
Apomictic Mountain Whitebeam (Sorbus austriaca, Rosaceae) Comprises Several Genetically and Morphologically Divergent Lineages
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Amplified Fragment Length Polymorphism (AFLP)
2.3. Analysis of Nuclear Microsatellites
2.4. Plastid trnT–trnF Sequencing and Phylogenetic Analyses
2.5. Genome Size Estimation
2.6. Morphometric Analyses
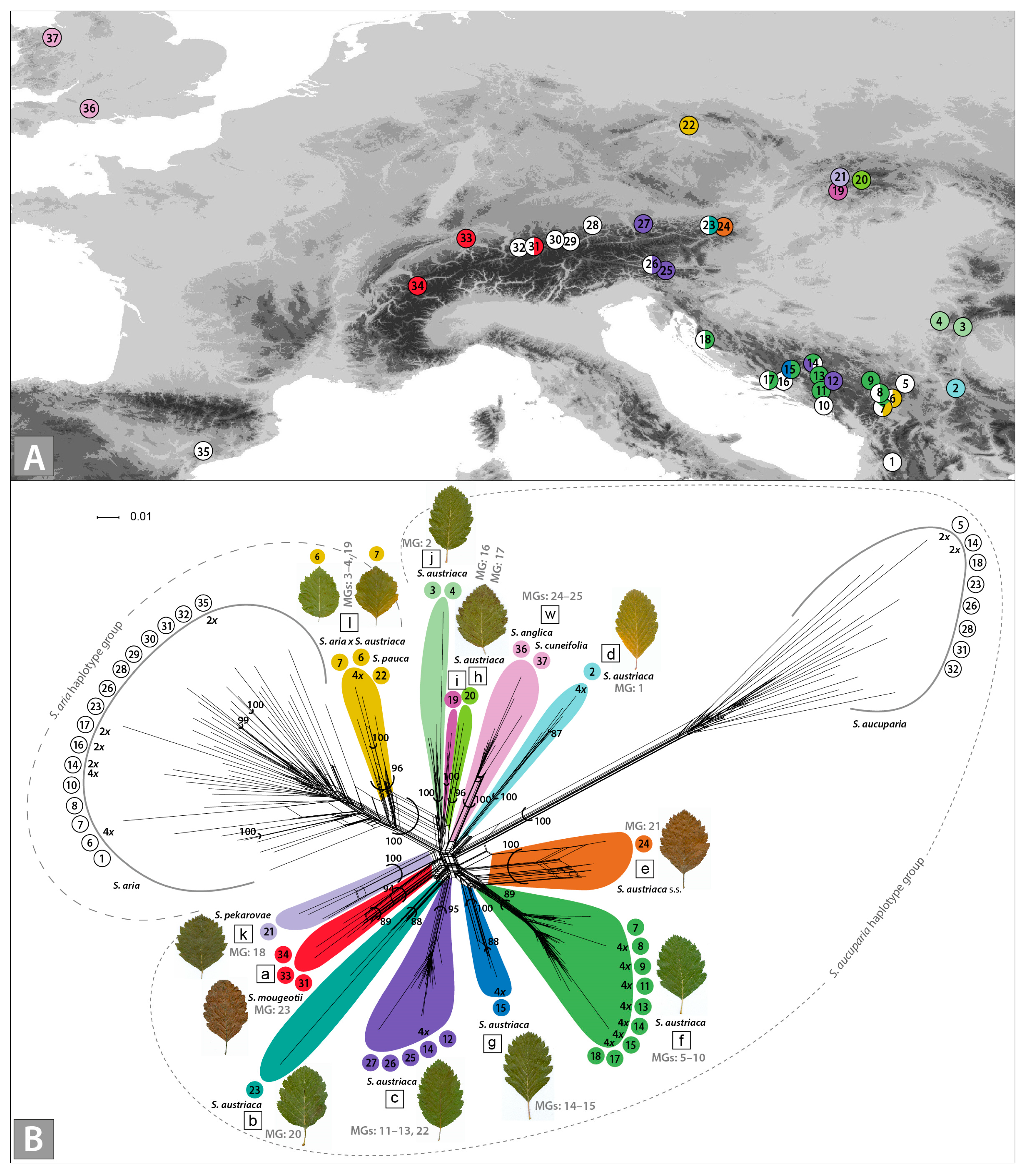
3. Results
3.1. AFLP Fingerprinting
3.2. Nuclear Microsatellites
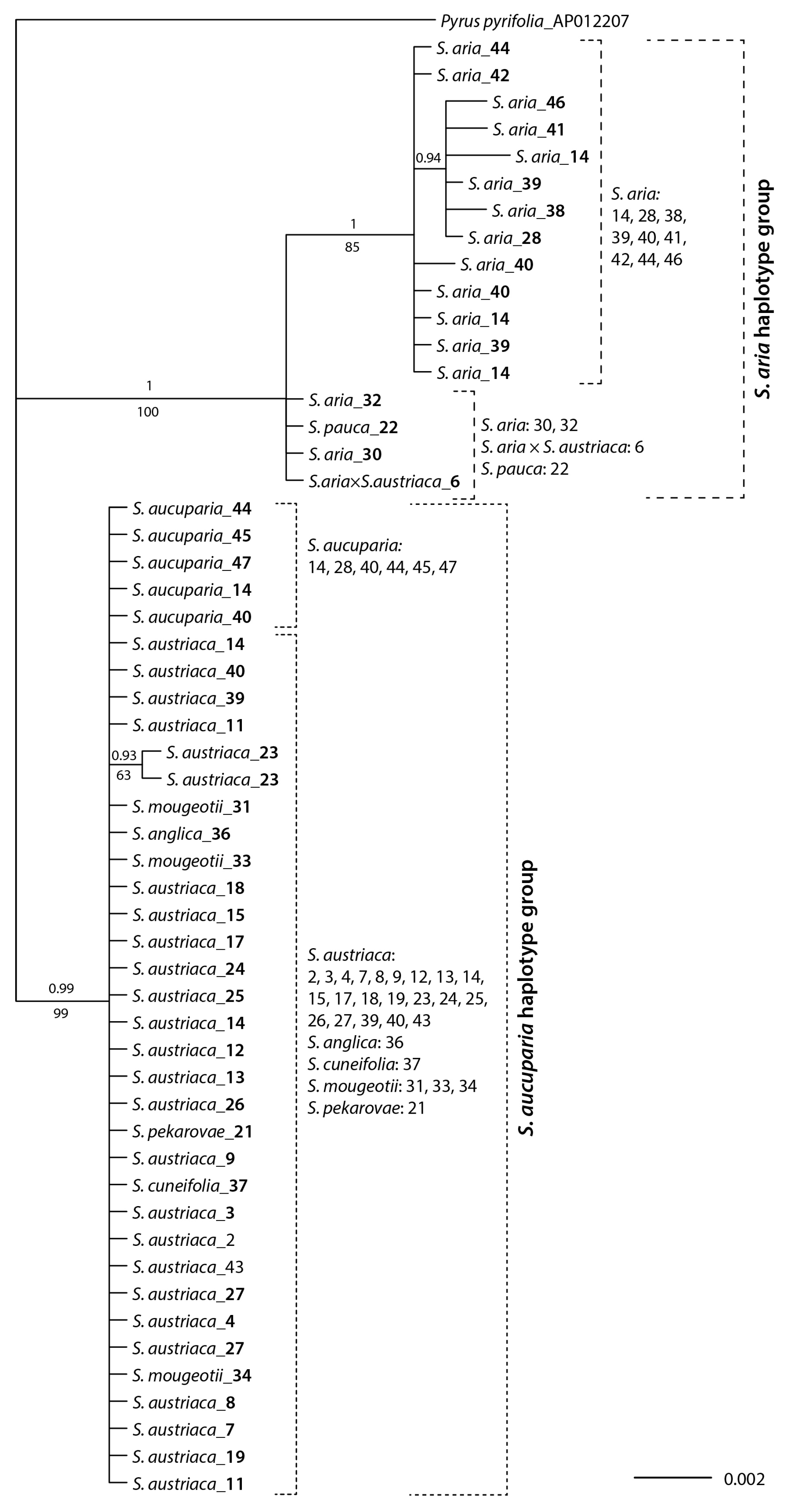
3.3. Plastid trnT–trnF Phylogenetic Relationships
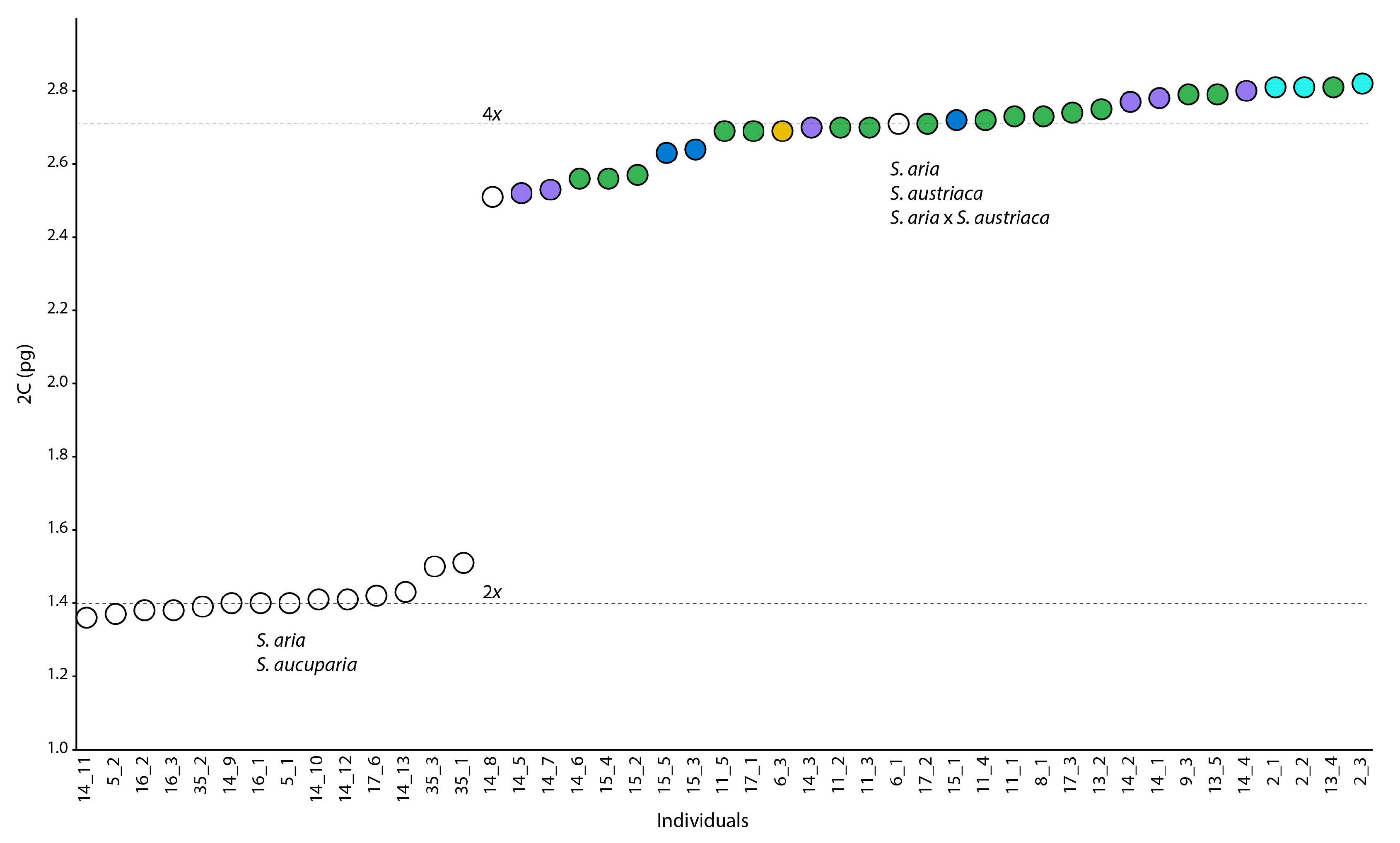
3.4. Genome Size, Ploidy Level and Reproduction Mode
3.5. Morphology
4. Discussion
4.1. Multiple Origins of S. austriaca Lineages
4.2. Mostly S. aucuparia, but also S. aria, Served as Maternal Parents of Hybridogenous Lineages
4.3. Taxonomic Considerations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soltis, D.E.; Albert, V.A.; Leebens-Mack, J.; Bell, C.D.; Paterson, A.H. Polyploidy and angiosperm diversification. Am. J. Bot. 2009, 96, 336–348. [Google Scholar] [CrossRef] [Green Version]
- Wood, T.E.; Takebayashi, N.; Barker, M.S.; Mayrose, I.; Greenspoon, P.B.; Rieseberg, L.H. The frequency of polyploid speciation in vascular plants. Proc. Natl. Acad. Sci. USA 2009, 106, 13875–13879. [Google Scholar] [CrossRef] [Green Version]
- Madlung, A. Polyploidy and its effect on evolutionary success: Old questions revisited with new tools. Heredity 2013, 110, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Peer, Y.; Ashman, T.-L.; Soltis, P.S.; Soltis, D.E. Polyploidy: An evolutionary and ecological force in stressful times. Plant Cell 2021, 33, 11–26. [Google Scholar] [CrossRef]
- Hegarty, M.J.; Hiscock, S.J. Genomic clues to the evolutionary success of polyploid plants. Curr. Biol. 2008, 18, 435–444. [Google Scholar] [CrossRef] [Green Version]
- Weiss-Schneeweiss, H.; Emadzade, K.; Jang, T.-S.; Schneeweiss, G.M. Evolutionary consequences, constraints and potential of polyploidy in plants. Cytogenet. Genome Res. 2013, 140, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Hörandl, E. The evolution of self-fertility in apomictic plants. Sex. Plant Reprod. 2010, 23, 73–86. [Google Scholar] [CrossRef] [Green Version]
- Hojsgaard, D.; Klatt, S.; Baier, R.; Carman, J.G.; Hörandl, E. Taxonomy and biogeography of apomixis in angiosperms and associated biodiversity characteristics. Crit. Rev. Plant Sci. 2014, 33, 414–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coughlan, J.M.; Han, S.; Stefanović, S.; Dickinson, T.A. Widespread generalist clones are associated with range and niche expansion in allopolyploids of Pacific Northwest Hawthorns (Crataegus L.). Mol. Ecol. 2017, 26, 5484–5499. [Google Scholar] [CrossRef]
- Ozias-Akins, P.; van Dijk, P.J. Mendelian genetics of apomixis in plants. Ann. Rev. Genet. 2007, 41, 509–537. [Google Scholar] [CrossRef]
