
Picks in the Fabric of a Polyploidy Complex: Integrative Species Delimitation in the Tetraploid Leucanthemum Mill. (Compositae, Anthemideae) Representatives

Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Taxon Selection
2.2. Morphological Analyses
2.3. Ecological Niche Modelling
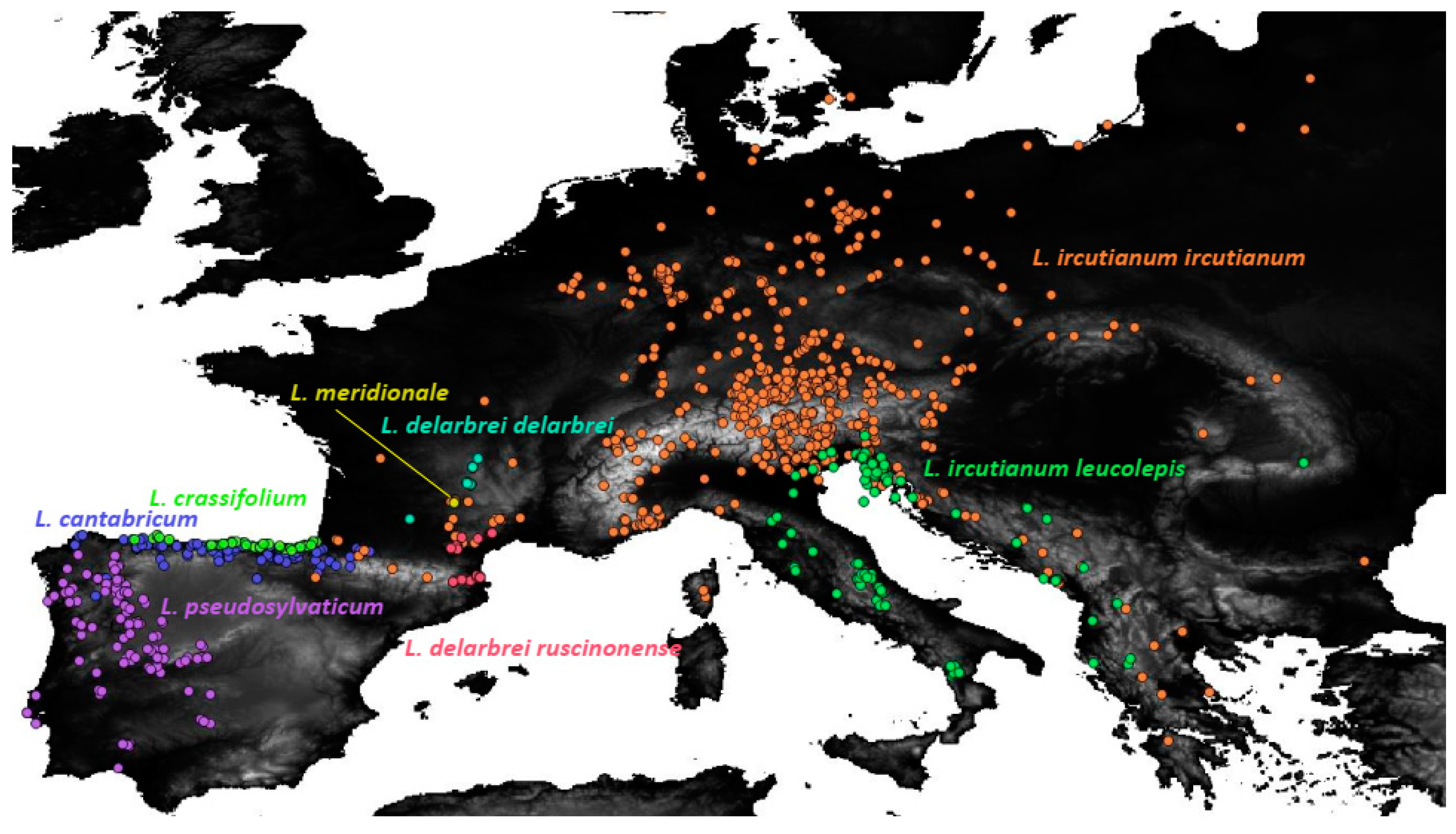
2.4. Geographical Distribution
2.5. RADseq Assembly
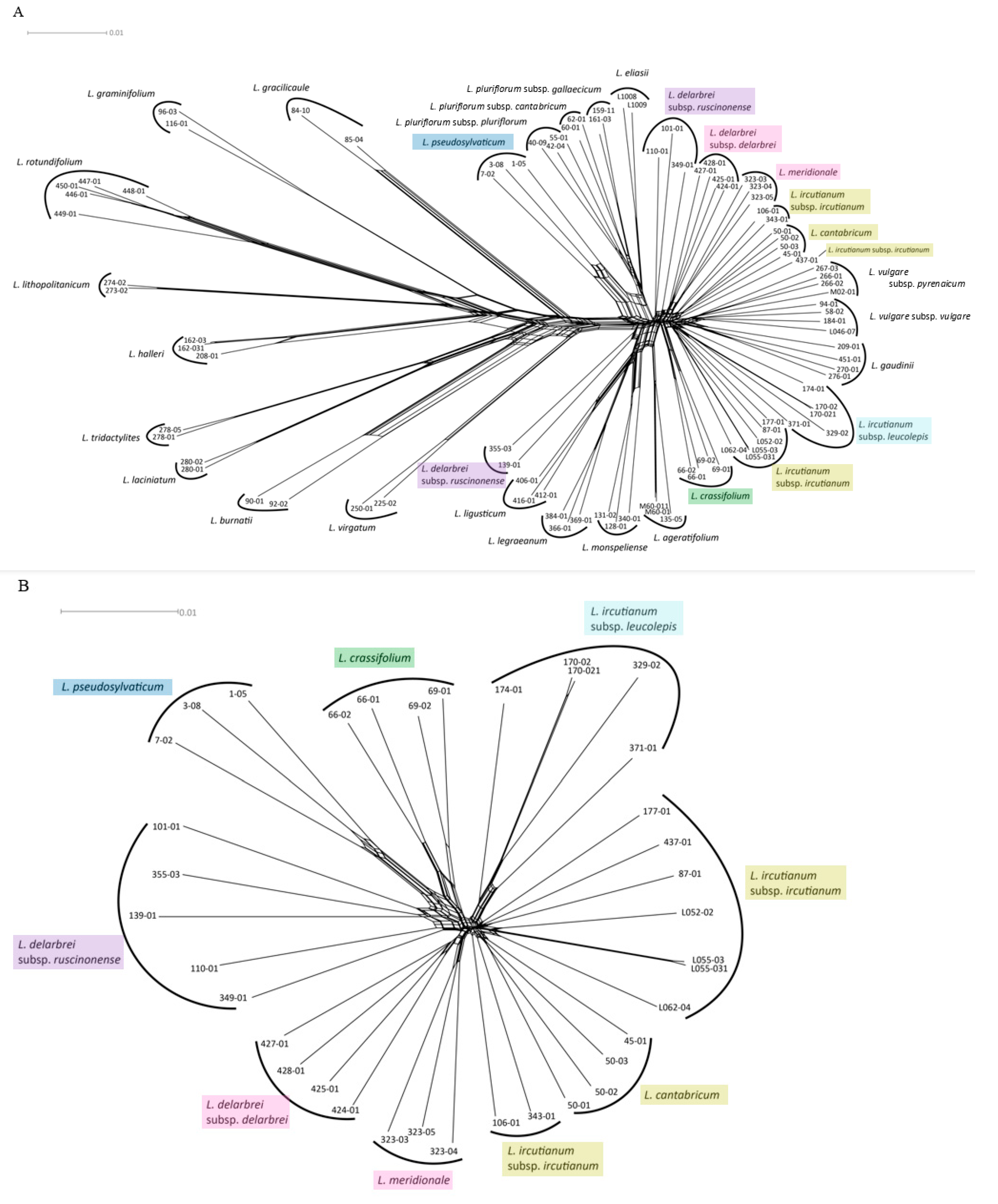
2.6. RADseq Network Analysis
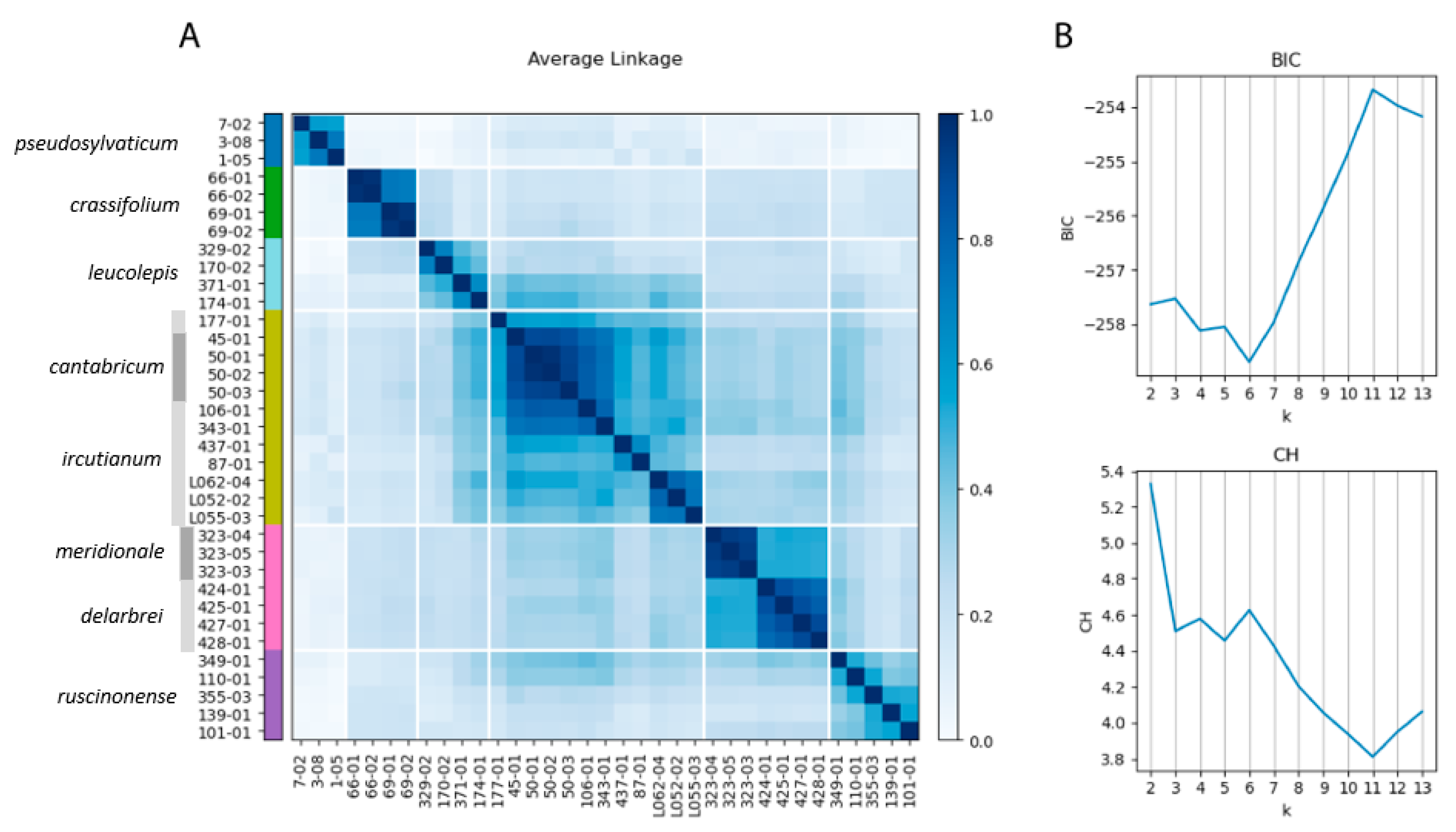
2.7. RADseq Consensus Clustering
2.8. Genealogical Species Delimitation
3. Results
3.1. Morphology
3.2. Ecological Niche Modelling and Geographical Range Overlap
3.3. RADseq Assembly and Analysis
3.4. Genealogical Species Delimitation
4. Discussion
4.1. Genealogical and Genetic Patterns
4.2. Morphological Patterns
4.3. Ecological and Geographical Patterns
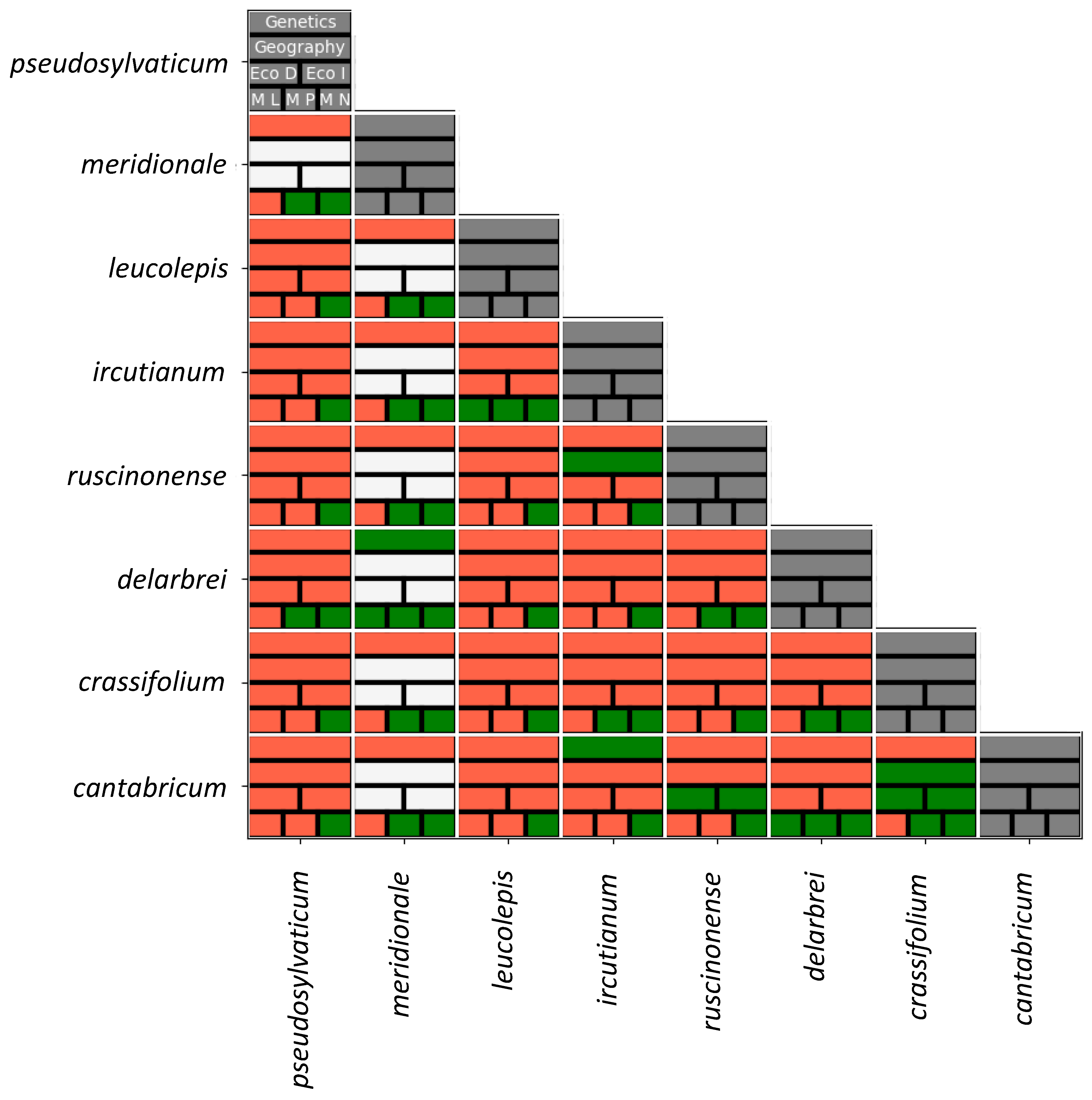
4.4. Integration of Sources of Evidence
5. Taxonomic Treatment
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stuessy, T.F. Ultrastructural data for the practicing plant systematist. Am. Zool. 1979, 19, 621. [Google Scholar] [CrossRef]
- Zachos, F.E.; Christidis, L.; Garnett, S.T. Mammalian species and the twofold nature of taxonomy: A comment on Taylor et al. Mammalia 2020, 84, 1. [Google Scholar] [CrossRef]
