
Molecular Actors of Inflammation and Their Signaling Pathways: Mechanistic Insights from Zebrafish
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Cellular Components of Inflammation in Zebrafish

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tool Name and Application | Target Cells | Description | References |
|---|---|---|---|
| Immune cell visualization (transgenic lines) | |||
| Tg(mpeg1:Gal4-VP16), Tg(mpeg1:GFP-caax), Tg(mpeg1:mCherry-F), Tg(mpeg1:GFP) | Macrophages | Reporter lines, mpeg1 promoter | [36,37,49] |
| Tg(mfap4:tdTomato), Tg(mfap4:mCherry-F) | Macrophages | Reporter lines, mfap4 promoter | [35,38] |
| Tg(tnfα:GFP-F) | Cells expressing TNFα | Reporter line, tnfα promoter | [50] |
| Tg(mpeg1:mCherry-F/tnfα:GFP-F) | M1-like macrophages | All macrophages are labelled with mCherry, pro-inflammatory macrophages are also labelled with GFP | [50] |
| Tg(il1β:eGFP-F), TgBAC(il1β:eGFP) | Cells expressing IL1β | Reporter lines, il1β promoter | [49,51,52] |
| Tg(mfap4:mCherry-F/tnfα:GFP-F) | M1 macrophages | All macrophages are labelled with mCherry, pro-inflammatory macrophages are also labelled with GFP | [53] |
| Tg(irg1:eGFP) | M1 macrophages | Activated macrophages are labelled with GFP | [54] |
| Tg(lysC:dsRed), Tg(lysC:eGFP) | Neutrophils | Reporter lines, lysC promoter | [40] |
| Tg(mpx:eGFP) | Neutrophils | Reporter line, mpx promoter | [39] |
| Tg(pu.1:GFP) | Myeloid cells | Reporter line, pu.1 promoter | [41] |
| Tg(coro1a:eGFP) | Macrophages, Neutrophils | Reporter line, coro1a promoter | [42] |
| Tg(lck:eGFP) | T cells | Reporter line, lck promoter | [46] |
| Tg(IgM1:eGFP) | B cells | Reporter line, IgM1 promoter | [47] |
| Tg(cd79:GFP) | B cells | Reporter line, cd79 promoter | [44] |
| Tg(krt4:nlsEGFP) | Superficial skin cells | Reporter line, krt4 promoter | [55] |
| Immune cell depletion system | |||
| Tg(mpeg1:Gal4/UAS-nfsb:mCherry) | Macrophages | Nitroreductase/MTZ treatment | |
| Tg(mpx:Gal4/UAS-nfsb:mCherry) | Neutrophils | Nitroreductase/MTZ treatment | [56] |
| Tg(lysC:Gal4/UAS-nfsb:mCherry) | Neutrophils | Nitroreductase/MTZ treatment | [57] |
| Clodronate liposomes | Macrophages | Liposome-mediated depletion | [36,58] |
| MO-csf3r | Neutrophils | Morpholino-based depletion | [37,59] |
| MO-irf8 | Macrophages | Morpholino-based depletion | [60] |
| Immune cell enrichment system | |||
| MO-irf8 | Neutrophils | irf8 knock-down leads to macrophage depletion and neutrophil enrichment | [60] |
| mRNA-irf8 | Macrophages | Macrophage enrichment through overexpression of irf8 | [60] |
| mRNA-gcsfa/b (csf3a/b) | Neutrophils | Neutrophil enrichment through overexpression of gcsfa or gcsfb | [61] |
| pu.1G242D mutant | Neutrophils | Suppression of pu.1 triggers neutrophil expansion | [62,63] |
3. Molecular Mediators of Inflammation in Zebrafish
3.1. Small Molecules and Free Radicals
3.2. Complement
3.3. Alarmins
3.3.1. S100 Proteins
3.3.2. HMGB1
3.3.3. NCAMP-1
3.3.4. HSPs
3.3.5. Cationic Antimicrobial Peptides (CAMPs)
3.4. Cytokines (Lymphotoxins and Interleukins)
3.5. Chemokines
3.6. Interferons
3.7. Inflammasomes
3.8. Proteases
3.9. Lipids
3.10. Other Pro-Resolving Mediators
4. Molecular Mechanisms and Signaling Pathways Governed by Inflammatory Mediators in Zebrafish
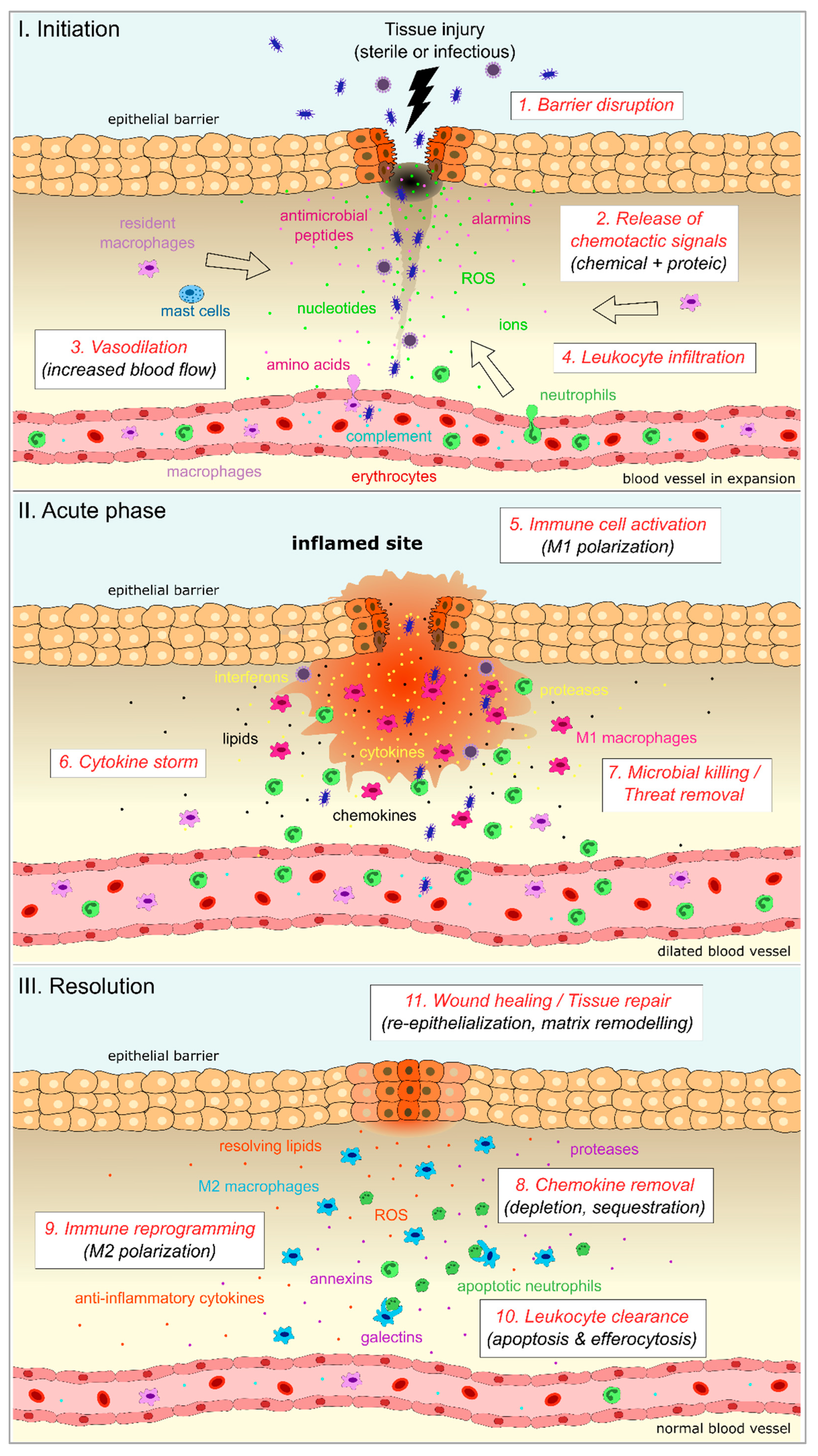
4.1. Induction of Inflammation


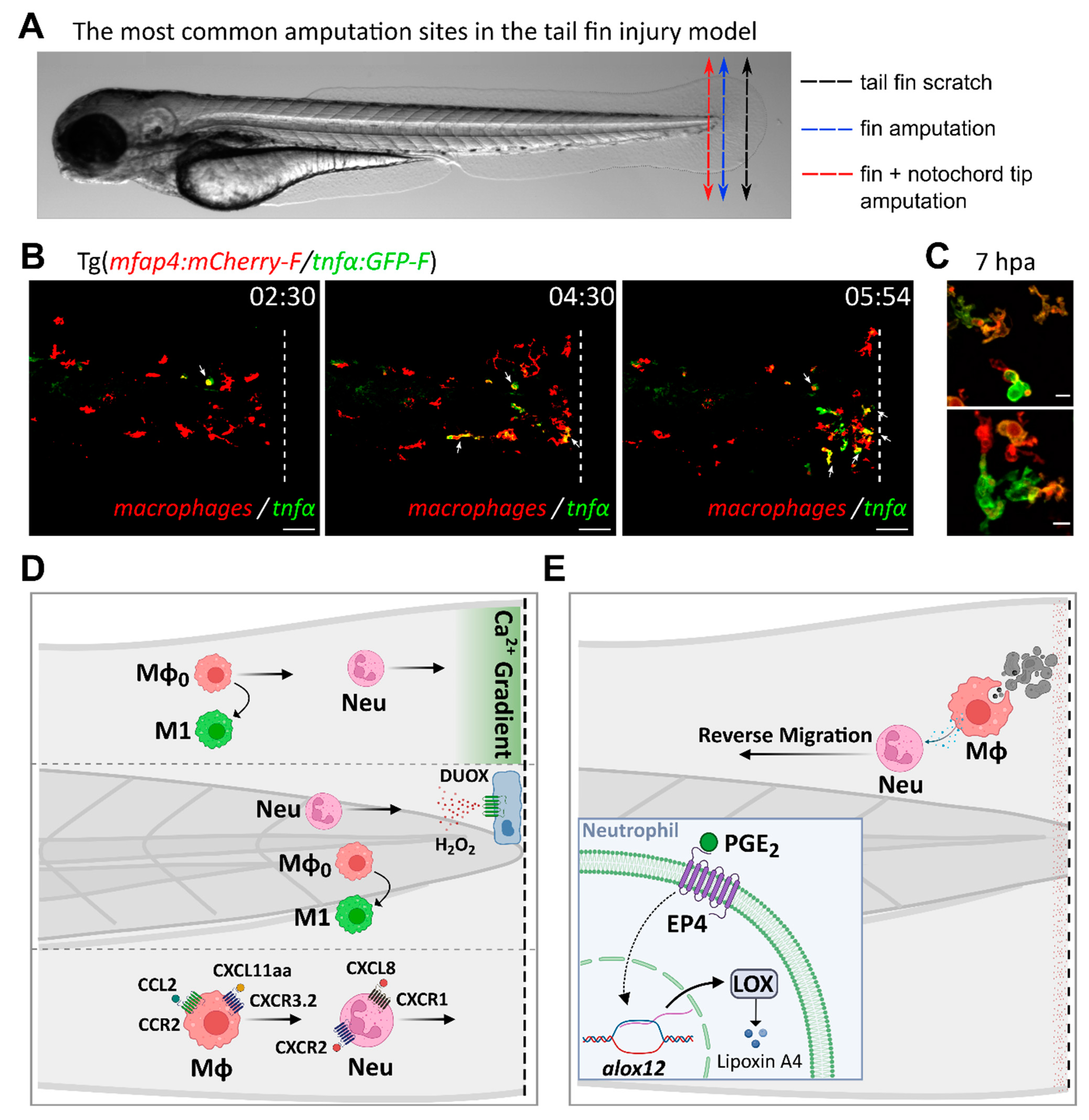
4.2. Neutrophil Migration and Activation
4.3. Macrophage Recruitment and Polarization
4.4. Resolution of Inflammation
4.5. Regeneration
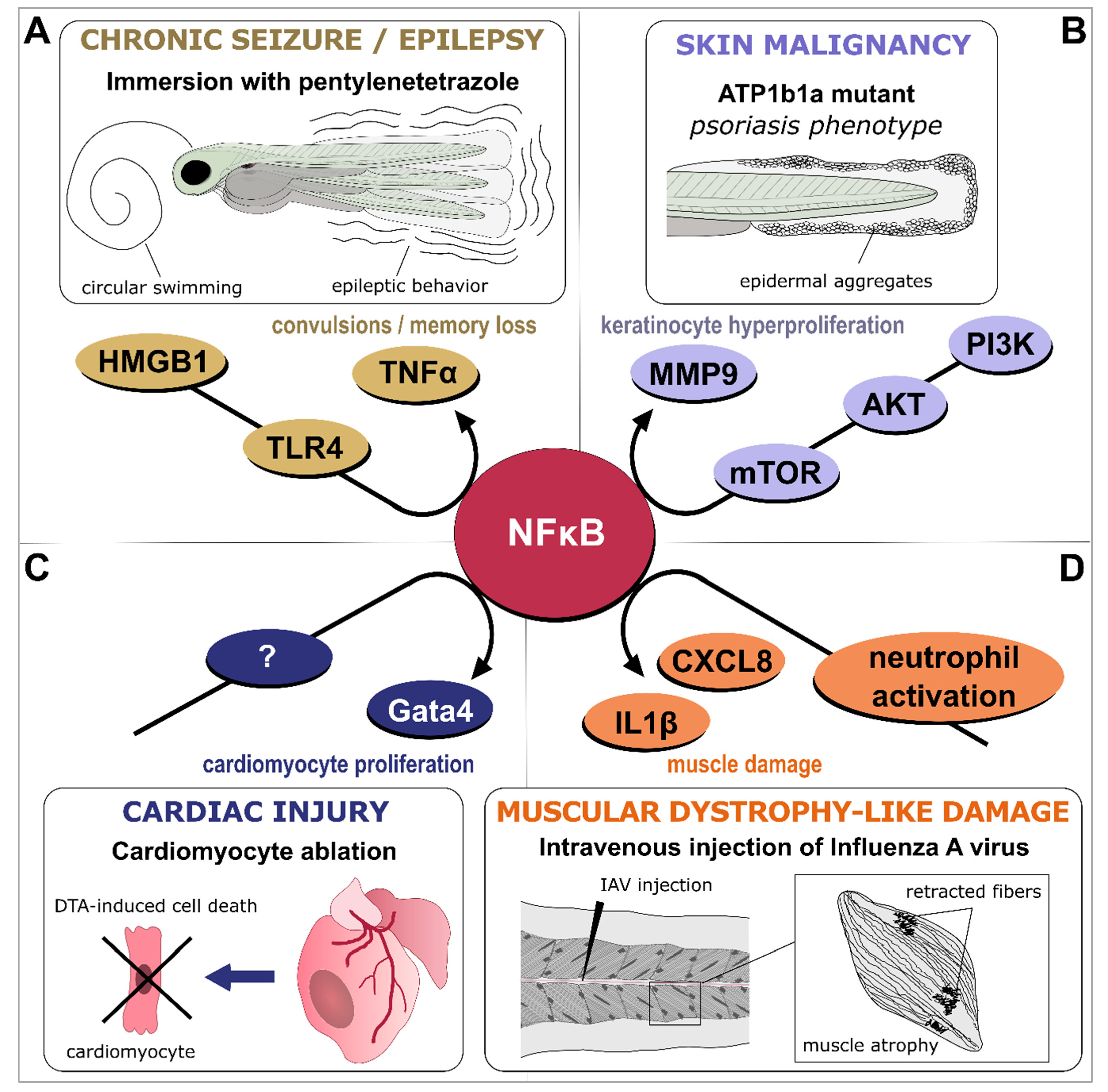
4.6. Infection-Driven Inflammation

5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Medzhitov, R. Origin and Physiological Roles of Inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Nunes, A.C.F. (Ed.) Introductory Chapter: Overview of the Cellular and Molecular Basis of Inflammatory Process. In Translational Studies on Inflammation; IntechOpen: London, UK, 2020; ISBN 978-1-78984-357-6. [Google Scholar]
- Netea, M.G.; Balkwill, F.; Chonchol, M.; Cominelli, F.; Donath, M.Y.; Giamarellos-Bourboulis, E.J.; Golenbock, D.; Gresnigt, M.S.; Heneka, M.T.; Hoffman, H.M.; et al. A Guiding Map for Inflammation. Nat. Immunol. 2017, 18, 826–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvão, I.; Sugimoto, M.A.; Vago, J.P.; Machado, M.G.; Sousa, L.P. Mediators of Inflammation. In Immunopharmacology and Inflammation; Riccardi, C., Levi-Schaffer, F., Tiligada, E., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 3–32. ISBN 978-3-319-77657-6. [Google Scholar]
- Mogensen, T.H. Pathogen Recognition and Inflammatory Signaling in Innate Immune Defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.Y.; Nuñez, G. Sterile Inflammation: Sensing and Reacting to Damage. Nat. Rev. Immunol. 2010, 10, 826–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedl, P.; Weigelin, B. Interstitial Leukocyte Migration and Immune Function. Nat. Immunol. 2008, 9, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.H.Y.; Heit, B.; Kubes, P. Molecular Regulators of Leucocyte Chemotaxis during Inflammation. Cardiovasc. Res. 2010, 86, 183–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luster, A.D.; Alon, R.; von Andrian, U.H. Immune Cell Migration in Inflammation: Present and Future Therapeutic Targets. Nat. Immunol. 2005, 6, 1182–1190. [Google Scholar] [CrossRef]
- Kohl, J. The Role of Complement in Danger Sensing and Transmission. Immunol. Res. 2006, 34, 157–176. [Google Scholar] [CrossRef]
- Gong, T.; Liu, L.; Jiang, W.; Zhou, R. DAMP-Sensing Receptors in Sterile Inflammation and Inflammatory Diseases. Nat. Rev. Immunol. 2020, 20, 95–112. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern Recognition Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef]
- Klos, A.; Tenner, A.J.; Johswich, K.-O.; Ager, R.R.; Reis, E.S.; Köhl, J. The Role of the Anaphylatoxins in Health and Disease. Mol. Immunol. 2009, 46, 2753–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neurath, M.F. Resolution of Inflammation: From Basic Concepts to Clinical Application. Semin. Immunopathol. 2019, 41, 627–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, M.A.; Sousa, L.P.; Pinho, V.; Perretti, M.; Teixeira, M.M. Resolution of Inflammation: What Controls Its Onset? Front. Immunol. 2016, 7, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandri, A.L.; Sousa, L.P.; Lucas, C.D.; Rossi, A.G.; Pinho, V.; Teixeira, M.M. Resolution of Inflammation: Mechanisms and Opportunity for Drug Development. Pharmacol. Ther. 2013, 139, 189–212. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The Zebrafish Reference Genome Sequence and Its Relationship to the Human Genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Postlethwait, J.H.; Yan, Y.-L.; Gates, M.A.; Horne, S.; Amores, A.; Brownlie, A.; Donovan, A.; Egan, E.S.; Force, A.; Gong, Z.; et al. Vertebrate Genome Evolution and the Zebrafish Gene Map. Nat. Genet. 1998, 18, 345–349. [Google Scholar] [CrossRef]
- Rafferty, S.A.; Quinn, T.A. A Beginner’s Guide to Understanding and Implementing the Genetic Modification of Zebrafish. Prog. Biophys. Mol. Biol. 2018, 138, 3–19. [Google Scholar] [CrossRef]
- Wiley, D.S.; Redfield, S.E.; Zon, L.I. Chemical Screening in Zebrafish for Novel Biological and Therapeutic Discovery. In Methods in Cell Biology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 138, pp. 651–679. ISBN 978-0-12-803473-6. [Google Scholar]
- Cavone, L.; McCann, T.; Drake, L.K.; Aguzzi, E.A.; Oprişoreanu, A.-M.; Pedersen, E.; Sandi, S.; Selvarajah, J.; Tsarouchas, T.M.; Wehner, D.; et al. A Unique Macrophage Subpopulation Signals Directly to Progenitor Cells to Promote Regenerative Neurogenesis in the Zebrafish Spinal Cord. Dev. Cell 2021, 56, 1617–1630.e6. [Google Scholar] [CrossRef]
- Buchan, K.D.; van Gent, M.; Prajsnar, T.K.; Ogryzko, N.V.; de Jong, N.W.M.; Kolata, J.; Foster, S.J.; van Strijp, J.A.G.; Renshaw, S.A. Human-Specific Staphylococcal Virulence Factors Enhance Pathogenicity in a Humanised Zebrafish C5a Receptor Model. J. Cell Sci. 2021, 134, jcs252205. [Google Scholar] [CrossRef]
- Choi, T.-Y.; Choi, T.-I.; Lee, Y.-R.; Choe, S.-K.; Kim, C.-H. Zebrafish as an Animal Model for Biomedical Research. Exp. Mol. Med. 2021, 53, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Meijer, A.H.; Schaaf, M.J.M. Modeling Inflammation in Zebrafish for the Development of Anti-Inflammatory Drugs. Front. Cell Dev. Biol. 2021, 8, 620984. [Google Scholar] [CrossRef] [PubMed]
