
Marine Plankton during the Polar Night: Environmental Predictors of Spatial Variability
 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
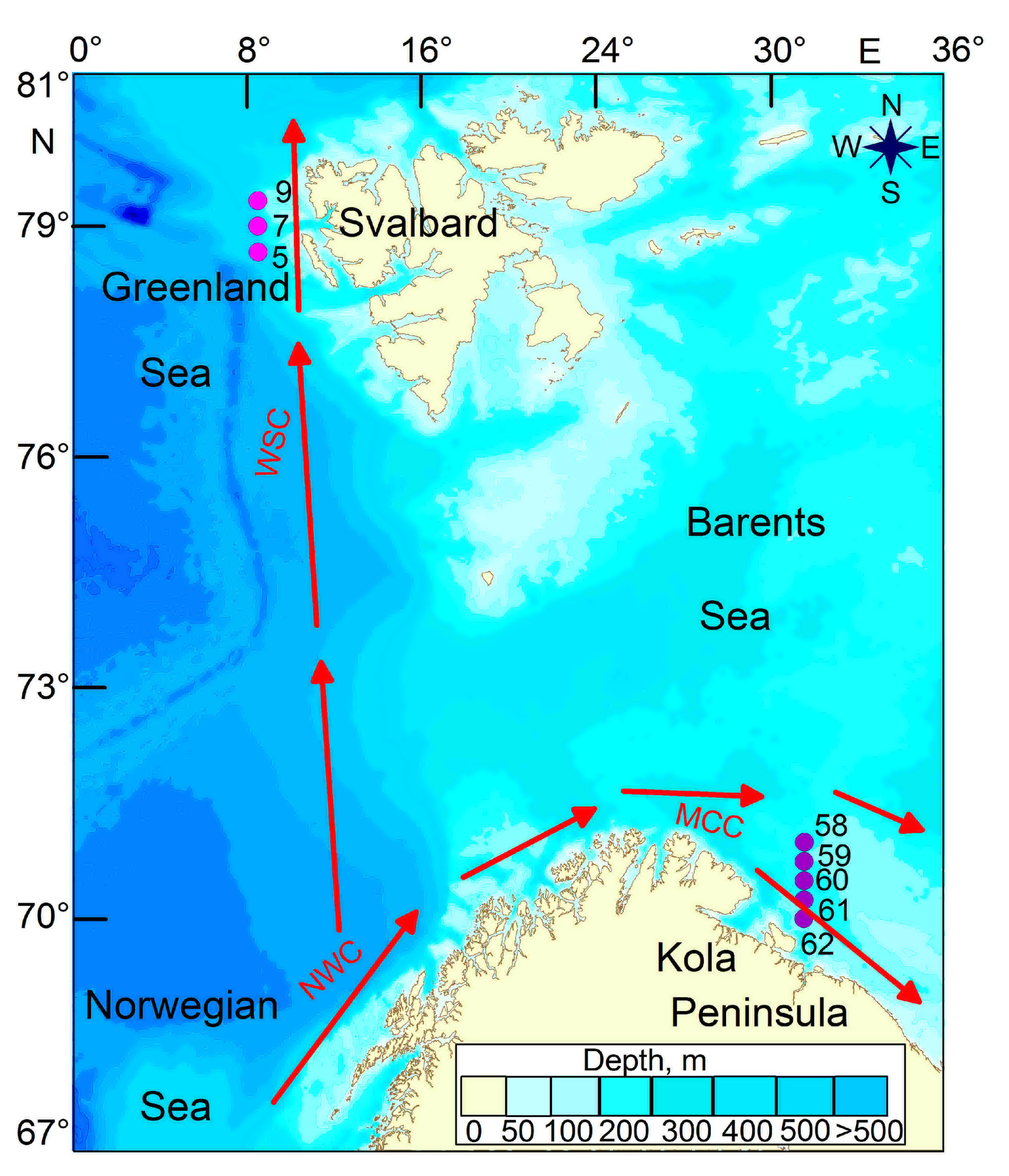
2.1. Sampling and Processing
2.2. Laboratory Procedures
2.3. Statistical Analyses
3. Results
3.1. Environmental Conditions
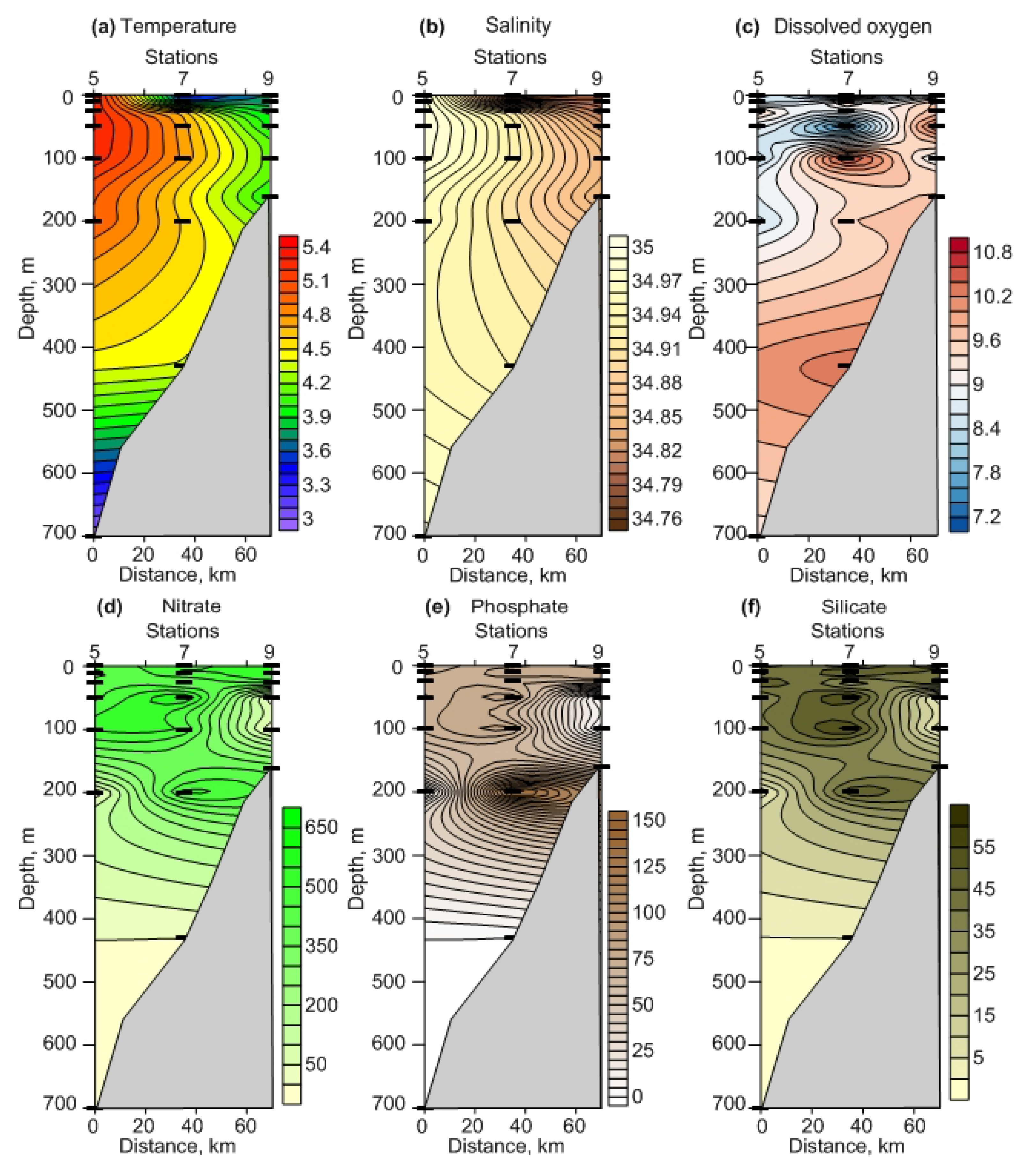
3.1.1. Hydrology
3.1.2. Dissolved Oxygen and Nutrients
3.2. Plankton Distribution
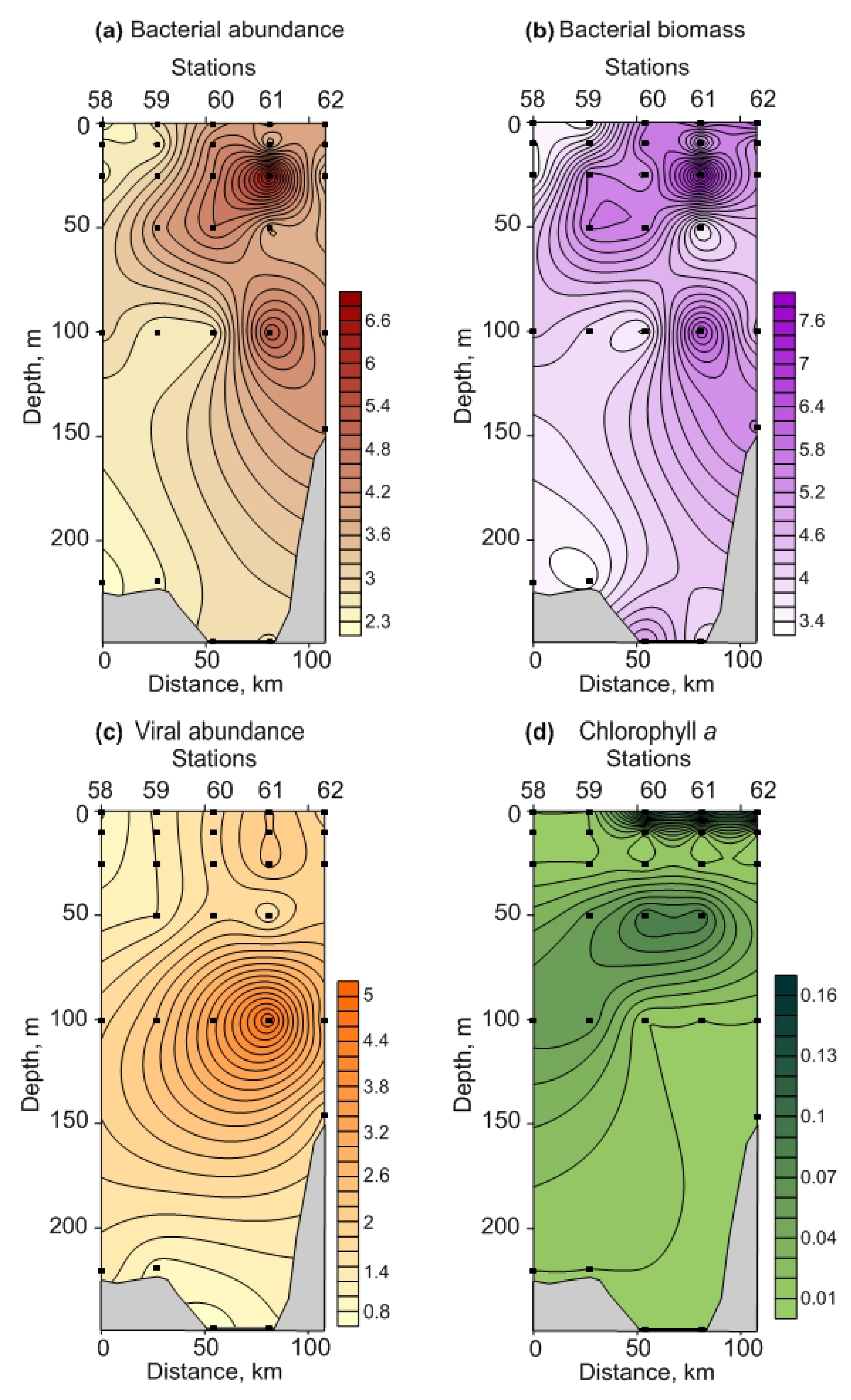
3.2.1. Microbial Parameters
3.2.2. Chlorophyll a
3.2.3. Zooplankton
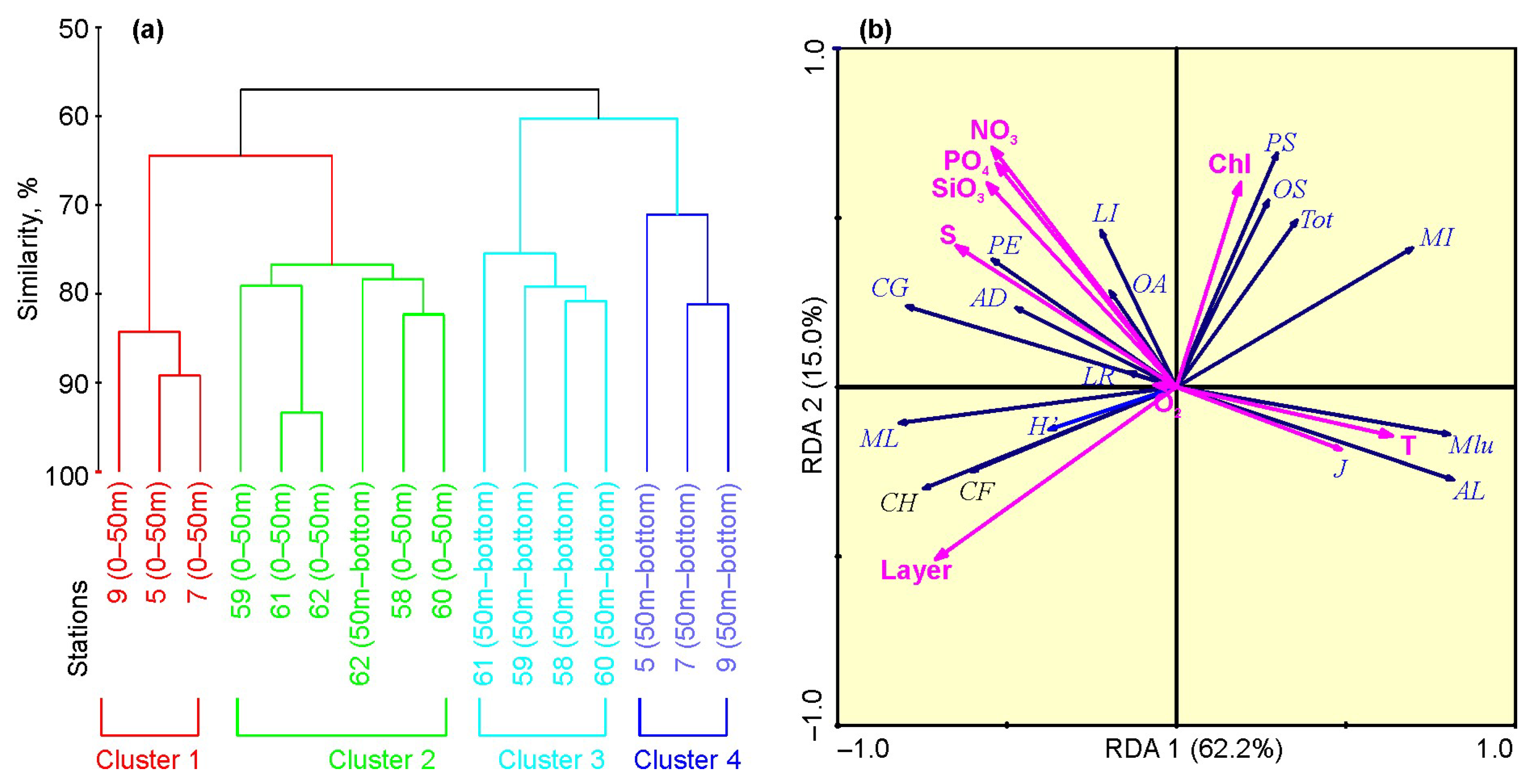
3.2.4. Spatial Separation of the Plankton
3.3. Environmental Influences on Plankton and Biotic Interactions
4. Discussion
4.1. Environmental Conditions
4.2. Microbial Parameters
4.3. Phytoplankton
4.4. Zooplankton
4.5. Environmental Control of Plankton and Biotic Interactions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polyakov, I.V.; Pnyushkov, A.; Alkire, M.; Ashik, I.M.; Baumann, T.M.; Carmack, E.C.; Goszczko, I.; Guthrie, J.D.; Ivanov, V.V.; Kanzow, T.; et al. Greater role for Atlantic inflows on sea-ice loss in the Eurasian Basin of the Arctic Ocean. Science 2017, 356, 285–291. [Google Scholar] [CrossRef] [Green Version]
- ICES. Working Group on the Integrated Assessments of the Barents Sea (WGIBAR). ICES Sci. Rep. 2021, 3, 236. [Google Scholar]
- Isaksen, K.; Nordli, Ø.; Ivanov, B.; Køltzow, M.A.; Aaboe, S.; Gjelten, H.M.; Mezghani, A.; Eastwoodl, S.; Førland, E.; Benestad, R.E.; et al. Exceptional warming over the Barents area. Sci. Rep. 2022, 12, 9371. [Google Scholar] [CrossRef]
- Rantanen, M.; Karpechko, A.Y.; Lipponen, A.; Nordling, K.; Hyvärinen, O.; Ruosteenoja, K.; Vihma, T.; Laaksonen, A. The Arctic has warmed nearly four times faster than the globe since 1979. Commun. Earth Environ. 2022, 3, 168. [Google Scholar] [CrossRef]
- ICES. Working Group on the Integrated Assessments of the Barents Sea (WGIBAR). ICES Sci. Rep. 2022, 4, 1–235. [Google Scholar]
- Polyakov, I.V.; Alkire, M.B.; Bluhm, B.A.; Brown, K.A.; Carmack, E.C.; Chierici, M.; Danielson, S.L.; Ellingsen, I.; Ershova, E.A.; Gårdfeldt, K.; et al. Borealization of the Arctic Ocean in response to anomalous advection from sub-arctic seas. Front. Mar. Sci. 2020, 7, 491. [Google Scholar] [CrossRef]
- Arrigo, K.R.; van Dijken, G.L. Continued increases in Arctic Ocean primary production. Prog. Oceanogr. 2015, 136, 60–70. [Google Scholar] [CrossRef]
- Lewis, K.M.; Van Dijken, G.L.; Arrigo, K.R. Changes in phytoplankton concentration now drive increased Arctic Ocean primary production. Science 2020, 369, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Dalpadado, P.; Arrigo, K.R.; van Dijken, G.L.; Skjoldal, H.R.; Bagøien, E.; Dolgov, A.V.; Prokopchuk, I.P.; Sperfeld, E. Climate effects on temporal and spatial dynamics of phytoplankton and zooplankton in the Barents Sea. Progr. Oceanogr. 2020, 182, 102320. [Google Scholar] [CrossRef]
- Slagstad, D.; Wassmann, P.; Ellingsen, I. Physical constrains and productivity in the future Arctic Ocean. Front. Mar. Sci. 2015, 2, 85. [Google Scholar] [CrossRef] [Green Version]
- Dvoretsky, V.G.; Vodopianova, V.V.; Bulavina, A.S. Effects of Climate Change on Chlorophyll a in the Barents Sea: A Long-Term Assessment. Biology 2023, 12, 119. [Google Scholar] [CrossRef]
- Lind, S.; Ingvaldsen, R.B.; Furevik, T. Arctic warming hotspot in the northern Barents Sea linked to declining sea-ice import. Nat. Clim. Chang. 2018, 8, 634–639. [Google Scholar] [CrossRef]