- Cosendai, A.C.; Wagner, J.; Ladinig, U.; Rosche, C.; Hörandl, E. Geographical parthenogenesis and population genetic structure in the alpine species Ranunculus kuepferi (Ranunculaceae). Heredity 2013, 110, 560–569. [Google Scholar] [CrossRef] [Green Version]
- Majeský, L.; Vašut, R.J.; Kitner, M.; Trávníček, B. The pattern of genetic variability in apomictic clones of Taraxacum officinale indicates the alternation of asexual and sexual histories of apomicts. PLoS ONE 2013, 7, e41868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paun, O.; Stuessy, T.F.; Hörandl, E. The role of hybridization, polyploidization and glaciation in the origin and evolution of the apomictic Ranunculus cassubicus complex. New Phytol. 2006, 171, 223–236. [Google Scholar] [CrossRef]
- Hörandl, E. The complex causality of geographical parthenogenesis. New Phytol. 2006, 171, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Hojsgaard, D.; Hörandl, E. Apomixis as a facilitator of range expansion and diversification in plants. In Evolutionary Biology: Biodiversification from Genotype to Phenotype, 1st ed.; Pontarotti, P., Ed.; Springer: Cham, Switzerland, 2015; pp. 305–327. [Google Scholar] [CrossRef]
- Dickinson, T.A.; Yan, B.X.; Han, S.; Zarrei, M. Niche shifts, hybridization, polyploidy and geographic parthenogenesis in western North American hawthorns (Crataegus subg. Sanguineae, Rosaceae). Agronomy 2021, 11, 2133. [Google Scholar] [CrossRef]
- Kurtto, A.; Webber, H.E.; Lampinen, R.; Sennikov, A.N. (Eds.) Atlas Florae Europaeae. Distribution of Vascular Plants in Europe. 15. Rosaceae (Rubus), 1st ed.; The Committee for Mapping the Flora of Europe & Societas Biologica Fennica Vanamo: Helsinki, Finland, 2010. [Google Scholar]
- Rich, T.C.G.; Houston, L.; Robertson, A.; Proctor, M.C.F. Whitebeams, Rowans and Service Trees of Britain and Ireland. A monograph of British and Irish Sorbus L., 1st ed.; B.S.B.I. Handbook No. 14; Botanical Society of the British Isles: London, UK, 2010. [Google Scholar]
- Kurtto, A.; Sennikov, A.N.; Lampinen, R. (Eds.) Atlas Florae Europaeae. Distribution of Vascular Plants in Europe. 17. Rosaceae (Sorbus s. lato), 1st ed.; The Committee for Mapping the Flora of Europe & Societas Biologica Fennica Vanamo: Helsinki, Finland, 2018. [Google Scholar]
- Vašut, R.J. Taraxacum sect. Erythrosperma in Moravia (Czech Republic): Taxonomic notes and the distribution of previously described species. Preslia 2003, 75, 311–338. [Google Scholar]
- Robertson, A.; Sydes, C. Sorbus pseudomeinichii, a new endemic Sorbus (Rosaceae) microspecies from Arran, Scotland. Watsonia 2006, 26, 85591923. [Google Scholar]
- Rich, T.C.G.; Charles, C.; Houston, L.; Tillotson, A. The diversity of Sorbus L. (Rosaceae) in the Lower Wye Valley. Watsonia 2009, 27, 301–313. [Google Scholar]
- Lepší, M.; Vit, P.; Lepší, P.; Boublik, K.; Suda, J. Sorbus milensis, a new hybridogenous species from northwestern Bohemia. Preslia 2008, 80, 229–244. [Google Scholar]
- Lepší, M.; Vit, P.; Lepší, P.; Boublik, K.; Kolar, F. Sorbus portae-bohemicae and Sorbus albensis, two new apomictic endemic species recognized based on revision of Sorbus bohemica. Preslia 2009, 81, 63–89. [Google Scholar]
