
Effects of Tricaine Methanesulphonate (MS-222) on Physiological Stress and Fresh Quality of Sea Bass (Lateolabrax maculatus) under Simulated High-Density and Long-Distance Transport Stress


Abstract
:Simple Summary
Abstract
1. Introduction

{kind=link}
{kind=link}
{kind=link}
| Anesthetic | Characteristics | References |
|---|---|---|
| Tricaine Methanesulphonate (MS-222) |
| Liu et al. [8] |
| Clove Oil |
| Kamble et al. [22] |
| 2-Phenoxyethanol (C8H10O2) |
| Priborsky et al. [23] |
| CO2 |
| Oberg et al. [24] |
2. Materials and Methods
2.1. Preparation of Sea Bass
2.2. Experimental Design
2.3. Pre-Treatment of Samples
2.4. Determination of Indicators
2.4.1. Determination of Water Quality Indicators
2.4.2. Determination of Biochemical Blood Indicators
2.4.3. Determination of Nutrients in Fish
2.4.4. Determination of Flavor Substances in Fish
3. Statistical Analyses
4. Results and Discussion
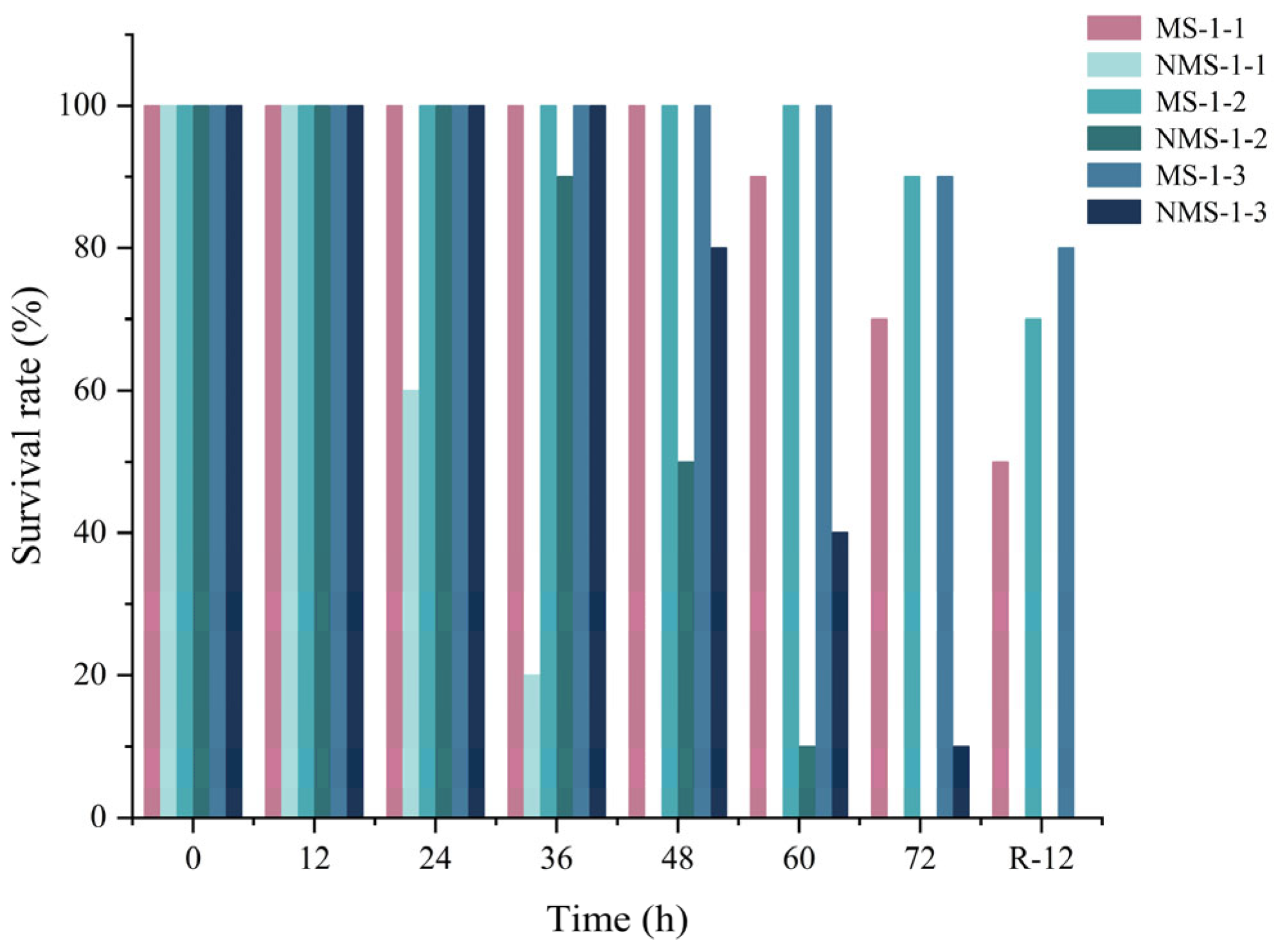
4.1. Survival Rate
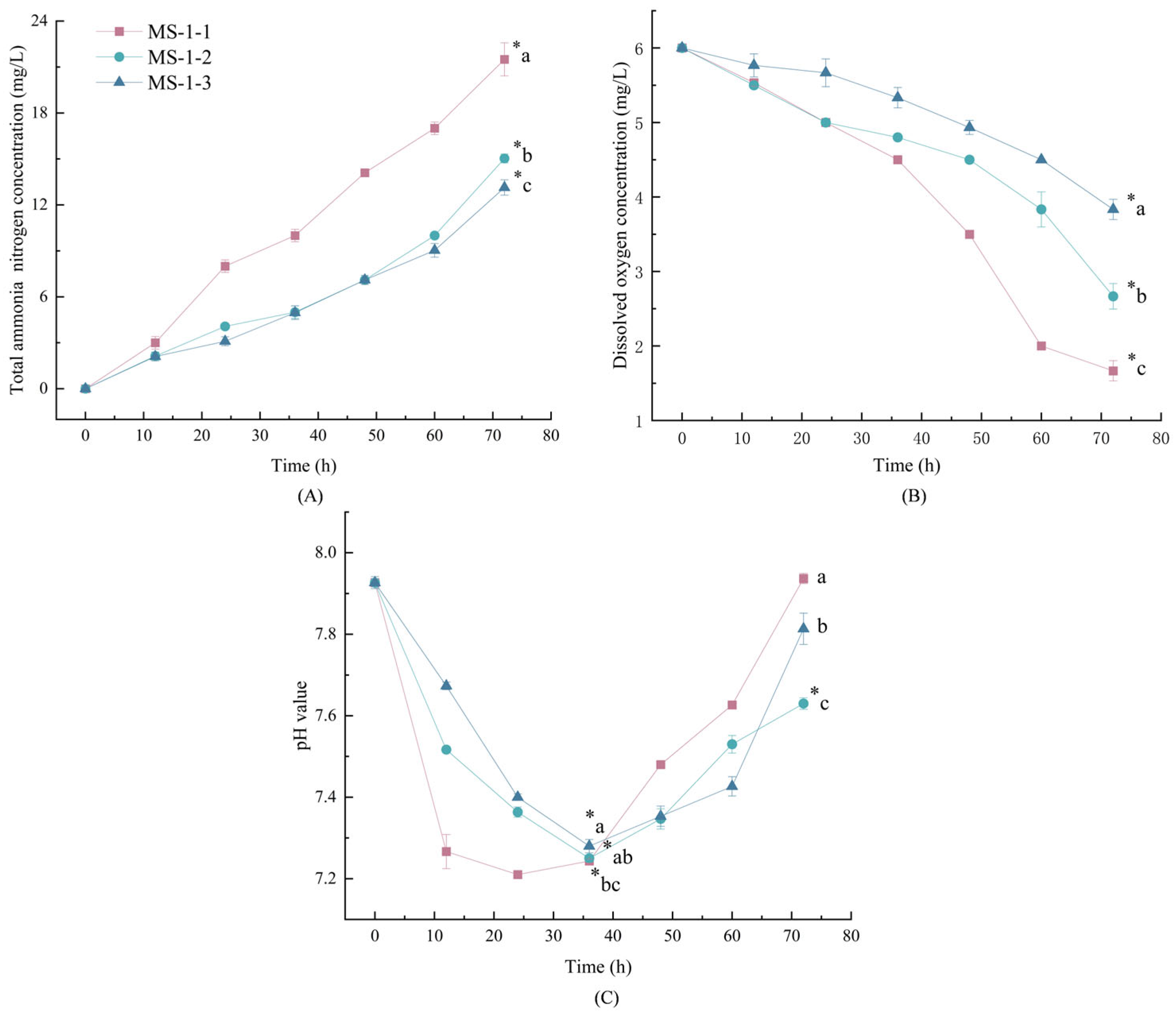
4.2. Water Quality Indicators
4.3. Blood Biochemical Indicators
4.4. Basic Nutrients
4.5. Nitrogenous Flavor Compounds: Free Amino Acids and Nucleotides
4.6. Non-Nitrogenous Flavor Compounds: Organic Acids and Inorganic Salts Ions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisheries Administration; Ministry of Agriculture. China Fishery Statistics Yearbook 2022; China Agriculture Press: Beijing, China, 2022.
- Liu, J.; Lan, W.; Sun, X.; Xie, J. Effects of chitosan grafted phenolic acid coating on microbiological, physicochemical and protein changes of sea bass (Lateolabrax japonicus) during refrigerated storage. J. Food Sci. 2020, 85, 2506–2515. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Q.; Mei, J.; Xie, J. Shelf-Life Extension of Refrigerated Turbot (Scophthalmus maximus) by Using Weakly Acidic Electrolyzed Water and Active Coatings Containing Daphnetin Emulsions. Front. Nutr. 2021, 8, 696212. [Google Scholar] [CrossRef]
- Nie, X.; Zhang, F.; Wang, T.; Zheng, X.; Li, Y.; Huang, B.; Zhang, C. Physiological and morphological changes in Turbot (Psetta maxima) gill tissue during waterless storage. Aquaculture 2019, 508, 30–35. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, Y.; Liu, Y.; Adanyi, N.; Zhang, X. Effects of waterless live transportation on survivability, physiological responses and flesh quality in Chinese farmed sturgeon (Acipenser schrenckii). Aquaculture 2020, 518, 734834. [Google Scholar] [CrossRef]
- Martins, T.; Valentim, A.; Pereira, N.; Antunes, L.M. Anaesthetics and analgesics used in adult fish for research: A review. Lab. Anim. 2019, 53, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Bowker, J.D.; Trushenski, J.T.; Bowman, M. Efficacy of eugenol to lightly sedate freshwater salmonids for an extended time period. N. A. J. Aquac. 2019, 81, 40–46. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhou, X.-W.; Ding, H.-T.; Dong, X.-J.; Zhang, J.-J.; Zheng, Y.-C.; Chen, X.-N.; Cheng, H.-L.; Ding, Z.-J.; Xu, J.-H. Effects of tricaine methanesulfonate (MS-222) on sedation and responses of yellow catfish (Pelteobagrus fulvidraco) subjected to simulated transportation stress. Aquaculture 2022, 549, 737789. [Google Scholar] [CrossRef]
