
Established and Emerging Research Trends in Norway Lobster, Nephrops norvegicus

 , , , , , , , and
, , , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
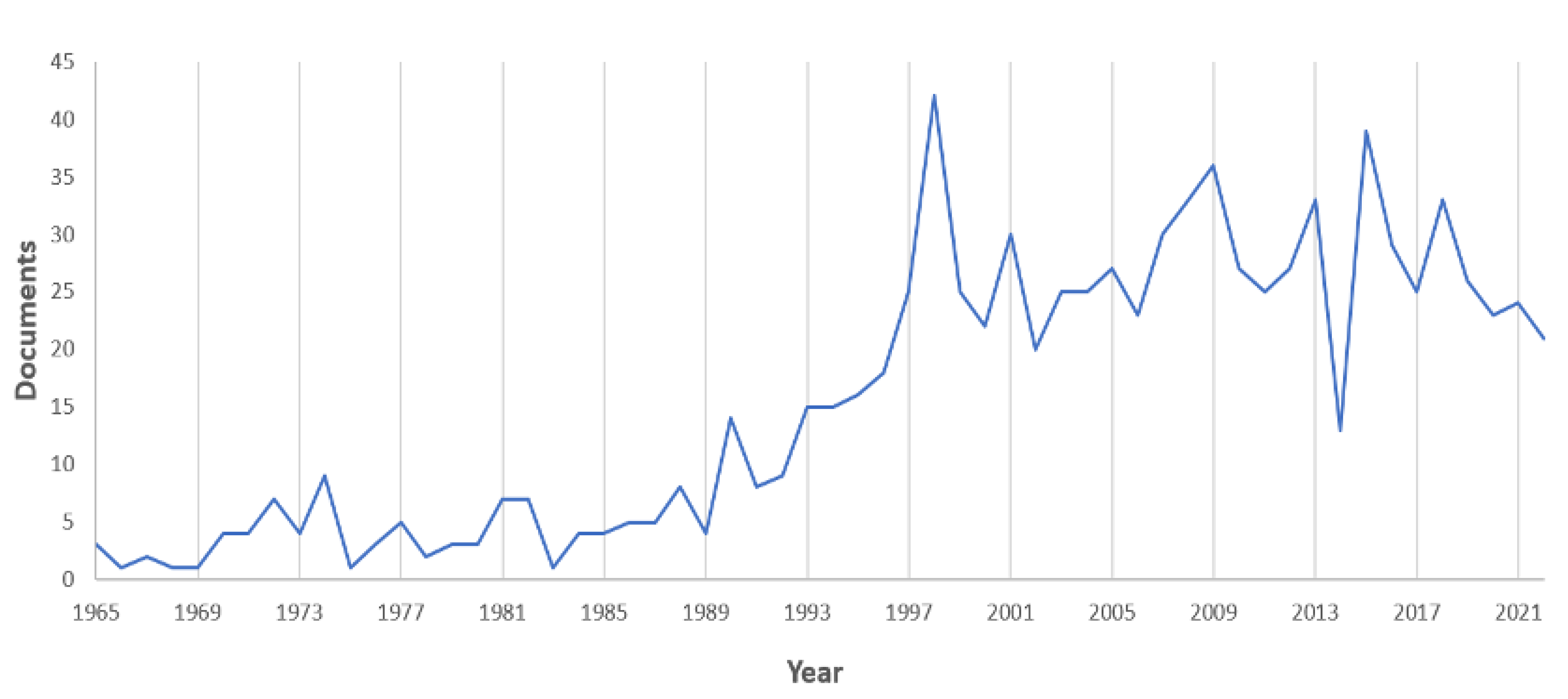
3. Results
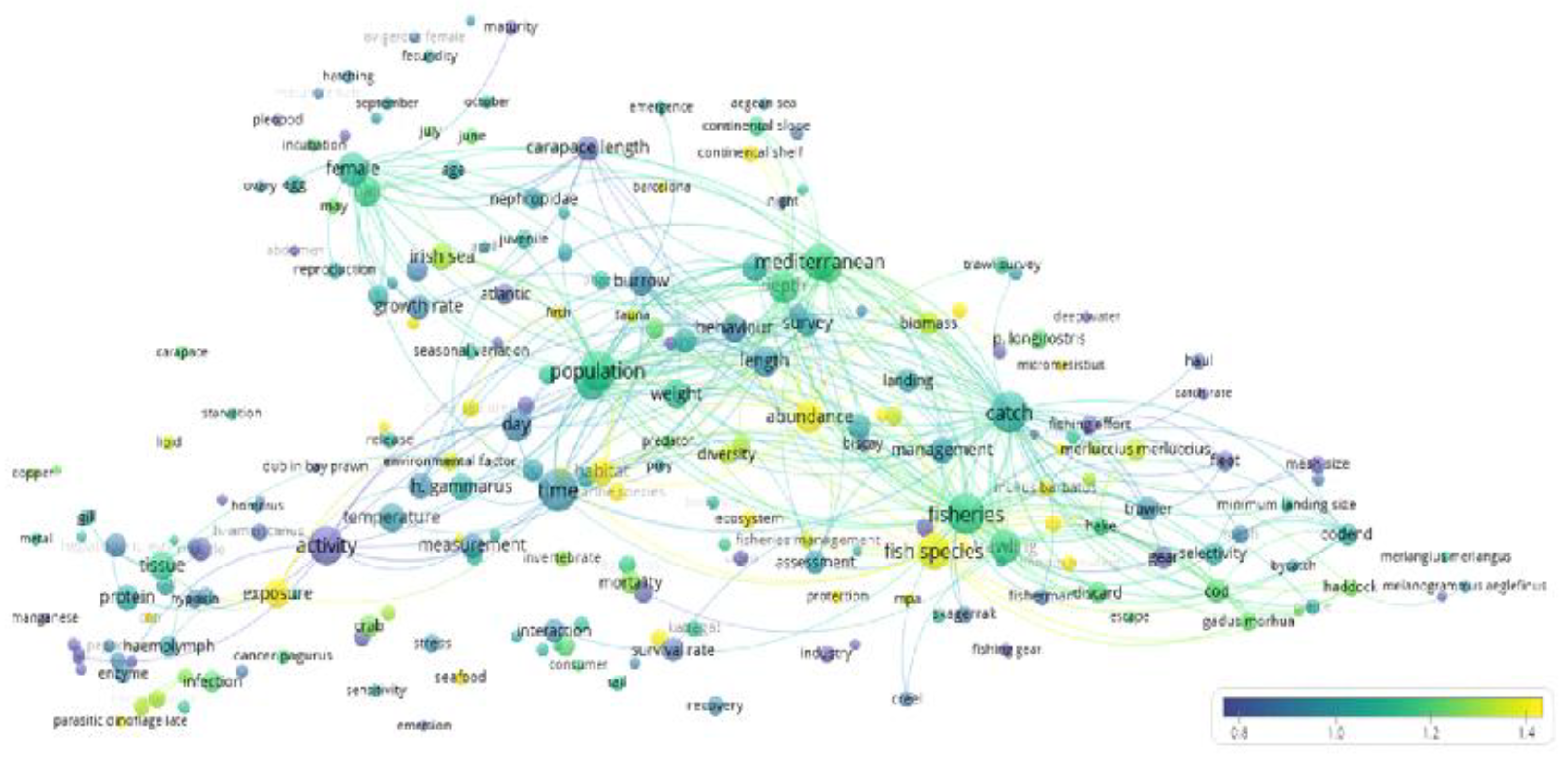
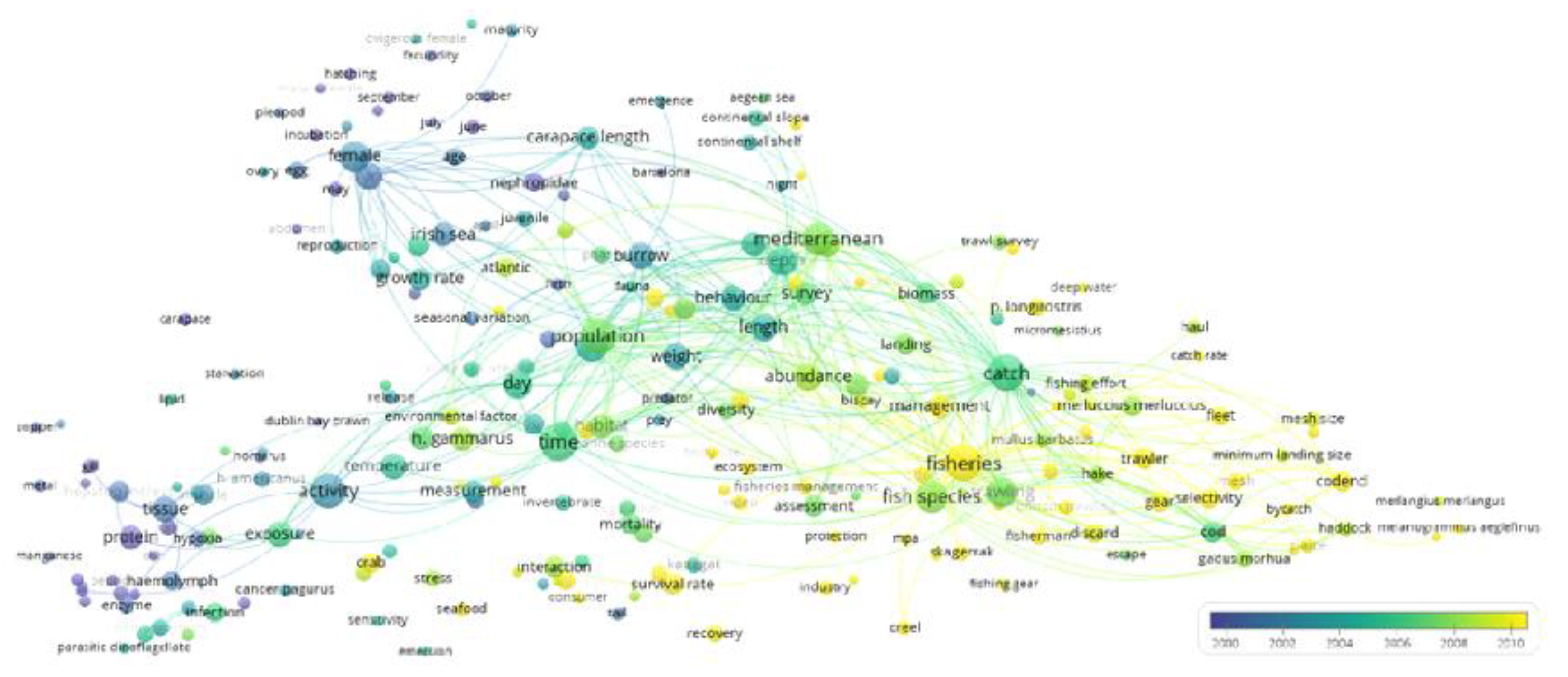
The Areas of Biological and Ecological Research
4. Discussion
4.1. The Research Status
4.2. A Conservation-Oriented Fishery Research
4.3. A Traditional Stock Assessment with No New Technological Components
4.4. The Status of Physiological Research
4.5. The Food Industry as Emerging Research Involving Ecotoxicology
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aguzzi, J.; Maynou, F.; Rotllant, G.; Sardà, F. The dimensional units of Nephrops norvegicus (L.) distribution: From burrows to populations. Int. J. Integr. Biogeogr. 2004, 25, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.; Tuck, I.; Dobby, H. Nephrops species. In Lobsters: Biology, Management, Aquaculture and Fisheries; Wiley: Hoboken, NJ, USA, 2013; pp. 357–413. [Google Scholar] [CrossRef]
- Farmer, A.S.D. Field assessments of diurnal activity in Irish Sea populations of the Norway Lobster Nephrops norvegicus (L.) (Decapoda: Nephropidae). Estuar. Coast. Mar. Sci. 1974, 2, 37–47. [Google Scholar] [CrossRef]
- Bailey, N.; Chapman, C.J.; Alfonso-Dias, M.; Turrell, W. The influence of hydrographic factors on Nephrops distribution and biology. ICES CM 1995, 17, 13. [Google Scholar]
- Tully, O.; Hillis, J.P. Causes and spatial scales of variability in population structure of Nephrops norvegicus (L.) in the Irish Sea. Fish. Res. 1995, 21, 329–347. [Google Scholar] [CrossRef]
- Maynou, F.X.; Sardà, F. Nephrops norvegicus population and morphometrical characteristic in relation to substrate heterogeneity. Fish. Res. 1997, 30, 139–149. [Google Scholar] [CrossRef]
- Froglia, C.; Atkinson, R.J.; Tuck, I.; Arneri, E. Underwater television survey, a tool to estimate Nephrops stock biomass on the Adriatic trawling grounds. In Tisucu Godina Prvoga Spomena Ribarstva u Hrvata; Hrvatska Akademija Znanosti i Umjetnosti: Zagreb, Croatia, 1997; pp. 657–667. [Google Scholar]
- Hugens, D.J.; Atkinson, R.J.A. A towed video survey of megafaunal bioturbation in the North-eastern Irish Sea. J. Mar. Biol. Assoc. 1997, 77, 635–653. [Google Scholar] [CrossRef]
- Tuck, I.D.; Atkinson, R.J.A.; Chapman, C.J. The structure and seasonal variability in the spatial distribution of Nephrops norvegicus burrows. Ophelia 1994, 40, 13–25. [Google Scholar] [CrossRef]
- Tuck, I.D.; Chapman, C.J.; Atkinson, R.J.A. Population biology of the Norway lobster, Nephrops norvegicus (L.) in the Firth of Clyde, Scotland-I: Growth and density. ICES J. Mar. Sci. 1997, 54, 125–135. [Google Scholar] [CrossRef]
- Issifu, I.; Alava, J.J.; Lam, V.W.; Sumaila, U.R. Impact of Ocean Warming, Overfishing and Mercury on European Fisheries: A Risk Assessment and Policy Solution Framework. Front. Mar. Sci. 2022, 8, 770805. [Google Scholar] [CrossRef]