- Ardyna, M.; Arrigo, K. Phytoplankton dynamics in a changing Arctic Ocean. Nat. Clim. Change 2020, 10, 892–903. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Copepod assemblages in a large Arctic coastal area: A baseline summer study. Diversity 2023, 15, 81. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Epiplankton in the Barents Sea: Summer variations of mesozooplankton biomass, community structure and diversity. Cont. Shelf Res. 2013, 52, 1–11. [Google Scholar] [CrossRef]
- Ingvaldsen, R.B.; Assmann, K.M.; Primicerio, R.; Fossheim, M.; Polyakov, I.V.; Dolgov, A.V. Physical manifestations and ecological implications of Arctic Atlantification. Nat. Rev. Earth Environ. 2021, 2, 874–889. [Google Scholar] [CrossRef]
- Tesi, T.; Muschitiello, F.; Mollenhauer, G.; Miserocchi, S.; Langone, L.; Ceccarelli, C.; Capotondi, L. Rapid Atlantification along the Fram Strait at the beginning of the 20th century. Sci. Adv. 2021, 7, eabj2946. [Google Scholar] [CrossRef]
- Evseeva, O.Y.; Ishkulova, T.G.; Dvoretsky, A.G. Environmental drivers of an intertidal bryozoan community in the Barents Sea: A case study. Animals 2022, 12, 552. [Google Scholar] [CrossRef]
- Evseeva, O.Y.; Dvoretsky, A.G. Shallow-water bryozoan communities in a glacier fjord of West Svalbard, Norway: Species composition and effects of environmental factors. Biology 2023, 12, 185. [Google Scholar] [CrossRef]
- Eriksen, E.; Skjoldal, H.R.; Gjøsæter, H.; Primicerio, R. Spatial and temporal changes in the Barents Sea pelagic compartment during the recent warming. Prog. Oceanogr. 2017, 151, 206–226. [Google Scholar] [CrossRef]
- Weydmann, A.; Walczowski, W.; Carstensen, J.; Kwaśniewski, S. Warming of Subarctic waters accelerates development of a key marine zooplankton Calanus Finmarchicus. Glob. Chang. Biol. 2018, 24, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Dvoretsky, V.G.; Dvoretsky, A.G. Coastal mesozooplankton assemblages during spring bloom in the eastern Barents Sea. Biology 2022, 11, 204. [Google Scholar] [CrossRef] [PubMed]
- Dvoretsky, V.G.; Dvoretsky, A.G. Mesozooplankton in the Kola Transect (Barents Sea): Autumn and winter structure. J. Sea Res. 2018, 142, 18–22. [Google Scholar] [CrossRef]
- Raymont, J.E.G. Plankton and Productivity in the Oceans, 2nd ed.; Pergamon Press: Southampton, UK, 1983; Volume 2. [Google Scholar]
- Castellani, C.; Edwards, M. Marine Plankton: A Practical Guide to Ecology, Methodology, and Taxonomy; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Matishov, G.G. (Ed.) Life and Environmental Conditions in the Pelagial of the Barents Sea; Kola Science Center: Apatity, Russia, 1985. (In Russian) [Google Scholar]
- Wassmann, P.; Reigstad, M.; Haug, T.; Rudels, B.; Carroll, M.L.; Hop, H.; Gabrielsen, G.W.; Falk-Petersen, S.; Denisenko, S.G.; Arashkevich, E.; et al. Food webs and carbon flux in the Barents Sea. Progr. Oceanogr. 2006, 71, 232–287. [Google Scholar] [CrossRef]
- Jakobsen, T.; Ozhigin, V.K. (Eds.) The Barents Sea: Ecosystem, Resources, Management: Half a Century of Russian-Norwegian Cooperation; Tapir Academic Press: Trondheim, Norway, 2011. [Google Scholar]
- Sakshaug, E.; Johnsen, G.; Kovacs, K. (Eds.) Ecosystem Barents Sea; Tapir Academic Press: Trondheim, Norway, 2009. [Google Scholar]
- Makarevich, P.; Druzhkova, E.; Larionov, V. Primary producers of the Barents Sea. In Diversity of Ecosystems; Mahamane, A., Ed.; In Tech: Rijeka, Croatia, 2012; pp. 367–392. [Google Scholar]
- Pavlova, L.V.; Zuyev, Y.A.; Dvoretsky, A.G. Shallow-water benthic communities on soft bottoms of a sub-arctic fjord (Southern Barents Sea, Russia) along a gradient of ecological factors. Diversity 2023, 15, 84. [Google Scholar] [CrossRef]
- Pavlova, L.V.; Dvoretsky, G. Prey selectivity in juvenile red king crabs from the coastal Barents Sea. Diversity 2022, 14, 568. [Google Scholar] [CrossRef]
- Azam, F.; Malfatti, F. Microbial structuring of marine ecosystems. Nat. Rev. Microbiol. 2007, 5, 782–791. [Google Scholar] [CrossRef]