- Wagner, G.N.; Singer, T.D.; McKinley, R.S. The ability of clove oil and MS-222 to minimize handling stress in rainbow trout (Oncorhynchus mykiss Walbaum). Aquac. Res. 2003, 34, 1139–1146. [Google Scholar] [CrossRef]
- Ouyang, M.; Wen, B.; Ma, H.; Chen, C.; Gao, J.; Zhang, Y.; Chen, Z. Minimally invasive evaluation of the anaesthetic efficacy of MS-222 for ornamental discus fish using skin mucus biomarkers. Aquac. Res. 2020, 51, 2926–2935. [Google Scholar] [CrossRef]
- Coyle, S.D.; Durrorow, R.M.; Tidwell, H.J. Effects of an aesthetics on the water parameters in a simulated transport experiment of platy fish (Xiphorus maculatus). Aquac. Res. 2004, 26, 256–271. [Google Scholar] [CrossRef]
- Zhao, J.; Zhu, Y.; He, Y.; Chen, J.; Feng, X.; Li, X.; Xiong, B.; Yang, D. Effects of Temperature Reduction and MS-222 on Water Quality and Blood Biochemistry in Simulated Transport Experiment of Largemouth Bronze Gudgeon, Coreius guichenoti. J. World Aquac. Soc. 2014, 45, 493–507. [Google Scholar] [CrossRef]
- De Oliveira, C.P.B.; da Paixão Lemos, C.H.; Vidal, L.V.O.; Couto, R.D.; Pereira, D.S.P.; Copatti, C.E. Anaesthesia with eugenol in hybrid Amazon catfish (Pseudoplatystoma reticulatum× Leiarius marmoratus) handling: Biochemical and haematological responses. Aquaculture 2019, 501, 255–259. [Google Scholar] [CrossRef]
- Hong, J.; Chen, X.; Liu, S.; Fu, Z.; Han, M.; Wang, Y.; Gu, Z.; Ma, Z. Impact of fish density on water quality and physiological response of golden pompano (Trachinotus ovatus) flingerlings during transportation. Aquaculture 2019, 507, 260–265. [Google Scholar] [CrossRef]
- Cléach, J.; Soret, M.; Grard, T.; Lencel, P. Assessment of fish freshness based on fluorescence measurement of mitochondrial membrane potential. Food Control 2020, 115, 107301. [Google Scholar] [CrossRef]
- Cao, J.; Wang, Q.; Mei, J.; Xie, J. Effect of 3-Aminobenzoic Acid Ethyl Ester Methanesulfonate (MS-222) on Quality of Marine Cultured Turbot (Scophthalmus maximus) during Simulated Transport in Water. Fishes 2021, 6, 20. [Google Scholar] [CrossRef]
- Wu, B.; Wang, Q.; Cao, J.; Mei, J.; Xie, J. Effects of Ascorbic Acid and beta-1,3-Glucan on Survival, Physiological Response and Flesh Quality of Cultured Tiger Grouper (Epinephelus fuscoguttatus) during Simulated Transport in Water. Biology 2020, 9, 37. [Google Scholar] [CrossRef] [Green Version]
- Espinoza-Ramos, L.A.; Pepe-Victoriano, R.; Huanacuni, J.I.; Nande, M. Effect of transportation time and stocking density on seawater quality and survival of Anisotremus scapularis (Perciformes: Haemulidae). J. World Aquac. Soc. 2022, 53, 1042–1050. [Google Scholar] [CrossRef]
- Garcés-Rodríguez, Y.; Sánchez-Velasco, L.; Parés-Sierra, A.; Jiménez-Rosenberg, S.P.A.; Márquez-Artavia, A.; Flores-Morales, A.L. FISH larvae distribution and transport on the thermal fronts in the Midriff Archipelago region, Gulf of California. Cont. Shelf Res. 2021, 218, 104384. [Google Scholar] [CrossRef]
- Vanderzwalmen, M.; Carey, P.; Snellgrove, D.; Sloman, K.A. Benefits of enrichment on the behaviour of ornamental fishes during commercial transport. Aquaculture 2020, 526, 735360. [Google Scholar] [CrossRef]
- Wu, S.M.; Chen, J.-R.; Chang, C.-y.; Tseng, Y.-J.; Pan, B.S. Potential benefit of I-Tiao-Gung (Glycine tomentella) extract to enhance ornamental fish welfare during live transport. Aquaculture 2021, 534, 736304. [Google Scholar] [CrossRef]