- Lepší, M.; Lepší, P.; Sádlo, J.; Koutecký, P.; Vít, P.; Petřík, P. Sorbus pauca species nova, the first endemic species of the Sorbus hybrida group for the Czech Republic. Preslia 2013, 85, 63–80. [Google Scholar]
- Lepší, M.; Lepší, P.; Koutecký, P.; Bílá, J.; Vít, P. Taxonomic revision of Sorbus subgenus Aria occurring in the Czech Republic. Preslia 2015, 87, 109–162. [Google Scholar]
- Aldasoro, J.J.; Aedo, C.; Navarro, C.; Garmendia, F.M. The genus Sorbus (Maloideae, Rosaceae) in Europe and in North Africa: Morphological analysis and systematics. Syst. Bot. 1998, 23, 189–212. [Google Scholar] [CrossRef]
- Karbstein, K.; Tomasello, S.; Hodač, L.; Dunkel, F.G.; Daubert, M.; Hörandl, E. Phylogenomics supported by geometric morphometrics reveals delimitation of sexual species within the polyploid apomictic Ranunculus auricomus complex (Ranunculaceae). Taxon 2020, 69, 1191–1220. [Google Scholar] [CrossRef]
- Zarrei, M.; Stefanović, S.; Dickinson, T.A. Reticulate evolution in North American black-fruited hawthorns (Crataegus section Douglasia; Rosaceae): Evidence from nuclear ITS2 and plastid sequences. Ann. Bot. 2014, 114, 253–269. [Google Scholar] [CrossRef] [Green Version]
- Paule, J.; Scherbantin, A.; Dobeš, C. Implications of hybridisation and cytotypic differentiation in speciation assessed by AFLP and plastid haplotypes—A case study of Potentilla alpicola La Soie. BMC Evol. Biol. 2012, 12, 132. [Google Scholar] [CrossRef] [Green Version]
- Šarhanová, P.; Sharbel, T.F.; Sochor, M.; Vašut, R.; Dančák, M.; Trávniček, B. Hybridization drives evolution of apomicts in Rubus subgenus Rubus: Evidence from microsatellite markers. Ann. Bot. 2017, 120, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Nelson-Jones, E.B.; Briggs, D.; Smith, A.G. The origin of intermediate species of the genus Sorbus. Theor. Appl. Genet. 2002, 105, 953–963. [Google Scholar] [CrossRef]
- Robertson, A.; Newton, A.C.; Ennos, R.A. Multiple hybrid origins, genetic diversity and population genetic structure of two endemic Sorbus taxa on the Isle of Arran, Scotland. Mol. Ecol. 2004, 13, 123–134. [Google Scholar] [CrossRef]
- Robertson, A.; Rich, T.C.G.; Allen, A.M.; Houston, L.; Roberts, C.; Bridle, J.R.; Harris, S.A.; Hiscock, S.J. Hybridization and polyploidy as drivers of continuing evolution and speciation in Sorbus. Mol. Ecol. 2010, 19, 1675–1690. [Google Scholar] [CrossRef]
- Ludwig, S.; Robertson, A.; Rich, T.C.G.; Djordjevic, M.; Cerovic, R.; Houston, L.; Harris, S.A.; Hiscock, S.J. Breeding systems, hybridisation and continuing evolution in Avon Gorge Sorbus. Ann. Bot. 2013, 111, 563–575. [Google Scholar] [CrossRef] [Green Version]
- Hamston, T.J.; de Vere, N.; King, A.R.; Pellicer, J.; Fay, M.F.; Creswell, J.E.; Stevens, J.R. Apomixis and hybridization drives reticulate evolution and phyletic differentiation in Sorbus L.: Implications for conservation. Front. Plant Sci. 2018, 9, 1796. [Google Scholar] [CrossRef] [PubMed]