- Ence, D.D.; Carstens, B.C. SpedeSTEM: A rapid and accurate method for species delimitation. Mol. Ecol. Resour. 2011, 11, 473. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.L.; Carstens, B.C. Species delimitation using molecular data. In Species Problems and Beyond–Contemporate Issues in Philosophy and Practice; Wilkins, J.S., Zachos, F.E., Pavlinov, I.Y., Eds.; CRC Press: Boca Raton, FL, USA, 2022; pp. 145–159. [Google Scholar]
- De Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879. [Google Scholar] [CrossRef] [PubMed]
- Dayrat, B. Towards integrative taxonomy. Biol. J. Linn. Soc. 2005, 85, 407. [Google Scholar] [CrossRef]
- Will, K.W.; Mishler, B.D.; Wheeler, Q.D. The perils of DNA barcoding and the need for integrative taxonomy. Syst. Biol. 2005, 54, 844. [Google Scholar] [CrossRef]
- Guillot, G.; Renaud, S.; Ledevin, R.; Michaux, J.; Claude, J. A unifying model for the analysis of phenotypic, genetic, and geographical data. Syst. Biol. 2012, 61, 897. [Google Scholar] [CrossRef] [PubMed]
- Zapata, F.; Jiménez, I. Species delimitation: Inferring gaps in morphology across geography. Syst. Biol. 2012, 61, 179. [Google Scholar] [CrossRef]
- Vásquez-Cruz, M.; Vovides, A.P.; Sosa, V. Disentangling species limits in the Vauquelinia corymbosa complex (Pyreae, Rosaceae). Syst. Bot. 2017, 42, 1. [Google Scholar] [CrossRef]
- Solís-Lemus, C.; Knowles, L.L.; Ané, C. Bayesian species delimitation combining multiple genes and traits in a unified framework. Evolution 2015, 69, 492. [Google Scholar] [CrossRef] [Green Version]
- Hausdorf, B.; Hennig, C. Species delimitation and geography. Mol. Ecol. Resour. 2020, 20, 950. [Google Scholar] [CrossRef] [PubMed]