- Kamble, A.D.; Saini, V.; Ojha, M. The efficacy of clove oil as anesthetic in common carp (Cyprinus carpio) and its potential metabolism reducing capacity. Int. J. Fauna Biol. Stud. 2014, 1, 1–6. [Google Scholar]
- Priborsky, J.; Velisek, J. A review of three commonly used fish anesthetics. Rev. Fish. Sci. Aquac. 2018, 26, 417–442. [Google Scholar] [CrossRef]
- Oberg, E.; Perez, K.; Fuiman, L. Carbon dioxide is an effective anesthetic for multiple marine fish species. Fish. Res. 2015, 165, 22–27. [Google Scholar] [CrossRef]
- De Oliveira, C.P.B.; Lemos, C.H.d.P.; e Silva, A.F.; de Souza, S.A.; Albinati, A.C.L.; Lima, A.O.; Copatti, C.E. Use of eugenol for the anaesthesia and transportation of freshwater angelfish (Pterophyllum scalare). Aquaculture 2019, 513, 734409. [Google Scholar] [CrossRef]
- Salbego, J.; Toni, C.; Becker, A.G.; Zeppenfeld, C.C.; Menezes, C.C.; Loro, V.L.; Heinzmann, B.M.; Baldisserotto, B. Biochemical parameters of silver catfish (Rhamdia quelen) after transport with eugenol or essential oil of Lippia alba added to the water. Braz. J. Biol. 2017, 77, 696–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Mei, J.; Xie, J. Effects of multi-frequency ultrasound on the freezing rates, quality properties and structural characteristics of cultured large yellow croaker (Larimichthys crocea). Ultrason. Sonochem. 2021, 76, 105657. [Google Scholar] [CrossRef]
- Favero, G.C.; Silva, W.d.S.e.; Boaventura, T.P.; Paes Leme, F.d.O.; Luz, R.K. Eugenol or salt to mitigate stress during the transport of juvenile Lophiosilurus alexandri, a Neotropical carnivorous freshwater catfish. Aquaculture 2019, 512, 734321. [Google Scholar] [CrossRef]
- Boaventura, T.P.; Souza, C.F.; Ferreira, A.L.; Favero, G.C.; Baldissera, M.D.; Heinzmann, B.M.; Baldisserotto, B.; Luz, R.K. The use of Ocimum gratissimum L. essential oil during the transport of Lophiosilurus alexandri: Water quality, hematology, blood biochemistry and oxidative stress. Aquaculture 2021, 531, 735964. [Google Scholar] [CrossRef]
- Zhang, H.; Fang, D.; Mei, J.; Xie, J.; Qiu, W. A Preliminary Study on the Effects of Nitrite Exposure on Hematological Parameters, Oxidative Stress, and Immune-Related Responses in Pearl Gentian Grouper. Fishes 2022, 7, 235. [Google Scholar] [CrossRef]
- Cao, J.; Wang, Q.; Ma, T.; Bao, K.; Yu, X.; Duan, Z.; Shen, X.; Li, C. Effect of EGCG-gelatin biofilm on the quality and microbial composition of tilapia fillets during chilled storage. Food Chem. 2020, 305, 125454. [Google Scholar] [CrossRef]
- Chamberlain, I.; Adams, K.; Le, S. ICP-MS determination of trace elements in fish. At. Spectrosc. 2000, 21, 118–122. [Google Scholar]
- Oshima, H.; Ueno, E.; Saito, I.; Matsumoto, H. A comparative study of cadmium, lead, mercury, arsenic, selenium, manganese, copper and zinc in brown rice and fish by inductively coupled plasma-mass spectrometry (ICP-MS) and atomic absorption spectrometry. J. Food Hyg. Soc. Jpn. 2004, 45, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Ki-Hwan, S.; Jong-Ho, L.; Yeoung-Lae, H.; Shang-Do, C.; Kwon-Il, S.; Ok-Soo, J. Changes of organic acid contents on heating conditions of fishes. J. Korean Soc. Food Nutr. 1994, 23, 939–944. [Google Scholar]