- Torraca, V.; Mostowy, S. Zebrafish Infection: From Pathogenesis to Cell Biology. Trends Cell Biol. 2018, 28, 143–156. [Google Scholar] [CrossRef] [Green Version]
- Kierdorf, K.; Dionne, M.S. The Software and Hardware of Macrophages: A Diversity of Options. Dev. Cell 2016, 38, 122–125. [Google Scholar] [CrossRef] [Green Version]
- Herbomel, P.; Thisse, B.; Thisse, C. Ontogeny and Behaviour of Early Macrophages in the Zebrafish Embryo. Development 1999, 126, 3735–3745. [Google Scholar] [CrossRef] [PubMed]
- Herbomel, P.; Thisse, B.; Thisse, C. Zebrafish Early Macrophages Colonize Cephalic Mesenchyme and Developing Brain, Retina, and Epidermis through a M-CSF Receptor-Dependent Invasive Process. Dev. Biol. 2001, 238, 274–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oosterhof, N.; Kuil, L.E.; van der Linde, H.C.; Burm, S.M.; Berdowski, W.; van Ijcken, W.F.J.; van Swieten, J.C.; Hol, E.M.; Verheijen, M.H.G.; van Ham, T.J. Colony-Stimulating Factor 1 Receptor (CSF1R) Regulates Microglia Density and Distribution, but Not Microglia Differentiation In Vivo. Cell Rep. 2018, 24, 1203–1217.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, J.Y.; Kim, A.D.; Violette, E.P.; Stachura, D.L.; Cisson, J.L.; Traver, D. Definitive Hematopoiesis Initiates through a Committed Erythromyeloid Progenitor in the Zebrafish Embryo. Development 2007, 134, 4147–4156. [Google Scholar] [CrossRef] [Green Version]
- Murayama, E.; Kissa, K.; Zapata, A.; Mordelet, E.; Briolat, V.; Lin, H.-F.; Handin, R.I.; Herbomel, P. Tracing Hematopoietic Precursor Migration to Successive Hematopoietic Organs during Zebrafish Development. Immunity 2006, 25, 963–975. [Google Scholar] [CrossRef] [Green Version]
- Lieschke, G.J.; Oates, A.C.; Crowhurst, M.O.; Ward, A.C.; Layton, J.E. Morphologic and Functional Characterization of Granulocytes and Macrophages in Embryonic and Adult Zebrafish. Blood 2001, 98, 3087–3096. [Google Scholar] [CrossRef]
- Le Guyader, D.; Redd, M.J.; Colucci-Guyon, E.; Murayama, E.; Kissa, K.; Briolat, V.; Mordelet, E.; Zapata, A.; Shinomiya, H.; Herbomel, P. Origins and Unconventional Behavior of Neutrophils in Developing Zebrafish. Blood 2008, 111, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, Q.T.; Sipka, T.; Gonzalez, C.; Levraud, J.-P.; Lutfalla, G.; Nguyen-Chi, M. Neutrophils Use Superoxide to Control Bacterial Infection at a Distance. PLoS Pathog. 2018, 14, e1007157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernut, A.; Herrmann, J.-L.; Kissa, K.; Dubremetz, J.-F.; Gaillard, J.-L.; Lutfalla, G.; Kremer, L. Mycobacterium Abscessus Cording Prevents Phagocytosis and Promotes Abscess Formation. Proc. Natl. Acad. Sci. USA 2014, 111, E943–E952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellett, F.; Pase, L.; Hayman, J.W.; Andrianopoulos, A.; Lieschke, G.J. Mpeg1 Promoter Transgenes Direct Macrophage-Lineage Expression in Zebrafish. Blood 2011, 117, e49–e56. [Google Scholar] [CrossRef] [Green Version]
- Walton, E.M.; Cronan, M.R.; Beerman, R.W.; Tobin, D.M. The Macrophage-Specific Promoter Mfap4 Allows Live, Long-Term Analysis of Macrophage Behavior during Mycobacterial Infection in Zebrafish. PLoS ONE 2015, 10, e0138949. [Google Scholar] [CrossRef] [Green Version]
- Renshaw, S.A.; Loynes, C.A.; Trushell, D.M.I.; Elworthy, S.; Ingham, P.W.; Whyte, M.K.B. A Transgenic Zebrafish Model of Neutrophilic Inflammation. Blood 2006, 108, 3976–3978. [Google Scholar] [CrossRef]
- Hall, C.; Flores, M.V.; Storm, T.; Crosier, K.; Crosier, P. The Zebrafish Lysozyme C Promoter Drives Myeloid-Specific Expression in Transgenic Fish. BMC Dev. Biol. 2007, 7, 42. [Google Scholar] [CrossRef] [Green Version]
- Hsu, K.; Traver, D.; Kutok, J.L.; Hagen, A.; Liu, T.-X.; Paw, B.H.; Rhodes, J.; Berman, J.N.; Zon, L.I.; Kanki, J.P.; et al. The Pu.1 Promoter Drives Myeloid Gene Expression in Zebrafish. Blood 2004, 104, 1291–1297. [Google Scholar] [CrossRef]
- Li, L.; Yan, B.; Shi, Y.-Q.; Zhang, W.-Q.; Wen, Z.-L. Live Imaging Reveals Differing Roles of Macrophages and Neutrophils during Zebrafish Tail Fin Regeneration. J. Biol. Chem. 2012, 287, 25353–25360. [Google Scholar] [CrossRef] [Green Version]
- Meijer, A.H.; Spaink, H.P. Host-Pathogen Interactions Made Transparent with the Zebrafish Model. Curr. Drug Targets 2011, 12, 1000–1017. [Google Scholar] [CrossRef]
- Liu, X.; Li, Y.-S.; Shinton, S.A.; Rhodes, J.; Tang, L.; Feng, H.; Jette, C.A.; Look, A.T.; Hayakawa, K.; Hardy, R.R. Zebrafish B Cell Development without a Pre–B Cell Stage, Revealed by CD79 Fluorescence Reporter Transgenes. J. Immunol. 2017, 199, 1706–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Xu, J.; Feng, S.; He, S.; Zhao, S.; Zhu, L.; Jin, W.; Dai, Y.; Luo, L.; Qu, J.Y.; et al. The First Wave of T Lymphopoiesis in Zebrafish Arises from Aorta Endothelium Independent of Hematopoietic Stem Cells. J. Exp. Med. 2017, 214, 3347–3360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wiest, D.L. Using the Zebrafish Model to Study T Cell Development. In T-Cell Development; Bosselut, R.S., Vacchio, M., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2016; Volume 1323, pp. 273–292. ISBN 978-1-4939-2808-8. [Google Scholar]
- Page, D.M.; Wittamer, V.; Bertrand, J.Y.; Lewis, K.L.; Pratt, D.N.; Delgado, N.; Schale, S.E.; McGue, C.; Jacobsen, B.H.; Doty, A.; et al. An Evolutionarily Conserved Program of B-Cell Development and Activation in Zebrafish. Blood 2013, 122, e1–e11. [Google Scholar] [CrossRef] [Green Version]
- Hernández, P.P.; Strzelecka, P.M.; Athanasiadis, E.I.; Hall, D.; Robalo, A.F.; Collins, C.M.; Boudinot, P.; Levraud, J.-P.; Cvejic, A. Single-Cell Transcriptional Analysis Reveals ILC-like Cells in Zebrafish. Sci. Immunol. 2018, 3, eaau5265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen-Chi, M.; Phan, Q.T.; Gonzalez, C.; Dubremetz, J.-F.; Levraud, J.-P.; Lutfalla, G. Transient Infection of the Zebrafish Notochord with E. Coli Induces Chronic Inflammation. Dis. Model. Mech. 2014, 7, 871–882. [Google Scholar] [CrossRef] [Green Version]
- Nguyen-Chi, M.; Laplace-Builhe, B.; Travnickova, J.; Luz-Crawford, P.; Tejedor, G.; Phan, Q.T.; Duroux-Richard, I.; Levraud, J.-P.; Kissa, K.; Lutfalla, G.; et al. Identification of Polarized Macrophage Subsets in Zebrafish. eLife 2015, 4, e07288. [Google Scholar] [CrossRef] [PubMed]
- Ogryzko, N.V.; Lewis, A.; Wilson, H.L.; Meijer, A.H.; Renshaw, S.A.; Elks, P.M. Hif-1α-Induced Expression of Il-1β Protects against Mycobacterial Infection in Zebrafish. J. Immunol. 2019, 202, 494–502. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, T.; Hall, C.J.; Crosier, P.S.; Abe, G.; Kawakami, K.; Kudo, A.; Kawakami, A. Transient Inflammatory Response Mediated by Interleukin-1β Is Required for Proper Regeneration in Zebrafish Fin Fold. eLife 2017, 6, e22716. [Google Scholar] [CrossRef]
- Sipka, T.; Peroceschi, R.; Hassan-Abdi, R.; Groß, M.; Ellett, F.; Begon-Pescia, C.; Gonzalez, C.; Lutfalla, G.; Nguyen-Chi, M. Damage-Induced Calcium Signaling and Reactive Oxygen Species Mediate Macrophage Activation in Zebrafish. Front. Immunol. 2021, 12, 636585. [Google Scholar] [CrossRef]
- Sanderson, L.E.; Chien, A.-T.; Astin, J.W.; Crosier, K.E.; Crosier, P.S.; Hall, C.J. An Inducible Transgene Reports Activation of Macrophages in Live Zebrafish Larvae. Dev. Comp. Immunol. 2015, 53, 63–69. [Google Scholar] [CrossRef]
- Chen, C.-F.; Chu, C.-Y.; Chen, T.-H.; Lee, S.-J.; Shen, C.-N.; Hsiao, C.-D. Establishment of a Transgenic Zebrafish Line for Superficial Skin Ablation and Functional Validation of Apoptosis Modulators In Vivo. PLoS ONE 2011, 6, e20654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, A.L.; Holmes, G.R.; Bojarczuk, A.N.; Burgon, J.; Loynes, C.A.; Chimen, M.; Sawtell, A.K.; Hamza, B.; Willson, J.; Walmsley, S.R.; et al. A Zebrafish Compound Screen Reveals Modulation of Neutrophil Reverse Migration as an Anti-Inflammatory Mechanism. Sci. Transl. Med. 2014, 6, 225ra29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elks, P.M.; van Eeden, F.J.; Dixon, G.; Wang, X.; Reyes-Aldasoro, C.C.; Ingham, P.W.; Whyte, M.K.B.; Walmsley, S.R.; Renshaw, S.A. Activation of Hypoxia-Inducible Factor-1α (Hif-1α) Delays Inflammation Resolution by Reducing Neutrophil Apoptosis and Reverse Migration in a Zebrafish Inflammation Model. Blood 2011, 118, 712–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooijen, N.V.; Sanders, A. Liposome Mediated Depletion of Macrophages: Mechanism of Action, Preparation of Liposomes and Applications. J. Immunol. Methods 1994, 174, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Liongue, C.; Hall, C.J.; O’Connell, B.A.; Crosier, P.; Ward, A.C. Zebrafish Granulocyte Colony-Stimulating Factor Receptor Signaling Promotes Myelopoiesis and Myeloid Cell Migration. Blood 2009, 113, 2535–2546. [Google Scholar] [CrossRef]