- Doyen, J.T.; Slobodchikoff, C.N. An operational approach to species classification. Syst. Zool. 1974, 23, 239. [Google Scholar] [CrossRef]
- Schlick-Steiner, B.C.; Steiner, F.M.; Seifert, B.; Stauffer, C.; Christian, E.; Crozier, R.H. Integrative taxonomy: A multisource approach to exploring biodiversity. Annu. Rev. Entomol. 2010, 55, 421. [Google Scholar] [CrossRef] [PubMed]
- Padial, J.M.; Miralles, A.; De la Riva, I.; Vences, M. The integrative future of taxonomy. Front. Zool. 2010, 7, 16. [Google Scholar] [CrossRef]
- Oberprieler, C. The Wettstein tesseract: A tool for conceptualising species-rank decisions and illustrating speciation trajectories. Taxon. [CrossRef]
- Sukumaran, J.; Holder, M.T.; Knowles, L.L. Incorporating the speciation process into species delimitation. PLoS Comput. Biol. 2021, 17, e1008924. [Google Scholar] [CrossRef] [PubMed]
- Hörandl, E. Novel approaches for species concepts and delimitation in polyploids and hybrids. Plants 2022, 11, 204. [Google Scholar] [CrossRef]
- Ott, T.; Schall, M.; Vogt, R.; Oberprieler, C. The warps and wefts of a polyploid complex: Integrative species delimitation of the diploid Leucanthemum (Compositae, Anthemideae) representatives. Plants 2022, 11, 1878. [Google Scholar] [CrossRef]
- Euro+Med (2006): Euro+Med PlantBase-the information resource for Euro-Mediterranean plant diversity. Available online: http://ww2.bgbm.org/EuroPlusMed/ (accessed on 24 March 2022).
