
Antibiotic Resistance, Virulence Gene Detection, and Biofilm Formation in Aeromonas spp. Isolated from Fish and Humans in Egypt
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Sample Collection
2.2. Isolation and Identification of Aeromonas spp.
2.3. Molecular Identification and Virulence Genes Detection
2.4. Antimicrobial Susceptibility Test
2.5. Biofilm Formation Assay
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morshdy, A.E.M.; Abdelhameed, N.S.A.; El Bayomi, R.M.; Abdallah, K. Prevalence of Antibiotic Resistant Aeromonas and Molecular Identification of Aeromonas hydrophila Isolated from Some Marketed Fish in Egypt. J. Adv. Vet. Res. 2022, 12, 717–721. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Carvalho, M.J.; Martínez-Murcia, A.; Esteves, A.C.; Correia, A.; Saavedra, M.J. Phylogenetic diversity, antibiotic resistance and virulence traits of Aeromonas spp. from untreated waters for human consumption. Int. J. Food Microbiol. 2012, 159, 230–239. [Google Scholar] [CrossRef]
- Dong, H.T.; Techatanakitarnan, C.; Jindakittikul, P.; Thaiprayoon, A.; Taengphu, S.; Charoensapsri, W.; Khunrae, P.; Rattanarojpong, T.; Senapin, S. Aeromonas jandaei and Aeromonas veronii caused disease and mortality in Nile tilapia, Oreochromis niloticus (L.). J. Fish Dis. 2017, 40, 1395–1403. [Google Scholar] [CrossRef]
- Nahar, S.; Rahman, M.M.; Ahmed, G.U.; Faruk, M.A.R. Isolation, identification, and characterization of Aeromonas hydrophila from juvenile farmed pangasius (Pangasianodon hypophthalmus). Int. J. Fish. Aquat. Sci. 2016, 4, 52–60. [Google Scholar]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [Green Version]
- Plumb, J.A.; Hanson, L.A. Health Maintenance and Principal Microbial Diseases of Cultured Fishes; John Wiley & Sons: Hoboken, NJ, USA, 2010; p. 483. [Google Scholar]
- Abdel-Latif, H.M.; Khafaga, A.F. Natural co-infection of cultured Nile tilapia Oreochromis niloticus with Aeromonas hydrophila and Gyrodactylus cichlidarum experiencing high mortality during summer. Aquac. Res. 2020, 51, 1880–1892. [Google Scholar] [CrossRef]
- Stratev, D.; Odeyemi, O.A. An overview of motile Aeromonas septicaemia management. Aquac. Int. 2017, 25, 1095–1105. [Google Scholar] [CrossRef]
- Hu, M.; Wang, N.; Pan, Z.; Lu, C.; Liu, Y. Identity and Virulence Properties of Aeromonas isolates from diseased fish, Healthy Controls and Water Environment in China. Lett. Appl. Microbiol. 2012, 55, 224–233. [Google Scholar] [CrossRef]
- Salunke, G.; Namshikar, V.; Gaonkar, R.; Gaonkar, T. A case of Aeromonas hydrophila meningitis in septic shock. Trop. J. Med. Res. 2015, 18, 54–57. [Google Scholar] [CrossRef]