- Li, L.; Jin, H.; Xu, J.; Shi, Y.; Wen, Z. Irf8 Regulates Macrophage versus Neutrophil Fate during Zebrafish Primitive Myelopoiesis. Blood 2011, 117, 1359–1369. [Google Scholar] [CrossRef] [Green Version]
- Stachura, D.L.; Svoboda, O.; Campbell, C.A.; Espín-Palazón, R.; Lau, R.P.; Zon, L.I.; Bartunek, P.; Traver, D. The Zebrafish Granulocyte Colony-Stimulating Factors (Gcsfs): 2 Paralogous Cytokines and Their Roles in Hematopoietic Development and Maintenance. Blood 2013, 122, 3918–3928. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Li, L.; Xu, J.; Zhen, F.; Zhu, L.; Liu, P.P.; Zhang, M.; Zhang, W.; Wen, Z. Runx1 Regulates Embryonic Myeloid Fate Choice in Zebrafish through a Negative Feedback Loop Inhibiting Pu.1 Expression. Blood 2012, 119, 5239–5249. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Liu, W.; Li, L.; Chen, J.; Wu, M.; Zhang, Y.; Leung, A.Y.H.; Zhang, W.; Wen, Z.; Liao, W. Suppression of Pu.1 Function Results in Expanded Myelopoiesis in Zebrafish. Leukemia 2013, 27, 1913–1917. [Google Scholar] [CrossRef]
- Presland, R.B.; Jurevic, R.J. Making Sense of the Epithelial Barrier: What Molecular Biology and Genetics Tell Us About the Functions of Oral Mucosal and Epidermal Tissues. J. Dent. Educ. 2002, 66, 564–574. [Google Scholar] [CrossRef]
- Andrews, C.; McLean, M.H.; Durum, S.K. Cytokine Tuning of Intestinal Epithelial Function. Front. Immunol. 2018, 9, 1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Neriah, Y.; Schmidt-Supprian, M. Epithelial NF-ΚB Maintains Host Gut Microflora Homeostasis. Nat. Immunol. 2007, 8, 479–481. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.K.; Freisinger, C.M.; LeBert, D.C.; Huttenlocher, A. Early Redox, Src Family Kinase, and Calcium Signaling Integrate Wound Responses and Tissue Regeneration in Zebrafish. J. Cell Biol. 2012, 199, 225–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordeiro, J.V.; Jacinto, A. The Role of Transcription-Independent Damage Signals in the Initiation of Epithelial Wound Healing. Nat. Rev. Mol. Cell Biol. 2013, 14, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, A.M.; Saraiva, L.R.; Korsching, S.I. Structural and Functional Diversification in the Teleost S100 Family of Calcium-Binding Proteins. BMC Evol. Biol. 2008, 8, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gahring, L.C.; Buckley, A.; Daynes, R.A. Presence of Epidermal-Derived Thymocyte Activating Factor/Interleukin 1 in Normal Human Stratum Corneum. J. Clin. Investig. 1985, 76, 1585–1591. [Google Scholar] [CrossRef] [Green Version]
- Kupper, T.S.; Ballard, D.W.; Chua, A.O.; McGuire, J.S.; Flood, P.M.; Horowitz, M.C.; Langdon, R.; Lightfoot, L.; Gubler, U. Human Keratinocytes Contain MRNA Indistinguishable from Monocyte Interleukin 1 Alpha and Beta MRNA. Keratinocyte Epidermal Cell-Derived Thymocyte-Activating Factor Is Identical to Interleukin 1. J. Exp. Med. 1986, 164, 2095–2100. [Google Scholar] [CrossRef] [Green Version]
- Eisenhoffer, G.T.; Slattum, G.; Ruiz, O.E.; Otsuna, H.; Bryan, C.D.; Lopez, J.; Wagner, D.S.; Bonkowsky, J.L.; Chien, C.-B.; Dorsky, R.I.; et al. A Toolbox to Study Epidermal Cell Types in Zebrafish. J. Cell Sci. 2016, 130, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Kanther, M.; Sun, X.; Mühlbauer, M.; Mackey, L.C.; Flynn, E.J.; Bagnat, M.; Jobin, C.; Rawls, J.F. Microbial Colonization Induces Dynamic Temporal and Spatial Patterns of NF-ΚB Activation in the Zebrafish Digestive Tract. Gastroenterology 2011, 141, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Kuri, P.; Schieber, N.L.; Thumberger, T.; Wittbrodt, J.; Schwab, Y.; Leptin, M. Dynamics of in Vivo ASC Speck Formation. J. Cell Biol. 2017, 216, 2891–2909. [Google Scholar] [CrossRef]
- Varela, M.; van der Vaart, M.; Groenewoud, A.; Meijer, A.H. Extracellular Mycobacterial DNA Drives Disease Progression by Triggering Caspase-11-Dependent Pyroptosis of Infected Macrophages. bioRxiv 2019. bioRxiv:514125. [Google Scholar]
- Isles, H.M.; Loynes, C.A.; Alasmari, S.; Kon, F.C.; Henry, K.M.; Kadochnikova, A.; Hales, J.; Muir, C.F.; Keightley, M.-C.; Kadirkamanathan, V.; et al. Pioneer Neutrophils Release Chromatin within in Vivo Swarms. eLife 2021, 10, e68755. [Google Scholar] [CrossRef] [PubMed]
- Palha, N.; Guivel-Benhassine, F.; Briolat, V.; Lutfalla, G.; Sourisseau, M.; Ellett, F.; Wang, C.-H.; Lieschke, G.J.; Herbomel, P.; Schwartz, O.; et al. Real-Time Whole-Body Visualization of Chikungunya Virus Infection and Host Interferon Response in Zebrafish. PLoS Pathog. 2013, 9, e1003619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maarifi, G.; Smith, N.; Maillet, S.; Moncorgé, O.; Chamontin, C.; Edouard, J.; Sohm, F.; Blanchet, F.P.; Herbeuval, J.-P.; Lutfalla, G.; et al. TRIM8 Is Required for Virus-Induced IFN Response in Human Plasmacytoid Dendritic Cells. Sci. Adv. 2019, 5, eaax3511. [Google Scholar] [CrossRef] [Green Version]
- Sieger, D.; Moritz, C.; Ziegenhals, T.; Prykhozhij, S.; Peri, F. Long-Range Ca2+ Waves Transmit Brain-Damage Signals to Microglia. Dev. Cell 2012, 22, 1138–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, M.J.; Pisharath, H.; Yusuff, S.; Moore, J.C.; Siekmann, A.F.; Lawson, N.; Leach, S.D. Notch-Responsive Cells Initiate the Secondary Transition in Larval Zebrafish Pancreas. Mech. Dev. 2009, 126, 898–912. [Google Scholar] [CrossRef] [Green Version]
- Coombs, C.; Georgantzoglou, A.; Walker, H.A.; Patt, J.; Merten, N.; Poplimont, H.; Busch-Nentwich, E.M.; Williams, S.; Kotsi, C.; Kostenis, E.; et al. Chemokine Receptor Trafficking Coordinates Neutrophil Clustering and Dispersal at Wounds in Zebrafish. Nat. Commun. 2019, 10, 5166. [Google Scholar] [CrossRef] [Green Version]
- Torraca, V.; Otto, N.A.; Tavakoli-Tameh, A.; Meijer, A.H. The Inflammatory Chemokine Cxcl18b Exerts Neutrophil-Specific Chemotaxis via the Promiscuous Chemokine Receptor Cxcr2 in Zebrafish. Dev. Comp. Immunol. 2017, 67, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Glass, T.J.; Lund, T.C.; Patrinostro, X.; Tolar, J.; Bowman, T.V.; Zon, L.I.; Blazar, B.R. Stromal Cell–Derived Factor-1 and Hematopoietic Cell Homing in an Adult Zebrafish Model of Hematopoietic Cell Transplantation. Blood 2011, 118, 766–774. [Google Scholar] [CrossRef]
- Gamba, L.; Cubedo, N.; Ghysen, A.; Lutfalla, G.; Dambly-Chaudière, C. Estrogen Receptor ESR1 Controls Cell Migration by Repressing Chemokine Receptor CXCR4 in the Zebrafish Posterior Lateral Line System. Proc. Natl. Acad. Sci. USA 2010, 107, 6358–6363. [Google Scholar] [CrossRef] [Green Version]
- Nguyen-Chi, M.; Laplace-Builhé, B.; Travnickova, J.; Luz-Crawford, P.; Tejedor, G.; Lutfalla, G.; Kissa, K.; Jorgensen, C.; Djouad, F. TNF Signaling and Macrophages Govern Fin Regeneration in Zebrafish Larvae. Cell Death Dis. 2017, 8, e2979. [Google Scholar] [CrossRef] [PubMed]
- Espín, R.; Roca, F.J.; Candel, S.; Sepulcre, M.P.; González-Rosa, J.M.; Alcaraz-Pérez, F.; Meseguer, J.; Cayuela, M.L.; Mercader, N.; Mulero, V. TNF Receptors Regulate Vascular Homeostasis in Zebrafish through a Caspase-8, Caspase-2 and P53 Apoptotic Program That Bypasses Caspase-3. Dis. Model. Mech. 2013, 6, 383–396. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Kuja-Panula, J.; Rouhiainen, A.; Chen, Y.; Panula, P.; Rauvala, H. High Mobility Group Box-1 (HMGB1; Amphoterin) Is Required for Zebrafish Brain Development. J. Biol. Chem. 2011, 286, 23200–23213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itou, J.; Taniguchi, N.; Oishi, I.; Kawakami, H.; Lotz, M.; Kawakami, Y. HMGB Factors Are Required for Posterior Digit Development through Integrating Signaling Pathway Activities. Dev. Dyn. 2011, 240, 1151–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, S.; Reyes-Aldasoro, C.C.; Candel, S.; Renshaw, S.A.; Mulero, V.; Calado, Â. Cxcl8 (IL-8) Mediates Neutrophil Recruitment and Behavior in the Zebrafish Inflammatory Response. J. Immunol. 2013, 190, 4349–4359. [Google Scholar] [CrossRef] [Green Version]
