
Environmental Enrichment and Metformin Improve Metabolic Functions, Hippocampal Neuron Survival, and Hippocampal-Dependent Memory in High-Fat/High-Sucrose Diet-Induced Type 2 Diabetic Rats


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Ethics Statement
2.2. Diets
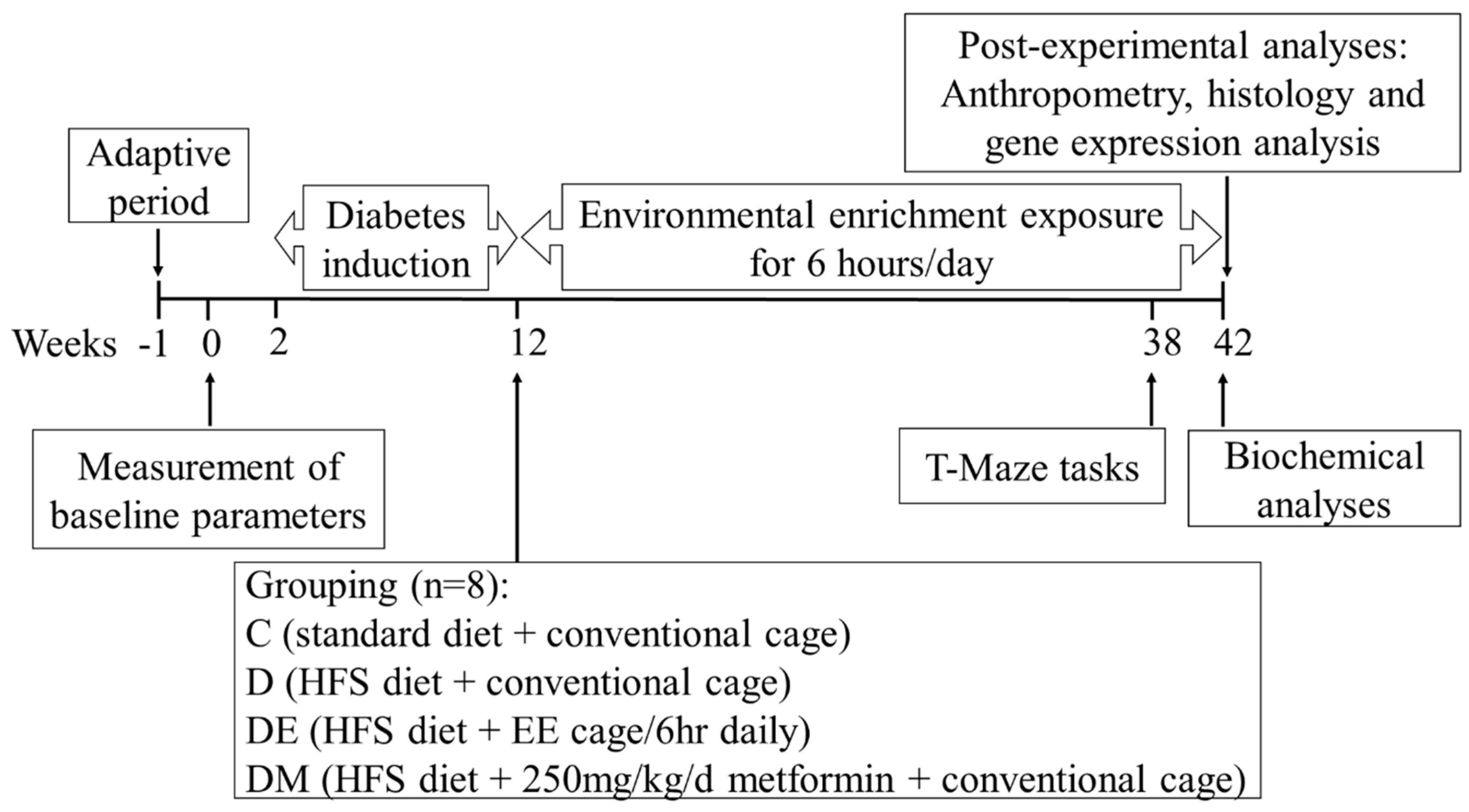
2.3. Experimental Design
2.4. Environmental Enrichment (EE) Cages
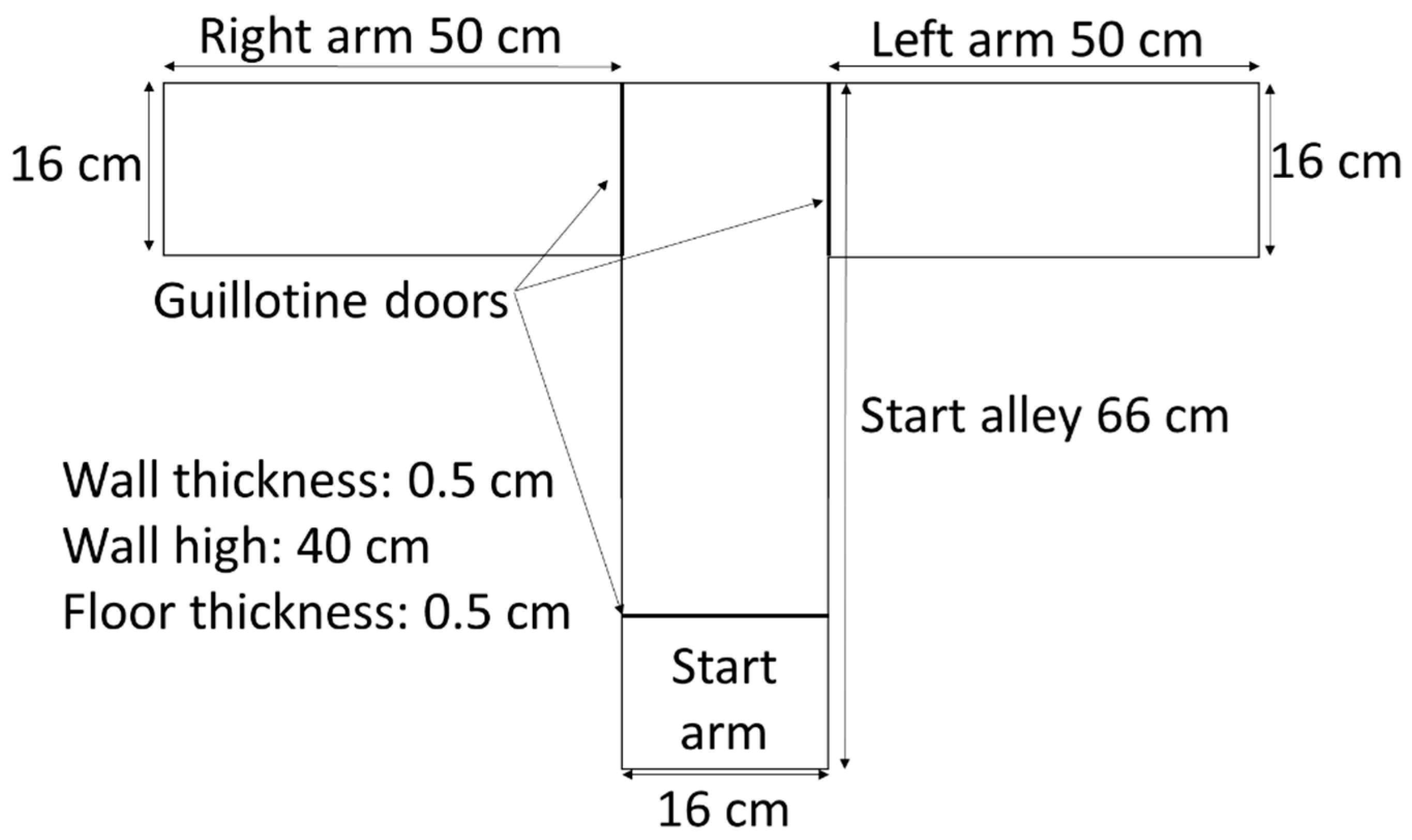
2.5. T-Maze Tasks: Spontaneous Alternation and Rewarded Alternation
2.6. Spontaneous Alternation Task
2.7. Rewarded Alternation Task
2.8. Measurement of Anthropometric Parameters
2.9. Measurement of Fasting Blood Glucose (FBG)
2.10. Measurement of C-Peptide
2.11. Measurement of the Lipid Profile
2.12. Measurement of Oxidative Status
2.13. Measurement of Serum BDNF
2.14. Tissue Preparation
2.15. Histology
2.16. Image Morphometry
2.17. Morphometric Measurement of Adipose Tissue
2.18. Morphometric Measurement of Pancreatic Islets
2.19. Morphometric Measurement of Hippocampal Neurons
2.20. RNA Extraction
2.21. Analysis of Gene Expression by the Real-Time PCR
2.22. Statistical Analysis
3. Results
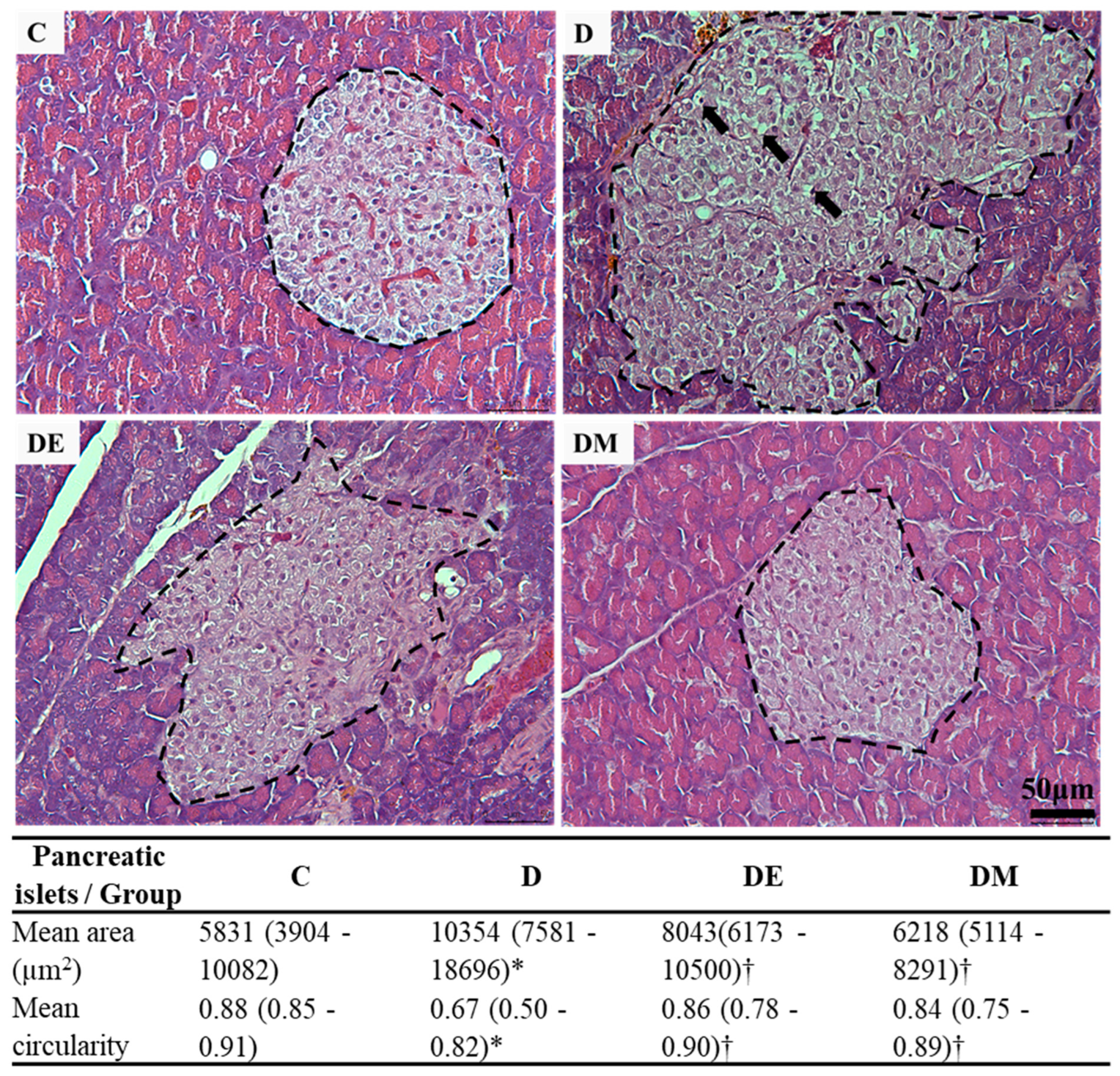
3.1. Effects of EE Exposure and Metformin Treatment on Fasting Blood Glucose (FBG), Serum C-Peptide, Histological Features, and Morphometric Measurement of Pancreatic Islets in HFS Diet-Induced T2D Rats
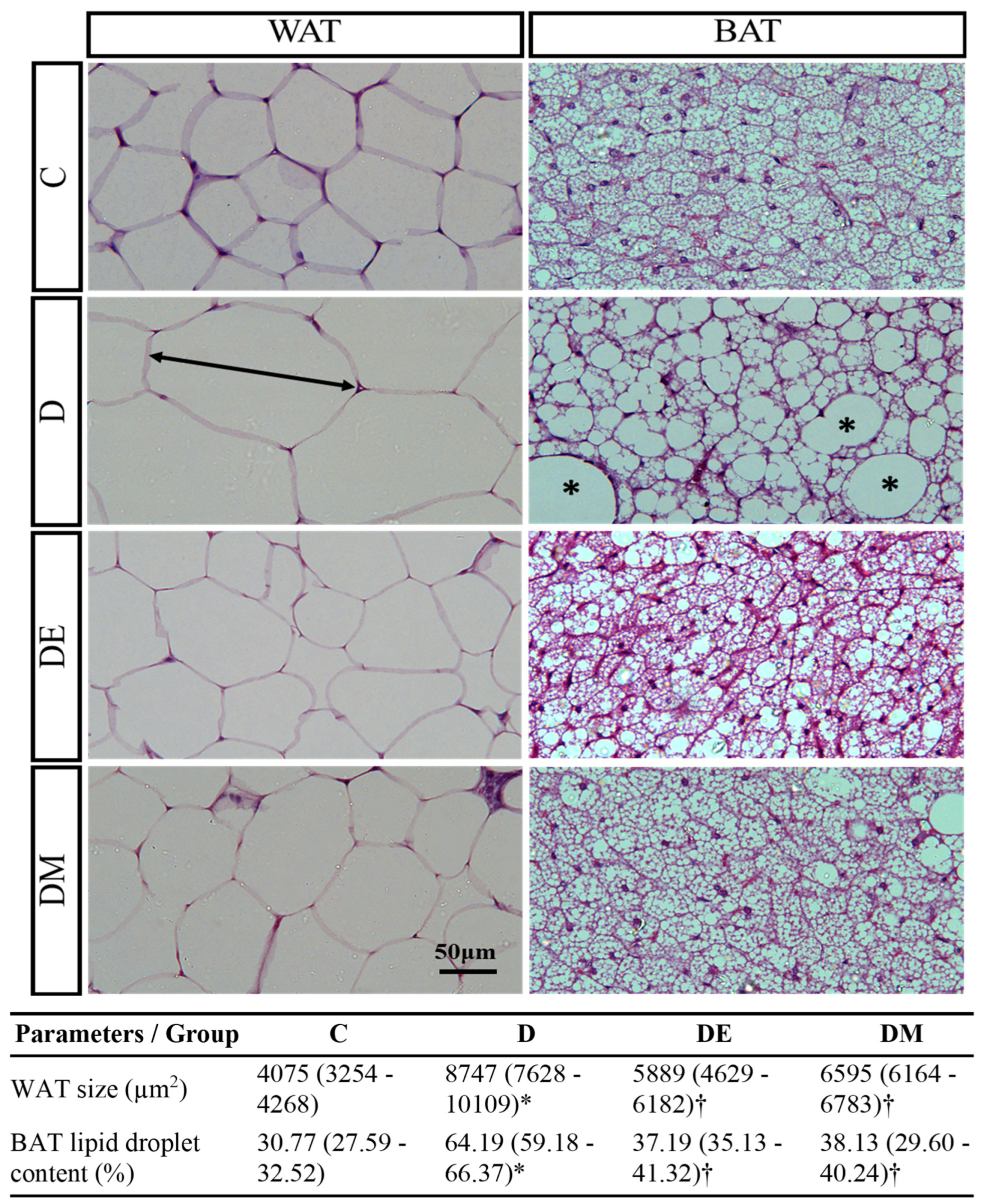
3.2. Effects of EE Exposure and Metformin Treatment on Serum Lipid Profile, Anthropometric Parameters, Histological Features, and Morphometric Measurement of Adipose Tissue in HFS Diet-Induced T2D Rats
3.3. Effects of EE Exposure and Metformin Treatment on Oxidative Status in HFS Diet-Induced T2D Rats
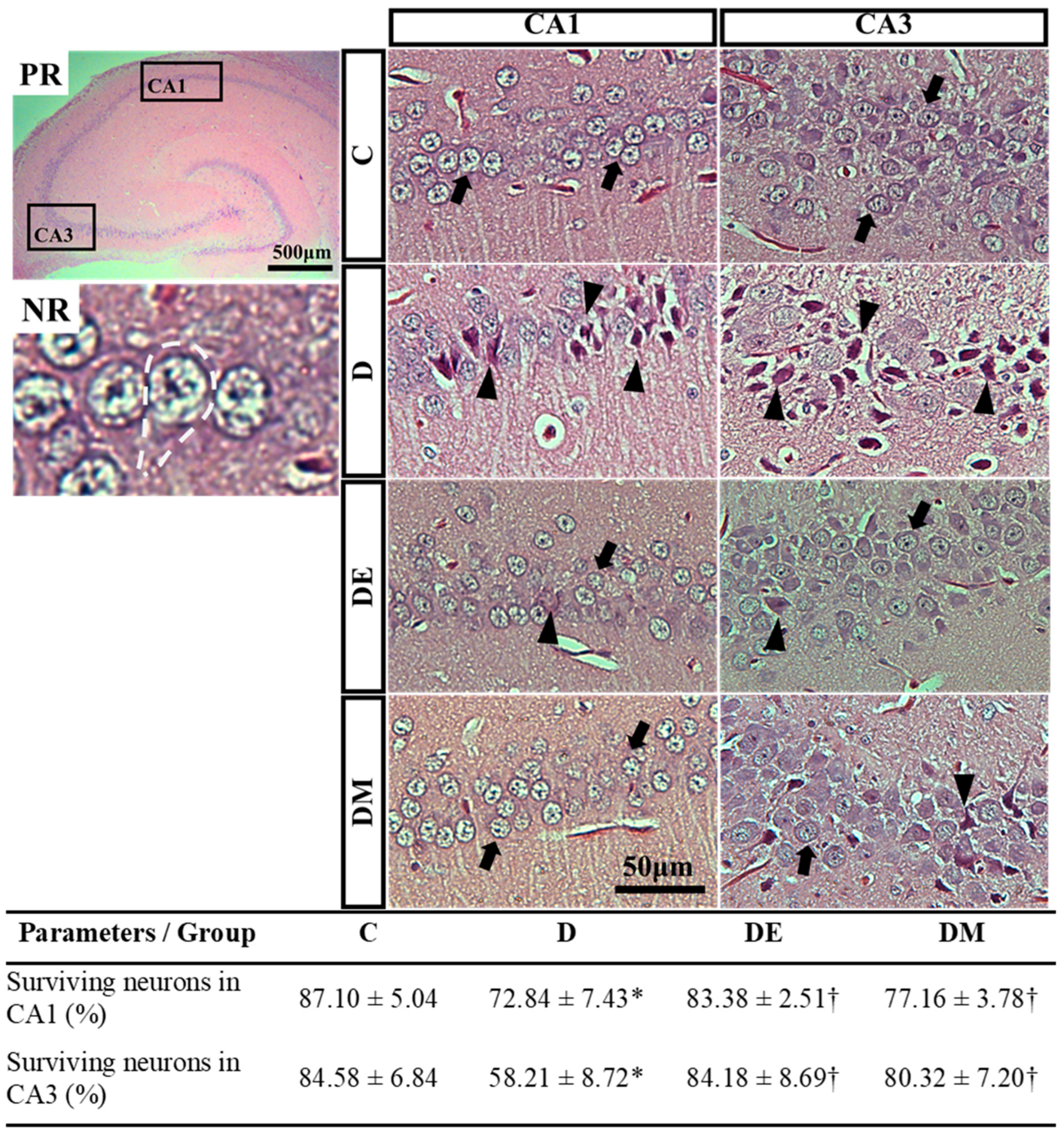
3.4. Effects of EE Exposure and Metformin Treatment on Spatial Learning and Memory Function, Serum BDNF, Histological Features, and Morphometric Measurement of Hippocampal Neurons in HFS Diet-Induced T2D Rats
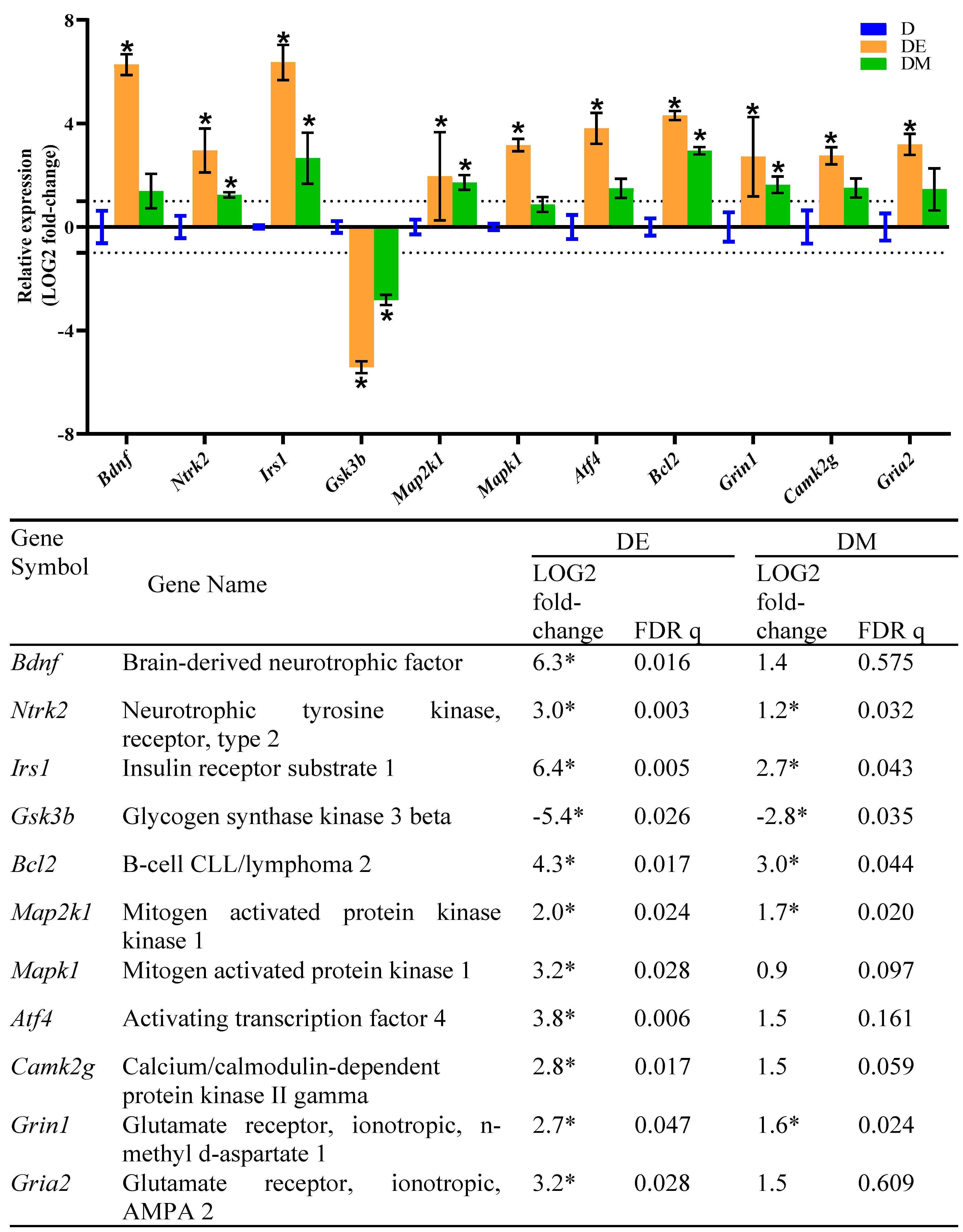
3.5. Effects of EE Exposure and Metformin Treatment on Gene Expression in the Hippocampus of HFS Diet-Induced T2D Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, M.A.B.; Hashim, M.J.; King, J.K.; Govender, R.D.; Mustafa, H.; Al Kaabi, J. Epidemiology of type 2 diabetes–global burden of disease and forecasted trends. J. Epidemiol. Glob. Health 2020, 10, 107. [Google Scholar] [CrossRef] [Green Version]