- Meusel, H.; Jäger, E.J. Vergleichende Chorologie der Zentraleuropäischen Flora; Gustav Fischer Verlag: Jena, Germany; Stuttgart, Germany; New York, NY, USA, 1992; Volume 3. [Google Scholar]
- Vogt, R. Die Gattung Leucanthemum Mill. (Compositae—Anthemideae) auf der Iberischen Halbinsel. Ruizia 1991, 10, 1–261. [Google Scholar]
- Vogt, R. Leucanthemum Mill. In Flora Iberica; Benedí, C., Buira, A., Rico, E., Crespo, M.B., Quintanar, A., Aedo, C., Eds.; Real Jardín Botánico, CSIC: Madrid, Spain, 2019; Volume 16, pp. 1848–1880. [Google Scholar]
- Oberprieler, C.; Greiner, R.; Konowalik, K.; Vogt, R. The reticulate evolutionary history of the polyploid NW Iberian Leucanthemum pluriflorum clan (Compositae, Anthemideae) as inferred from nrDNA ETS sequence diversity and eco-climatological niche-modelling. Mol. Phylogenet. Evol. 2014, 70, 478. [Google Scholar] [CrossRef]
- Konowalik, K.; Wagner, F.; Tomasello, S.; Vogt, R.; Oberprieler, C. Detecting reticulate relationships among diploid Leucanthemum Mill. (Compositae, Anthemideae) taxa using multilocus species tree reconstruction methods and AFLP fingerprinting. Mol. Phylogenet. Evol. 2015, 92, 308. [Google Scholar] [CrossRef] [PubMed]
- Wagner, F.; Härtl, S.; Vogt, R.; Oberprieler, C. ‘Fix Me Another Marguerite!’: Species delimitation in a group of intensively hybridizing lineages of ox-eye daisies (Leucanthemum Mill., Compositae-Anthemideae). Mol. Ecol. 2017, 26, 4260. [Google Scholar] [CrossRef] [PubMed]