- Fuhrman, J.A.; Cram, J.A.; Needham, D.M. Marine microbial community dynamics and their ecological interpretation. Nat. Rev. Microbiol. 2015, 13, 133–146. [Google Scholar] [CrossRef]
- Wommack, K.E.; Colwell, R.R. Virioplankton: Viruses in aquatic ecosystems. Microbiol. Mol. Biol. Rev. 2000, 64, 69–114. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, A.E.; Howard-Varona, C.; Needham, D.M.; John, S.G.; Worden, A.Z.; Sullivan, M.B.; Waldbauer, J.R.; Coleman, M.L. Metabolic and biogeochemical consequences of viral infection in aquatic ecosystems. Nat. Rev. Microbiol. 2020, 18, 21–34. [Google Scholar] [CrossRef]
- Hop, H.; Falk-Petersen, S.; Svendsen, H.; Kwasniewski, S.; Pavlov, V.; Pavlova, O.; Soreide, J.E. Physical and biological characteristics of the pelagic system across Fram Strait to Kongsfjorden. Prog. Oceanogr. 2006, 71, 182–231. [Google Scholar] [CrossRef]
- Randelhoff, A.; Reigstad, M.; Chierici, M.; Sundfjord, A.; Ivanov, V.; Cape, M.; Vernet, M.; Tremblay, J.É.; Bratbak, G.; Kristiansen, S. Seasonality of the Physical and Biogeochemical Hydrography in the Inflow to the Arctic Ocean through Fram Strait. Front. Mar. Sci. 2018, 5, 224. [Google Scholar] [CrossRef] [Green Version]
- Dvoretsky, A.G.; Dvoretsky, V.G. Inter-annual dynamics of the Barents Sea red king crab (Paralithodes camtschaticus) stock indices in relation to environmental factors. Polar Sci. 2016, 10, 541–552. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibiotic communities of common crab species in the coastal Barents Sea: Biodiversity and infestation patterns. Diversity 2022, 14, 6. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibionts of an introduced king crab in the Barents Sea: A second five-year study. Diversity 2023, 15, 29. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Commercial fish and shellfish in the Barents Sea: Have introduced crab species affected the population trajectories of commercial fish? Rev. Fish Biol. Fish. 2015, 25, 297–322. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Effects of environmental factors on the abundance, biomass, and individual weight of juvenile red king crabs in the Barents Sea. Front. Mar. Sci. 2020, 7, 726. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Structure of mesozooplankton community in the Barents Sea and adjacent waters in August 2009. J. Nat. Hist. 2013, 47, 2095–2114. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Arctic marine mesozooplankton at the beginning of the polar night: A case study for southern and south-western Svalbard waters. Polar Biol. 2020, 43, 71–79. [Google Scholar] [CrossRef]
- Nöthig, E.M.; Ramondenc, S.; Haas, A.; Hehemann, L.; Walter, A.; Bracher, A.; Lalande, C.; Metfies, K.; Peeken, I.; Bauerfeind, E.; et al. Summertime Chlorophyll a and Particulate Organic Carbon Standing Stocks in Surface Waters of the Fram Strait and the Arctic Ocean (1991–2015). Front. Mar. Sci. 2020, 7, 350. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Checklist of fauna found in zooplankton samples from the Barents Sea. Polar Biol. 2010, 33, 991–1005. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Spatial variations in reproductive characteristics of the small copepod Oithona similis in the Barents Sea. Mar. Ecol. Prog. Ser. 2009, 386, 133–146. [Google Scholar]
- Gluchowska, M.; Dalpadado, P.; Beszczynska-Möller, A.; Olszewska, A.; Ingvaldsen, R.B.; Kwasniewski, S. Interannual zooplankton variability in the main pathways of the Atlantic water flow into the Arctic Ocean (Fram Strait and Barents Sea branches). ICES J. Mar. Sci. 2017, 74, 1921–1936. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Early winter mesozooplankton of the coastal south-eastern Barents Sea. Estuar. Coast. Shelf Sci. 2015, 152, 116–123. [Google Scholar]