- Attal, K.; Wickman, J.A.; Kojima, S.; Blythe, S.N.; Toporikova, N. Time-restricted feeding drives periods of rapid food consumption in rats fed a high-fat diet with liquid sucrose. Obes. Med. 2021, 24, 100347. [Google Scholar] [CrossRef]
- Balasubramanian, P.; Kiss, T.; Tarantini, S.; Nyúl-Tóth, Á.; Ahire, C.; Yabluchanskiy, A.; Csipo, T.; Lipecz, A.; Tabak, A.; Institoris, A. Obesity-induced cognitive impairment in older adults: A microvascular perspective. Am. J. Physiol.-Heart Circ. Physiol. 2021, 320, H740–H761. [Google Scholar] [CrossRef]
- Di Liegro, C.M.; Schiera, G.; Proia, P.; Di Liegro, I. Physical activity and brain health. Genes 2019, 10, 720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannan, A.J. Environmental enrichment and brain repair: Harnessing the therapeutic effects of cognitive stimulation and physical activity to enhance experience-dependent plasticity. Neuropathol. Appl. Neurobiol. 2014, 40, 13–25. [Google Scholar] [CrossRef]
- Gergerlioglu, H.S.; Oz, M.; Demir, E.A.; Nurullahoglu-Atalik, K.E.; Yerlikaya, F.H. Environmental enrichment reverses cognitive impairments provoked by Western diet in rats: Role of corticosteroid receptors. Life Sci. 2016, 148, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Sampedro-Piquero, P.; Álvarez-Suárez, P.; Moreno-Fernández, R.D.; García-Castro, G.; Cuesta, M.; Begega, A. Environmental Enrichment Results in Both Brain Connectivity Efficiency and Selective Improvement in Different Behavioral Tasks. Neuroscience 2018, 388, 374–383. [Google Scholar] [CrossRef]
- Fischer, A. Environmental enrichment as a method to improve cognitive function. What can we learn from animal models? NeuroImage 2016, 131, 42–47. [Google Scholar] [CrossRef]
- Sritawan, N.; Suwannakot, K.; Naewla, S.; Chaisawang, P.; Aranarochana, A.; Sirichoat, A.; Pannangrong, W.; Wigmore, P.; Welbat, J.U. Effect of metformin treatment on memory and hippocampal neurogenesis decline correlated with oxidative stress induced by methotrexate in rats. Biomed. Pharmacother. 2021, 144, 112280. [Google Scholar] [CrossRef]
- Ponnulakshmi, R.; Shyamaladevi, B.; Vijayalakshmi, P.; Selvaraj, J. In silico and in vivo analysis to identify the antidiabetic activity of beta sitosterol in adipose tissue of high fat diet and sucrose induced type-2 diabetic experimental rats. Toxicol. Mech. Methods 2019, 29, 276–290. [Google Scholar] [CrossRef]