- STECF. Monitoring the Performance of the Common Fisheries Policy (STECF-Adhoc-20-01); Publications Office of the European Union: Luxembourg, 2020; ISBN 978-92-76-18115-6. [Google Scholar] [CrossRef]
- Roberge, J.M.; Angelstam, P. Usefulness of the umbrella species concept as a conservation tool. Conserv. Biol. 2004, 18, 76–85. [Google Scholar] [CrossRef]
- Aguzzi, J.; Bahamon, N.; Marotta, L. The influence of light availability and predatory behavior of the decapod crustacean Nephrops norvegicus on the activity rhythms of continental margin prey decapods. Mar. Ecol. 2009, 30, 366–375. [Google Scholar] [CrossRef]
- Bell, M.C.; Redant, F.; Tuck, I. Nephrops species. In Lobsters: Biology, Management, Aquaculture and Fisheries; Phillips, B.F., Ed.; Blackwell Publishing: Oxford, UK, 2006; pp. 412–461. [Google Scholar] [CrossRef]
- Aguzzi, J.; Sardà, F. A history of recent advancements on Nephrops norvegicus behavioral and physiological rhythms. Rev. Fish Biol. Fish. 2008, 18, 235–248. [Google Scholar] [CrossRef]
- Katho, E.; Sbragaglia, V.; Aguzzi, J.; Breithaupt, T. Sensory biology and behaviour of Nephrops norvegicus. Adv. Mar. Biol. 2013, 64, 66–106. [Google Scholar] [CrossRef]
- Aguzzi, J.; Bahamon, N.; Doyle, J.; Lordan, C.; Tuck, I.D.; Chiarini, M.; Martinelli, M. Burrow emergence rhythms of Nephrops norvegicus by UWTV and surveying biases. Sci. Rep. 2021, 11, 5797. [Google Scholar] [CrossRef] [PubMed]
- Aguzzi, J.; Chatzievangelou, D.; Robinson, N.J.; Bahamon, N.; Berry, A.; Carreras, M.; Company, J.B.; Costa, C.; Del Río, J.; Falahzadeh, A.; et al. Advancing fishery-independent stock assessments for the Norway lobster (Nephrops norvegicus) with new monitoring technologies. Front. Mar. Sci. 2022, 9, 969071. [Google Scholar] [CrossRef]
- Main, J.; Sangster, G.I. The Behaviour of the Norway Lobster Nephrops norvegicus (L) During Trawling; Department of Agriculture and Fisheries for Scotland: Aberdeen, UK, 1985; Volume 34, pp. 1–23. [Google Scholar]
- Aguzzi, J.; Abelló, P.; García, J.A. Daily activity patterns of the mud shrimp Solenocera membranacea (Decapoda: Penaeoidea: Solenoceridae) in the western Mediterranean: A Comparison by Depth and Season. Bull. Mar. Sci. 2006, 79, 353–364. [Google Scholar]
- Sardà, F.; Aguzzi, J. A review of burrow counting as an alternative to other typical methods of assessment of Norway lobster populations. Rev. Fish Biol. Fish. 2012, 22, 409–422. [Google Scholar] [CrossRef]
- Chapman, C.J.; Johnstone, A.D.F.; Rice, A.L. The behaviour and ecology of the Norway lobster Nephrops norvegicus (L). In Proceedings of the 9th European Marine Biology Symposium, Oban, UK, 2–8 October 1974; Barnes, H.B., Ed.; Aberdeen University Press: Aberdeen, UK, 1975; pp. 59–74. [Google Scholar]
- Moller, T.H.; Naylor, E. Environmental influence on locomotor activity in Nephrops norvegicus (Crustacea:Decapoda). J. Mar. Biol. Assoc. UK 1980, 60, 103–113. [Google Scholar] [CrossRef]