- Nöthig, E.M.; Bracher, A.; Engel, A.; Metfies, K.; Niehoff, B.; Peeken, I.; Bauerfeind, E.; Cherkasheva, A.; Gabler-Schwarz, S.; Hardge, K.; et al. Summertime plankton ecology in Fram Strait a compilation of long- and short-term observations. Polar Res. 2015, 34, 23349. [Google Scholar]
- Dvoretsky, V.G.; Dvoretsky, A.G. Ecology and distribution of red king crab larvae in the Barents Sea: A review. Water 2022, 14, 2328. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Estimated copepod production rate and structure of mesozooplankton communities in the coastal Barents Sea during summer–autumn 2007. Polar Biol. 2012, 35, 1321–1342. [Google Scholar] [CrossRef]
- Trudnowska, E.; Sagan, S.; Błachowiak-Samołyk, K. Spatial variability and size structure of particles and plankton in the Fram Strait. Prog. Oceanogr. 2018, 168, 1–12. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Mesozooplankton structure in Dolgaya Bay (Barents Sea). Polar Biol. 2010, 33, 703–708. [Google Scholar]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer mesozooplankton distribution near Novaya Zemlya (eastern Barents Sea). Polar Biol. 2009, 32, 719–731. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer mesozooplankton structure in the Pechora Sea (south-eastern Barents Sea). Estuar. Coast. Shelf Sci. 2009, 84, 11–20. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer-fall macrozooplankton assemblages in a large Arctic estuarine zone (south-eastern Barents Sea): Environmental drivers of spatial distribution. Mar. Environ. Res. 2022, 173, 105498. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Life cycle of Oithona similis (Copepoda: Cyclopoida) in Kola Bay (Barents Sea). Mar. Biol. 2009, 156, 1433–1446. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Venger, M.P.; Vashchenko, A.V.; Maksimovskaya, T.M.; Ishkulova, T.G.; Vodopianova, V.V. Pelagic bacteria and viruses in a high Arctic region: Environmental control in the autumn period. Biology 2022, 11, 845. [Google Scholar] [CrossRef] [PubMed]
- Berge, J.; Renaud, P.E.; Darnis, G.; Cottier, F.; Last, K.; Gabrielsen, T.M.; Johnsen, G.; Seuthe, L.; Weslawski, J.M.; Leu, E.; et al. In the dark: A review of ecosystem processes during the Arctic polar night. Prog. Oceanogr. 2015, 139, 258–271. [Google Scholar] [CrossRef] [Green Version]
- Berge, J.; Daase, M.; Hobbs, L.; Falk-Petersen, S.; Darnis, G.; Søreide, J.E. Zooplankton in the polar night. In Polar Night Marine Ecology; Springer: Cham, Switzerland, 2020; pp. 113–159. [Google Scholar]
- Grenvald, J.C.; Callesen, T.A.; Daase, M.; Hobbs, L.; Darnis, G.; Renaud, P.E.; Cottier, F.; Nielsen, T.G.; Berge, J. Plankton community composition and vertical migration during polar night in Kongsfjorden. Polar Biol. 2016, 39, 1879–1895. [Google Scholar] [CrossRef]
- Barth-Jensen, C.; Daase, M.; Ormańczyk, M.R.; Varpe; Kwaśniewski, S.; Svensen, C. High abundances of small copepods early developmental stages and nauplii strengthen the perception of a non-dormant Arctic winter. Polar Biol. 2022, 45, 675–690. [Google Scholar] [CrossRef]
- Wietz, M.; Bienhold, C.; Metfies, K.; Torres-Valdés, S.; von Appen, W.J.; Salter, I.; Boetius, A. The polar night shift: Seasonal dynamics and drivers of Arctic Ocean microbiomes revealed by autonomous sampling. ISME Comm. 2021, 1, 76. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Chemical and Biological Methods for Sea Water Analysis; Pergamon Press: New York, NY, USA, 1992. [Google Scholar]
- Aminot, A.; Rey, F. Standard Procedure for the Determination of Chlorophyll A by Spectroscopic Methods; International Council for the Exploration of the Sea: Copenhagen, Denmark, 2000. [Google Scholar]