- Sarris, M.; Masson, J.-B.; Maurin, D.; Van der Aa, L.M.; Boudinot, P.; Lortat-Jacob, H.; Herbomel, P. Inflammatory Chemokines Direct and Restrict Leukocyte Migration within Live Tissues as Glycan-Bound Gradients. Curr. Biol. 2012, 22, 2375–2382. [Google Scholar] [CrossRef] [Green Version]
- Powell, D.; Lou, M.; Barros Becker, F.; Huttenlocher, A. Cxcr1 Mediates Recruitment of Neutrophils and Supports Proliferation of Tumor-Initiating Astrocytes in Vivo. Sci. Rep. 2018, 8, 13285. [Google Scholar] [CrossRef] [Green Version]
- Knaut, H.; Werz, C.; Geisler, R.; Nüsslein-Volhard, C. Tübingen 2000 Screen Consortium A Zebrafish Homologue of the Chemokine Receptor Cxcr4 Is a Germ-Cell Guidance Receptor. Nature 2003, 421, 279–282. [Google Scholar] [CrossRef]
- Dambly-Chaudière, C.; Cubedo, N.; Ghysen, A. Control of Cell Migration in the Development of the Posterior Lateral Line: Antagonistic Interactions between the Chemokine Receptors CXCR4 and CXCR7/RDC1. BMC Dev. Biol. 2007, 7, 23. [Google Scholar] [CrossRef] [Green Version]
- Lu, I.-N.; Chiang, B.-L.; Lou, K.-L.; Huang, P.-T.; Yao, C.-C.J.; Wang, J.-S.; Lin, L.-D.; Jeng, J.-H.; Chang, B.-E. Cloning, Expression and Characterization of CCL21 and CCL25 Chemokines in Zebrafish. Dev. Comp. Immunol. 2012, 38, 203–214. [Google Scholar] [CrossRef]
- Tyrkalska, S.D.; Candel, S.; Pérez-Oliva, A.B.; Valera, A.; Alcaraz-Pérez, F.; García-Moreno, D.; Cayuela, M.L.; Mulero, V. Identification of an Evolutionarily Conserved Ankyrin Domain-Containing Protein, Caiap, Which Regulates Inflammasome-Dependent Resistance to Bacterial Infection. Front. Immunol. 2017, 8, 1375. [Google Scholar] [CrossRef] [PubMed]
- Vincent, W.J.B.; Freisinger, C.M.; Lam, P.-Y.; Huttenlocher, A.; Sauer, J.-D. Macrophages Mediate Flagellin Induced Inflammasome Activation and Host Defense in Zebrafish. Cell. Microbiol. 2016, 18, 591–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masumoto, J.; Zhou, W.; Chen, F.F.; Su, F.; Kuwada, J.Y.; Hidaka, E.; Katsuyama, T.; Sagara, J.; Taniguchi, S.; Ngo-Hazelett, P.; et al. Caspy, a Zebrafish Caspase, Activated by ASC Oligomerization Is Required for Pharyngeal Arch Development. J. Biol. Chem. 2003, 278, 4268–4276. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Zheng, X.; Chen, S.; Wang, Z.; Xu, W.; Tan, J.; Hu, T.; Hou, M.; Wang, W.; Gu, Z.; et al. Sensing of Cytosolic LPS through Caspy2 Pyrin Domain Mediates Noncanonical Inflammasome Activation in Zebrafish. Nat. Commun. 2018, 9, 3052. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Yan, B.; Lou, L.; Lin, X.; Yu, T.; Wu, S.; Lu, Q.; Liu, W.; Huang, Z.; Zhang, M.; et al. Nlrc3-like Is Required for Microglia Maintenance in Zebrafish. J. Genet. Genom. 2019, 46, 291–299. [Google Scholar] [CrossRef]
- Oehlers, S.H.; Flores, M.V.; Hall, C.J.; Swift, S.; Crosier, K.E.; Crosier, P.S. The Inflammatory Bowel Disease (IBD) Susceptibility Genes NOD1 and NOD2 Have Conserved Anti-Bacterial Roles in Zebrafish. Dis. Model. Mech. 2011, 4, 832–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hugot, J.-P.; Chamaillard, M.; Zouali, H.; Lesage, S.; Cézard, J.-P.; Belaiche, J.; Almer, S.; Tysk, C.; O’Morain, C.A.; Gassull, M.; et al. Association of NOD2 Leucine-Rich Repeat Variants with Susceptibility to Crohn’s Disease. Nature 2001, 411, 599–603. [Google Scholar] [CrossRef]
- Forn-Cuní, G.; Reis, E.S.; Dios, S.; Posada, D.; Lambris, J.D.; Figueras, A.; Novoa, B. The Evolution and Appearance of C3 Duplications in Fish Originate an Exclusive Teleost C3 Gene Form with Anti-Inflammatory Activity. PLoS ONE 2014, 9, e99673. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Fontaine, C.; Theveneau, E.; Tzekou, A.; Tada, M.; Woods, M.; Page, K.M.; Parsons, M.; Lambris, J.D.; Mayor, R. Complement Fragment C3a Controls Mutual Cell Attraction during Collective Cell Migration. Dev. Cell 2011, 21, 1026–1037. [Google Scholar] [CrossRef] [Green Version]
- Choksi, S.P.; Babu, D.; Lau, D.; Yu, X.; Roy, S. Systematic Discovery of Novel Ciliary Genes through Functional Genomics in the Zebrafish. Development 2014, 141, 3410–3419. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; van Steijn, L.; Li, C.; Verbeek, F.J.; Cao, L.; Merks, R.M.H.; Spaink, H.P. A Novel Function of TLR2 and MyD88 in the Regulation of Leukocyte Cell Migration Behavior During Wounding in Zebrafish Larvae. Front. Cell Dev. Biol. 2021, 9, 624571. [Google Scholar] [CrossRef] [PubMed]
- Bottiglione, F.; Dee, C.T.; Lea, R.; Zeef, L.A.H.; Badrock, A.P.; Wane, M.; Bugeon, L.; Dallman, M.J.; Allen, J.E.; Hurlstone, A.F.L. Zebrafish IL-4-like Cytokines and IL-10 Suppress Inflammation but Only IL-10 Is Essential for Gill Homeostasis. J. Immunol. 2020, 205, 994–1008. [Google Scholar] [CrossRef] [PubMed]
- Darvish, H.; Azcona, L.J.; Taghavi, S.; Firouzabadi, S.G.; Tafakhori, A.; Alehabib, E.; Mohajerani, F.; Zardadi, S.; Paisán-Ruiz, C. ANXA1 with Anti-Inflammatory Properties Might Contribute to Parkinsonism. Ann. Neurol. 2021, 90, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Matty, M.A.; Knudsen, D.R.; Walton, E.M.; Beerman, R.W.; Cronan, M.R.; Pyle, C.J.; Hernandez, R.E.; Tobin, D.M. Potentiation of P2RX7 as a Host-Directed Strategy for Control of Mycobacterial Infection. eLife 2019, 8, e39123. [Google Scholar] [CrossRef]
- Kenyon, A.; Gavriouchkina, D.; Zorman, J.; Napolitani, G.; Cerundolo, V.; Sauka-Spengler, T. Active Nuclear Transcriptome Analysis Reveals Inflammasome-Dependent Mechanism for Early Neutrophil Response to Mycobacterium Marinum. Sci. Rep. 2017, 7, 6505. [Google Scholar] [CrossRef] [Green Version]
- Charlton-Perkins, M.; Almeida, A.D.; MacDonald, R.B.; Harris, W.A. Genetic Control of Cellular Morphogenesis in Müller Glia. Glia 2019, 67, 1401–1411. [Google Scholar] [CrossRef] [Green Version]
- Aggad, D.; Mazel, M.; Boudinot, P.; Mogensen, K.E.; Hamming, O.J.; Hartmann, R.; Kotenko, S.; Herbomel, P.; Lutfalla, G.; Levraud, J.-P. The Two Groups of Zebrafish Virus-Induced Interferons Signal via Distinct Receptors with Specific and Shared Chains. J. Immunol. 2009, 183, 3924–3931. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, S.; López-Muñoz, A.; Candel, S.; Pelegrín, P.; Calado, Â.; Mulero, V. ATP Modulates Acute Inflammation In Vivo through Dual Oxidase 1–Derived H2O2 Production and NF-KB Activation. J. Immunol. 2014, 192, 5710–5719. [Google Scholar] [CrossRef] [Green Version]
- Niethammer, P.; Grabher, C.; Look, A.T.; Mitchison, T.J. A Tissue-Scale Gradient of Hydrogen Peroxide Mediates Rapid Wound Detection in Zebrafish. Nature 2009, 459, 996–999. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.K.; Starnes, T.W.; Deng, Q.; Huttenlocher, A. Lyn Is a Redox Sensor That Mediates Leukocyte Wound Attraction in Vivo. Nature 2011, 480, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Niethammer, P. The Early Wound Signals. Curr. Opin. Genet. Dev. 2016, 40, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Razzell, W.; Evans, I.R.; Martin, P.; Wood, W. Calcium Flashes Orchestrate the Wound Inflammatory Response through DUOX Activation and Hydrogen Peroxide Release. Curr. Biol. 2013, 23, 424–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, N.R.; Chen, Y.; Ishibashi, S.; Kritsiligkou, P.; Lea, R.; Koh, Y.; Gallop, J.L.; Dorey, K.; Amaya, E. Amputation-Induced Reactive Oxygen Species Are Required for Successful Xenopus Tadpole Tail Regeneration. Nat. Cell Biol. 2013, 15, 222–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klyubin, I.V.; Kirpichnikova, K.M.; Gamaley, I.A. Hydrogen Peroxide-Induced Chemotaxis of Mouse Peritoneal Neutrophils. Eur. J. Cell Biol. 1996, 70, 347–351. [Google Scholar] [PubMed]
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part I—Molecular Mechanisms of Activation and Regulation. Front. Immunol. 2015, 6, 262. [Google Scholar] [CrossRef] [Green Version]
- Lo, M.W.; Woodruff, T.M. Complement: Bridging the Innate and Adaptive Immune Systems in Sterile Inflammation. J. Leukoc. Biol. 2020, 108, 339–351. [Google Scholar] [CrossRef]
- Zhang, S.; Cui, P. Complement System in Zebrafish. Dev. Comp. Immunol. 2014, 46, 3–10. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, S.; Wang, G.; An, Y. Complement Activity in the Egg Cytosol of Zebrafish Danio Rerio: Evidence for the Defense Role of Maternal Complement Components. PLoS ONE 2008, 3, e1463. [Google Scholar] [CrossRef] [Green Version]
