray, A.; Vangergen, A.J.; Bergès, L.; Bohner, A.; Brooker, R.W.; De Cinti, B.; Dirnböck, T.; Grandin, U.; Hester, A.J.; et al. Functional traits and local environment predict vegetation response s to disturbance: A pan-European multi-site experiment. J. Ecol. 2011, 99, 777–787. [Google Scholar] [CrossRef]
- Stein, E.D.; Mattson, M.; Fetscher, A.E.; Halama, K.J. Influence of geologic setting on slope wetland hydrodynamics. Wetlands 2004, 24, 244–260. [Google Scholar] [CrossRef]
- Chimner, R.A.; Lemly, J.M.; Cooper, D.J. Mountain fen distribution, types and restoration priorities, San Juan Mountains, Colorado, USA. Wetlands 2010, 30, 763–771. [Google Scholar] [CrossRef]
- Batzer, D.P. The seemingly intractable ecological responses of invertebrates in North American wetlands: A Review. Wetlands 2013, 33, 1–15. [Google Scholar] [CrossRef]
- Holden, J.; Chapman, P.J.; Labadz, J.C. Artificial drainage of peatlands: Hydrological; and hydrochemical process and wetland restoration. Prog. Phys. Geogr. 2004, 28, 95–123. [Google Scholar] [CrossRef] [Green Version]
- Zeitz, J.; Velty, S. Soil properties of drained and rewetted fen soils. J. Plant. Nutr. Soil Sci. 2002, 165, 618–626. [Google Scholar] [CrossRef]
- Laiho, R. 2006. Decomposition in peatlands: Reconciling seemingly contrasting results on the impacts of lowered water levels. Soil Biol. Biochem. 2006, 38, 2011–2024. [Google Scholar] [CrossRef]
- Pitkänen, A.; Turunen, J.; Tahvanainen, T.; Simola, H. Carbon storage change in a partially forestry-drained boreal mire determined through peat column inventories. Boreal Environ. Res. 2013, 18, 223–234. [Google Scholar]
- Kopp, B.J.; Fleckenstein, J.H.; Roulet, N.T.; Humpreys, E.; Talbot, J.; Blodau, C. Impact of long-term drainage on summer groundwater flow patterns in the Mer Blue peatland, Ontario, Canada. Hydrol. Earth Syst. Sci. 2013, 17, 3485–3498. [Google Scholar] [CrossRef] [Green Version]
- Paoletti, M.G. The role of earthworms for assessment of sustainability and as bioindicators. Agric. Ecosyst. Environ. 1999, 74, 137–155. [Google Scholar] [CrossRef]
- Blouin, M.; Hodson, M.E.; Delgado, E.A.; Baker, G.; Brussard, L.; Butt, K.R.; Dai, J.; Dendooven, L.; Peres, G.; Tondoh, J.E.; et al. A review of earthworm impact on soil function and ecosystem services. Eur. J. Soil Sci. 2013, 64, 161–182. [Google Scholar] [CrossRef]
- Paoletti, M.G. Invertebrate Biodiversity as Bioindicators of Sustainable Landscapes: Practical Use of Invertebrates to Assess Sustainable Land Use; Elsevier Science: Amsterdam, The Netherlands, 2012; pp. 1–460. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2006; 1st update 2007; World Soil Resources Reports No. 103; FAO: Rome, Italy, 2007; pp. 1–128. [Google Scholar]
- Bouché, M.B. Lombriciens de France Écologie et Systématique; Institut National de la Recherche Agronomique: Paris, France, 1972; pp. 1–671. [Google Scholar]
- Hale, C.M.; Reich, P.B.; Frelich, L.E. Allometric Equations for Estimation of Ash-free Dry Mass from Length Measurements for Selected European Earthworm Species (Lumbricidae) in the Western Great Lakes Region. Am. Midl. Nat. 2004, 151, 179–185. [Google Scholar] [CrossRef]
- Csuzdi, C. Earthworm species, a searchable database. Opuscula Zoologica 2012, 43, 97–99. [Google Scholar] [CrossRef]
- DriloBASE TAXO, World Earthworms Database. 2014–2018 by Drilobase Project. Available online: http://taxo.drilobase.org (accessed on 10 October 2022).