- Porter, K.G.; Feig, Y.S. The use of DAPI for identifying and counting aquatic microflora. Limnol. Oceanogr. 1980, 25, 943–948. [Google Scholar] [CrossRef]
- Norland, S. The relationships between biomass and volume of bacteria. In Handbook of Methods in Aquatic Microbial Ecology; Kemp, P., Sherr, B., Sherr, E., Cole, J., Eds.; Lewis Publ.: Boca Raton, FL, USA, 1993; pp. 303–308. [Google Scholar]
- Noble, R.T.; Fuhrman, J.A. Virus decay and its causes in coastal waters. Appl. Environ. Microbiol. 1997, 63, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer macrozooplankton assemblages of Arctic shelf: A latitudinal study. Cont. Shelf Res. 2019, 188, 103967. [Google Scholar] [CrossRef]
- Chislenko, L.L. Nomogrammes to Determine Weights of Aquatic Organisms Based on the Size and Form of Their Bodies (Marine Mesobenthos and Plankton); Nauka Press: Leningrad, Russia, 1968. (In Russian) [Google Scholar]
- Berestovskij, E.G.; Anisimova, N.A.; Denisenk, C.G.; Luppowa, E.N.; Savinov, V.M.; Timofeev, S.F. Relationship between Size and Body Mass of Some Invertebrates and Fish of the North-East Atlantic; Academy of Sciences of the USSR, Murmansk Marine Biological Institute: Apatity, Russia, 1989. (In Russian) [Google Scholar]
- Hanssen, H. Mesozooplankton of the Laptev Sea and the adjacent eastern Nansen Basin–distribution and community structure in late summer. Ber. Polarforsch. 1997, 229, 1–131. [Google Scholar]
- Richter, C. Regional and seasonal variability in the vertical distribution of mesozooplankton in the Greenland Sea. Ber. Polarforsch. 1994, 154, 1–90. [Google Scholar]
- Harris, R.P.; Wiebe, P.H.; Lenz, J.; Skjoldal, H.R.; Huntley, M. (Eds.) ICES Zooplankton Methodology Manual; Academic Press: San Diego, CA, USA, 2000. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier Science: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Hammer; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Shannon, C.B.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1963. [Google Scholar]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Changes in Marine Communities: An Approach to Statistical Analysis and Interpretation; Plymouth Marine Laboratory, NERC: Plymouth, UK, 1994. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v5: User Manual/Tutorial; PRIMER-E Ltd.: Plymouth, UK, 2001. [Google Scholar]
- ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manualand CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice-Hall Inc.: Hoboken, NJ, USA, 1999. [Google Scholar]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models; Chapman and Hall: London, UK, 1983. [Google Scholar]
- Beszczynska-Möller, A.; Fahrbach, E.; Schauer, U.; Hansen, E. Variability in Atlantic water temperature and transport at the entrance to the Arctic Ocean, 1997–2010. ICES J. Mar. Sci. 2012, 69, 852–863. [Google Scholar] [CrossRef] [Green Version]
- Tuerena, R.E.; Hopkins, J.; Buchanan, P.J.; Ganeshram, R.S.; Norman, L.; von Appen, W.J.; Mahaffey, C. An Arctic strait of two halves: The changing dynamics of nutrient uptake and limitation across the Fram Strait. Glob. Biogeochem. Cycles 2021, 35, e2021GB006961. [Google Scholar] [CrossRef]
- Tuerena, R.E.; Mahaffey, C.; Henley, S.F.; De La Vega, C.; Norman, L.; Brand, T.; März, C. Nutrient pathways and their susceptibility to past and future change in the Eurasian Arctic Ocean. Ambio 2022, 51, 355–369. [Google Scholar] [CrossRef]
- Wulff, T.; Bauerfeind, E.; von Appen, W.J. Physical and ecological processes at a moving ice edge in the Fram Strait as observed with an AUV. Deep-Sea Res. I 2016, 115, 253–264. [Google Scholar] [CrossRef] [Green Version]