- Aguzzi, J.; Sardà, F.; Abelló, P.; Company, J.B.; Rotllant, G. Diel and seasonal patterns of Nephrops norvegicus (Decapoda: Nephropidae) catchability in the western Mediterranean. Mar. Ecol. Prog. Ser. 2003, 258, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Aguzzi, J.; Sardà, F.; Allué, R. Seasonal dynamics in Nephrops norvegicus (Decapoda: Nephropidae) catches off the Catalan Coasts (Western Mediterranean). Fish. Res. 2004, 69, 293–300. [Google Scholar] [CrossRef]
- Smyth, K.; Elliott, M. Effects of changing salinity on the ecology of the marine environment. In Stressors in the Marine Environment: Physiological and Ecological Responses; Societal Implications; Solan, M., Whiteley, N., Eds.; Oxford University Press: Oxford, UK, 2016; pp. 161–174. [Google Scholar] [CrossRef]
- McGeady, R.; Lordan, C.; Power, A.M. Shift in the larval phenology of a marine ectotherm due to ocean warming with consequences for larval transport. Limnol. Oceanogr. 2021, 66, 543–557. [Google Scholar] [CrossRef]
- Ridgway, I. Post Capture Physiology and Pathology of the Norway Lobster, Nephrops norvegicus. PhD Thesis, University of Glasgow, Glasgow, UK, 2005. [Google Scholar]
- Ridgway, I.D.; Small, H.J.; Atkinson, R.J.A.; Birkbeck, H.T.; Taylor, A.C.; Neil, D.M. Extracellular proteases and possible disease related virulence mechanisms of two marine bacteria implicated in an opportunistic bacterial infection of Nephrops norvegicus. J. Invertebr. Pathol. 2008, 99, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Stentiford, G.D.; Neil, D.M.; Atkinson, R.J.A. Infection by the dinoflagellate Hematodinium in the Norway lobster (Nephrops norvegicus L.) on the west coast of Scotland, United Kingdom. In Proceedings of the 1999 Annual Meeting of the National Shellfisheries Association, Halifax, NS, Canada, 18–22 April 1999. [Google Scholar]
- Stentiford, G.D.; Neil, D.M. Diseases of Nephrops and Metanephrops: A review. J. Invertebr. Pathol. 2011, 106, 92–109. [Google Scholar] [CrossRef] [PubMed]
- Stentiford, G.D.; Douglas, N.; Atkinson, R.J.A. Alteration of burrow-related behaviour of the Norway Lobster Nephrops norvegicus during infection by the parasitic dinoflagellate Hematodinium. Mar. Fresh. Behav. Physiol. 2001, 34, 139–156. [Google Scholar] [CrossRef]
- Van Eck, N.; Waltman, L. Software survey: VOSviewer, a computer program for bibliometric mapping. Scientometrics 2010, 84, 523–538. [Google Scholar] [CrossRef] [Green Version]
- Company, J.B.; Recasens, L.; García del Arco, J.A.; Bahamon, N.; Colmenero, A.I. Scenarios for the implementation of management measures reported. In Article 11.3. Management Practices for Bottom Trawling Fisheries (OTB) on Spanish Waters of the Western Mediterranean Sea (GSA6); Progress Technical Report; Catalan Institute for Ocean Governance Research (ICATMAR): Girona, Spain, 2020. [Google Scholar]
- FAO. The State of Mediterranean and Black Sea Fisheries 2020; General Fisheries Commission for the Mediterranean: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Costa, C.; Fanelli, E.; Marini, S.; Danovaro, R.; Aguzzi, J. Global deep-sea biodiversity research trends highlighted by science mapping approach. Front. Mar. Sci. 2020, 7, 384. [Google Scholar] [CrossRef]