- Ismail, T.R.; Yap, C.G.; Naidu, R.; Pamidi, N. Enrichment Protocol for Rat Models. Curr. Protoc. 2021, 1, e152. [Google Scholar] [CrossRef] [PubMed]
- Vittalrao, A.M.; Chakraborti, A.; Banawalikar, N. Evaluation of vigilance promoting drugs modafinil and caffeine on cognition enhancing activities in Wistar albino rats—A comparative study. Biomed. Pharmacol. J. 2019, 12, 1463–1470. [Google Scholar] [CrossRef]
- Abubakar, M.B.; Abdullah, W.Z.; Sulaiman, S.A.; Ang, B.S. The effects of exposure to petrol vapours on growth, haematological parameters and oxidative markers in sprague-dawley male rats. Malays. J. Med. Sci. MJMS 2015, 22, 23. [Google Scholar]
- Rabiu, A.; Wale, H.; Garba, K.; Sabo, A.; Hassan, Z.; Shugaba, A.; Egesie, U.; Odeh, S. Body mass index of male and female Wistar rats following administration of leptin hormone after a dietary regime. Ann. Bioanthropol. 2017, 5, 22–26. [Google Scholar] [CrossRef]
- Leopoldo, A.S.; Lima-Leopoldo, A.P.; Nascimento, A.F.; Luvizotto, R.A.M.; Sugizaki, M.M.; Campos, D.H.S.; da Silva, D.C.T.; Padovani, C.R.; Cicogna, A.C. Classification of different degrees of adiposity in sedentary rats. Braz. J. Med. Biol. Res. Rev. Bras. Pesqui. Med. E Biol. 2016, 49, e5028. [Google Scholar] [CrossRef] [Green Version]
- Bulboacă, A.E.; Boarescu, P.M.; Bolboacă, S.D.; Blidaru, M.; Feștilă, D.; Dogaru, G.; Nicula, C.A. Comparative effect of curcumin versus liposomal curcumin on systemic pro-inflammatory cytokines profile, mcp-1 and rantes in experimental diabetes mellitus. Int. J. Nanomed. 2019, 14, 8961. [Google Scholar] [CrossRef] [Green Version]
- Carito, V.; Ceccanti, M.; Cestari, V.; Natella, F.; Bello, C.; Coccurello, R.; Mancinelli, R.; Fiore, M. Olive polyphenol effects in a mouse model of chronic ethanol addiction. Nutrition 2017, 33, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Seyedsadjadi, N.; Berg, J.; Bilgin, A.A.; Tung, C.; Grant, R. Significant relationships between a simple marker of redox balance and lifestyle behaviours; relevance to the Framingham risk score. PLoS ONE 2017, 12, e0187713. [Google Scholar] [CrossRef] [Green Version]