- Carneiro, P.C.F.; Kaiseler, P.H.d.S.; Swarofsky, E.d.A.C.; Baldisserotto, B. Transport of jundiá Rhamdia quelen juveniles at different loading densities: Water quality and blood parameters. Neotrop. Ichthyol. 2009, 7, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Franklin, D.A.; Edward, L. Ammonia toxicity and adaptive response in marine fishes. Indian J. Geo-Mar. Sci. (IJMS) 2019, 48, 273–279. [Google Scholar]
- Harmon, T.S. Methods for reducing stressors and maintaining water quality associated with live fish transport in tanks: A review of the basics. Rev. Aquac. 2009, 1, 58–66. [Google Scholar] [CrossRef]
- Paterson, B.D.; Rimmer, M.A.; Meikle, G.M.; Semmens, G.L. Physiological responses of the Asian sea bass, Lates calcarifer to water quality deterioration during simulated live transport: Acidosis, red-cell swelling, and levels of ions and ammonia in the plasma. Aquaculture 2003, 218, 717–728. [Google Scholar] [CrossRef]
- Pramod, P.K.; Sajeevan, T.P.; Ramachandran, A.; Thampy, S.; Pai, S.S. Effects of Two Anesthetics on Water Quality during Simulated Transport of a Tropical Ornamental Fish, the Indian tiger barb Puntius filamentosus. N. Am. J. Aquac. 2010, 72, 290–297. [Google Scholar] [CrossRef]
- Velisek, J.; Stara, A.; Li, Z.-H.; Silovska, S.; Turek, J. Comparison of the effects of four anaesthetics on blood biochemical profiles and oxidative stress biomarkers in rainbow trout. Aquaculture 2011, 310, 369–375. [Google Scholar] [CrossRef]
- Palić, D.; Herolt, D.M.; Andreasen, C.B.; Menzel, B.W.; Roth, J.A. Anesthetic efficacy of tricaine methanesulfonate, metomidate and eugenol: Effects on plasma cortisol concentration and neutrophil function in fathead minnows (Pimephales promelas Rafinesque, 1820). Aquaculture 2006, 254, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Fatira, E.; Papandroulakis, N.; Pavlidis, M. Diel changes in plasma cortisol and effects of size and stress duration on the cortisol response in European sea bass (Dicentrarchus labrax). Fish Physiol. Biochem. 2014, 40, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Dong, Z.; Yang, Y.; Wang, J.; Yang, T.; Chen, X.; Liang, L.; Mu, W. Histology, physiology, and glucose and lipid metabolism of Lateolabrax maculatus under low temperature stress. J. Therm. Biol. 2022, 104, 103161. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Cao, X.; Wang, Y.; Kuang, S.; Hu, J.; Yang, Y.; Xu, S.; Zhang, M.; Sun, Y.; Gu, W. Reduced stress responses by MS-222 in juvenile silver pomfret (Pampus argenteus). J. World Aquac. Soc. 2020, 51, 1192–1207. [Google Scholar] [CrossRef]
- Wang, Q.; Mei, J.; Cao, J.; Xie, J. Effects of Melissa officinalis L. Essential Oil in Comparison with Anaesthetics on Gill Tissue Damage, Liver Metabolism and Immune Parameters in Sea Bass (Lateolabrax maculatus) during Simulated Live Transport. Biology 2021, 11, 11. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, N.; Pal, A.K.; Das, T.; Mohammed, M.S.; Sarma, K.; Venkateshwarlu, G.; Mukherjee, S.C. Secondary stress responses in Indian major carps Labeo rohita (Hamilton), Catla catla (Hamilton) and Cirrhinus mrigala (Hamilton) fry to increasing packing densities. Aquac. Res. 2006, 37, 472–476. [Google Scholar] [CrossRef]
- Kristoffersen, S.; Tobiassen, T.; Steinsund, V.; Olsen, R.L. Slaughter stress, postmortem muscle pH and rigor development in farmed Atlantic cod (Gadus morhua L.). Int. J. Food Sci. Technol. 2006, 41, 861–864. [Google Scholar] [CrossRef]