- Edaphobase. Senckenberg Museum für Naturkunde Görlitz. Occurrence Dataset. Available online: https://doi.org/10.15468/rk9xc7 (accessed on 6 December 2022).
- Garnier, E.; Cortez, J.; Billès, G.; Navas, M.L.; Roumet, C.; Debussche, M.; Laurent, G.; Blanchard, A.; Aubry, D.; Bellman, A.; et al. Plant functional markers capture ecosystem properties during secondary succession. Ecology 2004, 85, 2630–2637. [Google Scholar] [CrossRef]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003; pp. 1–282. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Baar, J.; ter Braak, C.J.F. Ectomycorrhizal sporocarp occurrence as affected by manipulation of litter and humus layers in Scots pine stands of different age. Appl. Soil Ecol. 1996, 4, 61–73. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier Science: Amsterdam, The Netherlands, 1998; pp. 1–852. [Google Scholar]
- Leifeld, J.; Steffens, M.; Galego, A. Sensitivity of peatland carbon loss to organic matter quality. Geophys. Res. Lett. 2012, 39, L14704. [Google Scholar] [CrossRef] [Green Version]
- Nicia, P.; Zadrożny, P.; Lamorski, T. 2009. General characteristic of selected soil profiles under the Caltho-Alnetum association in the Babiogórski National Park. Ecol. Chem. Eng. A 2009, 16, 839–843. [Google Scholar]
- Butt, K.R.; Lowe, C.N.; Frederckson, J.; Moffat, A.J. The development of sustainable earthworm populations at Calvert Landfill site, UK. Land Degrad. Dev. 2004, 15, 27–36. [Google Scholar] [CrossRef]
- Johnston, A.S.A.; Holmstrup, M.; Hodson, M.E.; Thorbek, M.E.; Alvarez, T.; Sibly, R.M. Earthworm distribution and abundance predicted by a process-based model. Appl. Soil Ecol. 2014, 84, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Marhan, S.; Scheu, S. The influence of mineral and organic fertilisers on the growth of the endogeic earthworm Octolasion tyrtaeum (Savigny). Pedobiologia 2005, 49, 239–249. [Google Scholar] [CrossRef]
- Eijsackers, H. Earthworms as colonizers of natural and cultivated soil environments. Appl. Soil Ecol. 2011, 50, 1–13. [Google Scholar] [CrossRef]
- Lee, K.E. Earthworms: Their Ecology and Relationship with Soils and Land Use; Academic Press: Sydney, Australia, 1985; pp. 1–411. [Google Scholar]
- Edwards, C.A.; Bohlen, P.J. Biology and Ecology of Earthworms, 3rd ed.; Chapman and Hall: London, UK, 1996; pp. 1–426. [Google Scholar]
- Von Groenigen, J.W.; Lubbers, I.M.; Vos, H.M.J.; Brown, G.G.; de Deyn, G.B.; van Groenigen, K.J. Earthworms increase plant production: A meta-analysis. Sci. Rep. 2014, 4, 6365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouché, M.B. Strategies lombricinnes. Ecol. Bull. 1977, 25, 122–132. [Google Scholar]
- Bohlen, P.J.; Edwards, W.M.; Edwards, C.A. Earthworm community structure and diversity in experimental agricultural watersheds in Norteastern Ohio. Plant Soil. 1995, 170, 233–239. [Google Scholar]
- Shekhostov, S.V.; Golovanova, E.V.; Peltek, S.E. Genetic diversity of the earthworm Octolasion tyrtaeum (Lumbricidae, Annelida). Pedobiologia 2014, 57, 245–250. [Google Scholar] [CrossRef]