- Levada, O.A.; Cherednichenko, N.V.; Trailin, A.V.; Troyan, A.S. Plasma Brain-Derived Neurotrophic Factor as a Biomarker for the Main Types of Mild Neurocognitive Disorders and Treatment Efficacy: A Preliminary Study. Dis. Markers 2016, 2016, 4095723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratt, R.D.; Brickman, C.; Nawab, A.; Cottrill, C.; Snoad, B.; Lakhani, H.V.; Jelcick, A.; Henderson, B.; Bhardwaj, N.N.; Sanabria, J.R.; et al. The Adipocyte Na/K-ATPase Oxidant Amplification Loop is the Central Regulator of Western Diet-Induced Obesity and Associated Comorbidities. Sci. Rep. 2019, 9, 7927. [Google Scholar] [CrossRef] [Green Version]
- van Dam, A.D.; Nahon, K.J.; Kooijman, S.; van den Berg, S.M.; Kanhai, A.A.; Kikuchi, T.; Heemskerk, M.M.; van Harmelen, V.; Lombès, M.; van den Hoek, A.M.; et al. Salsalate Activates Brown Adipose Tissue in Mice. Diabetes 2014, 64, 1544–1554. [Google Scholar] [CrossRef] [Green Version]
- Wahba, N.S.; Shaban, S.F.; Kattaia, A.A.; Kandeel, S.A. Efficacy of zinc oxide nanoparticles in attenuating pancreatic damage in a rat model of streptozotocin-induced diabetes. Ultrastruct. Pathol. 2016, 40, 358–373. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.T.; Gilbert, J.M.; Paiz, J.H.; Bowman, F.M.; Blum, B. Endocrine cell type sorting and mature architecture in the islets of Langerhans require expression of Roundabout receptors in β cells. Sci. Rep. 2018, 8, 10876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Khair, D.M.A.; El-Safti, F.E.-N.A.; Nooh, H.Z.; El-Mehi, A.E. A comparative study on the effect of high cholesterol diet on the hippocampal CA1 area of adult and aged rats. Anat. Cell Biol. 2014, 47, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kalaivani, K.; Sankaranarayanan, C. Isopulegol Ameliorates Dyslipidemia by Modulating Adipokine Secretion in High Fat Diet/Streptozotocin Induced Diabetic Rats. J. Drug Deliv. Ther. 2019, 9, 126–136. [Google Scholar] [CrossRef]
- El-Shiekh, R.A.; Al-Mahdy, D.A.; Mouneir, S.M.; Hifnawy, M.S.; Abdel-Sattar, E.A. Anti-obesity effect of argel (Solenostemma argel) on obese rats fed a high fat diet. J. Ethnopharmacol. 2019, 238, 111893. [Google Scholar] [CrossRef]
- Mehta, B.K.; Banerjee, S. Characterization of Cognitive Impairment in Type 2 Diabetic Rats. Indian J. Pharm. Sci. 2017, 79, 785–793. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E. Effect of High-Fat Diets on Oxidative Stress, Cellular Inflammatory Response and Cognitive Function. Nutrients 2019, 11, 2579. [Google Scholar] [CrossRef] [Green Version]