- Lepší, M.; Koutecký, P.; Nosková, J.; Lepší, P.; Urfus, T.; Rich, T.C.G. Versatility of reproductive modes and ploidy level interactions in Sorbus s.l. (Malinae, Rosaceae). Bot. J. Linn. Soc. 2019, 191, 502–522. [Google Scholar] [CrossRef]
- Jankun, A. Evolutive significance of apomixis in the genus Sorbus (Rosaceae). Fragm. Florist. Geobot. 1993, 38, 627–686. [Google Scholar]
- Hajrudinović, A.; Frajman, B.; Schönswetter, P.; Silajdžić, E.; Siljak-Yakovlev, S.; Bogunić, F. Towards a better understanding of polyploid Sorbus (Rosaceae) from Bosnia and Herzegovina (Balkan Peninsula), including description of a novel, tetraploid apomictic species. Bot. J. Linn. Soc. 2015, 178, 670–685. [Google Scholar] [CrossRef] [Green Version]
- Meyer, N.; Meierott, L. Ergänzende Beiträge zur Sorbus-Flora von Bayern. Bayer. Bot. Ges. 2021, 91, 21–48. [Google Scholar]
- Hajrudinović, A.; Siljak-Yakovlev, S.; Brown, S.C.; Pustahija, F.; Bourge, M.; Ballian, D.; Bogunić, F. When sexual meets apomict: Genome size, ploidy level and reproductive mode variation of Sorbus aria s.l. and S. austriaca (Rosaceae) in Bosnia and Herzegovina. Ann. Bot. 2015, 116, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Sennikov, A.N.; Kurtto, A. A phylogenetic checklist of Sorbus s.l. (Rosaceae) in Europe. Memo. Soc. Fauna. Flora. Fenn. 2017, 93, 1–78. [Google Scholar]
- Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W. (Eds.) The Global Flora. A Practical Flora to Vascular Plant Species of the World—Special Edition, 1st ed.; GLOVAP Nomenclature Part 1; Plant Gateway Ltd.: Bradford, UK, 2018; Volume 4. [Google Scholar]
- Meyer, N.; Meierott, L.; Schuwerk, H.; Angerer, O. Beiträge zur Gattung Sorbus in Bayern, 1st ed.; Berichte der Bayerischen Botanischen Gesellschaft zur Erforschung der heimischen Flora: Sonderband, Germany, 2005. [Google Scholar]
- Velebil, J.; Lepší, M.; Nosková, J.; Lepší, P. Taxonomic assessment of Sorbus subgenus Aria in the Malé Karpaty Mountains. Preslia 2022, 94, 305–334. [Google Scholar] [CrossRef]
- Warburg, E.F.; Kárpáti, Z.E. Sorbus L. In Flora Europaea 2; Tutin, T.G., Heywood, V.H., Burgess, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1968; pp. 67–71. [Google Scholar]
- Beck, G.M. Flora von Nieder-Österreich; Druck und Verlag von Carl Gerold’s Sohn: Wien, Austria, 1890; Volume 2, p. 714. [Google Scholar]
- Kovanda, M. Sorbus austriaca (Beck) Prain in Italy. Webbia 1998, 53, 27–29. [Google Scholar] [CrossRef]
- Rivers, M.C.; Beech, E. Sorbus austriaca (Beck) Hedl. IUCN Red List. Threat. Species 2017. [Google Scholar] [CrossRef]
- Májovský, J.; Bernátová, D. Sorbus pekarovae sp. nova: A new hybridogeneous species from the Veľká Fatra Mts. Biologia 1996, 51, 23–26. [Google Scholar]
- Rich, T.G.C.; Hjertson, M.; Moberg, R. Typification of some T. Hedlund names in Sorbus L. (Rosaceae). Taxon 2006, 55, 203–206. [Google Scholar] [CrossRef]
- Kurtto, A. Rosaceae (pro parte majore). In Euro+Med Plantbase—The Information Resource for Euro-Mediterranean Plant Diversity, 2009+; Available online: https://europlusmed.org/ (accessed on 9 November 2022).
- Jovanović, B. Rod Sorbus L. In Flora of SR Serbia 4, 1st ed.; Josifović, M., Ed.; Srpska akademija Nauka i Umetnosti, Odeljenje Prirodno-Matematičkih Nauka: Beograd, Srbija, 1972; pp. 150–160. [Google Scholar]
- Fischer, M.A.; Oswald, K.; Adler, W. Exkursionsflora für Österreich, Liechtenstein und Südtirol, 1st ed.; Biologiezentrum der Oberösterreichischen Landesmuseen: Linz, Austria, 2008. [Google Scholar]
- Lauber, K.; Wagner, G.; Gygax, A. Flora Helvetica–Illustrierte Flora der Schweiz, 2nd ed.; Haupt Verlag: Bern, Switzerland, 2018. [Google Scholar]
- Nikolić, T. Flora Croatica 3—Vaskularna Flora Republike Hrvatske, 1st ed.; ALFA d.d.: Zagreb, Croatia, 2020. [Google Scholar]
- Tel-zur, N.; Abbo, S.; Myslabodski, D.; Mizrahi, Y. Modified CTAB procedure for DNA isolation from epiphytic cacti of genera Hylocereus and Selenicereus (Cactaceae). Plant Mol. Biol. Rep. 1999, 17, 249–254. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [Green Version]
- Arrigo, N.; Tuszynski, J.W.; Ehrich, D.; Gerdes, T.; Alvarez, N. Evaluating the impact of scoring parameters on the structure of intra-specific genetic variation using RawGeno, an R package for automating AFLP scoring. BMC Bioinform. 2009, 10, 33. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: http://www.r-project.org/ (accessed on 9 November 2022).
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [Green Version]
- Van de Peer, Y.; De Wachter, R. Construction of evolutionary distance trees with TREECON for Windows: Accounting for variation in nucleotide substitution rate among sites. Comput. Appl. Biosci. 1997, 13, 227–230. [Google Scholar] [CrossRef]
- Huson, D.H. SplitsTree: A program for analyzing and visualizing evolutionary data. Bioinformatics 1998, 14, 68–73. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Meirmans, P.G.; Van Tienderen, P.H. GENOTYPE and GENODIVE: Two programs for the analysis of genetic diversity of asexual organisms. Mol. Ecol. Res. 2004, 4, 792–794. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Simmons, M.P.; Ochoterena, H. Gaps as characters in sequence-based phylogenetic analyses. Syst. Biol. 2000, 49, 369–381. [Google Scholar] [CrossRef] [Green Version]
- Müller, K. SeqState: Primer design and sequence statistics for phylogenetic DNA datasets. Appl. Bioinform. 2005, 4, 65–69. [Google Scholar] [CrossRef]
- Swofford, D. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylander, J.A.A. MrAIC.pl. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Lewis, P.O. A likelihood approach to estimating phylogeny from discrete morphological character data. Syst. Biol. 2001, 50, 913–925. [Google Scholar] [CrossRef] [Green Version]
- Marie, D.; Brown, S.C. A cytometric exercise in plant DNA histograms, with 2C values for 70 species. Biol. Cell 1993, 78, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Suda, J.; Krahulcová, A.; Trávníček, P.; Krahulec, F. Ploidy level versus DNA ploidy level: An appeal for consistent terminology. Taxon 2006, 55, 447–450. [Google Scholar] [CrossRef]
- Siljak-Yakovlev, S.; Pustahija, F.; Šolić, M.E.; Bogunić, F.; Muratović, E.; Bašić, N.; Catrice, O.; Brown, S.C. Towards a genome size and chromosome number database of Balkan flora: C-values in 343 taxa with novel values for 242. Adv. Sci. Lett. 2010, 3, 190–213. [Google Scholar] [CrossRef]
- Aldasoro, J.J.; Aedo, C.; Garmendia, F.M.; de la Hoz, F.P.; Navarro, C. Revision of Sorbus Subgenera Aria and Torminaria (Rosaceae-Maloideae). Syst. Bot. Monogr. 2004, 69, 1–148. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.; Stafford, S. Multivariate Statistics for Wildlife and Ecology Research, 1st ed.; Springer Science+Business Media Inc.: New York, NY, USA, 2000. [Google Scholar] [CrossRef]
- Valcárcel, V.; Vargas, P. Quantitative morphology and species delimitation under the general lineage concept: Optimization for Hedera (Araliaceae). Am. J. Bot. 2010, 97, 1555–1573. [Google Scholar] [CrossRef] [Green Version]
- Pellicer, J.; Clermont, S.; Houston, L.; Rich, T.C.G.; Fay, M.F. Cytotype diversity in the Sorbus complex (Rosaceae) in Britain: Sorting out the puzzle. Ann. Bot. 2012, 110, 1185–1193. [Google Scholar] [CrossRef] [Green Version]