- Markiewski, M.M.; Lambris, J.D. The Role of Complement in Inflammatory Diseases From Behind the Scenes into the Spotlight. Am. J. Pathol. 2007, 171, 715–727. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Han, Z.; Oppenheim, J.J. Alarmins and Immunity. Immunol. Rev. 2017, 280, 41–56. [Google Scholar] [CrossRef]
- Chan, J.K.; Roth, J.; Oppenheim, J.J.; Tracey, K.J.; Vogl, T.; Feldmann, M.; Horwood, N.; Nanchahal, J. Alarmins: Awaiting a Clinical Response. J. Clin. Investig. 2012, 122, 2711–2719. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, L.L.; Garrie, K.; Turner, M.D. Role of S100 Proteins in Health and Disease. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118677. [Google Scholar] [CrossRef] [PubMed]
- Donato, R.; Cannon, B.R.; Sorci, G.; Riuzzi, F.; Hsu, K.; Weber, D.J.; Geczy, C.L. Functions of S100 Proteins. Curr. Mol. Med. 2013, 13, 24–57. [Google Scholar] [CrossRef] [Green Version]
- Halawi, A.; Abbas, O.; Mahalingam, M. S100 Proteins and the Skin: A Review. J. Eur. Acad. Derm. Venereol. 2014, 28, 405–414. [Google Scholar] [CrossRef]
- Leśniak, W.; Graczyk-Jarzynka, A. The S100 Proteins in Epidermis: Topology and Function. Biochim. Biophys. Acta Gen. Subj. 2015, 1850, 2563–2572. [Google Scholar] [CrossRef] [PubMed]
- Sreejit, G.; Flynn, M.C.; Patil, M.; Krishnamurthy, P.; Murphy, A.J.; Nagareddy, P.R. S100 Family Proteins in Inflammation and Beyond. In Advances in Clinical Chemistry; Elsevier: Amsterdam, The Netherlands, 2020; Volume 98, pp. 173–231. ISBN 978-0-12-821558-6. [Google Scholar]
- Meijer, B.; Gearry, R.B.; Day, A.S. The Role of S100A12 as a Systemic Marker of Inflammation. Int. J. Inflamm. 2012, 2012, 907078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogl, T.; Eisenblätter, M.; Völler, T.; Zenker, S.; Hermann, S.; van Lent, P.; Faust, A.; Geyer, C.; Petersen, B.; Roebrock, K.; et al. Alarmin S100A8/S100A9 as a Biomarker for Molecular Imaging of Local Inflammatory Activity. Nat. Commun. 2014, 5, 4593. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, D.B.; Eubanks, J.O.; Ramakrishnan, D.; Criscitiello, M.F. Evolution of the S100 Family of Calcium Sensor Proteins. Cell Calcium 2013, 53, 170–179. [Google Scholar] [CrossRef]
- Speir, M.L.; Bhaduri, A.; Markov, N.S.; Moreno, P.; Nowakowski, T.J.; Papatheodorou, I.; Pollen, A.A.; Raney, B.J.; Seninge, L.; Kent, W.J.; et al. UCSC Cell Browser: Visualize Your Single-Cell Data. Bioinformatics 2021, 37, 4578–4580. [Google Scholar] [CrossRef]
- Yang, H.; Wang, H.; Andersson, U. Targeting Inflammation Driven by HMGB1. Front. Immunol. 2020, 11, 484. [Google Scholar] [CrossRef] [Green Version]
- Sha, Y.; Zmijewski, J.; Xu, Z.; Abraham, E. HMGB1 Develops Enhanced Proinflammatory Activity by Binding to Cytokines. J. Immunol. 2008, 180, 2531–2537. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Avalos, A.M.; Mao, S.-Y.; Chen, B.; Senthil, K.; Wu, H.; Parroche, P.; Drabic, S.; Golenbock, D.; Sirois, C.; et al. Toll-like Receptor 9–Dependent Activation by DNA-Containing Immune Complexes Is Mediated by HMGB1 and RAGE. Nat. Immunol. 2007, 8, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.E. HMGB1 Loves Company. J. Leukoc. Biol. 2009, 86, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Scaffidi, P.; Misteli, T.; Bianchi, M.E. Release of Chromatin Protein HMGB1 by Necrotic Cells Triggers Inflammation. Nature 2002, 418, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Schiraldi, M.; Raucci, A.; Muñoz, L.M.; Livoti, E.; Celona, B.; Venereau, E.; Apuzzo, T.; De Marchis, F.; Pedotti, M.; Bachi, A.; et al. HMGB1 Promotes Recruitment of Inflammatory Cells to Damaged Tissues by Forming a Complex with CXCL12 and Signaling via CXCR4. J. Exp. Med. 2012, 209, 551–563. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Tewary, P.; de la Rosa, G.; Wei, F.; Oppenheim, J.J. The Alarmin Functions of High-Mobility Group Proteins. Biochim. Biophys. Acta Gene Regul. Mech. 2010, 1799, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lu, Y.-Q. The Regulatory Role of High-Mobility Group Protein 1 in Sepsis-Related Immunity. Front. Immunol. 2021, 11, 601815. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wang, J.; Park, Y.-J.; Tsuruta, Y.; Lorne, E.F.; Zhao, X.; Abraham, E. High Mobility Group Protein-1 Inhibits Phagocytosis of Apoptotic Neutrophils through Binding to Phosphatidylserine. J. Immunol. 2008, 181, 4240–4246. [Google Scholar] [CrossRef] [Green Version]
- Venereau, E.; Casalgrandi, M.; Schiraldi, M.; Antoine, D.J.; Cattaneo, A.; De Marchis, F.; Liu, J.; Antonelli, A.; Preti, A.; Raeli, L.; et al. Mutually Exclusive Redox Forms of HMGB1 Promote Cell Recruitment or Proinflammatory Cytokine Release. J. Exp. Med. 2012, 209, 1519–1528. [Google Scholar] [CrossRef] [Green Version]
- Tirone, M.; Tran, N.L.; Ceriotti, C.; Gorzanelli, A.; Canepari, M.; Bottinelli, R.; Raucci, A.; Di Maggio, S.; Santiago, C.; Mellado, M.; et al. High Mobility Group Box 1 Orchestrates Tissue Regeneration via CXCR4. J. Exp. Med. 2018, 215, 303–318. [Google Scholar] [CrossRef]
- Moleri, S.; Cappellano, G.; Gaudenzi, G.; Cermenati, S.; Cotelli, F.; Horner, D.S.; Beltrame, M. The HMGB Protein Gene Family in Zebrafish: Evolution and Embryonic Expression Patterns. Gene Expr. Patterns 2011, 11, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.L.; Kaur, H.; Leary III, J.; Praveen, K.; Jaso-Friedmann, L. Molecular Characterization of a Novel Pattern Recognition Protein from Nonspecific Cytotoxic Cells: Sequence Analysis, Phylogenetic Comparisons and Anti-Microbial Activity of a Recombinant Homologue. Dev. Comp. Immunol. 2005, 29, 1049–1064. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.L.; Connor, M.A.; Moss, L.D.; Lackay, S.; Leary, J.H.; Krunkosky, T.; Jaso-Friedmann, L. Cellular Expression and Antimicrobial Function of a Phylogenetically Conserved Novel Histone 1x-like Protein on Mouse Cells: A Potential New Class of Pattern Recognition Receptor. J. Leukoc. Biol. 2009, 86, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Monette, M.M.; Evans, D.L.; Krunkosky, T.; Camus, A.; Jaso-Friedmann, L. Nonspecific Cytotoxic Cell Antimicrobial Protein (NCAMP-1): A Novel Alarmin Ligand Identified in Zebrafish. PLoS ONE 2015, 10, e0116576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaso-Friedmann, L.; Leary, J.H.; Camus, A.C.; Evans, D.L. The Teleost Acute-Phase Inflammatory Response and Caspase Activation by a Novel Alarmin-like Ligand. J. Leukoc. Biol. 2014, 95, 785–796. [Google Scholar] [CrossRef]
- Binder, R.J. Functions of Heat Shock Proteins in Pathways of the Innate and Adaptive Immune System. J. Immunol. 2014, 193, 5765–5771. [Google Scholar] [CrossRef] [Green Version]
- Calderwood, S.K.; Gong, J.; Murshid, A. Extracellular HSPs: The Complicated Roles of Extracellular HSPs in Immunity. Front. Immunol. 2016, 7, 159. [Google Scholar] [CrossRef] [Green Version]
- Asea, A.; Kraeft, S.-K.; Kurt-Jones, E.A.; Stevenson, M.A.; Chen, L.B.; Finberg, R.W.; Koo, G.C.; Calderwood, S.K. HSP70 Stimulates Cytokine Production through a CD14-Dependant Pathway, Demonstrating Its Dual Role as a Chaperone and Cytokine. Nat. Med. 2000, 6, 435–442. [Google Scholar] [CrossRef]
- Salari, S.; Seibert, T.; Chen, Y.-X.; Hu, T.; Shi, C.; Zhao, X.; Cuerrier, C.M.; Raizman, J.E.; O’Brien, E.R. Extracellular HSP27 Acts as a Signaling Molecule to Activate NF-ΚB in Macrophages. Cell Stress Chaperones 2013, 18, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Murshid, A.; Gong, J.; Prince, T.; Borges, T.J.; Calderwood, S.K. Scavenger Receptor SREC-I Mediated Entry of TLR4 into Lipid Microdomains and Triggered Inflammatory Cytokine Release in RAW 264.7 Cells upon LPS Activation. PLoS ONE 2015, 10, e0122529. [Google Scholar] [CrossRef]
- Murshid, A.; Gong, J.; Calderwood, S.K. The Role of Heat Shock Proteins in Antigen Cross Presentation. Front. Immun. 2012, 3, 63. [Google Scholar] [CrossRef] [PubMed]
- Van Eden, W.; Spiering, R.; Broere, F.; van der Zee, R. A Case of Mistaken Identity: HSPs Are No DAMPs but DAMPERs. Cell Stress Chaperones 2012, 17, 281–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Eden, W.; van der Zee, R.; Prakken, B. Heat-Shock Proteins Induce T-Cell Regulation of Chronic Inflammation. Nat. Rev. Immunol. 2005, 5, 318–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moudgil, K.D.; Chang, T.T.; Eradat, H.; Chen, A.M.; Gupta, R.S.; Brahn, E.; Sercarz, E.E. Diversification of T Cell Responses to Carboxy-Terminal Determinants within the 65-KD Heat-Shock Protein Is Involved in Regulation of Autoimmune Arthritis. J. Exp. Med. 1997, 185, 1307–1316. [Google Scholar] [CrossRef] [Green Version]