- Vadini, F.; Simeone, P.G.; Boccatonda, A.; Guagnano, M.T.; Liani, R.; Tripaldi, R.; Di Castelnuovo, A.; Cipollone, F.; Consoli, A.; Santilli, F. Liraglutide improves memory in obese patients with prediabetes or early type 2 diabetes: A randomized, controlled study. Int. J. Obes. 2020, 44, 1254–1263. [Google Scholar] [CrossRef]
- de Souza, R.M.; de Souza, L.; Machado, A.E.; de Bem Alves, A.C.; Rodrigues, F.S.; Aguiar, A.S.; Dos Santos, A.R.S.; de Bem, A.F.; Moreira, E.L.G. Behavioural, metabolic and neurochemical effects of environmental enrichment in high-fat cholesterol-enriched diet-fed mice. Behav. Brain Res. 2019, 359, 648–656. [Google Scholar] [CrossRef]
- Sgrò, P.; Emerenziani, G.P.; Antinozzi, C.; Sacchetti, M.; Di Luigi, L. Exercise as a drug for glucose management and prevention in type 2 diabetes mellitus. Curr. Opin. Pharmacol. 2021, 59, 95–102. [Google Scholar] [CrossRef]
- Borror, A.; Zieff, G.; Battaglini, C.; Stoner, L. The Effects of Postprandial Exercise on Glucose Control in Individuals with Type 2 Diabetes: A Systematic Review. Sport. Med. 2018, 48, 1479–1491. [Google Scholar] [CrossRef]
- Vela-Guajardo, J.E.; Garza-González, S.; García, N. Glucolipotoxicity-induced oxidative stress is related to mitochondrial dysfunction and apoptosis of pancreatic β-cell. Curr. Diabetes Rev. 2021, 17, 46–56. [Google Scholar] [CrossRef]
- Moraes-Silva, I.C.; Mostarda, C.; Moreira, E.D.; Silva, K.A.S.; dos Santos, F.; de Angelis, K.; Farah, V.d.M.A.; Irigoyen, M.C. Preventive role of exercise training in autonomic, hemodynamic, and metabolic parameters in rats under high risk of metabolic syndrome development. J. Appl. Physiol. 2013, 114, 786–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, M.V.; Vieira, A.B.; da Conceição, F.G.; Nascimento, A.R.; da Nóbrega, A.C.L.; Tibirica, E. Exercise training dose differentially alters muscle and heart capillary density and metabolic functions in an obese rat with metabolic syndrome. Exp. Physiol. 2017, 102, 1716–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xu, D. Effects of aerobic exercise on lipids and lipoproteins. Lipids Health Dis. 2017, 16, 132. [Google Scholar] [CrossRef] [Green Version]
- Lehnig, A.C.; Dewal, R.S.; Baer, L.A.; Kitching, K.M.; Munoz, V.R.; Arts, P.J.; Sindeldecker, D.A.; May, F.J.; Lauritzen, H.P.M.M.; Goodyear, L.J.; et al. Exercise Training Induces Depot-Specific Adaptations to White and Brown Adipose Tissue. iScience 2019, 11, 425–439. [Google Scholar] [CrossRef] [Green Version]