- Uhrinová, V.; Zozomova-Lihova, J.; Bernatova, D.; Paule, J.; Paule, L.; Gömöry, D. Origin and genetic differentiation of pink-flowered Sorbus hybrids in the Western Carpathians. Ann. Bot. 2017, 120, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Feulner, M.; Weig, A.; Paule, J.; Gregor, T.; Schott, L.F.; Aas, G. Genetic variability and morphology of tri- and tetraploid members of the Sorbus aria complex in northern Bavaria. Preslia 2017, 89, 275–290. [Google Scholar] [CrossRef] [Green Version]
- van Husen, D. Die Ostalpen in den Eiszeiten, 1st ed.; Geologische Bundesanstalt: Wien, Austria, 1987. [Google Scholar]
- Stepišnik, U.; Grlj, A.; Radoš, D.; Žebre, M. Geomorphology of Blidinje, Dinaric Alps (Bosnia and Herzegovina). J. Maps 2016, 12, 163–171. [Google Scholar] [CrossRef]
- Turril, W.B. The Plant Life of the Balkan Peninsula. Phytogeographical Study, 1st ed.; Oxford: Clarendon, UK, 1929. [Google Scholar]
- Horvat, I.; Glavač, V.; Ellenberg, H. Vegetation Südosteuropas, 1st ed.; Gustav Fischer Verlag: Stuttgart, Germany, 1974. [Google Scholar]
- Feulner, M.; Weig, A.; Voss, T.; Schott, L.F.; Aas, G. Central European polyploids of Sorbus subgenus Aria (Rosaceae) recurrently evolved from diploids of central and south-eastern Europe: Evidence from microsatellite data. Bot. J. Linn. Soc. 2019, 191, 315–324. [Google Scholar] [CrossRef]
- Oddou-Muratorio, S.; Guesnet, D.; Ozdemir, E.; Petit, R.J.; Demesure, B. Patterns of seed dispersal in a scattered forest tree species (Sorbus torminalis) based on multi-scale investigations of populations genetic structure for chloroplast DNA. In Genetic Response of Forest Systems to Changing Environmental Conditions, 1st ed.; Muller-Starck, G., Schubert, R., Eds.; Kluwer Academic Publisher: Dodrecht, The Netherlands, 2001; pp. 271–280. [Google Scholar]
- Šarhanová, P.; Vašut, R.J.; Dančák, M.; Bureš, P.; Trávniček, B. New insight into the variability of reproduction modes in European populations of Rubus subg. Rubus: How sexual are polyploid brambles? Sex. Plant Reprod. 2012, 25, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Richards, A.J. Apomixis in flowering plants: An overview. Phil. Trans. R. Soc. Lond. B 2003, 358, 1085–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spillane, C.; Steimer, A.; Grossniklauss, U. Apomixis in agriculture: The quest for clonal seeds. Sex. Plant Reprod. 2001, 14, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Ferreira de Carvalho, J.; Oplaat, C.; Pappas, N.; Derks, M.; Ridder, D.; de Verhoeven, J.F. Heritable gene expression differences between apomictic clone members in Taraxacum officinale: Insight into early stages of evolutionary divergence in asexual plants. BMC Genom. 2016, 17, 203. [Google Scholar] [CrossRef] [Green Version]
- Wilschut, R.A.; Oplaat, C.; Snoek, L.B.; Kirschner, J.; Verhoeven, K.J.F. Natural epigenetic variation contributes to heritable flowering divergence in a widespread asexual dandelion lineage. Mol. Ecol. 2016, 25, 1759–1768. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, J.; Gibbard, P.L.; Hughes, P.D. Quaternary Glaciations–Extent and Chronology: A Closer Look, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Hampe, A.; Arroyo, J.; Jordano, P.; Petit, R.J. Rangewide phylogeography of a bird-dispersed Eurasian shrub: Contrasting Mediterranean and temperate glacial refugia. Mol. Ecol. 2003, 12, 3415–3426. [Google Scholar] [CrossRef] [PubMed]
- Magri, D.; Vendramin, G.G.; Comps, B.; Dupanloup, I.; Geburek, T.; Gömöry, D.; Latałowa, M.; Litt, T.; Paule, L.; Roure, J.M.; et al. A new scenario for the Quaternary history of European beech populations: Palaeobotanical evidence and genetic consequences. New Phytol. 2006, 171, 199–221. [Google Scholar] [CrossRef]
- Grivet, D.; Petit, R.J. Chloroplast DNA phylogeography of the hornbeam in Europe: Evidence for a bottleneck at the outset of postglacial colonization. Conserv. Genet. 2003, 4, 47–56. [Google Scholar] [CrossRef]
- Mogensen, H.L. Invited special paper: The hows and whys of cytoplasmic inheritance in seed plants. Am. J. Bot. 1996, 83, 383–404. [Google Scholar] [CrossRef]
- Raspé, O. Inheritance of the chloroplast genome in Sorbus aucuparia L. (Rosaceae). J. Hered. 2001, 92, 507–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chester, M.; Cowan, R.S.; Fay, M.F.; Rich, T.C.G. Parentage of endemic Sorbus L. (Rosaceae) species in the British Isles: Evidence from plastid DNA. Bot. J. Linn. Soc. 2007, 154, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Karbstein, K.; Tomasello, S.; Hodač, L.; Wagner, N.; Marinček, P.; Barke, B.H.; Paetzold, C.; Hörandl, E. Untying Gordian knots: Unraveling reticulate polyploid plant evolution by genomic data using the large Ranunculus auricomus species complex. New Phytol. 2022, 235, 2081–2098. [Google Scholar] [CrossRef] [PubMed]