- Wagner, F.; Ott, T.; Zimmer, C.; Reichhart, V.; Vogt, R.; Oberprieler, C. ‘At the crossroads towards polyploidy’: Genomic divergence and extent of homoploid hybridization are drivers for the formation of the ox-eye daisy polyploid complex (Leucanthemum, Compositae-Anthemideae). New Phytol. 2019, 223, 2039. [Google Scholar] [CrossRef]
- Oberprieler, C.; Eder, C.; Meister, J.; Vogt, R. AFLP fingerprinting suggests an allopolyploid origin of two members of the Leucanthemum vulgare aggregate (Compositae, Anthemideae) in central Europe. Nord. J. Bot. 2011, 29, 370. [Google Scholar] [CrossRef]
- Greiner, R.; Oberprieler, C. The role of inter-ploidy block for reproductive isolation of the diploid Leucanthemum pluriflorum Pau (Compositae, Anthemideae) and its tetra- and hexaploid relatives. Flora 2012, 207, 629. [Google Scholar] [CrossRef]
- Greiner, R.; Vogt, R.; Oberprieler, C. Evolution of the polyploid north-west Iberian Leucanthemum pluriflorum clan (Compositae, Anthemideae) based on plastid DNA sequence variation and AFLP fingerprinting. Ann. Bot. 2013, 111, 1109. [Google Scholar] [CrossRef]
- Oberprieler, C.; Conti, F.; Dorfner, M.; Eder, S.M.; Heuschneider, A.; Ott, T.; Scheunert, A.; Vogt, R. The taxonomy of Leucanthemum ircutianum (Asteraceae, Anthemideae) in the Apennine Peninsula based on AFLP fingerprinting, plastid DNA sequence variation and eco-climatological niche reconstruction. Bot. J. Linn. Soc. 2022, 194, 830. [Google Scholar] [CrossRef]
- Kuhl, F.P.; Giardina, C.R. Elliptic Fourier features of a closed contour. Comput. Graph. Image Process. 1982, 18, 236. [Google Scholar] [CrossRef]