- Osako, K.; Fujii, A.; Ruttanapornvareesakul, Y.; Nagano, N.; Kuwahara, K.; Okamoto, A. Differences in free amino acid composition between testis and ovary of sea urchin Anthocidaris crassispina during gonadal development. Fish. Sci. 2007, 73, 660–667. [Google Scholar] [CrossRef]
- Dawei, Y.; Yanshun, X.; Regenstein, J.M.; Wenshui, X.; Fang, Y.; Qixing, J.; Bin, W. The effects of edible chitosan-based coatings on flavor quality of raw grass carp (Ctenopharyngodon idellus) fillets during refrigerated storage. Food Chem. 2018, 242, 412–420. [Google Scholar] [CrossRef]
- Bian, C.; Yu, H.; Yang, K.; Mei, J.; Xie, J. Effects of single-, dual-, and multi-frequency ultrasound-assisted freezing on the muscle quality and myofibrillar protein structure in large yellow croaker (Larimichthys crocea). Food Chem. X 2022, 15, 100362. [Google Scholar] [CrossRef]
- Shao, T.; Chen, X.; Zhai, D.; Wang, T.; Long, X.; Liu, Z. Evaluation of the effects of different stocking densities on growth and stress responses of juvenile hybrid grouper♀ Epinephelus fuscoguttatus × ♂ Epinephelus lanceo latus in recirculating aquaculture systems. J. Fish Biol. 2019, 95, 1022–1029. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Vatnikov, Y.A.; Kulikov, E.V.; Petrov, A.K.; Hoseinifar, S.H.; Van Doan, H. Effects of dietary arginine supplementation on ureagenesis and amino acid metabolism in common carp (Cyprinus carpio) exposed to ambient ammonia. Aquaculture 2019, 511, 734209. [Google Scholar] [CrossRef]
- De Aguiar Saldanha Pinheiro, A.C.; Urbinati, E.; Tappi, S.; Picone, G.; Patrignani, F.; Lanciotti, R.; Romani, S.; Rocculi, P. The impact of gas mixtures of Argon and Nitrous oxide (N2O) on quality parameters of sardine (Sardina pilchardus) fillets during refrigerated storage. Food Res. Int. 2019, 115, 268–275. [Google Scholar] [CrossRef]
- De-Wei, C.; Min, Z. Non-volatile taste active compounds in the meat of Chinese mitten crab (Eriocheir sinensis). Food Chem. 2007, 104, 1200–1205. [Google Scholar] [CrossRef]
- Hong, H.; Regenstein, J.M.; Luo, Y. The importance of ATP-related compounds for the freshness and flavor of post-mortem fish and shellfish muscle: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 1787–1798. [Google Scholar] [CrossRef] [PubMed]
- Wenxian, Y.; Wenzheng, S.; Shengnan, Z.; Yinghong, Q.; Zhihe, W. Research on the changes of water-soluble flavor substances in grass carp during steaming. J. Food Biochem. 2019, 43, e12993. [Google Scholar] [CrossRef]
- Cao, J.; Wang, Q.; Qiu, W.; Mei, J.; Xie, J. Transport and Recovery of Turbot (Scophthalmus maximus) Sedated with MS-222 and Eugenol: Effects on Intermediary Metabolism and Osmoregulation. Animals 2021, 11, 2228. [Google Scholar] [CrossRef]
- Mohan, C.O.; Ravishankar, C.N.; Kumar, K.A.; Gopal, T.K.S. Biogenic amines and nucleotide breakdown products of sodium acetate, sodium lactate, and sodium citrate treated seer fish (Scomberomorus commerson) during iced storage. J. Food Saf. 2019, 39, e12633. [Google Scholar] [CrossRef]
- Dong, Y.; Zhang, H.; Mei, J.; Xie, J.; Shao, C. Advances in Application of Ultrasound in Meat Tenderization: A review. Front. Sustain. Food Syst. 2022, 2022, 360. [Google Scholar] [CrossRef]