- Soltis, P.S.; Soltis, D.E. Ancient WGD events as drivers of key innovations in angiosperms. Curr. Opin. Plant Biol. 2016, 30, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Melichárková, A.; Španiel, S.; Marhold, K.; Hurdu, B.-I.; Drescher, A.; Zozomová-Lihová, J. Diversification and independent polyploid origins in the disjunct species Alyssum repens from the Southeastern Alps and the Carpathians. Am. J. Bot. 2019, 106, 1499–1518. [Google Scholar] [CrossRef]
- Cushman, K.R.; Burgess, M.B.; Doucette, E.T.; Nelson, G.A.; Campbell, C.S. Species delimitation in tetraploid, apomictic Amelanchier (Rosaceae). Syst. Bot. 2017, 42, 234–256. [Google Scholar] [CrossRef]
- Thapa, R.; Bayer, R.J.; Mandel, J.R. Genetic diversity, population structure, and ancestry estimation in the Antennaria rosea (Asteraceae: Gnaphalieae) polyploid agamic complex. Taxon 2021, 70, 139–152. [Google Scholar] [CrossRef]
- Liston, A.; Weitmier, K.A.; Letelier, L.; Podani, J.; Zong, Y.; Liu, L.; Dickinson, T.A. Phylogeny of Crataegus (Rosaceae) based on 257 nuclear loci and chloroplast genomes: Evaluating the impact of hybridization. PeerJ 2021, 9, e12418. [Google Scholar] [CrossRef]
- Majeský, L.; Krahulec, F.; Vašut, R.J. How apomictic taxa are treated in current taxonomy: A review. Taxon 2017, 66, 117–1140. [Google Scholar] [CrossRef] [Green Version]
- Levin, J.; Fay, M.F.; Pellicer, J.; Hedrén, M. Multiple independent origins of intermediate species between Sorbus aucuparia and S. hybrida (Rosaceae) in the Baltic region. Nord. J. Bot. 2018, 35. [Google Scholar] [CrossRef]
- Rich, T.C.G.; Gree, D.; Houston, L.; Lepší, M.; Ludwig, S.; Pellicer, J. British Sorbus (Rosaceae): Six new species, two hybrids and a new subgenus. New J. Bot. 2014, 4, 2–12. [Google Scholar] [CrossRef]
- Rieseberg, L.H. Hybrid origins of plant species. Ann. Rev. Ecol. Syst. 1997, 28, 359–389. [Google Scholar] [CrossRef] [Green Version]
- Hegarty, M.J.; Batstone, T.O.M.; Barker, G.L.; Edwards, K.J.; Abbott, R.J.; Hiscock, S.J. Nonadditive changes to cytosine methylation as a consequence of hybridization and genome duplication in Senecio (Asteraceae). Mol. Ecol. 2011, 20, 105–113. [Google Scholar] [CrossRef]
- Ennos, R.A.; French, G.C.; Hollingsworth, P.M. Conserving taxonomic complexity. Trends Ecol. Evol. 2005, 20, 164–168. [Google Scholar] [CrossRef]
- Ennos, R.A.; Whitlock, R.; Fay, M.F.; Jones, B.; Neaves, L.E.; Payne, R.; Taylor, I.; De Vere, N.; Hollingsworth, P.M. Process-based species action plans: An approach to conserve contemporary evolutionary processes that sustain diversity in taxonomically complex groups. Bot. J. Linn. Soc. 2012, 168, 194–203. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MG | N | AFLP Cluster | N Per Locality | Loci | Xo | Xe | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| MSS13 | MSS5 | CH01F02 | D11 | MSS16 | H10 | ||||||
| 1 | 5 | d | 2(5) | 189, 193, 195 | 124, 126, 130 | 186, 200 | 143, 151 | 154, 158, 162 | 86 | 3 | 4x a |
| 2 | 6 | j | 3(3), 4(3) | 193, 197 | 110, 124, 126 | 186, 190, 196, 200 | 137, 151 | 154, 158, 160, 162 | 92 | 4 | - |
| 3 | 2 | l | 6(2) | 191, 193, 200 | 124, 134 | 194, 196 | 149, 151 | 154, 158, 160 | 82, 88, 92 | 3 | 4x a |
| 4 | 1 | l | 7(1) | 193, 195, 201 | 122, 124, 126, 130 | 190, 196, 198, 200 | 151, 157 | 156, 158 | - | 4 | - |
| 5 | 23 | f | 7(1), 11(4), 13(5), 14(1), 15(2), 17(5), 18(5) | 191, 195, 201 | 124, 126, 150 | 186, 190, 196, 200 | 143, 151 | 156, 158 | 104, 106 | 4 | 4x a |
| 6 | 1 | f | 7(1) | 191, 195, 201 | 124, 126, 150 | 186, 190, 196, 200 | 143, 151 | 154, 158 | 104, 106 | 4 | - |
| 7 | 2 | f | 8(2) | 193, 197, 201 | 124, 126 | 186, 196 | 143, 151 | 156, 158 | 104, 106 | 3 | 4x a |
| 8 | 2 | f | 8(2) | 191, 197, 201 | 124, 126 | 186, 190, 196 | 143, 151 | 156, 158 | 106 | 3 | 4x a |
| 9 | 5 | f | 9(5) | 191, 197, 201 | 124, 126 | 186, 190, 196 | 143, 151 | 154, 158, 166 | 104, 106 | 3 | 4x a |
| 10 | 1 | f | 11(1) | 191, 195, 201 | 124, 126, 150 | 186, 190, 196, 200 | 143, 151 | 156, 158 | 102, 104 | 4 | 4x a |
| 11 | 5 | c | 12(5) | 191, 193, 195 | 110, 112, 124 | 188, 196, 202 | 147, 153 | 154, 158, 162 | 92 | 3 | - |
| 12 | 9 | c | 14(4), 25(5) | 191, 193, 195 | 110, 112, 124 | 188, 196, 202 | 147, 151 | 154, 158, 162 | 92 | 3 | 4x a |