- Peres Valgas da Silva, C.; Hernández-Saavedra, D.; White, J.D.; Stanford, K.I. Cold and exercise: Therapeutic tools to activate brown adipose tissue and combat obesity. Biology 2019, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.H.; Kang, S.-W.; Kim, K.; Wi, S.; Lee, J.W.; Cho, S.-R. Environmental Enrichment Attenuates Oxidative Stress and Alters Detoxifying Enzymes in an A53T α-Synuclein Transgenic Mouse Model of Parkinson’s Disease. Antioxidants 2020, 9, 928. [Google Scholar] [CrossRef]
- Müller, P.; Duderstadt, Y.; Lessmann, V.; Müller, N.G. Lactate and BDNF: Key Mediators of Exercise Induced Neuroplasticity? J. Clin. Med. 2020, 9, 1136. [Google Scholar] [CrossRef]
- Soltesz, I.; Losonczy, A. CA1 pyramidal cell diversity enabling parallel information processing in the hippocampus. Nat. Neurosci. 2018, 21, 484–493. [Google Scholar] [CrossRef] [Green Version]
- Leal, G.; Bramham, C.; Duarte, C. BDNF and hippocampal synaptic plasticity. Vitam. Horm. 2017, 104, 153–195. [Google Scholar]
- Babri, S.; Habibi, P.; Nouri, F.; Khazaei, M.; Nayebi Rad, S.; Javani, G. Protective effect of swimming and genistein on the expression of microRNA 132, insulin growth factor 1, and brain-derived neurotrophic factor genes, as well as spatial memory, in the hippocampus of diabetic ovariectomized rats. Avicenna J. Neuropsychophysiol. 2021, 8, 178–185. [Google Scholar] [CrossRef]
- Yang, L.; Wang, H.; Liu, L.; Xie, A. The Role of Insulin/IGF-1/PI3K/Akt/GSK3β Signaling in Parkinson’s Disease Dementia. Front. Neurosci. 2018, 12, 73. [Google Scholar] [CrossRef] [Green Version]
- Tanokashira, D.; Fukuokaya, W.; Taguchi, A. Involvement of insulin receptor substrates in cognitive impairment and Alzheimer’s disease. Neural Regen. Res. 2019, 14, 1330. [Google Scholar]
- Srivastava, P.; Dhuriya, Y.K.; Kumar, V.; Srivastava, A.; Gupta, R.; Shukla, R.K.; Yadav, R.S.; Dwivedi, H.N.; Pant, A.B.; Khanna, V.K. PI3K/Akt/GSK3β induced CREB activation ameliorates arsenic mediated alterations in NMDA receptors and associated signaling in rat hippocampus: Neuroprotective role of curcumin. Neurotoxicology 2018, 67, 190–205. [Google Scholar] [CrossRef] [PubMed]