- Krone, P.H.; Lele, Z.; Sass, J.B. Heat Shock Genes and the Heat Shock Response in Zebrafish Embryos. Biochem. Cell Biol. 1997, 75, 487–497. [Google Scholar] [CrossRef]
- Desai, K.; Spikings, E.; Zhang, T. 103. Studies on the Role of Hsp47 and Hsp90 in Zebrafish (Danio Rerio) Embryos under Chilled Condition. Cryobiology 2011, 63, 334. [Google Scholar] [CrossRef]
- Murtha, J. Characterization of the Heat Shock Response in Mature Zebrafish (Danio Rerio). Exp. Gerontol. 2003, 38, 683–691. [Google Scholar] [CrossRef]
- Lam, P.; Harvie, E.A.; Huttenlocher, A. Heat Shock Modulates Neutrophil Motility in Zebrafish. PLoS ONE 2013, 8, e84436. [Google Scholar] [CrossRef] [Green Version]
- Han, B.; Luo, J.; Jiang, P.; Li, Y.; Wang, Q.; Bai, Y.; Chen, J.; Wang, J.; Zhang, J. Inhibition of Embryonic HSP 90 Function Promotes Variation of Cold Tolerance in Zebrafish. Front. Genet. 2020, 11, 541944. [Google Scholar] [CrossRef]
- Pei, W.; Tanaka, K.; Huang, S.C.; Xu, L.; Liu, B.; Sinclair, J.; Idol, J.; Varshney, G.K.; Huang, H.; Lin, S.; et al. Extracellular HSP60 Triggers Tissue Regeneration and Wound Healing by Regulating Inflammation and Cell Proliferation. NPJ Regen. Med. 2016, 1, 16013. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.L.; Pan, X.; Mudumana, S.P.; Wang, H.; Kee, P.W.; Gong, Z. Development of a Heat Shock Inducible Gfp Transgenic Zebrafish Line by Using the Zebrafish Hsp27 Promoter. Gene 2008, 408, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Halloran, M.C.; Sato-Maeda, M.; Warren, J.T.; Su, F.; Lele, Z.; Krone, P.H.; Kuwada, J.Y.; Shoji, W. Laser-Induced Gene Expression in Specific Cells of Transgenic Zebrafish. Development 2000, 127, 1953–1960. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, R.I.; Bevins, C.L.; Ganz, T. Defensins and Other Antimicrobial Peptides and Proteins. In Mucosal Immunology; Elsevier: Amsterdam, The Netherlands, 2005; pp. 95–110. ISBN 978-0-12-491543-5. [Google Scholar]
- Krensky, A.M.; Clayberger, C. Biology and Clinical Relevance of Granulysin. Tissue Antigens 2009, 73, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Zanetti, M. Cathelicidins, Multifunctional Peptides of the Innate Immunity. J. Leukoc. Biol. 2004, 75, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krijgsveld, J.; Zaat, S.A.J.; Meeldijk, J.; van Veelen, P.A.; Fang, G.; Poolman, B.; Brandt, E.; Ehlert, J.E.; Kuijpers, A.J.; Engbers, G.H.M.; et al. Thrombocidins, Microbicidal Proteins from Human Blood Platelets, Are C-Terminal Deletion Products of CXC Chemokines. J. Biol. Chem. 2000, 275, 20374–20381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehrer, R.I.; Barton, A.; Daher, K.A.; Harwig, S.S.; Ganz, T.; Selsted, M.E. Interaction of Human Defensins with Escherichia Coli. Mechanism of Bactericidal Activity. J. Clin. Investig. 1989, 84, 553–561. [Google Scholar] [CrossRef]
- Dürr, M.; Peschel, A. Chemokines Meet Defensins: The Merging Concepts of Chemoattractants and Antimicrobial Peptides in Host Defense. Infect. Immun. 2002, 70, 6515–6517. [Google Scholar] [CrossRef] [Green Version]
- Semple, F.; Dorin, J.R. β-Defensins: Multifunctional Modulators of Infection, Inflammation and More? J. Innate Immun. 2012, 4, 337–348. [Google Scholar] [CrossRef]
- Lillard, J.W.; Boyaka, P.N.; Chertov, O.; Oppenheim, J.J.; McGhee, J.R. Mechanisms for Induction of Acquired Host Immunity by Neutrophil Peptide Defensins. Proc. Natl. Acad. Sci. USA 1999, 96, 651–656. [Google Scholar] [CrossRef] [Green Version]
- Boniotto, M.; Jordan, W.J.; Eskdale, J.; Tossi, A.; Antcheva, N.; Crovella, S.; Connell, N.D.; Gallagher, G. Human β-Defensin 2 Induces a Vigorous Cytokine Response in Peripheral Blood Mononuclear Cells. Antimicrob. Agents Chemother. 2006, 50, 1433–1441. [Google Scholar] [CrossRef] [Green Version]
- Deng, A.; Chen, S.; Li, Q.; Lyu, S.; Clayberger, C.; Krensky, A.M. Granulysin, a Cytolytic Molecule, Is Also a Chemoattractant and Proinflammatory Activator. J. Immunol. 2005, 174, 5243–5248. [Google Scholar] [CrossRef] [PubMed]
- Mookherjee, N.; Hamill, P.; Gardy, J.; Blimkie, D.; Falsafi, R.; Chikatamarla, A.; Arenillas, D.J.; Doria, S.; Kollmann, T.R.; Hancock, R.E.W. Systems Biology Evaluation of Immune Responses Induced by Human Host Defence Peptide LL-37 in Mononuclear Cells. Mol. BioSyst. 2009, 5, 483. [Google Scholar] [CrossRef] [PubMed]
- Bian, T.; Li, H.; Zhou, Q.; Ni, C.; Zhang, Y.; Yan, F. Human β -Defensin 3 Reduces TNF-α-Induced Inflammation and Monocyte Adhesion in Human Umbilical Vein Endothelial Cells. Mediat. Inflamm. 2017, 2017, 8529542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armbruster, N.S.; Sidelmann Brinch, K.; Andersen, B.; Stange, E.F.; Malek, N.; Nordkild, P.; Wehkamp, J. P669 Human β-Defensin 2 Suppresses TNF-α Secretion in Human and Mouse Dendritic Cells Mediated by Chemokine Receptor-2. J. Crohn’s Colitis 2018, 12, S449. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Mercier, C.; Koussounadis, A.; Secombes, C. Discovery of Multiple Beta-Defensin like Homologues in Teleost Fish. Mol. Immunol. 2007, 44, 638–647. [Google Scholar] [CrossRef]
- García-Valtanen, P.; Martinez-Lopez, A.; Ortega-Villaizan, M.; Perez, L.; Coll, J.M.; Estepa, A. In Addition to Its Antiviral and Immunomodulatory Properties, the Zebrafish β-Defensin 2 (ZfBD2) Is a Potent Viral DNA Vaccine Molecular Adjuvant. Antivir. Res. 2014, 101, 136–147. [Google Scholar] [CrossRef]
- Tao, Y.; Song, C.F.; Li, W. Expression of the Zebrafish β-Defensin 3 Mature Peptide in Pichia Pastoris and Its Purification and Antibacterial Activity. Appl. Biochem. Microbiol. 2017, 53, 661–668. [Google Scholar] [CrossRef]
- Chen, C.; Wang, A.; Zhang, F.; Zhang, M.; Yang, H.; Li, J.; Su, P.; Chen, Y.; Yu, H.; Wang, Y. The Protective Effect of Fish-Derived Cathelicidins on Bacterial Infections in Zebrafish, Danio Rerio. Fish Shellfish. Immunol. 2019, 92, 519–527. [Google Scholar] [CrossRef]
- Pereiro, P.; Varela, M.; Diaz-Rosales, P.; Romero, A.; Dios, S.; Figueras, A.; Novoa, B. Zebrafish Nk-Lysins: First Insights about Their Cellular and Functional Diversification. Dev. Comp. Immunol. 2015, 51, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [Green Version]
- Kalliolias, G.D.; Ivashkiv, L.B. TNF Biology, Pathogenic Mechanisms and Emerging Therapeutic Strategies. Nat. Rev. Rheumatol. 2016, 12, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Overview of the IL-1 Family in Innate Inflammation and Acquired Immunity. Immunol. Rev. 2018, 281, 8–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, A.; Mukaiyama, A.; Itoh, Y.; Nagase, H.; Thogersen, I.B.; Enghild, J.J.; Sasaguri, Y.; Mori, Y. Degradation of Interleukin 1beta by Matrix Metalloproteinases. J. Biol. Chem. 1996, 271, 14657–14660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Vaart, M.; Spaink, H.P.; Meijer, A.H. Pathogen Recognition and Activation of the Innate Immune Response in Zebrafish. Adv. Hematol. 2012, 2012, 159807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.; Wang, Y.; Li, Z.; Ma, L.; Wei, X.; Yang, J.; Xiao, R.; Xia, C. The Unique Structure of the Zebrafish TNF-α Homotrimer. Dev. Comp. Immunol. 2021, 122, 104129. [Google Scholar] [CrossRef]
- Kinoshita, S.; Biswas, G.; Kono, T.; Hikima, J.; Sakai, M. Presence of Two Tumor Necrosis Factor (Tnf)-α Homologs on Different Chromosomes of Zebrafish (Danio Rerio) and Medaka (Oryzias Latipes). Mar. Genom. 2014, 13, 1–9. [Google Scholar] [CrossRef]
- Candel, S.; de Oliveira, S.; López-Muñoz, A.; García-Moreno, D.; Espín-Palazón, R.; Tyrkalska, S.D.; Cayuela, M.L.; Renshaw, S.A.; Corbalán-Vélez, R.; Vidal-Abarca, I.; et al. Tnfa Signaling Through Tnfr2 Protects Skin Against Oxidative Stress–Induced Inflammation. PLoS Biol. 2014, 12, e1001855. [Google Scholar] [CrossRef] [Green Version]
- Bernut, A.; Nguyen-Chi, M.; Halloum, I.; Herrmann, J.-L.; Lutfalla, G.; Kremer, L. Mycobacterium Abscessus-Induced Granuloma Formation Is Strictly Dependent on TNF Signaling and Neutrophil Trafficking. PLoS Pathog. 2016, 12, e1005986. [Google Scholar] [CrossRef] [Green Version]
- Clay, H.; Volkman, H.E.; Ramakrishnan, L. Tumor Necrosis Factor Signaling Mediates Resistance to Mycobacteria by Inhibiting Bacterial Growth and Macrophage Death. Immunity 2008, 29, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Kam, J.Y.; Hortle, E.; Krogman, E.; Warner, S.E.; Wright, K.; Luo, K.; Cheng, T.; Manuneedhi Cholan, P.; Kikuchi, K.; Triccas, J.A.; et al. Rough and Smooth Variants of Mycobacterium Abscessus Are Differentially Controlled by Host Immunity during Chronic Infection of Adult Zebrafish. Nat. Commun. 2022, 13, 952. [Google Scholar] [CrossRef]