| 13 | 2 | c | 14(2) | 191, 193, 195 | 110, 112, 124 | 188, 196, 202 | 147, 151 | 154, 158, 162 | 80, 90 | 3 | 4x a |
| 14 | 2 | g | 15(2) | 191, 193 | 112, 120, 126 | 186, 190, 196, 200 | 151 | 154, 158, 164 | 98, 100 | 4 | 4x a |
| 15 | 1 | g | 15(1) | 191, 193 | 112, 120, 126 | 186, 190, 196, 200 | 151 | 154, 158, 164 | 102 | 4 | 4x a |
| 16 | 2 | i | 19(2) | 191, 193, 195 | 112, 120, 126 | 188, 190, 196, 200 | 147, 149, 151 | 154, 158, 160, 162 | 92 | 4 | - |
| 17 | 3 | h | 20(3) | - | 110, 112, 126 | 186, 190, 196, 200 | 149, 151 | 154, 160 | 96 | 4 | 4x d |
| 18 | 4 | k | 21(4) | 193, 201 | 124, 126, 132 | 186, 190, 196 | 131, 151 | 154, 162 | 90 | 3 | 4x d |
| 19 | 1 | l | 22(1) | 191, 193, 201 | 112, 124, 126 | 190, 196, 200 | 149, 151 | 156, 158, 160, 162 | 92 | 4 | 4x c |
| 20 | 5 | b | 23(5) | 191, 193, 195 | 110, 112, 124 | 186, 196, 208 | 131, 151 | 154, 158, 162 | 102, 104 | 3 | - |
| 21 | 5 | e | 24(5) | 191, 193, 195 | 124 | 190, 196 | 141, 147, 151 | 154, 158, 160 | 98, 100 | 3 | - |
| 22 | 8 | c | 26(5), 27(3) | 191, 193, 195 | 110, 112, 124 | 186, 196, 200 | 147, 151 | 154, 158, 162 | 92 | 3 | - |
| 23 | 5 | a | 31(3), 33(1), 34(1) | 193, 197, 201 | 124, 132 | 186, 196 | 147, 151 | 154, 160, 162 | 100, 102 | 3 | - |
| 24 | 8 | w | 36(5), 37(3) | 191, 193, 195 | 118, 128, 134 | 186, 196, 200 | 147, 149, 151 | 154, 162 | 98, 100 | 3 | 4x b |
| 25 | 2 | w | 37(2) | 191, 193, 195 | 118, 128, 134 | 186, 196, 200 | 147, 149, 151 | 154, 162 | 96, 98 | 3 | 4x b |
| 110 | |||||||||||
| Locality No. | Affiliation in AFLP NNet | Taxon | N | Genome Size (2C pg) | Genome Size (1Cx pg) | DNA Ploidy Level (2n) | N Seeds | Embryo (2C pg) | Endosperm (2C pg) | Embryo: Endosperm Ploidy | Seed Origin |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | d | S. austriaca | 3 | 2.81–2.82 | 0.70–0.71 | 4x | - | ||||
| 5 | S. aucuparia | S. aucuparia | 2 | 1.37–1.40 | 0.69–0.70 | 2x | 2 | 1.36–1.41 | 2.07–2.14 | 2x:3x | Sexual |
| 6 | l | S. aria × S. austriaca | 1 | 2.69 | 0.67 | 4x | 1 | 2.67 | 8.13 | 4x:12x | Apomictic |
| S. aria | S. aria | 1 | 2.71 | 0.68 | 4x | 1 | 2.77 | 8.15 | 4x:12x | Apomictic | |
| 8 | f | S. austriaca | 1 | 2.73 | 0.68 | 4x | 1 | 2.73 | 8.08 | 4x:12x | Apomictic |
| 9 | f | S. austriaca | 1 | 2.79 | 0.70 | 4x | 1 | 2.73 | 8.10 | 4x:12x | Apomictic |
| 11 | f | S. austriaca | 5 | 2.69–2.73 | 0.67–0.68 | 4x | 1 | 2.72 | 8.32 | 4x:12x | Apomictic |
| 13 | f | S. austriaca | 3 | 2.75–2.81 | 0.69–0.70 | 4x | 2 | 2.73–2.74 | 8.09–8.23 | 4x:12x | Apomictic |
| 14 | f | S. austriaca | 1 | 2.56 | 0.64 | 4x | 1 | 2.81 | 8.46 | 4x:12x | Apomictic |
| c | S. austriaca | 6 | 2.52–2.80 | 0.63–0.70 | 4x | 19 (7 *) | 2.69–2.80 | 7.99–8.65 | 4x:12x | Apomictic | |
| 1 | 2.65 | 6.70 | 4x:10x | Apomictic | |||||||
| S. aria | S. aria | 2 | 1.40–1.41 | 0.70–0.71 | 2x | 2 | 1.41–1.43 | 2.12–2.15 | 2x:3x | Sexual | |
| 1 | 2.51 | 0.63 | 4x | 1 | 2.74 | 8.34 | 4x:12x | Apomictic | |||
| S. aucuparia | S. aucuparia | 3 | 1.36–1.43 | 0.68–0.72 | 2x | 3 | 1.36–1.41 | 2.03–2.13 | 2x:3x | Sexual | |
| 15 | f, g | S. austriaca | 5 | 2.56–2.72 | 0.64–0.68 | 4x | 1 | 2.67 | 8.12 | 4x:12x | Apomictic |
| 16 | S. aria | S. aria | 3 | 1.38–1.40 | 0.69–0.70 | 2x | 2 | 1.37–1.43 | 2.06–2.13 | 2x:3x | Sexual |
| 17 | f | S. austriaca | 3 | 2.69–2.74 | 0.67–0.69 | 4x | 1 | 2.66 | 8.11 | 4x:12x | Apomictic |
| S. aria | S. aria | 1 | 1.42 | 0.71 | 2x | - | |||||
| 35 | S. aria | S. aria | 3 | 1.39–1.51 | 0.70–0.76 | 2x | - | ||||
| 45 | 40 (7 *) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajrudinović-Bogunić, A.; Frajman, B.; Schönswetter, P.; Siljak-Yakovlev, S.; Bogunić, F. Apomictic Mountain Whitebeam (Sorbus austriaca, Rosaceae) Comprises Several Genetically and Morphologically Divergent Lineages. Biology 2023, 12, 380. https://doi.org/10.3390/biology12030380
Hajrudinović-Bogunić A, Frajman B, Schönswetter P, Siljak-Yakovlev S, Bogunić F. Apomictic Mountain Whitebeam (Sorbus austriaca, Rosaceae) Comprises Several Genetically and Morphologically Divergent Lineages. Biology. 2023; 12(3):380. https://doi.org/10.3390/biology12030380
Chicago/Turabian StyleHajrudinović-Bogunić, Alma, Božo Frajman, Peter Schönswetter, Sonja Siljak-Yakovlev, and Faruk Bogunić. 2023. "Apomictic Mountain Whitebeam (Sorbus austriaca, Rosaceae) Comprises Several Genetically and Morphologically Divergent Lineages" Biology 12, no. 3: 380. https://doi.org/10.3390/biology12030380