- Beaz-Hidalgo, R.; Figueras, M.J. Aeromonas spp. whole genomes and virulence factors implicated in fish disease. J. Fish Dis. 2013, 36, 371–388. [Google Scholar] [CrossRef]
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef]
- Odeyemi, O.; Asmat, A.; Usup, G. Antibiotics resistance and putative virulence factors of Aeromonas hydrophila isolated from estuary. J. Microbiol. Biotechnol. Food Sci. 2012, 1, 1339. [Google Scholar]
- Sharma, I.; Rabha, D.; Das, S.; Ningombam, D. Hemolytic activity and antibiotic resistance of Aeromonas sp. isolated from marketed fish. Eur. J. Pharm. Med. Res. 2015, 2, 304–312. [Google Scholar]
- Odeyemi, O.A.; Ahmad, A. Antibiotic resistance profiling and phenotyping of Aeromonas species isolated from aquatic sources. Saudi J. Bio. Sci. 2017, 24, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Didugu, H.; Krishnaiah, N.; Rao, M.T.; Ramanipushpa, R.N. Antibiogram of Aeromonas Species Isolated From Livestock Products. Int. J. Sci. Environ. Technol. 2016, 5, 475–478. [Google Scholar]
- Kolter, R.; Greenberg, E.P. Microbial sciences: The superficial life of microbes. Nature 2006, 441, 300–302. [Google Scholar] [CrossRef]
- Elhariry, H.M. Biofilm formation by Aeromonas hydrophila on green-leafy vegetables: Cabbage and lettuce. Foodborne Pathog. Dis. 2011, 8, 125–131. [Google Scholar] [CrossRef]
- Odeyemi, O.A.; Ahmad, A. Population dynamics, antibiotics resistance and biofilm formation of Aeromonas and Vibrio species isolated from aquatic sources in Northern Malaysia. Microb. Pathog. 2017, 103, 178–185. [Google Scholar] [CrossRef]
- Jahid, I.K.; Ha, S.D. A review of microbial biofilms of produce: Future challenge to food safety. Food Sci. Biotechnol. 2012, 21, 299–316. [Google Scholar] [CrossRef]
- FAO (Fisheries and Aquaculture Department). Global Aquaculture Production Statistics for the year. 2011. Available online: ftp://ftp.fao.org/FI/news/GlobalAquacultureProductionStatistics2011.pdf (accessed on 2 March 2014).
- De Silva, B.C.J.; Hossain, S.; Dahanayake, P.S.; Heo, G.J. Aeromonas spp. from marketed Yesso scallop (Patinopecten yessoensis): Molecular characterization, phylogenetic analysis, virulence properties and antimicrobial susceptibility. J. Appl. Microbiol. 2019, 126, 288–299. [Google Scholar] [CrossRef]
- CLSI Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2020. [Google Scholar]