- Mongeon, P.; Paul-Hus, A. The journal coverage of Web of Science and Scopus: A comparative analysis. Scientometrics 2016, 106, 213–228. [Google Scholar] [CrossRef]
- Ioannoni, V.; Vitale, T.; Costa, C.; Elliot, I. Depicting communities of Romani studies: On the who, when and where of Roma related scientific publications. Scientometrics 2020, 122, 1473–1490. [Google Scholar] [CrossRef] [Green Version]
- Manfredini, A.; Malusà, E.; Costa, C.; Pallottino, F.; Mocali, S.; Pinzari, F.; Canfora, L. Current methods, common practices, and perspectives in tracking and monitoring bioinoculants in soil. Front. Microbiol. 2021, 12, 698491. [Google Scholar] [CrossRef] [PubMed]
- Waltman, L.; Van Eck, N.J.; Noyons, E.C.M. A unified approach to mapping and clustering of bibliometric networks. J. Informetr. 2010, 4, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Waltman, L.; Van Eck, N.J. A smart local moving algorithm for large-scale modularity-based community detection. Eur. Phys. J. B 2013, 86, 471. [Google Scholar] [CrossRef]
- Hassan, S.U.; Visvizi, A.; Waheed, H. The ‘who’and the ‘what’ in international migration research: Data-driven analysis of Scopus-indexed scientific literature. Behav. Inf. Technol. 2019, 38, 924–939. [Google Scholar] [CrossRef]
- Van Eck, N.J.; Waltman, L.; Van Raan, A.F.; Klautz, R.J.; Peul, W.C. Citation analysis may severely underestimate the impact of clinical research as compared to basic research. PLoS ONE 2013, 8, e62395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, P.; Sabates, A. Spatio-temporal distribution pattern of the red band-fish Cepola rubescens Linnaeus at different stages of its life cycle in the northwestern Mediterranean. J. Fish Biol. 1991, 39, 549–557. [Google Scholar] [CrossRef]
- EU. Regulation (EU) 2017/1004 of the European Parliament and of the Council of 17 May 2017 on the establishment of a Union framework for the collection, management and use of data in the fisheries sector and support for scientific advice regarding the common fisheries policy and repealing Council Regulation (EC) No 199/2008 (recast). Off. J. Eur. Union 2017, 157, 1–21. [Google Scholar]
- ICES. Working Group on Nephrops Surveys (WGNEPS; outputs from 2021). ICES Sci. Rep. 2022, 4, 183. [Google Scholar]
- Sbrana, M.; Zupa, W.; Ligas, A.; Capezzuto, F.; Chatzispyrou, A.; Follesa, M.C.; Gancitano, V.; Guijarro, B.; Isajlovic, I.; Jadaud, A.; et al. Spatiotemporal abundance pattern of deep-water rose shrimp, Parapenaeus longirostris, and Norway lobster, Nephrops norvegicus, in European Mediterranean waters. Sci. Mar. 2019, 83, 71–80. [Google Scholar] [CrossRef] [Green Version]
- GFCM. Recommendation GFCM/44/2021/2 on the Establishment of a Fisheries Restricted Area in the Jabuka/Pomo Pit in the Adriatic Sea (Geographical Subarea 17), Amending Recommendation; GFCM: Roma, Italy, 2021; Available online: https://gfcm.sharepoint.com/:b:/g/CoC/EWi2Yh6-KaFDggZvMW58ddwBlUwcYFZobua33Dil-Z8CKA (accessed on 25 November 2022).