- Cardozo-Mino, M.G.; Fadeev, E.; Salman-Carvalho, V.; Boetius, A. Spatial Distribution of Arctic Bacterioplankton Abundance Is Linked to Distinct Water Masses and Summertime Phytoplankton Bloom Dynamics (Fram Strait, 79° N). Front. Microbiol. 2021, 12, 658803. [Google Scholar] [CrossRef] [PubMed]
- Seuthe, L.; Topper, B.; Reigstad, M.; Thyrhaug, R.; Vaquer-Sunyer, R. Microbial communities and processes in ice-covered Arctic waters of the northwestern Fram Strait (75 to 80 degrees N) during the vernal pre-bloom phase. Aquat. Microb. Ecol. 2011, 64, 253–266. [Google Scholar] [CrossRef]
- Rokkan Iversen, K.; Seuthe, L. Seasonal microbial processes in a high-latitude fjord (Kongsfjorden, Svalbard): I—Heterotrophic bacteria, picoplankton and nanoflagellates. Polar Biol. 2011, 34, 731–749. [Google Scholar] [CrossRef] [Green Version]
- Piontek, J.; Sperling, M.; Nöthig, E.M.; Engel, A. Regulation of bacterioplankton activity in Fram Strait (Arctic Ocean) during early summer: The role of organic matter supply and temperature. J. Mar. Syst. 2014, 132, 83–94. [Google Scholar] [CrossRef]
- Wilson, B.; Müller, O.; Nordmann, E.-L.; Seuthe, L.; Bratbak, G.; Øvreås, L. Changes in Marine Prokaryote Composition with Season and Depth Over an Arctic Polar Year. Front. Mar. Sci. 2017, 4, 95. [Google Scholar] [CrossRef] [Green Version]
- von Jackowski, A.; Grosse, J.; Nöthig, E.M.; Engel, A. Dynamics of organic matter and bacterial activity in the Fram Strait during summer and autumn. Phil. Trans. R. Soc. A 2020, 378, 20190366. [Google Scholar] [CrossRef]
- von Jackowski, A.; Becker, K.W.; Wietz, M.; Bienhold, C.; Zäncker, B.; Nöthig, E.M.; Engel, A. Variations of microbial communities and substrate regimes in the eastern Fram Strait between summer and fall. Environ. Microbiol. 2022, 24, 4124–4136. [Google Scholar] [CrossRef]
- Venger, M.P.; Kopylov, A.I.; Zabotkina, E.A.; Makarevich, P.R. The influence of viruses on bacterioplankton of the offshore and coastal parts of the Barents Sea. Rus. J. Mar. Biol. 2016, 42, 26–35. [Google Scholar] [CrossRef]
- Shirokolobova, T.I.; Zhichkin, A.P.; Venger, M.P.; Vodopyanova, V.V.; Moiseev, D.V. Bacteria and viruses of the ice-free aquatic area of the Barents sea at the beginning of polar night. Dokl. Biol. Sci. 2016, 469, 182–186. [Google Scholar] [CrossRef]
- Sherr, E.B.; Sherr, B.F.; Wheeler, P.A.; Thompson, K. Temporal and spatial variation in stocks of autotrophic and heterotrophic microbes in the upper water column of the central Arctic Ocean. Deep-Sea Res. I 2003, 50, 557–571. [Google Scholar] [CrossRef]
- Maranger, R.; Vaqué, D.; Nguyen, D.; Hébert, M.P.; Lara, E. Pan-Arctic patterns of planktonic heterotrophic microbial abundance and processes: Controlling factors and potential impacts of warming. Prog. Oceanogr. 2015, 139, 221–232. [Google Scholar] [CrossRef]
- Venger, M.P.; Shirikolobova, T.I.; Makarevich, P.R.; Vodopyanova, V.V. Viruses in the pelagic zone of the Barents Sea. Dokl. Biol. Sci. 2012, 446, 345–349. [Google Scholar] [CrossRef]
- Błachowiak-Samołyk, K.; Wiktor, J.M.; Hegseth, E.N.; Wold, A.; Falk-Petersen, S.; Kubiszyn, A.M. Winter Tales: The dark side of planktonic life. Polar Biol. 2015, 38, 23–36. [Google Scholar] [CrossRef]
- Basedow, S.L.; Sundfjord, A.; von Appen, W.J.; Halvorsen, E.; Kwasniewski, S.; Reigstad, M. Seasonal Variation in Transport of Zooplankton into the Arctic Basin through the Atlantic Gateway, Fram Strait. Front. Mar. Sci. 2018, 5, 194. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, P.; Hagen, W.; von Appen, W.J.; Niehoff, B.; Hildebrandt, N.; Auel, H. Effects of a Submesoscale Oceanographic Filament on Zooplankton Dynamics in the Arctic Marginal Ice Zone. Front. Mar. Sci. 2021, 8, 625395. [Google Scholar] [CrossRef]