- Lall, S.P.; Kaushik, S.J. Nutrition and Metabolism of Minerals in Fish. Animals 2021, 11, 2711. [Google Scholar] [CrossRef] [PubMed]
- Vasil’Eva, E.N.; Diubiuk, N.E.; Lychnikova, T.D. Mineral composition of certain types of fish and verification of the relationship between mineral and protein contents. Vopr. Pitan. 1961, 20, 54–59. [Google Scholar] [PubMed]
- Galan-Garcia, D.; Oliver-Pujol, R.; Estrany-Coda, F. Fish as food. Aliment. Equipos Y Tecnol. 2005, 24, 115–118. [Google Scholar]
- Martinez Para, M.C.; Masoud, T.A.; Aguilar, M.V.; Polo, M.I. Minerals in preserved fish and shellfish. Alimentaria 1983, 1983, 37–43. [Google Scholar]
- Goss, G.G.; Wood, C.M. The effects of acid and acid/aluminum exposure on circulating plasma cortisol levels and other blood parameters in the rainbow trout, Salmo gaivdneri. J. Fish Biol. 2010, 32, 63–76. [Google Scholar] [CrossRef]
- Fuke, S.; Konosu, S. Taste-active components in some foods: A review of Japanese research. Physiol. Behav. 1991, 49, 863–868. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Ma, X.; Xie, J. Review on natural preservatives for extending fish shelf life. Foods 2019, 8, 490. [Google Scholar] [CrossRef] [Green Version]



| Indicators | Samples | Simulated Transport Time (h) | ||
|---|---|---|---|---|
| 0 (CK) | 72 | Recover-12 | ||
| Cortisol (COR) (ng/L) | MS-1-1 | 178.37 ± 4.67 a | 264.20 ± 9.48 Ab | 227.54 ± 9.42 Ab |
| MS-1-2 | 240.87 ± 8.90 Ab | 224.62 ± 7.13 Aab | ||
| MS-1-3 | 201.29 ± 2.59 Ba | 189.62 ± 3.53 Aa | ||
| Glucose (GLU) (mmoL/L) | MS-1-1 | 1.48 ± 0.10 a | 8.34 ± 0.39 Ab | 3.46 ± 0.09 Ac |
| MS-1-2 | 4.29 ± 0.02 Bb | 2.69 ± 0.02 Bc | ||
| MS-1-3 | 3.54 ± 0.14 Cb | 1.95 ± 0.19 Cc | ||
| Lactic acid (LD) (mmoL/L) | MS-1-1 | 1.65 ± 0.01 a | 3.24 ± 0.00 Ab | 1.93 ± 0.04 Ac |
| MS-1-2 | 3.23 ± 0.04 Ab | 1.83 ± 0.01 Bc | ||
| MS-1-3 | 3.75 ± 0.12 Bb | 2.03 ± 0.04 Ac | ||
| Aspartate aminotransferase (AST) (U/L) | MS-1-1 | 94.23 ± 1.27 a | 198.44 ± 2.71 Ab | 123.50 ± 2.93 Ac |
| MS-1-2 | 176.86 ± 6.43 Bb | 107.71 ± 2.91 Bc | ||
| MS-1-3 | 171.71 ± 2.35 Bb | 110.01 ± 4.44 Bc | ||
| Alanine aminotransferase (ALT) (U/L) | MS-1-1 | 48.92 ± 1.55 a | 85.34 ± 2.42 Ab | 67.08 ± 2.54 Ac |
| MS-1-2 | 80.35 ± 2.84 Ab | 54.36 ± 2.64 Ba | ||
| MS-1-3 | 74.09 ± 13.07 Ab | 51.10 ± 2.67 Ba | ||
| Alkaline phosphatase (ALP) (U/L) | MS-1-1 | 14.986 ± 0.294 a | 9.01 ± 0.18 Ab | 13.89 ± 0.16 Ac |
| MS-1-2 | 8.37 ± 0.19 Bb | 13.28 ± 0.172 Bc | ||
| MS-1-3 | 7.62 ± 0.022 Cb | 13.97 ± 0.271 Ac | ||
| Indicators | Samples | Simulated Transport Time (h) | ||
|---|---|---|---|---|
| 0 (CK) | 72 | Recover-12 | ||
| moisture content (%) | MS-1-1 | 76.68 ± 0.20 a | 77.11 ± 0.106 Aa | 76.95 ± 0.436 Aa |
| MS-1-2 | 76.69 ± 0.174 BCa | 77.25 ± 0.28 Aa | ||
| MS-1-3 | 76.58 ± 0.212 Ca | 76.76 ± 0.28 Aa | ||
| Crude fat content (%) | MS-1-1 | 7.24 ± 0.09 a | 4.87 ± 0.11 Ab | 2.92 ± 0.06 Ac |
| MS-1-2 | 5.58 ± 0.15 Bb | 3.70 ± 0.16 Bc | ||
| MS-1-3 | 6.23 ± 0.39 Bb | 5.29 ± 0.11 Cc | ||
| Total protein content (μg/mL) | MS-1-1 | 573.96 ± 15.16 a | 416.94 ± 24.26 Ab | 475.82 ± 14.88 Ac |
| MS-1-2 | 479.39 ± 22.39 Bb | 481.77 ± 24.91 Ab | ||
| MS-1-3 | 496.64 ± 15.35 Bb | 509.13 ± 8.86 Ab | ||
| Muscle glycogen (mg/g) | MS-1-1 | 3.52 ± 0.03 a | 2.44 ± 0.02 Ab | 2.66 ± 0.01 Ac |
| MS-1-2 | 2.31 ± 0.01 Bb | 2.93 ± 0.01 Bc | ||
| MS-1-3 | 2.95 ± 0.00 Cb | 3.14 ± 0.10 Cc | ||
| Indicators | Samples | Simulated Transport Time (h) | ||
|---|---|---|---|---|
| 0 (CK) | 72 | Recover-12 | ||
| Aspartic Acid # (Asp) (mg/kg) | MS-1-1 | 2.68 ± 0.11 a | 2.89 ± 0.16 Aa | 2.77 ± 0.06 Aa |
| MS-1-2 | 2.76 ± 0.09 Aa | 2.60 ± 0.10 Aa | ||
| MS-1-3 | 2.83 ± 0.08 Aa | 2.79 ± 0.02 Aa | ||
| Threonine * (Thr) (mg/kg) | MS-1-1 | 9.63 ± 0.11 a | 11.13 ± 0.05 Ab | 9.86 ± 0.11 Aab |
| MS-1-2 | 10.60 ± 0.04 Ab | 9.73 ± 0.04 Aa | ||
| MS-1-3 | 10.29 ± 0.09 Aa | 9.70 ± 0.06 Aa | ||
| Serine * (Ser) (mg/kg) | MS-1-1 | 13.60 ± 0.05 a | 15.94 ± 0.05 Ab | 15.56 ± 0.06 Ab |
| MS-1-2 | 15.77 ± 0.04 Bb | 13.50 ± 0.03 Aa | ||
| MS-1-3 | 14.80 ± 0.00 Ba | 13.33 ± 0.25 Aa | ||
| Glutamic Acid # (Glu) (mg/kg) | MS-1-1 | 31.24 ± 0.02 a | 35.91 ± 0.04 Ab | 33.07 ± 0.02 Ac |
| MS-1-2 | 36.52 ± 0.19 Ab | 33.66 ± 0.09 Aab | ||
| MS-1-3 | 33.93 ± 0.01 Ab | 32.81 ± 0.04 Ac | ||
| Glycine * (Gly) (mg/kg) | MS-1-1 | 95.02 ± 0.01 a | 99.58 ± 0.01 Aa | 97.60 ± 0.07 Aa |
| MS-1-2 | 98.95 ± 0.04 Aa | 97.14 ± 0.35 Aa | ||
| MS-1-3 | 99.33 ± 0.33 Aa | 96.40 ± 0.71 Aa | ||
| Alanine * (Ala) (mg/kg) | MS-1-1 | 78.41 ± 0.02 a | 91.65 ± 0.04 Ab | 81.76 ± 0.50 Aa |
| MS-1-2 | 89.54 ± 0.01 Bb | 79.72 ± 0.03 Ac | ||
| MS-1-3 | 85.78 ± 0.09 Cb | 78.55 ± 0.48 Aa | ||
| Valine (Val) (mg/kg) | MS-1-1 | 11.70 ± 0.00 a | 13.19 ± 0.08 Ab | 12.27 ± 0.00 Aab |
| MS-1-2 | 12.60 ± 0.01 Aa | 12.00 ± 0.09 Aa | ||
| MS-1-3 | 12.81 ± 0.02 Aa | 11.93 ± 0.18 Aa | ||
| Methionine (Met) (mg/kg) | MS-1-1 | 6.24 ± 0.00 a | 7.86 ± 0.02 Ab | 6.66 ± 0.06 Aa |
| MS-1-2 | 7.58 ± 0.66 Ab | 6.50 ± 0.06 Aab | ||
| MS-1-3 | 7.15 ± 0.03 Ab | 6.11 ± 0.02 Aa | ||
| Isoleucine (Ile) (mg/kg) | MS-1-1 | 8.03 ± 0.01 a | 9.64 ± 0.10 Ab | 9.03 ± 0.00 Aab |
| MS-1-2 | 9.23 ± 0.05 Ab | 9.08 ± 0.01 Ab | ||
| MS-1-3 | 9.17 ± 0.00 Aa | 9.07 ± 0.13 Aa | ||
| Leucine (Leu) (mg/kg) | MS-1-1 | 14.76 ± 0.01 a | 16.95 ± 0.39 Aa | 16.08 ± 0.01 Aa |
| MS-1-2 | 16.80 ± 0.00 Ab | 15.89 ± 0.01 Ac | ||
| MS-1-3 | 16.26 ± 0.20 Aa | 15.59 ± 0.14 Aa | ||
| Tyrosine (Tyr) (mg/kg) | MS-1-1 | 3.03 ± 0.01 a | 5.28 ± 0.02 Ab | 3.61 ± 0.00 Ac |
| MS-1-2 | 4.99 ± 0.01 Ab | 3.47 ± 0.02 Ac | ||
| MS-1-3 | 4.08 ± 0.00 Bb | 3.19 ± 0.00 Bc | ||
| Phenylalanine (Phe) (mg/kg) | MS-1-1 | 2.93 ± 0.01 a | 4.99 ± 0.00 Ab | 4.06 ± 0.09 Aab |
| MS-1-2 | 4.79 ± 0.02 Ab | 3.36 ± 0.05 Aa | ||
| MS-1-3 | 3.88 ± 0.01 Ba | 3.23 ± 0.06 Aa | ||
| Lysine (Lys) #* (mg/kg) | MS-1-1 | 55.89 ± 0.02 a | 86.49 ± 0.19 Ab | 73.00 ± 0.16 Ac |
| MS-1-2 | 79.52 ± 0.01 Bb | 67.09 ± 0.04 Bc | ||
| MS-1-3 | 73.79 ± 0.03 Cb | 62.28 ± 0.01 Cc | ||
| Histidine (His) (mg/kg) | MS-1-1 | 8.04 ± 0.01 a | 13.19 ± 0.00 Ab | 10.55 ± 0.01 ABc |
| MS-1-2 | 13.61 ± 0.07 Ab | 11.94 ± 0.08 Ac | ||
| MS-1-3 | 11.74 ± 0.00 Bb | 10.04 ± 0.10 Bc | ||
| Arginine (Arg) (mg/kg) | MS-1-1 | 8.50 ± 0.11 a | 11.73 ± 0.20 Aa | 9.04 ± 0.02 Aa |
| MS-1-2 | 11.70 ± 0.00 Ab | 8.65 ± 0.00 Aa | ||
| MS-1-3 | 10.90 ± 0.10 Ab | 8.17 ± 0.07 Aa | ||
| Total (mg/kg) | MS-1-1 | 349.71 ± 5.00 a | 422.43 ± 20.19 Ab | 383.33 ± 11.66 Ac |
| MS-1-2 | 412.95 ± 6.49 Ab | 373.36 ± 10.12 Aa | ||
| MS-1-3 | 393.58 ± 9.89 Ab | 362.60 ± 22.63 Aab | ||
| IMP (mg/100g) | MS-1-1 | 283.44 ± 17.15 a | 319.35 ± 4.57 Ab | 292.10 ± 6.17 Aa |
| MS-1-2 | 304.14 ± 7.81 Ba | 280.84 ± 5.01 Aa | ||
| MS-1-3 | 299.29 ± 5.27 Ba | 285.38 ± 1.77 Aa | ||