- Heethoff, M.; Etzold, K.; Scheu, S. Mitochondrial COII sequences indicate that the parthenogenetic earthworm Octolasion tyrtaeum (Savigny 1826) constitutes of two lineages differing in body size and genotype. Pedobiologia 2004, 48, 9–13. [Google Scholar] [CrossRef]
- Klok, C.; Zorn, M.; Koolhaas, J.E.; Eijsackers, H.J.P.; van Gestel, C.A.M. Does reproductive plasticity in Lumbricus rubellus improve the recovery of populations in frequently inundated river floodplain. Soil Biol. Biochem. 2006, 38, 611–618. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, G.; Ni, J. Effects of topographical and edaphic factors on the distribution of plant communities in two subtropical karst forests, southwestern China. J. Mt. Sci. 2013, 10, 95–104. [Google Scholar] [CrossRef]
- Łajczak, A. Role of land-relief and structure on the formation of peat-bogs in mountain areas, as exemplified by the Polish Carpathians. Landf. Anal. 2013, 22, 61–73. [Google Scholar] [CrossRef]
- Glorvigen, P.; Andreassen, H.P.; Ims, R.A. Local and regional determinants of colonisation-extinction dynamics of a riparian mainland-island root vole metapopulation. PLoS ONE 2013, 8, e56462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matias, M.G.; Underwood, A.J.; Hochuli, D.F.; Coleman, R.A. Independent effect of patch size and structural complexity on diversity of benthic macroinvertebrates. Ecology 2010, 91, 1908–1915. [Google Scholar] [CrossRef] [PubMed]
- Bell, G.; Lechowicz, M.J.; Appenzeller, A.; Chandler, M.; DeBlois, E.; Jackson, L.; Mackenzie, B.; Preziosi, R.; Schallenberg, M.; Tinker, N. The spatial structure of physical environment. Oecologia 1993, 96, 114–121. [Google Scholar] [CrossRef]
- Stine, M.B.; Resler, L.M.; Campbell, J.B. Ecotone characteristics of southern Appalachian Mountain wetland. Catena 2011, 86, 57–65. [Google Scholar] [CrossRef]
- Clobert, J.; Baguette, M.; Benton, T.G.; Bullock, J.M. Dispersal Ecology and Evolution; Oxford University Press: Oxford, UK, 2012; pp. 1–462. [Google Scholar] [CrossRef]
- Jimenéz, J.J.; Decaëns, T.; Amézquita, E.; Rao, I.; Thomas, R.J.; Lavelle, P. Short-range spatial variability of soil physic-chemical variables related to earthworm clustering in a neotropical gallery forest. Soil Biol. Biochem. 2011, 43, 1071–1080. [Google Scholar] [CrossRef] [Green Version]
- Decaëns, T. Macroecological patterns in soil communities. Glob. Ecol. Biogeogr. 2010, 19, 287–302. [Google Scholar] [CrossRef]
- Salome, C.; Guenat, C.; Bullinger-Weber, G.; Gobat, J.M. Earthworm communities in alluvial forests: Influence of altitude, vegetation stage and soil parameters. Pedobiologia 2011, 54, 89–98. [Google Scholar] [CrossRef]
- Rożen, A.; Mysłajek, A.; Sobczyk, W. Altitude versus vegetation as the factors influencing the diversity and abundance of earthworms and other soil macrofauna in montane habitat (Silesian Beskid Mts, Western Carpathians). Pol. J. Ecol. 2013, 61, 145–156. [Google Scholar]