- Roca, F.J.; Whitworth, L.J.; Redmond, S.; Jones, A.A.; Ramakrishnan, L. TNF Induces Pathogenic Programmed Macrophage Necrosis in Tuberculosis through a Mitochondrial-Lysosomal-Endoplasmic Reticulum Circuit. Cell 2019, 178, 1344–1361.e11. [Google Scholar] [CrossRef] [PubMed]
- Ogryzko, N.V.; Hoggett, E.E.; Solaymani-Kohal, S.; Tazzyman, S.; Chico, T.J.A.; Renshaw, S.A.; Wilson, H.L. Zebrafish Tissue Injury Causes Upregulation of Interleukin-1 and Caspase-Dependent Amplification of the Inflammatory Response. Dis. Model. Mech. 2014, 7, 259–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, S.H.; Dóró, E.; Hammond, F.R.; Nguyen-Chi, M.E.; Lutfalla, G.; Wiegertjes, G.F.; Forlenza, M. Occurrence of Foamy Macrophages during the Innate Response of Zebrafish to Trypanosome Infections. eLife 2021, 10, e64520. [Google Scholar] [CrossRef] [PubMed]
- Mazon-Moya, M.J.; Willis, A.R.; Torraca, V.; Boucontet, L.; Shenoy, A.R.; Colucci-Guyon, E.; Mostowy, S. Septins Restrict Inflammation and Protect Zebrafish Larvae from Shigella Infection. PLoS Pathog. 2017, 13, e1006467. [Google Scholar] [CrossRef] [Green Version]
- Saraceni, P.R.; Romero, A.; Figueras, A.; Novoa, B. Establishment of Infection Models in Zebrafish Larvae (Danio Rerio) to Study the Pathogenesis of Aeromonas Hydrophila. Front. Microbiol. 2016, 7, 1219. [Google Scholar] [CrossRef] [Green Version]
- Varela, M.; Romero, A.; Dios, S.; van der Vaart, M.; Figueras, A.; Meijer, A.H.; Novoa, B. Cellular Visualization of Macrophage Pyroptosis and Interleukin-1β Release in a Viral Hemorrhagic Infection in Zebrafish Larvae. J. Virol. 2014, 88, 12026–12040. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Li, C.; Zhang, Y.; Lin, C.; Zhang, Y.; Shu, L.; Luo, L.; Zhuo, J.; Li, L. Macrophage-Derived IL-1β Regulates Emergency Myelopoiesis via the NF-ΚB and C/Ebpβ in Zebrafish. J. Immunol. 2020, 205, 2694–2706. [Google Scholar] [CrossRef]
- Tsarouchas, T.M.; Wehner, D.; Cavone, L.; Munir, T.; Keatinge, M.; Lambertus, M.; Underhill, A.; Barrett, T.; Kassapis, E.; Ogryzko, N.; et al. Dynamic Control of Proinflammatory Cytokines Il-1β and Tnf-α by Macrophages in Zebrafish Spinal Cord Regeneration. Nat. Commun. 2018, 9, 4670. [Google Scholar] [CrossRef] [Green Version]
- Varela, M.; Dios, S.; Novoa, B.; Figueras, A. Characterisation, Expression and Ontogeny of Interleukin-6 and Its Receptors in Zebrafish (Danio Rerio). Dev. Comp. Immunol. 2012, 37, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Dhanagovind, P.T.; Kujur, P.K.; Swain, R.K.; Banerjee, S. IL-6 Signaling Protects Zebrafish Larvae during Staphylococcus Epidermidis Infection in a Bath Immersion Model. J. Immunol. 2021, 207, 2129–2142. [Google Scholar] [CrossRef]
- Zhao, X.-F.; Wan, J.; Powell, C.; Ramachandran, R.; Myers, M.G.; Goldman, D. Leptin and IL-6 Family Cytokines Synergize to Stimulate Müller Glia Reprogramming and Retina Regeneration. Cell Rep. 2014, 9, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Gupta, V.; Karra, R.; Holdway, J.E.; Kikuchi, K.; Poss, K.D. Translational Profiling of Cardiomyocytes Identifies an Early Jak1/Stat3 Injury Response Required for Zebrafish Heart Regeneration. Proc. Natl. Acad. Sci. USA 2013, 110, 13416–13421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, Y.; Kiyooka, M.; Ohshima, T. Transcriptome Analyses Reveal IL6/Stat3 Signaling Involvement in Radial Glia Proliferation After Stab Wound Injury in the Adult Zebrafish Optic Tectum. Front. Cell Dev. Biol. 2021, 9, 668408. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, C.; Kim, C.H. Zebrafish as a Model for Infectious Disease and Immune Function. Fish Shellfish Immunol. 2008, 25, 341–350. [Google Scholar] [CrossRef]
- Zhang, D.-C.; Shao, Y.-Q.; Huang, Y.-Q.; Jiang, S.-G. Cloning, Characterization and Expression Analysis of Interleukin-10 from the Zebrafish (Danio Rerion). J. Biochem. Mol. Biol. 2005, 38, 571–576. [Google Scholar] [CrossRef] [Green Version]
- Ohtani, M.; Hayashi, N.; Hashimoto, K.; Nakanishi, T.; Dijkstra, J.M. Comprehensive Clarification of Two Paralogous Interleukin 4/13 Loci in Teleost Fish. Immunogenetics 2008, 60, 383–397. [Google Scholar] [CrossRef]
- Hughes, C.E.; Nibbs, R.J.B. A Guide to Chemokines and Their Receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef]
- López-Cotarelo, P.; Gómez-Moreira, C.; Criado-García, O.; Sánchez, L.; Rodríguez-Fernández, J.L. Beyond Chemoattraction: Multifunctionality of Chemokine Receptors in Leukocytes. Trends Immunol. 2017, 38, 927–941. [Google Scholar] [CrossRef]
- Alcami, A. Viral Mimicry of Cytokines, Chemokines and Their Receptors. Nat. Rev. Immunol. 2003, 3, 36–50. [Google Scholar] [CrossRef]
- Bird, S.; Tafalla, C. Teleost Chemokines and Their Receptors. Biology 2015, 4, 756–784. [Google Scholar] [CrossRef] [Green Version]
- Nomiyama, H.; Hieshima, K.; Osada, N.; Kato-Unoki, Y.; Otsuka-Ono, K.; Takegawa, S.; Izawa, T.; Yoshizawa, A.; Kikuchi, Y.; Tanase, S.; et al. Extensive Expansion and Diversification of the Chemokine Gene Family in Zebrafish: Identification of a Novel Chemokine Subfamily CX. BMC Genom. 2008, 9, 222. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xu, Q.; Wang, T.; Collet, B.; Corripio-Miyar, Y.; Bird, S.; Xie, P.; Nie, P.; Secombes, C.J.; Zou, J. Phylogenetic Analysis of Vertebrate CXC Chemokines Reveals Novel Lineage Specific Groups in Teleost Fish. Dev. Comp. Immunol. 2013, 41, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Sommer, F.; Torraca, V.; Meijer, A.H. Chemokine Receptors and Phagocyte Biology in Zebrafish. Front. Immunol. 2020, 11, 325. [Google Scholar] [CrossRef] [Green Version]
- Zuñiga-Traslaviña, C.; Bravo, K.; Reyes, A.E.; Feijóo, C.G. Cxcl8b and Cxcr2 Regulate Neutrophil Migration through Bloodstream in Zebrafish. J. Immunol. Res. 2017, 2017, 6530531. [Google Scholar] [CrossRef] [Green Version]
- Sommer, F.; Torraca, V.; Kamel, S.M.; Lombardi, A.; Meijer, A.H. Frontline Science: Antagonism between Regular and Atypical Cxcr3 Receptors Regulates Macrophage Migration during Infection and Injury in Zebrafish. J. Leukoc. Biol. 2020, 107, 185–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torraca, V.; Cui, C.; Boland, R.; Bebelman, J.-P.; van der Sar, A.M.; Smit, M.J.; Siderius, M.; Spaink, H.P.; Meijer, A.H. The CXCR3-CXCL11 Signaling Axis Mediates Macrophage Recruitment and Dissemination of Mycobacterial Infection. Dis. Model. Mech. 2015, 8, 253–269. [Google Scholar] [CrossRef] [Green Version]
- Isles, H.M.; Herman, K.D.; Robertson, A.L.; Loynes, C.A.; Prince, L.R.; Elks, P.M.; Renshaw, S.A. The CXCL12/CXCR4 Signaling Axis Retains Neutrophils at Inflammatory Sites in Zebrafish. Front. Immunol. 2019, 10, 1784. [Google Scholar] [CrossRef] [Green Version]
- Stratman, A.N.; Burns, M.C.; Farrelly, O.M.; Davis, A.E.; Li, W.; Pham, V.N.; Castranova, D.; Yano, J.J.; Goddard, L.M.; Nguyen, O.; et al. Chemokine Mediated Signalling within Arteries Promotes Vascular Smooth Muscle Cell Recruitment. Commun. Biol. 2020, 3, 734. [Google Scholar] [CrossRef]
- Sommer, F.; Ortiz Zacarías, N.V.; Heitman, L.H.; Meijer, A.H. Inhibition of Macrophage Migration in Zebrafish Larvae Demonstrates in Vivo Efficacy of Human CCR2 Inhibitors. Dev. Comp. Immunol. 2021, 116, 103932. [Google Scholar] [CrossRef]
- Naumann, U.; Cameroni, E.; Pruenster, M.; Mahabaleshwar, H.; Raz, E.; Zerwes, H.-G.; Rot, A.; Thelen, M. CXCR7 Functions as a Scavenger for CXCL12 and CXCL11. PLoS ONE 2010, 5, e9175. [Google Scholar] [CrossRef] [Green Version]
- Donà, E.; Barry, J.D.; Valentin, G.; Quirin, C.; Khmelinskii, A.; Kunze, A.; Durdu, S.; Newton, L.R.; Fernandez-Minan, A.; Huber, W.; et al. Directional Tissue Migration through a Self-Generated Chemokine Gradient. Nature 2013, 503, 285–289. [Google Scholar] [CrossRef]
- Zakrzewska, A.; Cui, C.; Stockhammer, O.W.; Benard, E.L.; Spaink, H.P.; Meijer, A.H. Macrophage-Specific Gene Functions in Spi1-Directed Innate Immunity. Blood 2010, 116, e1–e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaacs, A.; Lindenmann, J. Virus Interference. I. The Interferon. Proc. R. Soc. Lond. B. Biol. Sci. 1957, 147, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Secombes, C.J.; Zou, J. Evolution of Interferons and Interferon Receptors. Front. Immunol. 2017, 8, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazan, J.F. Haemopoietic Receptors and Helical Cytokines. Immunol. Today 1990, 11, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Nicola, N.A.; Hilton, D.J. General Classes and Functions of Four-Helix Bundle Cytokines. Adv. Protein Chem. 1998, 52, 1–65. [Google Scholar] [CrossRef]