- Yano, Y.; Hamano, K.; Tsutsui, I.; Aue-umneoy, D.; Ban, M.; Satomi, M. Occurrence, molecular characterization, and antimicrobial susceptibility of Aeromonas spp. in marine species of shrimps cultured at inland low salinity ponds. Food Microbiol. 2015, 47, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Yousr, A.H.; Napis, S.; Rusul, G.R.A.; Son, R. Detection of aerolysin and hemolysin genes in Aeromonas spp. isolated from environmental and shellfishsources by Polymerase Chain Reaction. ASEAN Food. J. 2007, 14, 115–122. [Google Scholar]
- CLSI. Clinical and Laboratory Standards Institute. Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria; Approved Guideline M45-P. Proposed Guideline. CLSI document M45-P; Clinical and Laboratory Standard Institute: Wayne, PA, USA, 2006; Volume 25. [Google Scholar]
- CLSI. Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria., 3rd ed.; CLSI guideline M45.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- CLSI. Clinical and Laboratory Standards Institute. M100 Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017; p. 296. [Google Scholar]
- Performance Standards for Antimicrobial Susceptibility Testing. CLSI supplement M100, 29th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- CASFM. Comite’ de l’Antibiogramme de la Socie´te´ Franc¸aise de Microbiologie; Proposer les Recommandations; CASFM: Littleton, CO, USA, 2013. [Google Scholar]
- Sarter, S.; Kha Nguyen, H.N.; Hung, L.T.; Lazard, J.; Montet, D. Antibiotic resistance in Gram-negative bacteria isolated from farmed catfish. Food Control 2007, 18, 1391–1396. [Google Scholar] [CrossRef]
- Krumperman, P.H. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizan, M.F.R.; Jahid, I.K.; Ha, S.D. Microbial biofilms in seafood: A food-hygiene challenge. Food Microbiol. 2015, 49, 41–55. [Google Scholar] [CrossRef]
- Elexson, N.; Afsah-Hejri, L.; Rukayadi, Y.; Soopna, P.; Lee, H.Y.; Zainazor, T.T.; Son, R. Effect of detergents as antibacterial agents on biofilm of antibiotics-resistant Vibrio parahaemolyticus isolates. Food Control 2014, 35, 378–385. [Google Scholar] [CrossRef]
- Skåre, J.U.; Brantsæter, A.L.; Frøyland, L.; Hemre, G.-I.; Knutsen, H.K.; Lillegaard, I.T.L.; Torstensen, B. Benefit-Risk Assessment of Fish and Fish Products in the Norwegian Diet—An Update; Norwegian Scientific Committee for Food Safety (VKM): Oslo, Norway, 2014. (In Norwegian) [Google Scholar]
- Abbas, E.M.; Soliman, T.; El-Magd, M.; Farrag, M.; Ismail, R.F.; Kato, M. Phylogeny and DNA barcoding of the family Sparidae inferred from mitochondrial DNA of the Egyptian waters. J. Fish Aquat. Sci. 2017, 12, 73–81. [Google Scholar] [CrossRef] [Green Version]
- El Deen, A.N.; Dorgham-Sohad, M.; Hassan-Azza, H.; Hakim, A. Studies on Aeromonas hydrophila in cultured Oreochromis niloticus at Kafr El Sheikh Governorate, Egypt with reference to histopathological alterations in some vital organs. World J. Fish Mar. Sci. 2014, 6, 233–240. [Google Scholar] [CrossRef]
- Hassan, M.A.; Noureldin, E.; Mahmoud, M.A.; Fita, N.A. Molecular identification and epizootiology of Aeromonas veronii infection among farmed Oreochromis niloticus in Eastern Province, KSA. J. Aquat. Res. 2017, 43, 161–167. [Google Scholar] [CrossRef]
- Ahmed, H.A.; Mohamed, M.E.; Rezk, M.M.; Gharieb, R.; Abdel-Maksoud, S.A. Aeromonas hydrophila in fish and humans; prevalence, virulotyping and antimicrobial resistance. Slov. Vet. Res. 2018, 55, 113–124. [Google Scholar] [CrossRef]
- Lopatek, M.; Wieczorek, K.; Osek, J. Antimicrobial resistance, virulence factors, and genetic profiles of Vibrio parahaemolyticus from seafood. Appl. Environ. Microbiol. 2018, 84, e00537–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahimi, E.; Raissy, M.; Razzaghimanesh, M.; Dastgerdi, A.A.; Shahraki, M.M. Occurrence of Aeromonas hydrophila in fish, shrimp, lobster and crab in Iran. Kafkas Univ. Vet. Fak. Derg. 2014, 20, 691–696. [Google Scholar] [CrossRef]
- Castro-Escarpulli, G.; Figueras, M.J.; Aguilera-Arreola, G.; Soler, L.; Fernández-Rendón, E.; Aparicio, G.O.; Guarro, J.; Chacón, M.R. Characterisation of Aeromonas spp. isolated from frozen fish intended for human consumption in Mexico. Int. J. Food Microbiol. 2003, 84, 41–49. [Google Scholar] [CrossRef] [PubMed]
- El-ghareeb, H.M.; Zahran, E.; Abd-Elghany, S.M. Occurrence, molecular characterization and antimicrobial resistance of pathogenic Aeromonas Hydrophila from retail fish. Alex. J. For Vet. Sci. 2019, 62, 172–181. [Google Scholar]
- Ashiru, A.W.; Uaboi-Egbeni, P.O.; Oguntowo, J.E.; Idika, C.N. Isolation and antibiotic profile of Aeromonas spp. from tilapia and catfish fish. Pak. J. Nutr. 2011, 10, 982–986. [Google Scholar] [CrossRef] [Green Version]
- Borchardt, M.A.; Stemper, M.E.; Standridge, J.H. Aeromonas isolates from human diarrheic stool and ground water compared by pulsed-field gel electrophoresis. Emerg. Infect Dis. 2003, 9, 224–228. [Google Scholar] [CrossRef] [Green Version]
- Tahoun, A.B.; Ahmed, H.A.; Abou Elez, R.M.; El-Gedawy, A.A.; Elsohaby, I.; Abd El-Ghafar, A.E. Molecular characterisation, genotyping and survival of Aeromonas hydrophila isolated from milk, dairy products and humans in Egypt. Int. Dairy J. 2016, 63, 52–58. [Google Scholar] [CrossRef]