- Van der Walt, S.; Schönberger, J.L.; Nunez-Iglesias, J.; Boulogne, F.; Warner, J.D.; Yager, N.; Gouillart, E.; Yu, T. scikit-image: Image processing in Python. PeerJ 2014, 2, e453. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array programming with NumPy. Nature 2020, 585, 357. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community ecology package–R package version 2.6-4, 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 31 January 2023).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302. [Google Scholar] [CrossRef]
- Hengl, T.; Jesus, J.M.; de Heuvelink, G.B.M.; Gonzalez, M.R.; Kilibarda, M.; Blagotić, A.; Shangguan, W.; Wright, M.N.; Geng, X.; Bauer-Marschallinger, B.; et al. SoilGrids250m: Global gridded soil information based on machine learning. PLoS ONE 2017, 12, e0169748. [Google Scholar] [CrossRef]
- Hijmans, R. Raster: Geographic Data Analysis and Modeling–R package Version 3.6-11. 2022. Available online: https://CRAN.R-project.org/package=raster (accessed on 31 January 2023).
- Warren, D.L.; Matzke, N.J.; Cardillo, M.; Baumgartner, J.B.; Beaumont, L.J.; Turelli, M.; Glor, R.E.; Huron, N.A.; Simões, M.; Iglesias, T.L.; et al. ENMTools 1.0: An R package for comparative ecological biogeography. Ecography 2021, 44, 504. [Google Scholar] [CrossRef]
- Phillips, S.; Dudík, M.; Schapire, R. Maxent Software for Modeling Species Niches and Distributions, Version 3.4.4. 2021. Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 31 January 2023).