- Kraft, N.J.B.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community assembly, coexistence and the environmental filtering metaphor. Funct. Ecol. 2015, 29, 592–599. [Google Scholar] [CrossRef]
- Bengtsson, J.; Angelstam, P.; Elmqvist, T.; Emanuelsson, U.; Folke, C.; Ihse, M.; Moberg, F.; Nyström, M. Reserves, resilience and dynamic landscapes. Ambio 2003, 32, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Ortmann-Ajkai, A.; Lóczy, D.; Gyenizse, P.; Pirkhoffer, E. Wetland habitat patches as ecological components of landscape memory in a highly modified floodplains. River Res. Appl. 2014, 30, 874–886. [Google Scholar] [CrossRef]
- Lindo, Z.; Whiteley, J.; Gonzales, A. Traits explain community disassembly and trophic contraction following experimental change. Glob. Chang. Biol. 2012, 18, 2448–2457. [Google Scholar] [CrossRef]
- Townsend, C.R.; Hildrew, A.G. Species traits in relation to a habitat templet for river systems. Freshw. Biol. 1994, 31, 265–275. [Google Scholar] [CrossRef]
- Snyder, R.E. Transient dynamics in altered disturbance regimes: Recovery may start quickly, than slow. Theor. Ecol. 2009, 2, 79–87. [Google Scholar] [CrossRef]
- Decaëns, T.; Margerie, P.; Aubert, M.; Hedde, M.; Bureau, F. Assembly rules within earthworm communities in North-Western France—A regional analysis. Appl. Soil Ecol. 2008, 39, 321–335. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hydrologic Conditions | Size (m2) | Slope (°) | Elevation (m a.s.l.) | Soil Type |
|---|---|---|---|---|
| Natural | ||||
| 1 | 1775 | 5 | 914 | Otni |
| 2 | 750 | 2 | 772 | POtni |
| 3 | 546 | 5 | 703 | POtni |
| Semi-natural | ||||
| 4 | 255 | 8 | 964 | POtni |
| 5 | 945 | 7 | 950 | POtni |
| 6 | 897 | 3 | 760 | POtni |
| Degraded | ||||
| 7 | 1284 | 3 | 962 | Mtni |
| 8 | 12,903 | 3 | 771 | Mtni |
| 9 | 739 | 6 | 740 | Ah |
| Traits | Data Type | Attribute Class | Code |
|---|---|---|---|
| Biological traits: | |||
| Length min/max at maturity stage (mm) | ordinal | 1—<50 mm | Lmin/max |
| 2—50–90 mm | |||
| 3—>90 mm | |||
| Prostomium | ordinal | 1—epilobic open | Prost |
| 2—epilobic closed | |||
| 3—tanylobic | |||
| Dispersal potential | ordinal | 1—high | Dispot |
| 2—intermediate | |||
| 3—low | |||
| Quiescence | binary | 1—no | Quisc |
| 2—yes | |||
| Performance traits: | |||
| Time to maturity (weeks) | binary | 1—<52 | Tmat |
| 2—>52 | |||
| Cocoons (number/year) | binary | 1—<36 | Cocons |
| 2—>36 | |||
| Ecological traits: | |||
| Vertical distribution | ordinal | 1—epigeic | Vdist |
| 2—epiendogeic | |||
| 3—endogeic | |||
| Habitat width | binary | 1—steno | Habwidth |
| 2—eury | |||
| Hygrophily | ordinal | 1—high | Hydro |
| 2—intermediate | |||
| 3—low | |||
| C/N soil preference | ordinal | 1—low | C/Npref |
| 2—eury | |||
| 3—high | |||
| Low pH tolerance | binary | 1—tolerant | pHtol |
| 2—euryvalent |
| Soil Properties | Natural | Semi-Natural | Degraded | |||
|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | |
| Mg2+ * (μmol kg−1 soil) | 86.11 | 8.68 | 52.78 | 6.01 | 21.06 | 16.07 |
| EC * | 0.19 | 0.08 | 0.12 | 0.04 | 0.06 | 0.02 |
| TEB * (μmol kg−1 soil) | 919.28 | 268.62 | 598.02 | 92.77 | 125.53 | 132.46 |
| Hh(=H+) * (μmol kg−1 soil) | 6.78 | 2.03 | 7.18 | 0.66 | 137.21 | 54.48 |
| CEC | 926.06 | 266.74 | 605.20 | 92.84 | 262.74 | 112.73 |
| BS * (%) | 99.13 | 0.55 | 98.79 | 0.19 | 72.34 | 5.33 |
| pHH2O | 6.50 | 0.45 | 5.28 | 0.29 | 4.72 | 0.66 |
| C/N | 22.68 | 3.64 | 19.08 | 2.32 | 20.09 | 2.69 |
| G.W.L. * (cm) | 3.53 | 0.78 | 14.39 | 3.08 | 22.17 | 13.43 |
| Species | Code | Natural | Semi-Natural | Degraded | HC | Site | ||
|---|---|---|---|---|---|---|---|---|
| F/K | p | F | p | |||||
| Aporrectodea caliginosa (Savigny, 1826) | Acalig | 8.00 ± 12.31 | 8.59 ± 15.21 | 8.30 ± 19.89 | 0.16 | 0.854 | 1.52 | 0.193 |
| Aporrectodea rosea (Savigny, 1826) | Arosea | 5.63 ± 8.07 | 2.07 ± 3.24 | 5.33 ± 12.54 | 0.53 | 0.595 | 1.42 | 0.228 |
| Dendrobaena octaedra (Savigny, 1826) | Docta | 58.67 ± 35.0 | 35.56 ± 26.06 | 53.33 ± 43.79 | 3.85 | 0.146 | ||
| Dendrodrilus rubidus (Savigny, 1826) | Drubid | 3.85 ± 7.50 | 4.15 ± 8.29 | 2.96 ± 4.92 | 0.002 | 0.998 | 1.04 | 0.412 |
| Eisenia lucens (Waga, 1875) | Eisluc | 4.15 ± 7.21 | 0.59 ± 1.72 | 2.67 ± 5.56 | 2.43 | 0.297 | ||
| Eiseniella tetraedra (Savigny, 1826) | Etetra | 124.44 ± 191.64 b | 69.33 ± 43.13 c | 3.85 ± 7.27 a | 19.61 | 0.001 * | ||
| Lumbricus rubellus Hoffmeister, 1845 | Lumru | 3.56 ± 6.84 ab | 1.85 ± 3.90 b | 5.04 ± 7.42 a | 3.23 | 0.049 * | 3.54 | 0.006 * |
| Octodrilus argoviensis (Bretscher, 1899) | Octarg | 0.30 ± 1.26 a | 14.52 ± 19.43 b | 0.00 ± 0.00 a | 20.95 | 0.000 * | ||
| Octodrilus transpadanus (Rosa, 1884) | Octtra | 1.19 ± 3.90 | 0.30 ± 1.26 | 0.00 ± 0.00 | 1.23 | 0.334 | 0.65 | 0.692 |
| Octolasion tyrtaeum (Savigny, 1826) | Otyrt | 40.59 ± 58.20 b | 13.33 ± 17.76 ab | 7.11 ± 11.57 a | 5.10 | 0.010 * | 6.53 | 0.000 * |
| A (total) | 250.37 ± 244.53 b | 149.63 ± 80.71 ab | 88.59 ± 49.06 a | 6.24 | 0.004 * | 6.94 | 0.000 * | |
| S | 4.22 ± 1.63 | 4.44 ± 1.20 | 3.56 ± 1.15 | 2.17 | 0.125 | 0.90 | 0.501 | |
| H’ | 1.07 ± 0.40 | 1.17 ± 0.25 | 0.84 ± 0.36 | 4.55 | 0.015 | 1.50 | 0.200 | |
| J’ | 0.77 ± 0.14 | 0.81 ± 0.09 | 0.68 ± 0.20 | 4.07 | 0.131 |
| Marginal Effect (Forward: Step 1) | Conditional Effect (Forward: Continued) | ||||
|---|---|---|---|---|---|
| Variable | λ1 | λa | Cum % | F | p |
| Size | 0.05 | 0.05 | 5 | 3.01 | 0.008 * |
| Semi-natural | 0.05 | 0.04 | 9 | 2.27 | 0.058 |
| Elevation | 0.03 | 0.09 | 18 | 5.43 | 0.002 * |
| Slope | 0.05 | 0.03 | 21 | 1.81 | 0.094 |
| Time (year 2010) | 0.02 | 0.03 | 24 | 1.55 | 0.116 |
| Natural | 0.03 | 0.02 | 26 | 1.40 | 0.198 |
| G.W.L | 0.03 | 0.04 | 30 | 2.74 | 0.018 |
| Time (year 2011) | 0.02 | 0.02 | 32 | 1.00 | 0.406 |
| Time (spring) | 0.01 | 0.01 | 33 | 0.63 | 0.710 |
| Degraded | 0.05 | ||||
| Time (year 2012) | 0.02 | ||||
| Time (spring) | 0.01 | ||||
| Time (autumn) | 0.01 | ||||
| Traits | Natural | Semi-Natural | Degraded | HC | Site | ||
|---|---|---|---|---|---|---|---|
| F/K | p | F | p | ||||
| Biological traits: | |||||||
| Lmin (mm) | 2.04 ± 0.03 | 2.03 ± 0.04 | 2.06 ± 0.06 | 1.60 | 0.219 | 1.10 | 0.357 |
| Lmax (mm) | 2.36 ± 0.03 | 2.34 ± 0.04 | 2.37 ± 0.05 | 6.32 | 0.042 * | ||
| Prostomium | 2.07 ± 0.08 | 2.10 ± 0.07 | 2.10 ± 0.10 | 0.71 | 0.499 | 2.23 | 0.056 |
| Dispersal potential | 2.16 ± 0.06 | 2.11 ± 0.06 | 2.14 ± 0.13 | 4.26 | 0.119 | ||
| Quiescence | 2.17 ± 0.08 | 2.11 ± 0.11 | 2.17 ± 0.14 | 1.46 | 0.244 | 2.56 | 0.032 * |
| Performance traits: | |||||||
| Time to maturity | 2.17 ± 0.09 | 2.17 ± 0.09 | 2.17 ± 0.14 | 0.01 | 0.990 | 2.09 | 0.073 |
| Cocoons (n/year) | 2.19 ± 0.07 | 2.17 ± 0.09 | 2.18 ± 0.13 | 0.77 | 0.681 | ||
| Ecological traits: | |||||||
| Vertical distribution | 2.18 ± 0.11 | 2.15 ± 0.10 | 2.18 ± 0.14 | 0.60 | 0.554 | 2.37 | 0.045 * |
| Habitat width | 2.37 ± 0.11 b | 2.28 ± 0.08 a | 2.44 ± 0.08 c | 24.83 | 0.000 * | 7.00 | 0.000 * |
| Hygrophily | 2.26 ± 0.09 b | 2.19 ± 0.07 a | 2.31 ± 0.07 c | 19.18 | 0.000 * | 6.43 | 0.000 * |
| C/N soil preference | 2.38 ± 0.06 | 2.36 ± 0.06 | 2.40 ± 0.08 | 1.62 | 0.209 | 2.83 | 0.020 * |
| Low pH tolerance | 2.27 ± 0.15 | 2.35 ± 0.05 | 2.20 ± 0.14 | 7.80 | 0.02 * | ||
| Marginal Effect (Forward: Step 1) | Conditional Effect (Forward: Continued) | ||||
|---|---|---|---|---|---|
| Variable | λ1 | λa | cum % | F | p |
| Semi-natural | 0.07 | 0.07 | 7 | 3.92 | 0.020 * |
| Elevation | 0.05 | 0.10 | 17 | 6.47 | 0.012 * |
| Size | 0.07 | 0.05 | 22 | 3.20 | 0.038 * |
| Time (year 2010) | 0.05 | 0.05 | 27 | 3.07 | 0.036 * |
| Time (spring) | 0.03 | 0.03 | 30 | 1.92 | 0.152 |
| G.W.L. | 0.02 | 0.03 | 33 | 2.15 | 0.086 |
| Natural | 0.01 | 0.03 | 36 | 2.01 | 0.124 |
| Slope | 0.05 | −0.00 | 36 | 0.58 | 0.616 |
| Time (year 2011) | 0.04 | 0.01 | 37 | 0.47 | 0.678 |
| Degraded | 0.04 | ||||
| Time (autumn) | 0.03 | ||||
| Time (year 2012) | 0.01 | ||||
| Coef. (±SD) | t | DFs. | F | p | Coef. (±SD) | t | DFs | F | p | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Biological traits | |||||||||||
| Lmin | |||||||||||
| null | 71.80 | null | 71.80 | ||||||||
| intercept | 4.695 (0.013) | 360.716 | intercept | 4.695 (0.013) | 360.716 | ||||||
| Ax1 | 71.80 | ns | Ax2 | 71.80 | ns | ||||||
| Lmax | |||||||||||
| null | 117.81 | null | 117.81 | ||||||||
| intercept | 5.428 (0.009) | 601.421 | intercept | 5.429 (0.009) | 602.353 | ||||||
| Ax1 | 0.051 (0.009) | 5.686 | 85.51 | 19.55 | <0.001 * | Ax2 | 117.81 | ns | |||
| Prost | |||||||||||
| null | 287.73 | null | 287.73 | ||||||||
| intercept | 4.821 (0.012) | 3.94.724 | intercept | 4.820 (0.012) | 393.649 | ||||||
| Ax1 | - | - | 287.73 | ns | Ax2 | 0.067 (0.012) | 5.508 | 257.46 | 5.87 | 0.020 * | |
| Disppot | |||||||||||
| null | 363.32 | null | 363.32 | ||||||||
| intercept | 4.938 (0.012) | 514.0.84 | intercept | 4.943 (0.011) | 429.858 | ||||||
| Ax1 | 0.101 (0.011) | 7.254 | 285.33 | 14.19 | <0.001 * | Ax2 | 363.32 | ns | |||
| Quisc | |||||||||||
| null | 560.97 | null | 560.97 | ||||||||
| intercept | 4.970 (0.011) | 436.517 | intercept | 4.977 (0.011) | 440.399 | ||||||
| Ax1 | 0.121 (0.011) | 10.745 | 445.96 | 13.20 | <0.001 * | Ax2 | 560.97 | ns | |||
| Performance traits | |||||||||||
| Tmat | |||||||||||
| null | 505.68 | null | 505.68 | ||||||||
| intercept | 5.015 (0.011) | 451.913 | intercept | 5.018 (0.011) | 453.465 | ||||||
| Ax1 | 0.075 (0.011) | 6.818 | 459.40 | 5.22 | 0.026 * | Ax2 | 505.68 | ns | |||
| Cocoons | |||||||||||
| null | 425.47 | null | 425.47 | ||||||||
| intercept | 5.044 (0.010) | 461.124 | intercept | 5.047 (0.011) | 462.549 | ||||||
| Ax1 | 0.071 (0.010) | 6.571 | 382.28 | 5.83 | 0.019 * | Ax2 | 425.47 | ns | |||
| Ecological preference | |||||||||||
| Vdist | |||||||||||
| null | 580.91 | null | 580.91 | ||||||||
| intercept | 5.013 (0.011) | 450.267 | intercept | 5.019 (0.011) | 453.895 | ||||||
| Ax1 | 0.115 (0.011) | 10.457 | 472.01 | 11.75 | <0.001 * | Ax2 | 580.91 | ns | |||
| Habwidth | |||||||||||
| null | 757.28 | null | 757.28 | ||||||||
| intercept | 5.466 (0.009) | 616.719 | intercept | 5.373 (0.013) | 411.439 | ||||||
| Ax1 | 0.091 (0.009) | 10.416 | 649.05 | 9.26 | 0.007 * | Ax2 | 0.023 (0.009) | 2.674 | 629.29 | 5.49 | 0.007 * |
| Hydro | |||||||||||
| null | 429.54 | null | 429.54 | ||||||||
| intercept | 5.201 (0.010) | 514.084 | intercept | 5.122 (0.015) | 344.297 | ||||||
| Ax1 | 0.073 (0.010) | 7.254 | 376.92 | 7.53 | 0.008 * | Ax2 | 0.023 (0.009) | 2.355 | 357.54 | 5.34 | 0.008 * |
| C/Npref | |||||||||||
| null | 284.91 | null | 284.91 | ||||||||
| intercept | 5.486 (0.008) | 626.358 | intercept | 5.486 (0.009) | 626.358 | ||||||
| Ax1 | 284.91 | ns | Ax2 | 284.91 | ns | ||||||
| pHtot | |||||||||||
| null | 927.18 | null | 927.18 | ||||||||
| intercept | 5.278 (0.010) | 543.321 | intercept | 5.363 (0.014) | 386.442 | ||||||
| Ax1 | 927.18 | ns | Ax2 | −0.081 (0.010) | −7.884 | 760.58 | 0.006 * | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pižl, V.; Sterzyńska, M.; Tajovský, K.; Starý, J.; Nicia, P.; Zadrożny, P.; Bejger, R. Effects of Hydrologic Regime Changes on a Taxonomic and Functional Trait Structure of Earthworm Communities in Mountain Wetlands. Biology 2023, 12, 482. https://doi.org/10.3390/biology12030482
Pižl V, Sterzyńska M, Tajovský K, Starý J, Nicia P, Zadrożny P, Bejger R. Effects of Hydrologic Regime Changes on a Taxonomic and Functional Trait Structure of Earthworm Communities in Mountain Wetlands. Biology. 2023; 12(3):482. https://doi.org/10.3390/biology12030482
Chicago/Turabian StylePižl, Václav, Maria Sterzyńska, Karel Tajovský, Josef Starý, Paweł Nicia, Paweł Zadrożny, and Romualda Bejger. 2023. "Effects of Hydrologic Regime Changes on a Taxonomic and Functional Trait Structure of Earthworm Communities in Mountain Wetlands" Biology 12, no. 3: 482. https://doi.org/10.3390/biology12030482