- Chen, S.N.; Gan, Z.; Hou, J.; Yang, Y.C.; Huang, L.; Huang, B.; Wang, S.; Nie, P. Identification and Establishment of Type IV Interferon and the Characterization of Interferon-υ Including Its Class II Cytokine Receptors IFN-ΥR1 and IL-10R2. Nat. Commun. 2022, 13, 999. [Google Scholar] [CrossRef]
- Su, J. The Discovery of Type IV Interferon System Revolutionizes Interferon Family and Opens up a New Frontier in Jawed Vertebrate Immune Defense. Sci. China Life Sci. 2022, 65, 2335–2337. [Google Scholar] [CrossRef]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I Interferons in Infectious Disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef]
- Kerner, G.; Rosain, J.; Guérin, A.; Al-Khabaz, A.; Oleaga-Quintas, C.; Rapaport, F.; Massaad, M.J.; Ding, J.-Y.; Khan, T.; Ali, F.A.; et al. Inherited Human IFN-γ Deficiency Underlies Mycobacterial Disease. J. Clin. Investig. 2020, 130, 3158–3171. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, G. The Molecular Basis for Differential Type I Interferon Signaling. J. Biol. Chem. 2017, 292, 7285–7294. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Gorgoglione, B.; Taylor, N.G.H.; Summathed, T.; Lee, P.-T.; Panigrahi, A.; Genet, C.; Chen, Y.-M.; Chen, T.-Y.; Ul Hassan, M.; et al. Salmonids Have an Extraordinary Complex Type I IFN System: Characterization of the IFN Locus in Rainbow Trout Oncorhynchus Mykiss Reveals Two Novel IFN Subgroups. J. Immunol. 2014, 193, 2273–2286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggad, D.; Stein, C.; Sieger, D.; Mazel, M.; Boudinot, P.; Herbomel, P.; Levraud, J.-P.; Lutfalla, G.; Leptin, M. In Vivo Analysis of Ifn-Γ1 and Ifn-Γ2 Signaling in Zebrafish. J. Immunol. 2010, 185, 6774–6782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manivasagam, S.; Klein, R.S. Type III Interferons: Emerging Roles in Autoimmunity. Front. Immunol. 2021, 12, 764062. [Google Scholar] [CrossRef] [PubMed]
- Platanias, L.C. Mechanisms of Type-I- and Type-II-Interferon-Mediated Signalling. Nat. Rev. Immunol. 2005, 5, 375–386. [Google Scholar] [CrossRef]
- Levraud, J.-P.; Jouneau, L.; Briolat, V.; Laghi, V.; Boudinot, P. IFN-Stimulated Genes in Zebrafish and Humans Define an Ancient Arsenal of Antiviral Immunity. J. Immunol. 2019, 203, 3361–3373. [Google Scholar] [CrossRef]
- Crow, Y.J.; Stetson, D.B. The Type I Interferonopathies: 10 Years On. Nat. Rev. Immunol. 2022, 22, 471–483. [Google Scholar] [CrossRef]
- Hamming, O.J.; Lutfalla, G.; Levraud, J.-P.; Hartmann, R. Crystal Structure of Zebrafish Interferons I and II Reveals Conservation of Type I Interferon Structure in Vertebrates. J. Virol. 2011, 85, 8181–8187. [Google Scholar] [CrossRef] [Green Version]
- Jamilloux, Y.; Henry, T. Les Inflammasomes: Plates-Formes de l’immunité Innée. Med. Sci. 2013, 29, 975–984. [Google Scholar] [CrossRef] [Green Version]
- Franchi, L.; Eigenbrod, T.; Muñoz-Planillo, R.; Nuñez, G. The Inflammasome: A Caspase-1-Activation Platform That Regulates Immune Responses and Disease Pathogenesis. Nat. Immunol. 2009, 10, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Downs, K.P.; Nguyen, H.; Dorfleutner, A.; Stehlik, C. An Overview of the Non-Canonical Inflammasome. Mol. Asp. Med. 2020, 76, 100924. [Google Scholar] [CrossRef] [PubMed]
- Forn-Cuní, G.; Meijer, A.H.; Varela, M. Zebrafish in Inflammasome Research. Cells 2019, 8, 901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spead, O.; Verreet, T.; Donelson, C.J.; Poulain, F.E. Characterization of the Caspase Family in Zebrafish. PLoS ONE 2018, 13, e0197966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Huang, Y.; Cao, X.; Yin, X.; Jin, X.; Liu, S.; Jiang, J.; Jiang, W.; Xiao, T.S.; Zhou, R.; et al. Functional and Structural Characterization of Zebrafish ASC. FEBS J. 2018, 285, 2691–2707. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Schiffer, P.H.; Zielinski, J.; Wiehe, T.; Laird, G.K.; Marioni, J.C.; Soylemez, O.; Kondrashov, F.; Leptin, M. Structure and Evolutionary History of a Large Family of NLR Proteins in the Zebrafish. Open Biol. 2016, 6, 160009. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, Y.; Cao, X.; Jin, X.; Jin, T. Pattern Recognition Receptors in Zebrafish Provide Functional and Evolutionary Insight into Innate Immune Signaling Pathways. Cell Mol. Immunol. 2017, 14, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Nie, L.; Zhang, Y.; Dong, W.; Xiang, L.; Shao, J. Involvement of Zebrafish RIG-I in NF-ΚB and IFN Signaling Pathways: Insights into Functional Conservation of RIG-I in Antiviral Innate Immunity. Dev. Comp. Immunol. 2015, 48, 95–101. [Google Scholar] [CrossRef]
- Sun, F.; Zhang, Y.-B.; Liu, T.-K.; Shi, J.; Wang, B.; Gui, J.-F. Fish MITA Serves as a Mediator for Distinct Fish IFN Gene Activation Dependent on IRF3 or IRF7. J. Immunol. 2011, 187, 2531–2539. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.M.; Zhang, J.; Li, P.W.; Hu, Y.W.; Cao, L.; Ouyang, S.; Bi, Y.H.; Nie, P.; Chang, M.X. NOD1 Promotes Antiviral Signaling by Binding Viral RNA and Regulating the Interaction of MDA5 and MAVS. J. Immunol. 2020, 204, 2216–2231. [Google Scholar] [CrossRef]
- Gabor, K.A.; Charette, J.R.; Pietraszewski, M.J.; Wingfield, D.J.; Shim, J.S.; Millard, P.J.; Kim, C.H. A DN-Mda5 Transgenic Zebrafish Model Demonstrates That Mda5 Plays an Important Role in Snakehead Rhabdovirus Resistance. Dev. Comp. Immunol. 2015, 51, 298–304. [Google Scholar] [CrossRef] [Green Version]
- Pham, C.T.N. Neutrophil Serine Proteases: Specific Regulators of Inflammation. Nat. Rev. Immunol. 2006, 6, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.J.; Campbell, M.A.; Owen, C.A. Bioactive Proteinase 3 on the Cell Surface of Human Neutrophils: Quantification, Catalytic Activity, and Susceptibility to Inhibition. J. Immunol. 2000, 165, 3366–3374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, C.A.; Campbell, M.A.; Sannes, P.L.; Boukedes, S.S.; Campbell, E.J. Cell Surface-Bound Elastase and Cathepsin G on Human Neutrophils: A Novel, Non-Oxidative Mechanism by Which Neutrophils Focus and Preserve Catalytic Activity of Serine Proteinases. J. Cell Biol. 1995, 131, 775–789. [Google Scholar] [CrossRef]
- Reeves, E.P.; Lu, H.; Jacobs, H.L.; Messina, C.G.M.; Bolsover, S.; Gabella, G.; Potma, E.O.; Warley, A.; Roes, J.; Segal, A.W. Killing Activity of Neutrophils Is Mediated through Activation of Proteases by K+ Flux. Nature 2002, 416, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Kessenbrock, K.; Dau, T.; Jenne, D.E. Tailor-Made Inflammation: How Neutrophil Serine Proteases Modulate the Inflammatory Response. J. Mol. Med. 2011, 89, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Champagne, B.; Tremblay, P.; Cantin, A.; St Pierre, Y. Proteolytic Cleavage of ICAM-1 by Human Neutrophil Elastase. J. Immunol. 1998, 161, 6398–6405. [Google Scholar] [CrossRef]
- Lévesque, J.P.; Takamatsu, Y.; Nilsson, S.K.; Haylock, D.N.; Simmons, P.J. Vascular Cell Adhesion Molecule-1 (CD106) Is Cleaved by Neutrophil Proteases in the Bone Marrow Following Hematopoietic Progenitor Cell Mobilization by Granulocyte Colony-Stimulating Factor. Blood 2001, 98, 1289–1297. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.M.; Betz, T.V.; Lamont, D.J.; Kim, M.B.; Shaw, S.K.; Froio, R.M.; Baleux, F.; Arenzana-Seisdedos, F.; Alon, R.; Luscinskas, F.W. Elastase Release by Transmigrating Neutrophils Deactivates Endothelial-Bound SDF-1α and Attenuates Subsequent T Lymphocyte Transendothelial Migration. J. Exp. Med. 2004, 200, 713–724. [Google Scholar] [CrossRef]
- Padrines, M.; Wolf, M.; Walz, A.; Baggiolini, M. Interleukin-8 Processing by Neutrophil Elastase, Cathepsin G and Proteinase-3. FEBS Lett. 1994, 352, 231–235. [Google Scholar] [CrossRef] [Green Version]
- Robache-Gallea, S.; Morand, V.; Bruneau, J.M.; Schoot, B.; Tagat, E.; Réalo, E.; Chouaib, S.; Roman-Roman, S. In Vitro Processing of Human Tumor Necrosis Factor-α. J. Biol. Chem. 1995, 270, 23688–23692. [Google Scholar] [CrossRef] [PubMed]
- Young, R.E.; Thompson, R.D.; Larbi, K.Y.; La, M.; Roberts, C.E.; Shapiro, S.D.; