Calumenin, a Ca2+ Binding Protein, Is Required for Dauer Formation in Caenorhabditis elegans


{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. C. elegans Strains and Cultivation
2.2. Temporal Expression of calu-1 by Quantitative RT-PCR
2.3. Construct and Microscopy
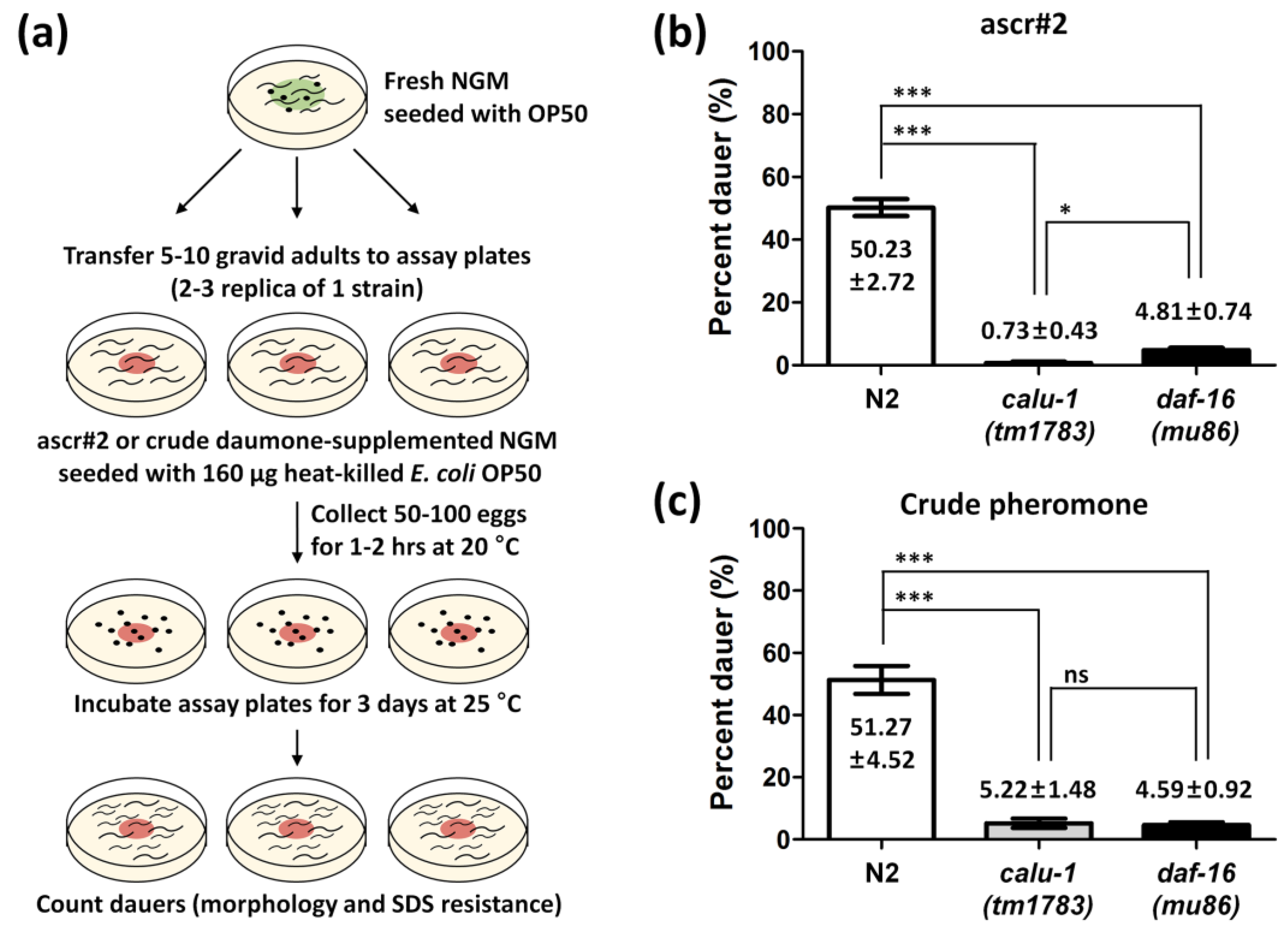
2.4. Dauer Formation Assay
2.5. Statistical Analysis
3. Results
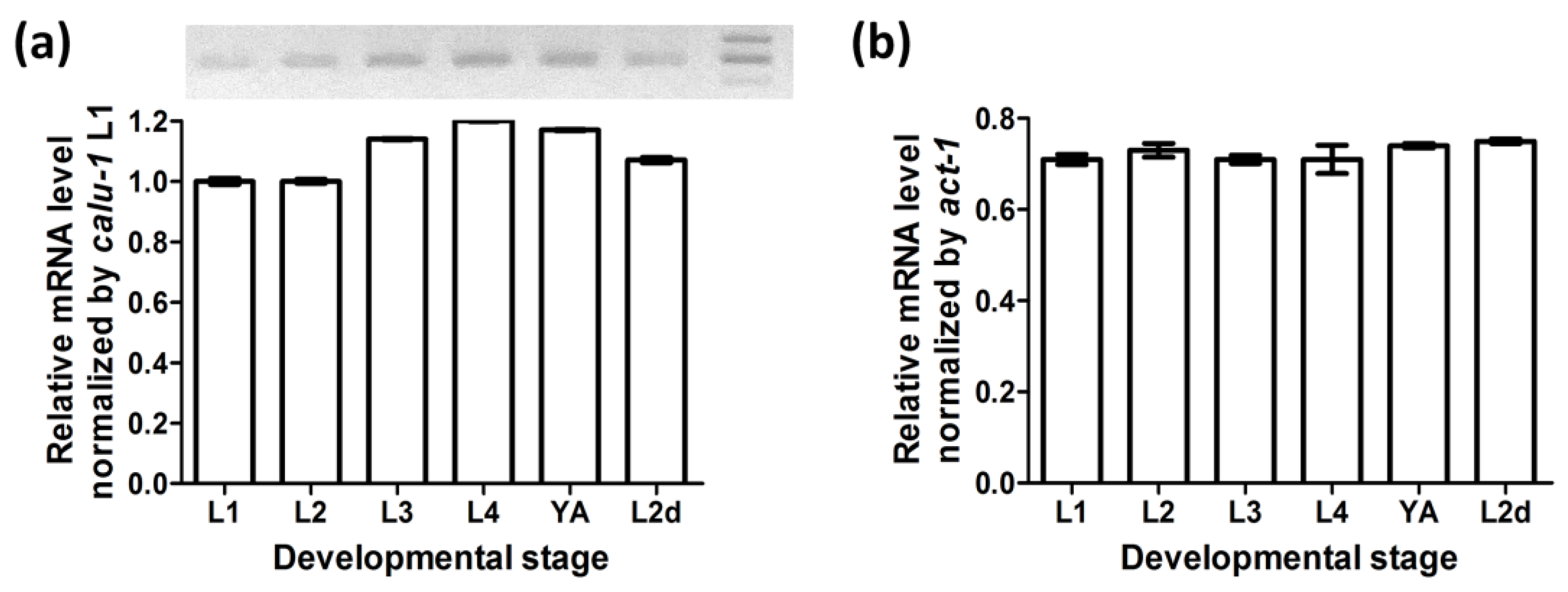
3.1. Calu-1 Is Expressed throughout Development—Including the Dauer Stage
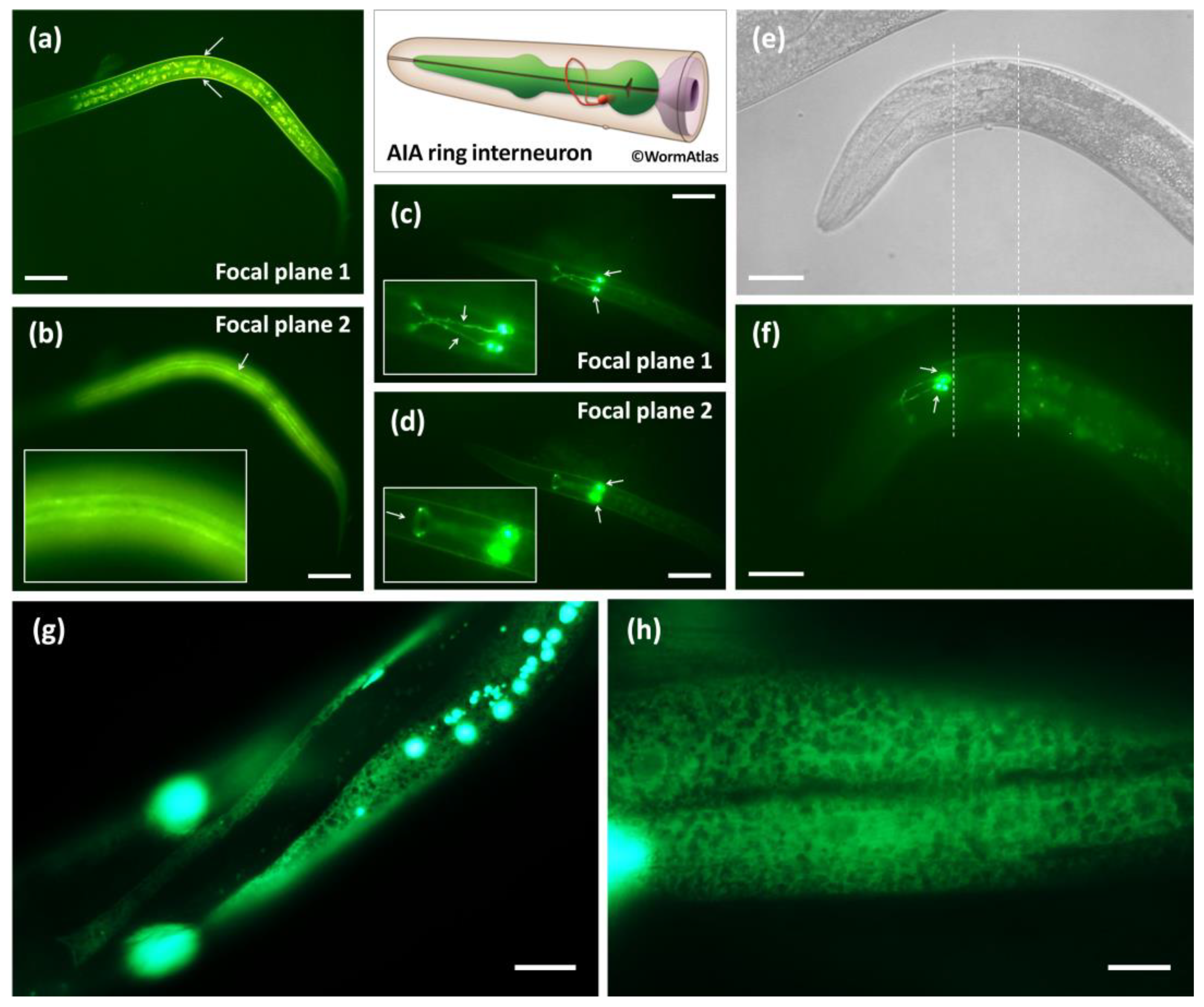
3.2. Calu-1 Is Expressed in Both Neuron and Effector Tissues That Mediate Dauer Entry
3.3. The calu-1(tm1783) Mutant Is Dauer-defective
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cassada, R.C.; Russell, R.L. The dauerlarva, a post-embryonic developmental variant of the nematode Caenorhabditis elegans. Dev. Biol. 1975, 46, 326–342. [Google Scholar] [CrossRef]
- Kelly, S.A.; Panhuis, T.M.; Stoehr, A.M. Phenotypic plasticity: Molecular mechanisms and adaptive significance. Compr. Physiol. 2012, 2, 1417–1439. [Google Scholar] [PubMed]
- Golden, J.W.; Riddle, D.L. A pheromone influences larval development in the nematode Caenorhabditis elegans. Science 1982, 218, 578–580. [Google Scholar] [CrossRef] [PubMed]
- Kramer, J.M. Extracellular Matrix. In C. elegans II, 2nd ed.; Riddle, D.L., Blumenthal, T., Meyer, B.J., Priess, J.R., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Wollow, C.A.; Hall, D.H. The Dauer Cuticle. WormAtlas. 2007. Available online: https://www.wormatlas.org/dauer/cuticle/Cutframeset.html (accessed on 22 February 2023).
- Albert, P.S.; Riddle, D.L. Developmental alterations in sensory neuroanatomy of the Caenorhabditis elegans dauer larva. J. Comp. Neurol. 1983, 219, 461–481. [Google Scholar] [CrossRef] [PubMed]
- Jeong, P.Y.; Jung, M.; Yim, Y.H.; Kim, H.; Park, M.; Hong, E.; Lee, W.; Kim, Y.H.; Kim, K.; Paik, Y.K. Chemical structure and biological activity of the Caenorhabditis elegans dauer-inducing pheromone. Nature 2005, 433, 541–545. [Google Scholar] [CrossRef]
- Butcher, R.A.; Fujita, M.; Schroeder, F.C.; Clardy, J. Small-molecule pheromones that control dauer development in Caenorhabditis elegans. Nat. Chem. Biol. 2007, 3, 420–422. [Google Scholar] [CrossRef]
- Bargmann, C.I.; Horvitz, H.R. Control of larval development by chemosensory neurons in Caenorhabditis elegans. Science 1991, 251, 1243–1246. [Google Scholar] [CrossRef]
- Kim, K.; Sato, K.; Shibuya, M.; Zeiger, D.M.; Butcher, R.A.; Ragains, J.R.; Clardy, J.; Touhara, K.; Sengupta, P. Two chemoreceptors mediate developmental effects of dauer pheromone in C. elegans. Science 2009, 326, 994–998. [Google Scholar] [CrossRef] [Green Version]
- Schackwitz, W.S.; Inoue, T.; Thomas, J.H. Chemosensory neurons function in parallel to mediate a pheromone response in C. elegans. Neuron 1996, 17, 719–728. [Google Scholar] [CrossRef] [Green Version]
- Fielenbach, N.; Antebi, A. C. elegans dauer formation and the molecular basis of plasticity. Genes Dev. 2008, 22, 2149–2165. [Google Scholar] [CrossRef] [Green Version]
- Gerisch, B.; Weitzel, C.; Kober-Eisermann, C.; Rottiers, V.; Antebi, A. A hormonal signaling pathway influencing C. elegans metabolism, reproductive development, and life span. Dev. Cell 2001, 1, 841–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottlieb, S.; Ruvkun, G. daf-2, daf-16 and daf-23: Genetically interacting genes controlling dauer formation in Caenorhabditis elegans. Genetics 1994, 137, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.; Albert, P.S.; Riddle, D.L. DAF-9, a cytochrome P450 regulating C. elegans larval development and adult longevity. Development 2002, 129, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Riddle, D.L.; Swanson, M.M.; Albert, P.S. Interacting genes in nematode dauer larva formation. Nature 1981, 290, 668–671. [Google Scholar] [CrossRef]
- Thomas, J.H.; Birnby, D.A.; Vowels, J.J. Evidence for parallel processing of sensory information controlling dauer formation in Caenorhabditis elegans. Genetics 1993, 134, 1105–1117. [Google Scholar] [CrossRef]
- Vowels, J.J.; Thomas, J.H. Genetic analysis of chemosensory control of dauer formation in Caenorhabditis elegans. Genetics 1992, 130, 105–123. [Google Scholar] [CrossRef]
- Honore, B. The rapidly expanding CREC protein family: Members, localization, function, and role in disease. Bioessays 2009, 31, 262–277. [Google Scholar] [CrossRef]
- Wu, W.; Tang, X.; Hu, W.; Lotan, R.; Hong, W.K.; Mao, L. Identification and validation of metastasis-associated proteins in head and neck cancer cell lines by two-dimensional electrophoresis and mass spectrometry. Clin. Exp. Metastasis 2002, 19, 319–326. [Google Scholar] [CrossRef]
- Voisin, S.N.; Krakovska, O.; Matta, A.; DeSouza, L.V.; Romaschin, A.D.; Colgan, T.J.; Siu, K.W. Identification of novel molecular targets for endometrial cancer using a drill-down LC-MS/MS approach with iTRAQ. PLoS ONE 2011, 6, e16352. [Google Scholar] [CrossRef] [Green Version]
- Torres, S.; Bartolome, R.A.; Mendes, M.; Barderas, R.; Fernandez-Acenero, M.J.; Pelaez-Garcia, A.; Pena, C.; Lopez-Lucendo, M.; Villar-Vazquez, R.; de Herreros, A.G.; et al. Proteome profiling of cancer-associated fibroblasts identifies novel proinflammatory signatures and prognostic markers for colorectal cancer. Clin. Cancer Res. 2013, 19, 6006–6019. [Google Scholar] [CrossRef] [Green Version]
- Kurpinska, A.; Suraj, J.; Bonar, E.; Zakrzewska, A.; Stojak, M.; Sternak, M.; Jasztal, A.; Walczak, M. Proteomic characterization of early lung response to breast cancer metastasis in mice. Exp. Mol. Pathol. 2019, 107, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, S.K.; Kim, D.H. Characterization of calumenin in mouse heart. BMB Rep. 2010, 43, 158–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahoo, S.K.; Kim, D.H. Calumenin interacts with SERCA2 in rat cardiac sarcoplasmic reticulum. Mol. Cells 2008, 26, 265–269. [Google Scholar] [PubMed]
- Cho, J.H.; Song, H.O.; Singaravelu, G.; Sung, H.; Oh, W.C.; Kwon, S.; Kim, D.H.; Ahnn, J. Pleiotropic roles of calumenin (calu-1), a calcium-binding ER luminal protein, in Caenorhabditis elegans. FEBS Lett. 2009, 583, 3050–3056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Altun, Z.F.; Hall, D.H. Introduction to C. elegans Anatomy. WormAtlas. 2009. Available online: https://www.wormatlas.org/hermaphrodite/introduction/Introframeset.html (accessed on 22 February 2023).
- Sapio, M.R.; Hilliard, M.A.; Cermola, M.; Favre, R.; Bazzicalupo, P. The Zona Pellucida domain containing proteins, CUT-1, CUT-3 and CUT-5, play essential roles in the development of the larval alae in Caenorhabditis elegans. Dev. Biol. 2005, 282, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Kim, K.Y.; Joo, H.J.; Kim, H.; Jeong, P.Y.; Paik, Y.K. Methods for evaluating the Caenorhabditis elegans dauer state: Standard dauer-formation assay using synthetic daumones and proteomic analysis of O-GlcNAc modifications. Methods Cell Biol. 2011, 106, 445–460. [Google Scholar]
- Park, J.Y.; Cheong, M.C.; Cho, J.Y.; Koo, H.S.; Paik, Y.K. A novel functional cross-interaction between opioid and pheromone signaling may be involved in stress avoidance in Caenorhabditis elegans. Sci. Rep. 2020, 10, 7524. [Google Scholar] [CrossRef]
- Schroeder, N.E.; Flatt, K.M. In vivo imaging of Dauer-specific neuronal remodeling in C. elegans. J. Vis. Exp. 2014, 91, e51834. [Google Scholar]
- Chai, C.M.; Torkashvand, M.; Seyedolmohadesin, M.; Park, H.; Venkatachalam, V.; Sternberg, P.W. Interneuron control of C. elegans developmental decision-making. Curr. Biol. 2022, 32, 2316–2324.e4. [Google Scholar] [CrossRef]
- Altun, Z.F.; Herndon, L.A.; Wolkow, C.A.; Crocker, C.; Lints, R.; Hall, D.H. WormAtlas. 2002–2023. Available online: http://www.wormatlas.org (accessed on 22 February 2023).
- Yabe, D.; Nakamura, T.; Kanazawa, N.; Tashiro, K.; Honjo, T. Calumenin, a Ca2+-binding protein retained in the endoplasmic reticulum with a novel carboxyl-terminal sequence, HDEF. J. Biol. Chem. 1997, 272, 18232–18239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorum, H.; Hager, H.; Christensen, B.M.; Nielsen, S.; Honore, B. Human calumenin localizes to the secretory pathway and is secreted to the medium. Exp. Cell Res. 1999, 248, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Coppinger, J.A.; Cagney, G.; Toomey, S.; Kislinger, T.; Belton, O.; McRedmond, J.P.; Cahill, D.J.; Emili, A.; Fitzgerald, D.J.; Maguire, P.B. Characterization of the proteins released from activated platelets leads to localization of novel platelet proteins in human atherosclerotic lesions. Blood 2004, 103, 2096–2104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Feng, H.; Zheng, P.; Shen, B.; Chen, L.; Liu, L.; Liu, X.; Hao, Q.; Wang, S.; Chen, J.; et al. The intracellular transport and secretion of calumenin-1/2 in living cells. PLoS ONE 2012, 7, e35344. [Google Scholar] [CrossRef] [PubMed]
- Jung, D.H.; Mo, S.H.; Kim, D.H. Calumenin, a multiple EF-hands Ca2+-binding protein, interacts with ryanodine receptor-1 in rabbit skeletal sarcoplasmic reticulum. Biochem. Biophys. Res. Commun. 2006, 343, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Shen, B.; Chen, L.; Zheng, P.; Feng, H.; Hao, Q.; Liu, X.; Liu, L.; Xu, S.; Chen, J.; et al. Extracellular calumenin suppresses ERK1/2 signaling and cell migration by protecting fibulin-1 from MMP-13-mediated proteolysis. Oncogene 2015, 34, 1006–1018. [Google Scholar] [CrossRef]