- Poland, J.A.; Brown, P.J.; Sorrells, M.E.; Jannink, J.L. Development of high-density genetic maps for barley and wheat using a novel two-enzyme genotyping-by-sequencing approach. PLoS ONE 2012, 7, e32253. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Dickson, E.E. Preservation of plant samples for DNA restriction endonuclease analysis. Taxon 1987, 36, 715. [Google Scholar] [CrossRef]
- Eaton, D.A.R.; Overcast, I. ipyrad: Interactive assembly and analysis of RADseq datasets. Bioinformatics 2020, 36, 2592. [Google Scholar] [CrossRef]
- Mastretta-Yanes, A.; Arrigo, N.; Alvarez, N.; Jorgensen, T.H.; Piñero, D.; Emerson, B.C. Restriction site-associated DNA sequencing, genotyping error estimation and de novo assembly optimization for population genetic inference. Mol. Ecol. Resour. 2015, 15, 28. [Google Scholar] [CrossRef]
- Joly, S.; Bruneau, A. Incorporating allelic variation for reconstructing the evolutionary history of organisms from multiple genes: An example from Rosa in North America. Syst. Biol. 2006, 55, 623. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; He, S.; Lin, Z.; Yang, F.; Zhou, Q.; Zhou, X. An adaptive robust semi-supervised clustering framework using weighted consensus of random k-means ensemble. IEEE Trans. Knowl. Data Eng. 2019, 33, 1877–1890. [Google Scholar] [CrossRef]
- Peralta, M.; Combes, M.C.; Cenci, A.; Lashermes, P.; Dereeper, A. SNiPloid: A utility to exploit high-throughput SNP data derived from RNA-Seq in allopolyploid species. Int. J. Plant Genom. 2013, 2013, 890123. [Google Scholar] [CrossRef] [PubMed]
- Wagner, N.D.; He, L.; Hörandl, E. Phylogenomic relationships and evolution of polyploid Salix species revealed by RAD sequencing data. Front. Plant Sci. 2020, 11, 1077. [Google Scholar] [CrossRef]
- He, L.; Wagner, N.D.; Hörandl, E. Restriction-site associated DNA sequencing data reveal a radiation of willow species (Salix L., Salicaceae) in the Hengduan Mountains and adjacent areas. J. Syst. Evol. 2021, 59, 44. [Google Scholar] [CrossRef]
- Marcussen, T.; Heier, L.; Brysting, A.K.; Oxelman, B.; Jakobsen, K.S. From gene trees to a dated allopolyploid network: Insights from the angiosperm genus Viola (Violaceae). Syst. Biol. 2015, 64, 84. [Google Scholar] [CrossRef]
- Kamneva, O.K.; Syring, J.; Liston, A.; Rosenberg, N.A. Evaluating allopolyploid origins in strawberries (Fragaria) using haplotypes generated from target capture sequencing. BMC Evol. Biol. 2017, 17, 1. [Google Scholar] [CrossRef]
- Sancho, R.; Inda, L.A.; Dáz-Pérez, A.; Des Marais, D.L.; Gordon, S.; Vogel, J.P.; Lusinska, J.; Hasterok, R.; Contreras-Moreira, B.; Catalán, P. Tracking the ancestry of known and ‘ghost’ homeologous subgenomes in model grass Brachypodium polyploids. Plant J. 2022, 109, 1535. [Google Scholar] [CrossRef]
- Greiner, R.; Vogt, R.; Oberprieler, C. Phylogenetic studies in the polyploid complex of the genus Leucanthemum (Compositae, Anthemideae) based on cpDNA sequence variation. Plant Syst. Evol. 2012, 298, 1407. [Google Scholar] [CrossRef]
- Wagner, F.; Ott, T.; Schall, M.; Lautenschlager, U.; Vogt, R.; Oberprieler, C. Taming the Red Bastards: Hybridisation and species delimitation in the Rhodanthemum arundanum-group (Compositae, Anthemideae). Mol. Phylogenet. Evol. 2020, 144, 106702. [Google Scholar] [CrossRef]
- Monti, S.; Tamayo, P.; Mesirov, J.; Golub, T. Consensus clustering: A resampling-based method for class discovery and visualization of gene expression microarray data. Mach. Learn. 2003, 52, 91. [Google Scholar] [CrossRef]
- Tison, J.M.; de Foucault, B. Flora Gallica: Flore de France; Biotope: Mèze, France, 2014. [Google Scholar]
- Lo Presti, R.M.; Oberprieler, C.; Vogt, R. Tribus VI–Anthemideae (gen. 56, 58-73, 75-79). In Flora d´Italia, 2nd, 3rd ed.; Pignatti, S., Ed.; Edagricole: Milano, Italy, 2018; pp. 815–881. [Google Scholar]