- Tayler, A.E.; Ayala, J.A.; Niumsup, P.; Westphal, K.; Baker, J.A.; Zhang, J.A.; Zhang, L.; Walsh, T.R.; Wiedemann, B.; Bennett, P.A.; et al. Induction of β-lactamase production in Aeromonas hydrophila is responsive to β-lactam-mediated changes in peptidoglycan composition. J. Microbiol. 2010, 156, 2327–2335. [Google Scholar] [CrossRef] [Green Version]
- Hafez, A.E.E.; Darwish, W.S.; Elbayomi, R.M.; Hussein, M.A.; El Nahal, S.M. Prevalence, antibiogram and molecular characterization of Aeromonas hydrophila isolated from frozen fish marketed in Egypt. Slov. Vet. Res. 2018, 55, 445–454. [Google Scholar]
- Dhanapala, P.M.; Kalupahana, R.S.; Kalupahana, A.W.; Wijesekera, D.P.H.; Kottawatta, S.A.; Jayasekera, N.K.; Jagoda, S.D.S. Characterization and antimicrobial resistance of environmental and clinical Aeromonas species isolated from fresh water ornamental fish and associated farming environment in Sri Lanka. J. Micro. 2021, 9, 2106. [Google Scholar] [CrossRef]
- Sarder, H.; Khan, T.; Saha, M.L.; Punom, N.J.; Mandal, S.C.; Rahman, M.S. Prevalence and antibiotic susceptibility of Aeromonas hydrophila isolated from freshwater fishes. J. Fish 2016, 4, 411–419. [Google Scholar] [CrossRef]
- McDermott, P.F.; Zhao, S.; Wagner, D.D.; Simjee, S.; Walker, R.D.; White, D.G. The food safety perspective of antibiotic resistance. Anim. Biotechnol. 2002, 13, 71–84. [Google Scholar] [CrossRef]
- Baron, S.; Granier, S.A.; Larvor, E.; Jouy, E.; Cineux, M.; Wilhelm, A.; Gassilloud, B.; Le Bouquin, S.; Kempf, I.; Chauvin, C. Aeromonas diversity and antimicrobial susceptibility in freshwater—An attempt to set generic epidemiological cut-off values . Front. Microbiol. 2017, 8, 503. [Google Scholar] [CrossRef] [Green Version]
- Usui, M.; Tagaki, C.; Fukuda, A.; Okubo, T.; Boonla, C.; Suzuki, S.; Seki, K.; Takada, H.; Tamura, Y. Use of Aeromonas spp. as general indicators of antimicrobial susceptibility among bacteria in aquatic environments in Thailand. Front. Microbiol. 2016, 7, 710. [Google Scholar] [CrossRef] [Green Version]
- Laith, A.; Najiah, M. Aeromonas hydrophila: Antimicrobial susceptibility and histopathology of isolates from diseased catfish, Clarias gariepinus (Burchell). J. Aquac. Res. Dev. 2014, 5, 215. [Google Scholar] [CrossRef] [Green Version]
- Matter, A.F.; El Asely, A.M.; Shaheen, A.A.; El-Gawad, E.A.A.; El-Abd, H.; Abbass, A.A. Phenotypic and molecular characterization of bacterial pathogens isolated from diseased freshwater fishes. Fish Aquat. Stud. Int. J. 2018, 6, 34–41. [Google Scholar]
- Jutfelt, F.; Sundh, H.; Glette, J.; Mellander, L.; Thrandur Björnsson, B.; Sundell, K. The involvement of Aeromonas salmonicida virulence factors in bacterial translocation across the rainbow trout, Oncorhynchus mykiss (Walbaum), intestine. J. Fish Dis. 2008, 31, 141–151. [Google Scholar] [CrossRef]
- Dar, G.H.; Dar, S.A.; Kamili, A.N.; Chishti, M.Z.; Ahmad, F. Detection and characterization of potentially pathogenic Aeromonas sobria isolated from fish Hypophthalmichthys molitrix (Cypriniformes: Cyprinidae). Microb. Pathog. 2016, 91, 136–140. [Google Scholar] [CrossRef]
- Yogananth, N.; Bhakyaraj, R.; Chanthuru, A.; Anbalagan, T.; Nila, K.M. Detection of Virulence Gene in Aeromonas hydrophila isolated from fish samples Using PCR Technique. Glob. J. Biotech. Biochem. 2009, 4, 51–53. [Google Scholar]
- Ramadan, H.; Ibrahim, N.; Samir, M.; Abd El-Moaty, A.; Gad, T. Aeromonas hydrophila from marketed mullet (Mugil cephalus) in Egypt: PCR characterization of β-lactam resistance and virulence genes. J. Appl. Microbiol. 2018, 124, 1629–1637. [Google Scholar] [CrossRef]
- Younes, A.M.; Gaafar, A.Y.; Awad, E.S. Virulence Determinants and Plasmid Profile of Aeromonas hydrophila Strains Isolated from Oreo-chromis niloticus. Glob. Vet. 2015, 15, 613–617. [Google Scholar]
- Simon, S.S.; Lalitha, K.V.; Joseph, T.C. Virulence properties of Aeromonas spp. from modified-atmosphere- and vacuum-packed milk fish (Chanos chanos Forsskal, 1775). Ann. Microbiol. 2016, 66, 1109–1115. [Google Scholar] [CrossRef]
- Abd-Elall, A.; Abd-El-Kader, M.; Atia, A. Occurrence, seasonal variations and virulence of Aeromonas hydrophila and Aeromonas caviae in fish farms at East Delta, Egypt. Glob. Vet. 2014, 13, 328–336. [Google Scholar]
- Hoel, S.; Vadstein, O.; Jakobsen, A.N. Species distribution and prevalence of putative virulence factors in mesophilic Aeromonas spp. isolated from fresh retail sushi. Front. Microbiol. 2017, 8, 931. [Google Scholar] [CrossRef] [Green Version]
- El-Gohary, F.A.; Zahran, E.; Abd El-Gawad, E.A.; El-Gohary, A.H.M.; Abdelhamid, F.; El-Mleeh, A.; Elmahallawy, E.K.; Elsayed, M.M. Investigation of the Prevalence, Virulence Genes, and Antibiogram of Motile Aeromonads Isolated from Nile Tilapia Fish Farms in Egypt and Assessment of their Water Quality. J. Anim. 2020, 10, 1432. [Google Scholar] [CrossRef]
- Dias, C.; Borges, A.; Saavedra, M.J.; Simões, M. Biofilm formation and multidrug-resistant Aeromonas spp. from wild animals. J. Glob. Antimicrob. Resist. 2018, 12, 227–234. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
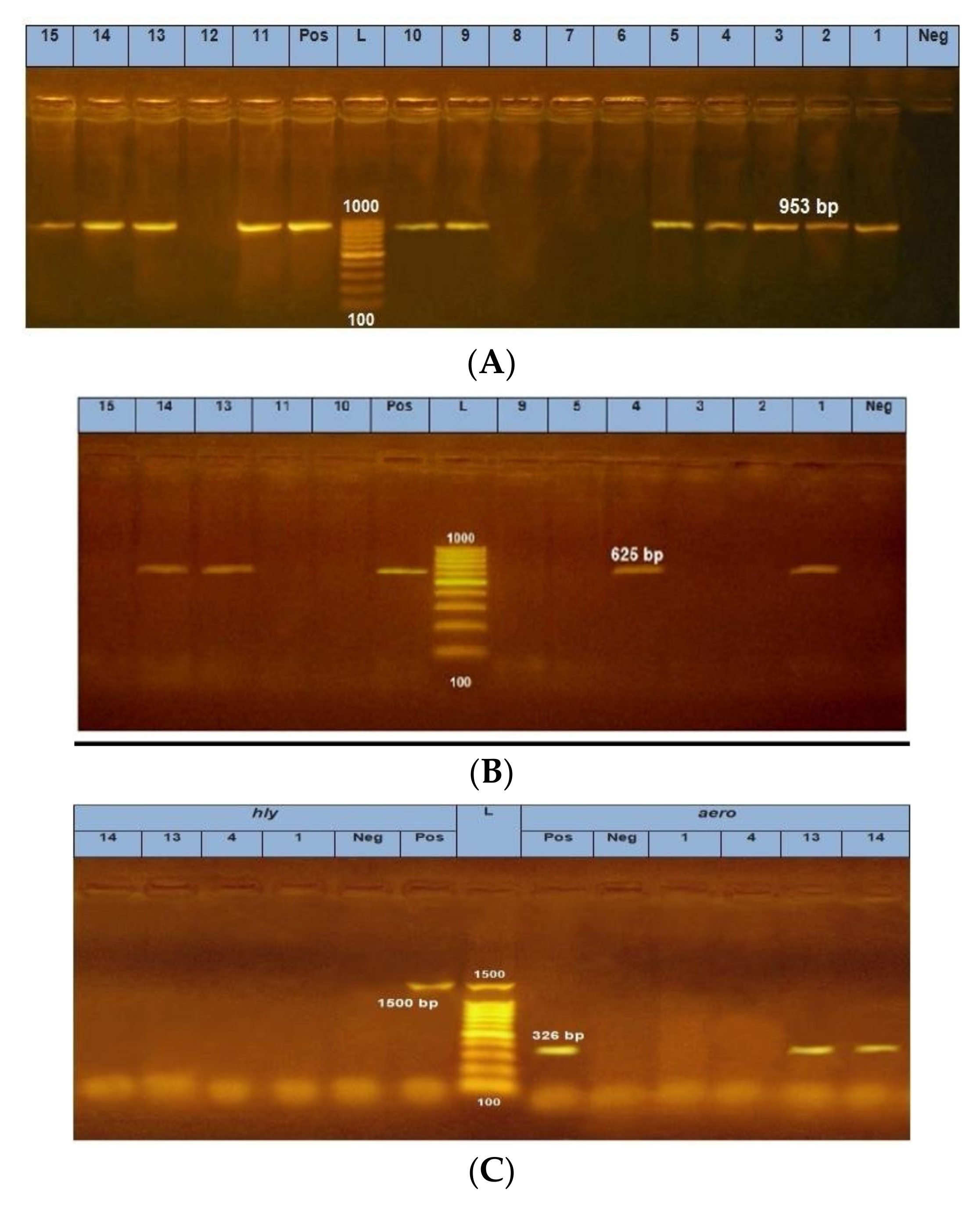
| Gene | Primer | Length (bp) | Reference |
|---|---|---|---|
| 16S rRNA (genus-specific) | 953 | [20] | |