| AMP (mg/100g) | MS-1-1 | 26.30 ± 2.16 a | 18.19 ± 2.29 Ab | 23.34 ± 1.67 Aa |
| MS-1-2 | 19.91 ± 0.65 Ab | 24.20 ± 2.27 Aab | ||
| MS-1-3 | 21.10 ± 0.44 Ab | 25.72 ± 1.70 Aa | ||
| Indicators | Samples | Simulated Transport Time (h) | ||
|---|---|---|---|---|
| 0 h (CK) | 72 | Recover-12 | ||
| Lactic acid (mg/kg) | MS-1-1 | 161.11 ± 0.96 a | 334.49 ± 1.73 Ab | 227.64 ± 0.15 Ac |
| MS-1-2 | 300.20 ± 0.56 Bb | 224.85 ± 1.02 Ac | ||
| MS-1-3 | 281.97 ± 0.79 Cb | 209.78 ± 2.18 Bc | ||
| Acetic acid (mg/kg) | MS-1-1 | 97.25 ± 1.74 a | 141.74 ± 1.54 Ab | 128.64 ± 2.15 Ac |
| MS-1-2 | 135.58 ± 1.72 Bb | 114.61 ± 4.55 Bc | ||
| MS-1-3 | 135.34 ± 1.68 Bb | 113.42 ± 0.06 Bc | ||
| Fumaric acid (mg/kg) | MS-1-1 | 1.07 ± 0.01 a | 1.60 ± 0.01 Ab | 1.30 ± 0.00 Ac |
| MS-1-2 | 1.41 ± 0.00 Bb | 1.29 ± 0.01 ABc | ||
| MS-1-3 | 1.30 ± 0.00 Cb | 1.26 ± 0.01 Bc | ||
| Na (mg/kg) | MS-1-1 | 480.20 ± 52.09 a | 764.98 ± 28.92 Ab | 628.77 ± 6.51 Ac |
| MS-1-2 | 821.05 ± 9.97 Ab | 783.19 ± 41.69 Bb | ||
| MS-1-3 | 664.91 ± 123.52 Aa | 652.22 ± 28.27 Aa | ||
| Mg (mg/kg) | MS-1-1 | 483.92 ± 35.52 a | 602.21 ± 23.97 Ab | 561.72 ± 6.61 Ab |
| MS-1-2 | 556.93 ± 42.39 Aa | 532.18 ± 19.61 Aa | ||
| MS-1-3 | 561.01 ± 86.36 Aa | 521.93 ± 24.51 Aa | ||
| P (mg/kg) | MS-1-1 | 2729.06 ± 71.26 a | 3309.95 ± 190.99 Aa | 3054.43 ± 121.93 Aa |
| MS-1-2 | 3214.50 ± 330.87 Aa | 2821.68 ± 74.99 Aa | ||
| MS-1-3 | 2949.47 ± 257.73 Aa | 2829.35 ± 97.49 Aa | ||
| K (mg/kg) | MS-1-1 | 7358.79 ± 202.85 a | 9260.77 ± 445.85 Ab | 8033.81 ± 242.31 Aa |
| MS-1-2 | 8568.97 ± 406.25 ABb | 7447.34 ± 256.76 ABa | ||
| MS-1-3 | 7880.18 ± 504.78 Ba | 7410.30 ± 219.16 Ba | ||
| Ca (mg/kg) | MS-1-1 | 60.67 ± 7.80 a | 61.38 ± 6.56 Aa | 59.31 ± 3.38 Aa |
| MS-1-2 | 60.19 ± 0.73 Aa | 60.83 ± 5.91 Aa | ||
| MS-1-3 | 59.86 ± 8.21 Aa | 61.09 ± 1.79 Aa | ||
| Fe * (mg/kg) | MS-1-1 | 15.70 ± 1.91 a | 14.06 ± 2.25 Aa | 13.45 ± 0.82 Aa |
| MS-1-2 | 15.22 ± 0.79 Aa | 19.45 ± 6.70 Aa | ||
| MS-1-3 | 16.86 ± 1.82 Aab | 12.32 ± 0.60 Aac | ||
| Cu * (mg/kg) | MS-1-1 | 4.65 ± 0.02 a | 4.40 ± 0.09 Aa | 4.49 ± 0.10 Aa |
| MS-1-2 | 4.45 ± 0.09 Aa | 4.54 ± 0.07 Aa | ||
| MS-1-3 | 4.40 ± 0.08 Aa | 4.55 ± 0.04 Aa | ||
| Zn * (mg/kg) | MS-1-1 | 8.06 ± 1.47 a | 8.80 ± 1.83 Aa | 8.44 ± 0.96 Aa |
| MS-1-2 | 8.73 ± 0.80 Aa | 8.19 ± 0.81 Aa | ||
| MS-1-3 | 8.59 ± 1.86 Aa | 8.18 ± 1.00 Aa | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Wang, Q.; Dong, Y.; Mei, J.; Xie, J. Effects of Tricaine Methanesulphonate (MS-222) on Physiological Stress and Fresh Quality of Sea Bass (Lateolabrax maculatus) under Simulated High-Density and Long-Distance Transport Stress. Biology 2023, 12, 223. https://doi.org/10.3390/biology12020223
Zhang H, Wang Q, Dong Y, Mei J, Xie J. Effects of Tricaine Methanesulphonate (MS-222) on Physiological Stress and Fresh Quality of Sea Bass (Lateolabrax maculatus) under Simulated High-Density and Long-Distance Transport Stress. Biology. 2023; 12(2):223. https://doi.org/10.3390/biology12020223
Chicago/Turabian StyleZhang, Hongzhi, Qi Wang, Yixuan Dong, Jun Mei, and Jing Xie. 2023. "Effects of Tricaine Methanesulphonate (MS-222) on Physiological Stress and Fresh Quality of Sea Bass (Lateolabrax maculatus) under Simulated High-Density and Long-Distance Transport Stress" Biology 12, no. 2: 223. https://doi.org/10.3390/biology12020223