- Seberg, O. Genome analysis, phylogeny, and classification. Plant Syst. Evol. 1989, 166, 159. [Google Scholar] [CrossRef]
- Zachos, F.E. Species Concepts in Biology; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar]
- Stankowski, S.; Ravinet, M. Defining the speciation continuum. Evolution 2021, 75, 1256. [Google Scholar] [CrossRef] [PubMed]
- Von Wettstein, R. Grundzüge der Geographisch-Morphologischen Methode der Pflanzensystematik; Gustav Fischer Verlag: Jena, Germany, 1898. [Google Scholar]
- Wiley, E.O. The evolutionary species concept reconsidered. Syst. Biol. 1978, 27, 17. [Google Scholar] [CrossRef]
- Sukumaran, J.; Knowles, L.L. Multispecies coalsecent delimits structure, not species. Proc. Natl. Acad. Sci. USA 2017, 114, 1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, J.M. Pugillus plantarum imrimis hispanicarum quas in itinere 1851-52 legit Joh. Lange, Vidensk. Meddel. Dan. Naturhist. Kjöbenhaven 1861, 3, 33–116. [Google Scholar]
- Sennen, F. Diagnoses des Nouveautés Parues dans les Exsiccata Plantes d´Espagne et du Maroc de 1928 à Imp; Anglada: Vic, Spain, 1936. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| cantabricum | crassifolium | delarbrei | ruscinonense | ircutianum | leucolepis | meridionale | pseudosylvaticum | |
|---|---|---|---|---|---|---|---|---|
| cantabricum | <0.01 | 0.21 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| crassifolium | 1.0/1.0 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | |
| delarbrei | 1.0/1.0 | 0.44/1.0 | <0.01 | <0.01 | <0.01 | 1.0 | <0.01 | |
| ruscinonense | 0.04/<0.01 | 0.02/<0.01 | 1.0/1.0 | <0.01 | <0.01 | <0.01 | <0.01 | |
| ircutianum | <0.01/<0.01 | 0.17/<0.01 | 0.02/0.83 | <0.01/<0.01 | 1.0 | <0.01 | <0.01 | |
| leucolepis | <0.01/<0.01 | <0.01/<0.01 | <0.01/<0.01 | <0.01/<0.01 | 1.0/<0.01 | <0.01 | <0.01 | |
| meridionale | 1.0/1.0 | 1.0/1.0 | 1.0/0.83 | 1.0/1.0 | 1.0/<0.01 | 0.38/<0.01 | <0.01 | |
| pseudosylvaticum | <0.01/<0.01 | <0.01/<0.01 | 0.06/<0.01 | 0.01/<0.01 | <0.01/<0.01 | <0.01/<0.01 | 1.0/1.0 |
| cantabricum | crassifolium | delarbrei | ruscinonense | ircutianum | leucolepis | meridionale | pseudosylvaticum | |
|---|---|---|---|---|---|---|---|---|
| cantabricum | 0.52 | 0 | 0.32 | 0 | 0 | n/a | 0 | |
| crassifolium | 0.31 | 0 | 0 | 0 | 0 | n/a | 0 | |
| delarbrei | 0 | 0 | 0 | 0 | 0 | n/a | 0 | |
| ruscinonense | 0.52 | 0 | 0 | 0 | 0 | n/a | 0 | |
| ircutianum | 0 | 0 | 0 | 0 | 0 | n/a | 0 | |
| leucolepis | 0 | 0 | 0 | 0 | 0 | n/a | 0 | |
| meridionale | n/a | n/a | n/a | n/a | n/a | n/a | n/a | |
| pseudosylvaticum | 0 | 0 | 0 | 0 | 0 | 0 | n/a |
| cantabricum | crassifolium | delarbrei | ruscinonense | ircutianum | leucolepis | meridionale | pseudosylvaticum | |
|---|---|---|---|---|---|---|---|---|
| cantabricum | 1.0 | 0 | 0 | 0 | 0 | n/a | 0 | |
| crassifolium | 0 | 0 | 0 | 0 | n/a | 0 | ||
| delarbrei | 0 | 0 | 0 | n/a | 0 | |||
| ruscinonense | 0.05 | 0 | n/a | 0 | ||||
| ircutianum | 0 | n/a | 0 | |||||
| leucolepis | n/a | 0 | ||||||
| meridionale | n/a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oberprieler, C.; Ott, T.; Vogt, R. Picks in the Fabric of a Polyploidy Complex: Integrative Species Delimitation in the Tetraploid Leucanthemum Mill. (Compositae, Anthemideae) Representatives. Biology 2023, 12, 288. https://doi.org/10.3390/biology12020288
Oberprieler C, Ott T, Vogt R. Picks in the Fabric of a Polyploidy Complex: Integrative Species Delimitation in the Tetraploid Leucanthemum Mill. (Compositae, Anthemideae) Representatives. Biology. 2023; 12(2):288. https://doi.org/10.3390/biology12020288
Chicago/Turabian StyleOberprieler, Christoph, Tankred Ott, and Robert Vogt. 2023. "Picks in the Fabric of a Polyploidy Complex: Integrative Species Delimitation in the Tetraploid Leucanthemum Mill. (Compositae, Anthemideae) Representatives" Biology 12, no. 2: 288. https://doi.org/10.3390/biology12020288