- Chiarini, M.; Guicciardi, S.; Zacchetti, L.; Domenichetti, F.; Canduci, G.; Angelini, S.; Belardinelli, A.; Croci, C.; Giuliani, G.; Scarpini, P.; et al. Looking for a Simple Assessment Tool for a Complex Task: Short-Term Evaluation of Changes in Fisheries Management Measures in the Pomo/Jabuka Pits Area (Central Adriatic Sea). Sustainability 2022, 14, 7742. [Google Scholar] [CrossRef]
- Roberts, C.M.; Hawkins, J.P. Establishment of fish stock recovery areas. In Report for European Parliament’s Committee on Fisheries; European Parliament: Strasbourg, France, 2012. [Google Scholar]
- DEFRA (Department for Environment, Food and Rural Affairs). Establishing a Multi-Annual Plan for Demersal Stocks and Nephrops in the North Sea and the Fisheries Exploiting Those Stocks. Summary of Responses and Defra’s Response; DEFRA: London, UK, 2017. [Google Scholar]
- Ungfors, A.; Bell, E.; Johson, M.L.; Cowing, D.; Dobson, N.C.; Bublitz, R.; Sandell, J. Habitat and ecology of Nephrops norvegicus. Adv. Mar. Biol. 2013, 64, 248–306. [Google Scholar]
- Letschert, J.; Stollberg, N.; Rambo, H.; Kempf, A.; Berkenhagen, J.; Stelzenmüller, V. The uncertain future of the Norway lobster fisheries in the North Sea calls for new management strategies. ICES J. Mar. Sci 2021, 78, 3639–3649. [Google Scholar] [CrossRef]
- FAO-GFCM. Fishery and Aquaculture Statistics. GFCM Capture Production 1970-2020 (FishStatJ). In FAO Fisheries and Aquaculture Division. Rome. Updated 2022. Available online: www.fao.org/fishery/statistics/software/fishstatj/en (accessed on 11 November 2022).
- Worm, B.; Hilborn, R.; Baum, J.K.; Branch, T.A.; Collie, J.S.; Costello, C.; Fogarty, M.J.; Fulton, E.A.; Hutchings, J.A.; Jennings, S.; et al. Rebuilding Global Fisheries. Science 2009, 325, 578–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- STECF. Stock Assessments: Demersal Stocks in the Western Mediterranean Sea; Scientific, Technical and Economic Committee for Fisheries (STECF-22-09); Publications Office of the European Union: Luxembourg, 2022. [Google Scholar]
- Bahamon, N.; Sarda, F.; Aguzzi, J. Fuzzy diel patterns in catchability of deep-water species on the continental margin. ICES J. Mar. Sci. 2009, 66, 2211–2218. [Google Scholar] [CrossRef] [Green Version]
- McCauley, D.J.; DeSalles, P.A.; Young, H.S.; Gardner, J.P.A.; Micheli, F. Use of high-resolution acoustic cameras to study reef shark behavioral ecology. J. Exp. Mar. Biol. Ecol. 2016, 482, 128–133. [Google Scholar] [CrossRef] [Green Version]
- Marini, S.; Bonofiglio, F.; Corgnati, L.P.; Bordone, A.; Schiaparelli, S.; Peirano, A. Long-term automated visual monitoring of Antarctic benthic fauna. Methods Ecol. Evol. 2022, 13, 1746–1764. [Google Scholar] [CrossRef]
- Aguzzi, J.; Navarro, J.; Bahamon, N.; Rotllant, G.; García, J.A.; Río, J.D.; Gomàriz, S.; Masmitja, I.; Fanelli, E.; Marini, S.; et al. New monitoring technologies to produce ancillary data on Nephrops stock assessment. Report of the Working Group on Nephrops Surveys. ICES WGNEPS Rep. 2019, 5, 90–91. [Google Scholar]
- Masmitja, I.; Navarro, J.; Gomaríz, S.; Aguzzi, J.; Kieft, B.; O’Reilly, T.; Katija, K.; Bouvet, P.J.; Fannjiang, C.; Vigo, C.; et al. Mobile robotic platforms for the acoustic tracking of deep-sea demersal fishery resources. Sci. Robot. 2020, 5, eabc3701. [Google Scholar] [CrossRef] [PubMed]
- Rountree, R.A.; Juanes, F.; Bolgan, M. Temperate freshwater soundscapes: A cacophony of undescribed biological sounds now threatened by anthropogenic noise. PLoS ONE 2020, 15, e0221842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuck, I.D.; Parsons, D.M.; Hartill, B.W.; Chiswell, S.M. Scampi (Metanephrops challengeri) emergence patterns and catchability. ICES J. Mar. Sci. 2015, 72 (Suppl. S1), i199–i210. [Google Scholar] [CrossRef] [Green Version]
- Vigo, M.; Navarro, J.; Giménez, J.; Andón, N.; Martínez-Lage, A.; Company, J.B.; Rotllant, G. Using molecular and stable isotope markers to identify the main predators of Nephrops norvegicus in Mediterranean deep-water ecosystems. Mar. Ecol. Prog. Ser. 2022, 695, 95–108. [Google Scholar] [CrossRef]