- Svensen, C.; Seuthe, L.; Vasilyeva, Y.; Pasternak, A.; Hansen, E. Zooplankton distribution across Fram Strait in autumn: Are small copepods and protozooplankton important? Prog. Oceanogr. 2011, 91, 534–544. [Google Scholar] [CrossRef]
- Dvoretsky, V.G.; Dvoretsky, A.G. Summer population structure of the copepods Paraeuchaeta spp. in the Kara Sea. J. Sea Res. 2015, 96, 18–22. [Google Scholar] [CrossRef]
- Ducklow, H.W. Bacterial production and biomass in the oceans. In Microbial Ecology of the Oceans; Kirchman, D.L., Ed.; John Wiley and Sons: Hoboken, NJ, USA, 2000; pp. 85–120. [Google Scholar]
- Kirchman, D.L.; Morán, X.A.G.; Ducklow, H. Microbial growth in the polar oceans—Role of temperature and potential impact of climate change. Nat. Rev. Microbiol. 2009, 7, 451–459. [Google Scholar] [CrossRef]
- Turner, J.T. The importance of small planktonic copepods and their roles in pelagic marine food webs. Zool. Stud. 2004, 43, 255–266. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Date | Region | N | E | Depth, m (Number of Sampling Layers) | Number of Samples (Viruses/ Bacteria/ Chl a/ Zooplankton) |
|---|---|---|---|---|---|---|
| 5 | 24 November 2021 | Fram Strait | 78°40′ | 08°30′ | 700 (7) | 5/7/6/2 |
| 7 | 24 November 2021 | Fram Strait | 79°00′ | 08°30′ | 430 (7) | 5/7/6/2 |
| 9 | 25 November 2021 | Fram Strait | 79°20′ | 08°30′ | 161 (6) | 5/6/8/2 |
| 58 | 16 December 2021 | Southern Barents Sea | 70°60′ | 33°31′ | 220 (6) | 3/5/5/2 |
| 59 | 16 December 2021 | Southern Barents Sea | 70°45′ | 33°30′ | 219 (6) | 4/6/5/2 |
| 60 | 17 December 2021 | Southern Barents Sea | 70°30′ | 33°30′ | 248 (5) | 3/6/5/2 |
| 61 | 17 December 2021 | Southern Barents Sea | 70°15′ | 33°30′ | 248 (5) | 5/6/6/2 |
| 62 | 18 December 2021 | Southern Barents Sea | 70°00′ | 33°29′ | 146 (5) | 3/5/5/2 |
| Fram Strait | Barents Sea | |||
|---|---|---|---|---|
| Abundance | Biomass | Abundance | Biomass | |
| Taxon | 0–50 m/ 50 m–bottom | 0–50 m/ 50 m–bottom | 0–50 m/ 50 m–bottom | 0–50 m/ 50 m–bottom |
| Copepoda | ||||
| Acartia longiremis | 2 ± 0/<1 | <0.1/<0.1 | 44 ± 18/7 ± 3 | 0.4 ± 0.2/0.1 ± 0 |
| Aetideopsis armatus | -/<1 | -/<0.1 | -/- | -/- |
| Bradyidius similis | <1/<1 | <0.1/<0.1 | -/<1 | -/<0.1 |
| Calanus finmarchicus | 20 ± 7/22 ± 10 | 3 ± 1/3 ± 1 | 8 ± 4/162 ± 42 | 1.3 ± 0.8/34 ± 9.2 |
| Calanus glacialis | 38 ± 21/14 ± 4 | 6.8 ± 4/3.7 ± 1.5 | 1 ± 1/17 ± 4 | 0.2 ± 0.1/3.6 ± 1.1 |
| Calanus hyperboreus | 1 ± 0/3 ± 1 | 0.1 ± 0/0.9 ± 0.5 | <1/2 ± 1 | <1/0.3 ± 0.1 |
| Centropages hamatus | -/- | -/- | 2 ± 1/1 ± 0 | <0.1/<0.1 |
| Chiridius obtusifrons | -/<1 | -/<0.1 | -/- | -/- |
| Copepoda nauplii | <1/<1 | <0.1/<0.1 | -/- | -/- |
| Gaetanus tenuispinus | -/1 ± 0 | -/0.2 ± 0.1 | -/- | -/- |
| Gaidius brevispinus | -/<1 | -/<0.1 | -/- | -/- |
| Heterorhabdus norvegicus | -/<1 | -/<0.1 | -/- | -/- |
| Jaschnovia brevis | -/<1 | -/<0.1 | -/- | -/- |
| Metridia longa | 13 ± 3/11 ± 3 | 1.4 ± 0.4/1.1 ± 0.2 | 1 ± 0/46 ± 18 | 0.1 ± 0.1/4.5 ± 1.9 |
| Metridia lucens | 14 ± 3/4 ± 1 | 0.5 ± 0.2/0.1 ± 0 | 129 ± 48/27 ± 11 | 3.5 ± 1.4/0.7 ± 0.4 |
| Microcalanus pusillus | 32 ± 7/7 ± 4 | <0.1/<0.1 | 76 ± 10/32 ± 16 | 0.1 ± 0/<0.1 |
| Microcalanus pygmaeus | 88 ± 8/12 ± 2 | 0.1 ± 0/<0.1 | 51 ± 8/35 ± 7 | 0.1 ± 0/<0.1 |
| Microsetella norvegica | -/- | -/- | <1/- | <0.1/- |
| Oithona atlantica | 10 ± 3/7 ± 4 | <0.1/<0.1 | 3 ± 1/10 ± 4 | <0.1/<0.1 |
| Oithona similis | 364 ± 66/67 ± 33 | 0.4 ± 0.1/0.1 ± 0 | 177 ± 25/122 ± 34 | 0.2 ± 0/0.1 ± 0 |
| Triconia borealis | 1 ± 1/1 ± 0 | <0.1/<0.1 | -/- | -/- |
| Paraeuchaeta spp. I–IV | -/1 ± 0 | -/0.2 ± 0.1 | -/<1 | -/<0.1 |