- Pasini, S.; Corona, C.; Liu, J.; Greene, L.A.; Shelanski, M.L. Specific downregulation of hippocampal ATF4 reveals a necessary role in synaptic plasticity and memory. Cell Rep. 2015, 11, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Sritawan, N.; Prajit, R.; Chaisawang, P.; Sirichoat, A.; Pannangrong, W.; Wigmore, P.; Welbat, J.U. Metformin alleviates memory and hippocampal neurogenesis decline induced by methotrexate chemotherapy in a rat model. Biomed. Pharmacother. 2020, 131, 110651. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT2 qPCR Primer Assay Catalogue No. | Gene | NCBI Reference Sequence: | Official Name |
|---|---|---|---|
| PPR45425C | Akt1 | NM_033230 | v-akt murine thymoma viral oncogene homolog 1 |
| PPR42749A | Atf4 | NM_024403 | activating transcription factor 4 |
| PPR06577B | Bcl2 | NM_016993 | B-cell CLL/lymphoma 2 |
| PPR45333A | Bdnf | NM_012513 | Brain-derived neurotrophic factor |
| PPR43356B | Calm1 | NM_001007614 | calmodulin 1 |
| PPR45142B | Camk2g | NM_133605 | calcium/calmodulin-dependent protein kinase II gamma |
| PPR43063B | Grb2 | NM_030846 | growth factor receptor-bound protein 2 |
| PPR06753B | Gria2 | NM_001083811 | glutamate receptor, ionotropic, AMPA 2 |
| PPR06847D | Grin1 | NM_017010 | glutamate receptor, ionotropic, N-methyl D-aspartate 1 |
| PPR44848A | Gsk3b | NM_032080 | glycogen synthase kinase 3 beta |
| PPR44872B | Irs1 | NM_012969 | insulin receptor substrate 1 |
| PPR47860F | Kras | NM_031515 | KRAS proto-oncogene, GTPase |
| PPR43465A | Map2k1 | NM_031643 | mitogen-activated protein kinase 1 |
| PPR48780A | Mapk1 | NM_053842 | mitogen-activated protein kinase 1 |
| PPR45322B | Ntrk2 | NM_001163168 | neurotrophic tyrosine kinase, receptor, type 2 |
| PPR59984F | Pik3ca | NM_133399 | phosphatidylinositol-4,5-bisphosphate 3-kinase, catalytic subunit alpha |
| PPR45212A | Sh2b1 | NM_001048180 | SH2B adaptor protein 1 |
| PPR06570C | Actb | NM_031144 | actin, beta |
| PPR42247F | Hprt1 | NM_012583 | hypoxanthine phosphoribosyltransferase 1 |
| PPR56649C | Pgk1 | NM_053291 | phosphoglycerate kinase 1 |
| PPR47412A | Tbp | NM_001004198 | TATA box binding protein |
| PPR63338A | GDC | Genomic DNA control | |
| PPX63339A | PPC | PCR positive control | |
| PPX63340A | RTC | Reverse transcription control |
| Parameters/Group | C | D | DE | DM |
|---|---|---|---|---|
| FBG (mmol/L) | 5.90 ± 0.16 | 8.26 ± 0.48 * | 6.76 ± 0.10 † | 6.76 ± 0.34 † |
| C-peptide (ng/mL) | 26.19 ± 13.65 | 94.30 ± 26.26 * | 53.03 ± 33.31 † | 40.35 ± 22.15 † |
| Parameters/Group | C | D | DE | DM |
|---|---|---|---|---|
| TG (mmol/L) | 0.98 (0.97–1.13) | 2.54 (1.97–3.48) * | 1.58 (1.22–1.66) † | 1.69 (1.20–1.78) † |
| TC (mmol/L) | 2.59 (2.59–2.60) | 3.25 (3.14–3.37) * | 2.63 (2.61–2.64) † | 2.69 (2.66–2.78) † |
| HDL-C (mmol/L) | 1.98 (1.87–2.05) | 0.87 (0.68–1.24) * | 1.28 (1.24–1.34) | 1.30 (1.29–1.39) |
| Initial Body Weight (g) | 240.7 ± 18.62 | 258.9 ± 37.52 | 271.5 ± 28.89 | 265.7 ± 43.99 |
| Final Body Weight (g) | 385.9 ± 33.75 | 509.0 ± 48.85 * | 437.4 ± 21.80 † | 457.7 ± 59.02 |
| PWG (%) | 60.3 ± 8.60 | 98.0 ± 13.12 * | 62.0 ± 9.63 † | 73.4 ± 13.91 † |
| BMI (g/cm2) | 0.71 ± 0.04 | 0.87 ± 0.03 * | 0.74 ± 0.02 † | 0.78 ± 0.03 † |
| AI (%) | 3.76 ± 0.75 | 8.91 ± 0.56 * | 4.87 ± 0.46 † | 5.51 ± 1.17 † |
| Parameters/Group | C | D | DE | DM |
|---|---|---|---|---|
| FORT (mmol H2O2 Eq/L) | 2.03 ± 0.10 | 2.91 ± 0.18 * | 2.49 ± 0.03 † | 2.49 ± 0.09 † |
| FORD (mmol/L Trolox Eq/L) | 1.43 ± 0.16 | 1.43 ± 0.15 | 1.64 ± 0.24 | 1.44 ± 0.17 |
| Redox balance | 0.71 ± 0.05 | 0.49 ± 0.03 * | 0.66 ± 0.09 † | 0.58 ± 0.05 |
| Parameters/Group | C | D | DE | DM |
|---|---|---|---|---|
| Spontaneous alternation (%) | 84.52 ± 6.234 | 67.86 ± 4.64 * | 85.12 ± 5.30 † | 85.71 ± 5.82 † |
| Correct response (%) | 86.31 ± 5.75 | 67.26 ± 3.75 * | 80.36 ± 4.64 † | 79.76 ± 2.87 † |
| Serum BDNF (ng/mL) | 14.45 ± 3.43 | 9.55 ± 1.67 * | 14.89 ± 1.00 † | 15.35 ± 2.27 † |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ismail, T.R.; Yap, C.G.; Naidu, R.; Pamidi, N. Environmental Enrichment and Metformin Improve Metabolic Functions, Hippocampal Neuron Survival, and Hippocampal-Dependent Memory in High-Fat/High-Sucrose Diet-Induced Type 2 Diabetic Rats. Biology 2023, 12, 480. https://doi.org/10.3390/biology12030480
Ismail TR, Yap CG, Naidu R, Pamidi N. Environmental Enrichment and Metformin Improve Metabolic Functions, Hippocampal Neuron Survival, and Hippocampal-Dependent Memory in High-Fat/High-Sucrose Diet-Induced Type 2 Diabetic Rats. Biology. 2023; 12(3):480. https://doi.org/10.3390/biology12030480
Chicago/Turabian StyleIsmail, Teh Rasyidah, Christina Gertrude Yap, Rakesh Naidu, and Narendra Pamidi. 2023. "Environmental Enrichment and Metformin Improve Metabolic Functions, Hippocampal Neuron Survival, and Hippocampal-Dependent Memory in High-Fat/High-Sucrose Diet-Induced Type 2 Diabetic Rats" Biology 12, no. 3: 480. https://doi.org/10.3390/biology12030480