| F; 5′- CTA TGA AAA AAC TAA AAA TAA CTG - | ′3 | ||
| R; 5′- CAG TAT AAG TGG GGA AAT GGA AAG - | ′3 | ||
| 16S rRNA (species-specific) | 625 | [24,25] | |
| F; 5′- CAC AGC CAA TAT GTC GGT GAA G -′3 | |||
| R; 5′- GTC ACC TTC TCG CTC AGG C--′3 |
| Types of Samples | No. of Examined Samples | No. of Infected Samples (Positive Samples) | Percentage of Positivesamples | |
|---|---|---|---|---|
| Tilapia: (n = 160) | Viscera | 57 | 6 | 10.5% |
| Liver | 57 | 8 | 14.0% | |
| Muscles | 46 | 2 | 4.3% | |
| Mugil: (n = 105) | Viscera | 38 | 8 | 21.0% |
| Liver | 25 | 1 | 4.0% | |
| Muscles | 42 | 5 | 11.9% | |
| Hand swabs (n = 51) | 51 | 12 | 23.5% | |
| Patient Stool (n = 27) | 27 | 0 | 0 | |
| Total | 343 | 42 | 12.2% | |
| Code of Isolates | Source of Isolates | Molecularly Identified Aeromonas spp. | Molecularly Identified A. hydrophila | Hly Gene | aer Gene |
|---|---|---|---|---|---|
| Aer 1 | Tilapia V * | + | + | - | - |
| Aer 2 | Tilapia V | + | - | - | - |
| Aer 3 | Mugil V * | + | - | - | - |
| Aer 4 | Mugil V | + | + | - | - |
| Aer 5 | Mugil M * | + | - | - | - |
| Aer 6 | Mugil M | - | - | - | - |
| Aer 7 | Mugil M | - | - | - | - |
| Aer 8 | Mugil L * | - | - | - | - |
| Aer 9 | Mugil V | + | - | - | - |
| Aer 10 | Hand swabs | + | - | - | - |
| Aer 11 | Hand swabs | + | - | - | - |
| Aer 12 | Hand swabs | - | - | - | |
| Aer 13 | Tilapia V | + | + | - | + |
| Aer 14 | Mugil V | + | + | - | + |
| Aer 15 | Tilapia M * | + | - | - | - |
| Antibiotics | Aeromonas spp. Isolates (no = 11) RI * S * | ||
|---|---|---|---|
| Ceftriaxone (CRO 30) | 4 (36.4%) | 5 (45.4%) | 2 (18.2%) |
| Cefotaxime (CTX 30) | 2 (18.2%) | 7 (63.6%) | 2 (18.2%) |
| Ceftazidime (CAZ 30) | 3 (27.3%) | 6 (54.5) | 2 (18.2%) |
| Cefixime (CFM 5) | 9 (81.8%) | - | 2 (18.2%) |
| Imipenem (IPM 10) | - | - | 11 (100%) |
| Gentamycin (CN 10) | - | 2 (18.2%) | 9 (81.8%) |
| Tobramycin (TOB 10) | 5 (45.4%) | 4 (36.4%) | 2 (18.2%) |
| Kanamycin (K 30) | 3 (27.3%) | 5 (45.4%) | 3 (27.3%) |
| Streptomycin (S 10) | 1 (9.1%) | 1 (9.1%) | 9 (81.8%) |
| Tetracycline (TE 30) | 10 (90.9%) | 1 (9.1%) | - |
| Ciprofloxacin (CIP 5) | 9 (81.8%) | 2 (18.2%) | - |
| Norfloxacin (NOR) | 2 (18.2%) | 5 (45.4%) | 4 (36.4%) |
| Nalidixic acid (NA 30) | 5 (45.4%) | 3 (27.3%) | 3 (27.3%) |
| Chloramphenicol (C) | - | 1 (9.1%) | 10 (90.9%) |
| Bacterial Code | Isolates Source | No. of Resistant Antibiotics | Resistance Profile | Number of Antibiotic Classes | MAR Resi Index | Stance Level |
|---|---|---|---|---|---|---|
| Aer1 | Tilapia Viscera | 4 | CFM, TE, CIP, NOR. | 2 | 0.285 | DR |
| Aer2 | Tilapia Viscera | 3 | CFM, TE, CIP. | 2 | 0.214 | DR |