- Florko, K.R.N.; Davidson, E.R.; Lees, K.J.; Hammer, L.J.; Lavoie, M.F.; Lennox, R.J.; Simard, É.; Archambault, P.; Auger-Méthé, M.; McKindsey, C.W.; et al. Tracking movements of decapod crustaceans: A review of a half-century of telemetry-based studies. Mar. Ecol. Prog. Ser. 2021, 679, 219–239. [Google Scholar] [CrossRef]
- Matley, J.K.; Klinard, N.V.; Barbosa Martins, A.P.; Aarestrup, K.; Aspillaga, E.; Cooke, S.J.; Cowley, P.D.; Heupel, M.R.; Lowe, C.G.; Lowerre-Barbieri, S.K.; et al. Global trends in aquatic animal tracking with acoustic telemetry. Trends Ecol. Evol. 2022, 37, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Smullen, R.P.; Bentley, M.G. Studies on crustacean hyperglycaemic hormone of the Norway lobster Nephrops norvegicus (L.). Invertebr. Reprod. Dev. 1994, 26, 23–32. [Google Scholar] [CrossRef]
- Edomi, P.; Azzoni, E.; Mettulio, R.; Pandolfelli, N.; Ferrero, E.A.; Giulianini, P.G. Gonad-inhibiting hormone of the Norway lobster (Nephrops norvegicus): cDNA cloning, expression, recombinant protein production, and immunolocalization. Gene 2002, 284, 93–102. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Rotllant, G.E.; Cummins, S.F.; Elizur, A.; Ventura, T. Insights Into Sexual Maturation and Reproduction in the Norway Lobster (Nephrops norvegicus) via in Silico Prediction and Characterization of Neuropeptides and G Protein-coupled Receptors. Front. Endocrinol. 2018, 9, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loew, E.R. Light, and photoreceptor degeneration in the Norway lobster, Nephrops norvegicus (L.). Proc. R. Soc. B 1976, 193, 31–44. [Google Scholar] [CrossRef]
- Shelton, P.M.J.; Gaten, E.; Chapman, C.J. Light and retinal damage in Nephrops norvegicus (L.) (Crustacea). Proc. R. Soc. B 1985, 226, 217–236. [Google Scholar] [CrossRef]
- Gaten, E. Light-induced damage to the dioptric apparatus of Nephrops norvegicus (L.) and the quantitative assessment of the damage. Mar. Freshw. Behav. Physiol. 1988, 13, 169–183. [Google Scholar] [CrossRef]
- Chapman, C.J.; Shelton, P.M.J.; Shanks, A.M.; Gaten, E. Survival and growth of the Norway lobster Nephrops norvegicus in relation to light-induced eye damage. Mar. Biol. 2000, 136, 233–241. [Google Scholar] [CrossRef]
- Gaten, E.; Moss, S.; Johnson, M.L. Chapter Four—The Reniform Reflecting Superposition Compound Eyes of Nephrops norvegicus: Optics, Susceptibility to Light-Induced Damage, Electrophysiology and a Ray Tracing Model. Adv. Mar. Biol. 2013, 64, 107–148. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.S.; Brown, J.N.; Jourdain, N.C.; van Sebille, E.; Ganachaud, A.; Vergés, A. Episodic and non-uniform shifts of thermal habitats in a warming ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 113, 59–72. [Google Scholar] [CrossRef]
- Poloczanska, E.S.; Burrows, M.T.; Brown, C.J.; García Molinos, J.; Halpern, B.S.; Hoegh-Guldberg, O.; Kappel, V.; Moore, P.J.; Richardson, A.J.; Schoeman, D.S.; et al. Responses of Marine Organisms to Climate Change across Oceans. Front. Mar. Sci. 2016, 3, 62. [Google Scholar] [CrossRef] [Green Version]
- Chaikin, S.; Dubiner, S.; Belmaker, J. Cold-water species deepen to escape warm water temperatures. Glob. Ecol. Biogeogr. 2021, 31, 75–88. [Google Scholar] [CrossRef]
- Eriksson, S.P.; Hernroth, B.; Baden, S.P. Chapter Five—Stress Biology and Immunology in Nephrops norvegicus. Adv. Mar. Biol. 2001, 64, 149–200. [Google Scholar] [CrossRef]
- Hernroth, B.; Sköld, H.N.; Wiklander, K.; Jutfelt, F.; Baden, S. Simulated climate change causes immune suppression and protein damage in the crustacean Nephrops norvegicus. Fish Shellfish Immunol. 2012, 33, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Wood, H.L.; Eriksson, S.P.; Nordborg, M.; Styf, H.K. The effect of environmental stressors on the early development of the Norway lobster Nephrops norvegicus (L.). J. Exp. Mar. Biol. Ecol. 2015, 473, 35–42. [Google Scholar] [CrossRef]
- Chiarini, M.; Guicciardi, S.; Angelini, S.; Tuck, I.D.; Grilli, F.; Penna, P.; Domenichetti, F.; Canduci, G.; Belardinelli, A.; Santojanni, A.; et al. Accounting for environmental and fishery management factors when standardizing CPUE data from a scientific survey: A case study for Nephrops norvegicus in the Pomo Pits area (Central Adriatic Sea). PLoS ONE 2022, 17, e0270703. [Google Scholar] [CrossRef] [PubMed]
- Baden, S.P.; Pihl, L.; Rosenberg, R. Effects of oxygen depletion on the ecology, blood physiology and fishery of the Norway lobster Nephrops norvegicus. Mar. Ecol. Prog. Ser. 1990, 67, 141–155. [Google Scholar] [CrossRef]
- Spicer, J.; Baden, S. Environmental hypoxia and haemocyanin between-individual variability in Norway lobsters Nephrops norvegicus (L.). Mar. Biol. 2001, 139, 727–734. [Google Scholar] [CrossRef]
- Scott, K.N. Ocean acidification: A due diligence obligation under the LOSC. Int. J. Mar. Coast 2020, 35, 382–408. [Google Scholar] [CrossRef]
- Cooley, S.R.; Kite-Powell, H.L.; Doney, S.C. Ocean acidification’s potential to alter global marine ecosystem services. Oceanography 2009, 22, 172–181. [Google Scholar] [CrossRef] [Green Version]
- Cattano, C.; Claudet, J.; Domenici, P.; Milazzo, M. Living in a high CO2 world: A global meta-analysis shows multiple trait-mediated fish responses to ocean acidification. Ecol. Monogr. 2018, 88, 320–335. [Google Scholar] [CrossRef]
- Kroeker, K.J.; Kordas, R.L.; Crim, R.N.; Singh, G.G. Meta-analysis reveals negative yet variable effects of ocean acidification on marine organisms. Ecol. Lett. 2010, 13, 1419–1434. [Google Scholar] [CrossRef] [PubMed]
- Bednaršek, N.; Ambrose, R.; Calosi, P.; Childers, R.K.; Feely, R.A.; Litvin, S.Y.; Long, W.C.; Spicer, J.I.; Štrus, J.; Taylor, J.; et al. Synthesis of Thresholds of Ocean Acidification Impacts on Decapods. Front. Mar. Sci. 2021, 8, 651102. [Google Scholar] [CrossRef]
- Styf, H.K.; Sköld, H.N.; Eriksson, S.P. Embryonic response to long-term exposure of the marine crustacean Nephrops norvegicus to ocean acidification and elevated temperature. Ecol. Evol. 2013, 3, 5055–5065. [Google Scholar] [CrossRef] [PubMed]
- Saba, G.K.; Wright-Fairbanks, E.; Miles, T.N.; Chen, B.; Cai, W.J.; Wang, K.; Jones, C.P. Developing a profiling glider pH sensor for high resolution coastal ocean acidification monitoring. In Proceedings of the OCEANS 2018 MTS/IEEE Charleston, Charleston, SC, USA, 22–25 October 2018; pp. 1–8. [Google Scholar]
- Santana, C.S.; Olivares, D.M.M.; Silva, V.H.; Luzardo, F.H.; Velasco, F.G.; De Jesus, R.M. Assessment of water resources pollution associated with mining activity in a semi-arid region. J. Environ. Manag. 2020, 273, 111148. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.L. Theme and variations: Amphibious air-breathing intertidal fishes. J. Fish Biol. 2014, 84, 577–602. [Google Scholar] [CrossRef] [PubMed]
- Puig, P.; Canals, M.; Company, J.B.; Martín, J.; Amblas, D.; Lastras, G.; Palanques, A. Ploughing the deep sea floor. Nature 2012, 489, 286–289. [Google Scholar] [CrossRef] [Green Version]
- Matabos, M.; Aguzzi, J.; Robert, K.; Costa, C.; Menesatti, P.; Company, J.B.; Juniper, K. Multi-parametric study of behavioural modulation in demersal decapods at the VENUS cabled observatory in Saanich Inlet, British Columbia, Canada. J. Exp. Mar. Biol. Ecol. 2011, 401, 89–96. [Google Scholar] [CrossRef]
- Ozhikandathil, J.; Badilescu, S.; Packirisamy, M. A brief review on microfluidic platforms for hormones detection. J. Neural Transm. 2017, 124, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Violino, S.; Pallottino, F.; Sperandio, G.; Figorilli, S.; Ortenzi, L.; Tocci, F.; Vasta, S.; Imperi, G.; Costa, C. A full technological traceability system for extra virgin olive oil. Foods 2020, 9, 624. [Google Scholar] [CrossRef]
- Slesser, H.W.; Trueman, C.N. Traceability of the Norway Lobster Nephrops norvegicus in UK Shelf Seas: A Stable Isotope Approach. J. Shellfish Res. 2021, 40, 153–160. [Google Scholar] [CrossRef]
- Costa, C.; Pinsino, A.; Bonaventura, R.; Russo, R.; Zito, F.; Matranga, V. A pilot study for an innovative approach highlighting Actin and COI mRNAs as potential biomarkers of quality of the edible crustacean Nephrops norvegicus (Linnaeus, 1758). Food Control 2018, 97, 58–66. [Google Scholar] [CrossRef]
- Welden, N.A.; Cowie, P.R. Long-term microplastic retention causes reduced body condition in the langoustine, Nephrops norvegicus. Environ. Pollut. 2016, 218, 895–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carreras-Colom, E.; Cartes, J.E.; Rodríguez-Romeu, O.; Padrós, F.; Solé, M.; Grelaud, M.; Ziveri, P.; Palet, C.; Soler-Membrives, A.; Carrassón, M. Anthropogenic pollutants in Nephrops norvegicus (Linnaeus, 1758) from the NW Mediterranean Sea: Uptake assessment and potential impact on health. Environ. Pollut. 2022, 314, 120230. [Google Scholar] [CrossRef]
- Cau, A.; Avio, C.G.; Dessì, C.; Moccia, D.; Pusceddu, A.; Regoli, F.; Cannas, R.; Follesa, M.C. Benthic Crustacean Digestion Can Modulate the Environmental Fate of Microplastics in the Deep Sea. Environ. Sci. Technol. 2020, 54, 4886–4892. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, M.; Gomiero, A.; Guicciardi, S.; Frapiccini, E.; Strafella, P.; Angelini, S.; Domenichetti, F.; Belardinelli, A.; Colella, S. Preliminary results on the occurrence and anatomical distribution of microplastics in wild populations of Nephrops norvegicus from the Adriatic Sea. Environ. Pollut. 2021, 278, 116872. [Google Scholar] [CrossRef]
- Joyce, H.; Frias, J.; Kavanagh, F.; Lynch, R.; Pagter, E.; White, J.; Nash, R. Plastics, prawns, and patterns: Microplastic loadings in Nephrops norvegicus and surrounding habitat in the North East Atlantic. Sci. Total Environ. 2022, 826, 154036. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Affiliation | Country | Documents | % |
|---|---|---|---|
| Instituto de Ciencias del Mar (ICM-CSIC) | Spain | 80 | 5.3 |
| University of Glasgow | UK | 74 | 4.9 |
| Havforskningsinstituttet | Norway | 62 | 4.1 |
| Marine Scotland Information (MSI; Marine and Fisheries of the Scottish Government) | UK | 45 | 3.0 |
| Göteborgs Universitet | Sweden | 43 | 2.8 |
| Technical University of Denmark | Denmark | 42 | 2.8 |
| University Marine Biological Station Milliport | UK | 40 | 2.6 |
| Centre for the Environment Fisheries and Aquaculture Science | UK | 37 | 2.4 |
| Instituto Español de Oceanografía (IEO) | Spain | 34 | 2.2 |
| Consiglio Nazionale delle Ricerche (CNR) | Italy | 34 | 2.2 |
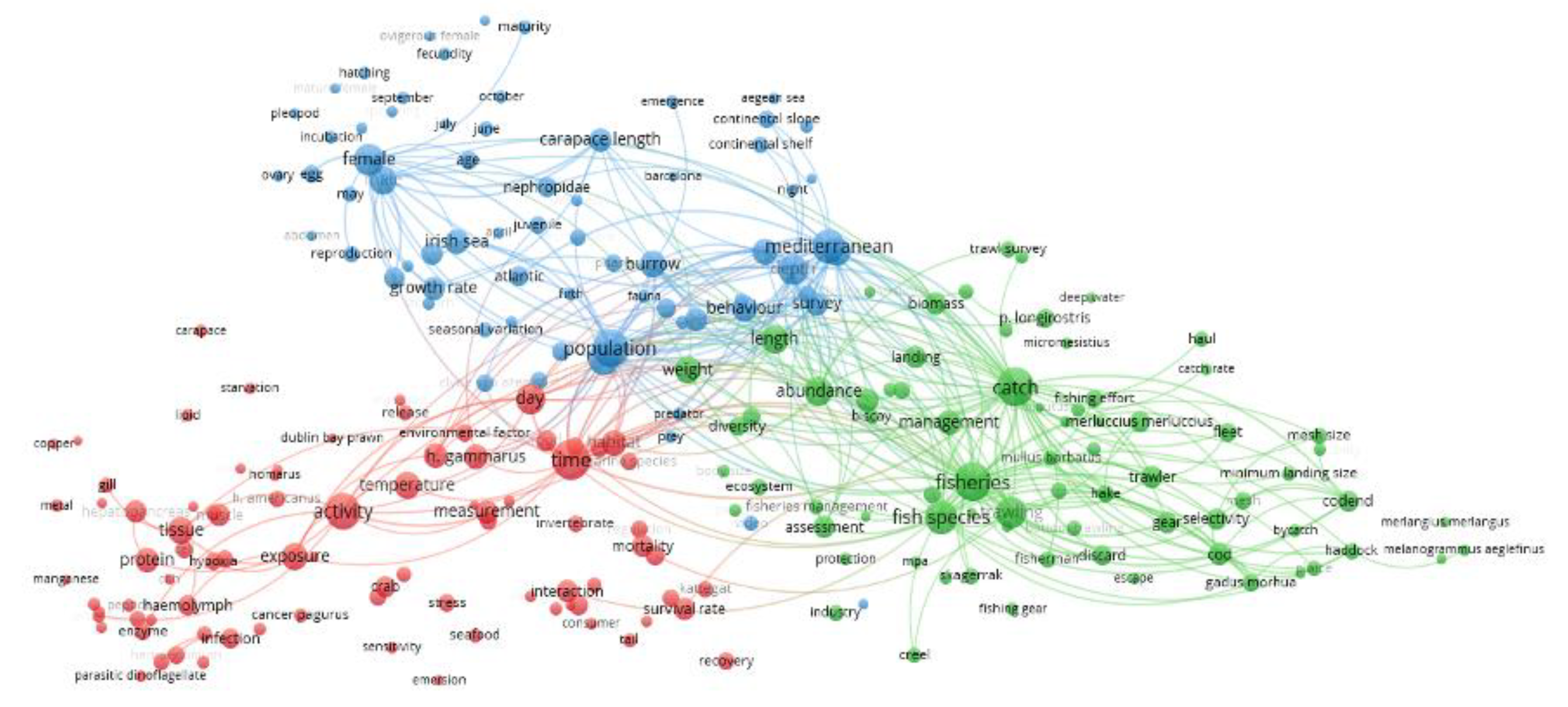
| Term | Score | Close to (or Visible) | Into the Cluster |
|---|---|---|---|
| microplastic | 39.842 | to "measurement" | RED |
| ingestion | 24.547 | to "release" | RED |
| ecosystem | 19.063 | VISIBLE | GREEN |
| stomach | 18.964 | to "growth rate" | BLUE |
| marine species | 18.117 | VISIBLE | RED |
| cephalopod | 16.703 | among "fisheries", "fish species" and "trawling" | GREEN |
| Mullus barbatus | 16.214 | VISIBLE | GREEN |
| Micromesistius | 15.859 | VISIBLE | GREEN |
| commercial species | 15.027 | right side of "fisheries" and "trawling" | GREEN |
| habitat | 14.838 | VISIBLE | RED |
| exposure | 14.799 | VISIBLE | RED |
| bottom trawling | 14.685 | VISIBLE | GREEN |
| management measure | 14.408 | right side of "catch" and "management" | GREEN |
| Barcelona | 14.152 | VISIBLE | BLUE |
| Aristeus antennatus | 14.139 | right side of "biomass" | GREEN |
| continental shelf | 14.088 | VISIBLE | BLUE |
| seafood | 13.925 | VISIBLE | RED |
| protection | 13.852 | VISIBLE | GREEN |
| parasitic dinoflagellate | 13.807 | VISIBLE | RED |
| abundance | 13.801 | VISIBLE | GREEN |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguzzi, J.; Violino, S.; Costa, C.; Bahamon, N.; Navarro, J.; Chatzievangelou, D.; Robinson, N.J.; Doyle, J.; Martinelli, M.; Lordan, C.; et al. Established and Emerging Research Trends in Norway Lobster, Nephrops norvegicus. Biology 2023, 12, 225. https://doi.org/10.3390/biology12020225
Aguzzi J, Violino S, Costa C, Bahamon N, Navarro J, Chatzievangelou D, Robinson NJ, Doyle J, Martinelli M, Lordan C, et al. Established and Emerging Research Trends in Norway Lobster, Nephrops norvegicus. Biology. 2023; 12(2):225. https://doi.org/10.3390/biology12020225
Chicago/Turabian StyleAguzzi, Jacopo, Simona Violino, Corrado Costa, Nixon Bahamon, Joan Navarro, Damianos Chatzievangelou, Nathan J. Robinson, Jennifer Doyle, Michela Martinelli, Colm Lordan, and et al. 2023. "Established and Emerging Research Trends in Norway Lobster, Nephrops norvegicus" Biology 12, no. 2: 225. https://doi.org/10.3390/biology12020225