| Paraeuhaeta glacialis V–VI | <1/<1 | <0.1/0.7 ± 0.2 | -/<1 | -/<0.1 |
| Paraeuhaeta norvegica V–VI | <1/1 ± 0 | <0.1/0.8 ± 0.5 | -/<1 | -/<0.1 |
| Pseudocalanus spp. I–IV | 175 ± 33/33 ± 4 | 0.6 ± 0.1/0.1 ± 0 | 87 ± 8/14 ± 6 | 0.3 ± 0/0.1 ± 0 |
| Pseudocalanus minutus V–VI | 108 ± 23/31 ± 3 | 1 ± 0.2/0.3 ± 0 | 12 ± 2/14 ± 5 | 0.1 ± 0/0.1 ± 0.1 |
| Pseudocalanus acuspes V–VI | 41 ± 7/14 ± 3 | 0.3 ± 0.1/0.1 ± 0 | 8 ± 2/9 ± 6 | 0.1 ± 0/0.1 ± 0 |
| Scolecithricella minor | -/1 ± 0 | -/<0.1 | -/- | -/- |
| Temora longicornis | -/- | -/- | <1/<1 | <0.1/<0.1 |
| Ostracoda | ||||
| Boroecia borealis | <1/1 ± 0 | <0.1/<0.1 | -/- | -/- |
| Discoconchoecia elegans | -/<1 | -/<0.1 | -/- | -/- |
| Boroecia maxima | -/<1 | -/<0.1 | -/- | -/- |
| Amphipoda | ||||
| Themisto abyssorum | -/- | -/- | -/<1 | -/<0.1 |
| Themisto libellula | -/<1 | -/0.1 ± 0.1 | -/- | -/- |
| Euphausiacea | ||||
| Meganyctyphanes norvegica | -/<1 | -/0.4 ± 0.1 | -/<1 | -/0.2 ± 0.1 |
| Thysanoessa inermis | -/<1 | -/0.6 ± 0.2 | -/<1 | -/0.1 ± 0 |
| Thysanoessa longicaudata | <1/<1 | <0.1/0.2 ± 0.1 | <1/<1 | <0.1/<0.1 |
| Nematoscelis megalops | -/<1 | -/<0.1 | -/- | -/- |
| Themisto abyssorum | -/- | -/- | -/<1 | -/<0.1 |
| Themisto libellula | -/<1 | -/0.1 ± 0.1 | -/- | -/- |
| Decapoda | ||||
| Pandalus borealis larvae | -/<1 | -/<0.1 | -/- | -/- |
| Cnidaria | ||||
| Aglantha digitale | <1/1 ± 0 | <0.1/0.2 ± 0 | <1/<1 | <0.1/0.2 ± 0 |
| Dimophyes arctica | <1/<1 | <0.1/<0.1 | -/- | -/- |
| Rathkea octopunctata | <1/- | <0.1/- | -/- | -/- |
| Physophora hydrostatica necthophore | -/<1 | -/<0.1 | <1/<1 | <0.1/<0.1 |
| Pandalus borealis larvae | -/<1 | -/<0.1 | -/- | -/- |
| Ctenophora | ||||
| Beroe cucumis | -/- | -/- | <1/<1 | <0.1/<0.1 |
| Mertensia ovum | <1/<1 | <0.1/0.1 ± 0.1 | <1/<1 | <0.1/<0.1 |
| Pleurobrachia pileus | -/<1 | -/<0.1 | -/- | -/- |
| Polychaeta | ||||
| Pelagobia longicirrata | <1/<1 | <0.1/<0.1 | -/<1 | -/<0.1 |
| Tomopteris helgolandica | -/<1 | -/<0.1 | -/<1 | -/<0.1 |
| Polychaeta larvae | 1 ± 0/- | <0.1/- | -/<1 | -/<0.1 |
| Nemertini | ||||
| Nemertini juv. | -/- | -/- | -/<1 | -/<0.1 |
| Gastropoda | ||||
| Clione limacina larvae | <1/<1 | <0.1/<0.1 | -/- | -/- |
| Clione limacina | <1/<1 | <0.1/0.3 ± 0.1 | -/- | -/- |
| Limacina helicina larvae | 4 ± 1/2 ± 1 | 0.2 ± 0.1/0.1 ± 0.1 | 1 ± 0/1 ± 0 | <0.1/0.1 ± 0 |
| Limacina helicina | 1 ± 0/1 ± 0 | 0.1 ± 0.1/0.1 ± 0 | <1/1 ± 0 | <0.1/0.2 ± 0 |
| Limacina retroversa | 1 ± 0/<1 | 0.1 ± 0/<0.1 | <1/1 ± 0 | <0.1/0.1 ± 0 |
| Gastropoda larvae | -/<1 | -/<0.1 | <1/<1 | <0.1/<0.1 |
| Chaetognatha | ||||
| Eukrohnia hamata | -/<1 | -/0.4 ± 0.2 | -/<1 | -/0.3 ± 0.2 |
| Parasagitta elegans | 2 ± 0/3 ± 2 | 1.3 ± 0.3/1.6 ± 1 | <1/<1 | <0.1/0.8 ± 0.3 |
| Appendicularia | ||||
| Oikopleura juv. | 2 ± 0/<1 | <0.1/<0.1 | <1/- | <0.1/- |
| Oikopleura vanhoeffeni | <1/<1 | <0.1/<0.1 | -/- | -/- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dvoretsky, V.G.; Venger, M.P.; Vashchenko, A.V.; Vodopianova, V.V.; Pastukhov, I.A.; Maksimovskaya, T.M. Marine Plankton during the Polar Night: Environmental Predictors of Spatial Variability. Biology 2023, 12, 368. https://doi.org/10.3390/biology12030368
Dvoretsky VG, Venger MP, Vashchenko AV, Vodopianova VV, Pastukhov IA, Maksimovskaya TM. Marine Plankton during the Polar Night: Environmental Predictors of Spatial Variability. Biology. 2023; 12(3):368. https://doi.org/10.3390/biology12030368
Chicago/Turabian StyleDvoretsky, Vladimir G., Marina P. Venger, Anastasya V. Vashchenko, Veronika V. Vodopianova, Ivan A. Pastukhov, and Tatyana M. Maksimovskaya. 2023. "Marine Plankton during the Polar Night: Environmental Predictors of Spatial Variability" Biology 12, no. 3: 368. https://doi.org/10.3390/biology12030368