- Choi, T.W.; Cho, J.H.; Ahnn, J.; Song, H.O. Novel Findings of Anti-Filarial Drug Target and Structure-Based Virtual Screening for Drug Discovery. Int. J. Mol. Sci. 2018, 19, 3579. [Google Scholar] [CrossRef] [Green Version]
- Barret, A.; Varol, E.; Weinreb, A.; Taylor, S.R.; McWhirter, R.M.; Cros, C.; Basaravaju, M.; Poff, A.; Tipps, J.A.; Majeed, M.; et al. Integrating bulk and single cell RNa-seq refines transcriptomic profiles of specific C. elegans neurons. bioRxiv 2022. [Google Scholar] [CrossRef]
- White, J.G.; Southgate, E.; Thomson, J.N.; Brenner, S. The structure of the nervous system of the nematode Caenorhabditis elegans. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1986, 314, 1–340. [Google Scholar] [PubMed]
- Wakabayashi, T.; Kitagawa, I.; Shingai, R. Neurons regulating the duration of forward locomotion in Caenorhabditis elegans. Neurosci. Res. 2004, 50, 103–111. [Google Scholar] [CrossRef]
- Shinkai, Y.; Yamamoto, Y.; Fujiwara, M.; Tabata, T.; Murayama, T.; Hirotsu, T.; Ikeda, D.D.; Tsunozaki, M.; Iino, Y.; Bargmann, C.I.; et al. Behavioral choice between conflicting alternatives is regulated by a receptor guanylyl cyclase, GCY-28, and a receptor tyrosine kinase, SCD-2, in AIA interneurons of Caenorhabditis elegans. J. Neurosci. 2011, 31, 3007–3015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomioka, M.; Adachi, T.; Suzuki, H.; Kunitomo, H.; Schafer, W.R.; Iino, Y. The insulin/PI 3-kinase pathway regulates salt chemotaxis learning in Caenorhabditis elegans. Neuron 2006, 51, 613–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrity, P.A.; Goodman, M.B.; Samuel, A.D.; Sengupta, P. Running hot and cold: Behavioral strategies, neural circuits, and the molecular machinery for thermotaxis in C. elegans and Drosophila. Genes Dev. 2010, 24, 2365–2382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, J.; von Reuss, S.H.; Bose, N.; Zaslaver, A.; Mahanti, P.; Ho, M.C.; O’Doherty, O.G.; Edison, A.S.; Sternberg, P.W.; Schroeder, F.C. A modular library of small molecule signals regulates social behaviors in Caenorhabditis elegans. PLoS Biol. 2012, 10, e1001237. [Google Scholar] [CrossRef] [Green Version]
- Macosko, E.Z.; Pokala, N.; Feinberg, E.H.; Chalasani, S.H.; Butcher, R.A.; Clardy, J.; Bargmann, C.I. A hub-and-spoke circuit drives pheromone attraction and social behaviour in C. elegans. Nature 2009, 458, 1171–1175. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Kim, K. Neuropeptides. Wormbook 2008, 25, 1–36. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.E.; Cho, J.H.; Song, H.-O. Calumenin, a Ca2+ Binding Protein, Is Required for Dauer Formation in Caenorhabditis elegans. Biology 2023, 12, 464. https://doi.org/10.3390/biology12030464
Lee KE, Cho JH, Song H-O. Calumenin, a Ca2+ Binding Protein, Is Required for Dauer Formation in Caenorhabditis elegans. Biology. 2023; 12(3):464. https://doi.org/10.3390/biology12030464
Chicago/Turabian StyleLee, Kyung Eun, Jeong Hoon Cho, and Hyun-Ok Song. 2023. "Calumenin, a Ca2+ Binding Protein, Is Required for Dauer Formation in Caenorhabditis elegans" Biology 12, no. 3: 464. https://doi.org/10.3390/biology12030464