| Aer3 | Mugil Viscera | 2 | CFM, TE. | 2 | 0.142 | DR |
| Aer4 | Mugil Viscera | 4 | CFM, TE, CRO, CAZ | 2 | 0.285 | DR |
| Aer5 | Mugil Muscles | 3 | TE, CIP, TOB | 2 | 0.214 | DR |
| Aer9 | Mugil Viscera | 5 | CFM, TE, CIP, CAZ, TOB | 3 | 0.357 | MDR |
| Aer10 | Hand swabs | 5 | TE, CIP, NOR, NA, TOB | 2 | 0.357 | DR |
| Aer11 | Hand swabs | 8 | CFM, TE, CIP, NA, CRO, CAZ, TOB, K | 3 | 0.571 | MDR |
| Aer13 | Tilapia Viscera | 6 | CFM, CIP, CRO, CTX, K, NA | 3 | 0.428 | MDR |
| Aer14 | Mugil Viscera | 9 | CFM, TE, CIP, CRO, CTX, TOB, K, S, NA | 3 | 0.642 | MDR |
| Aer15 | Tilapia Muscles | 4 | CFM, TE, CIP, NA | 2 | 0.285 | DR |
| Bacterial Code | Source of Isolates | Mean OD ± SD | Mean * ± SD | Degree of Biofilm |
|---|---|---|---|---|
| Control | 0.103 ± 0.01 c | |||
| Aer 1 | Tilapia Viscera | 0.124. ± 0.02 c | 0.021 ± 0.021 | Non |
| Aer 2 | Tilapia Viscera | 0.126 ± 0.007 c | 0.023 ± 0.007 | Non |
| Aer 3 | Mugil Viscera | 0.108 ± 0.005 c | 0.005 ± 0.005 | Non |
| Aer 4 | Mugil Viscera | 0.119 ± 0.012 c | 0.016 ± 0.012 | Non |
| Aer 5 | Mugil Muscle | 0.117 ± 0.013 c | 0.014 ± 0.013 | Non |
| Aer 9 | Mugil Viscera | 0.106 ± 0.002 c | 0.003 ± 0.002 | Non |
| Aer 10 | Hand Swabs | 0.149 ± 0.004 c | 0.046 ± 0.004 | Non |
| Aer 11 | Hand Swabs | 0.155 ± 0.009 c | 0.052 ± 0.009 | Non |
| Aer 13 | Tilapia Viscera | 0.135 ± 0.029 c | 0.032 ± 0.029 | Non |
| Aer 14 | Mugil Viscera | 0.27 ± 0.009 b | 0.167 ± 0.009 | Weak |
| Aer 15 | Tilapia muscles | 0.701 ± 0.089 a | 0.598 ± 0.089 | Strong |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Hossary, D.; Mahdy, A.; Elariny, E.Y.T.; Askora, A.; Merwad, A.M.A.; Saber, T.; Dahshan, H.; Hakami, N.Y.; Ibrahim, R.A. Antibiotic Resistance, Virulence Gene Detection, and Biofilm Formation in Aeromonas spp. Isolated from Fish and Humans in Egypt. Biology 2023, 12, 421. https://doi.org/10.3390/biology12030421
El-Hossary D, Mahdy A, Elariny EYT, Askora A, Merwad AMA, Saber T, Dahshan H, Hakami NY, Ibrahim RA. Antibiotic Resistance, Virulence Gene Detection, and Biofilm Formation in Aeromonas spp. Isolated from Fish and Humans in Egypt. Biology. 2023; 12(3):421. https://doi.org/10.3390/biology12030421
Chicago/Turabian StyleEl-Hossary, Dalia, Asmaa Mahdy, Eman Y. T. Elariny, Ahmed Askora, Abdallah M. A. Merwad, Taisir Saber, Hesham Dahshan, Nora Y. Hakami, and Rehab A. Ibrahim. 2023. "Antibiotic Resistance, Virulence Gene Detection, and Biofilm Formation in Aeromonas spp. Isolated from Fish and Humans in Egypt" Biology 12, no. 3: 421. https://doi.org/10.3390/biology12030421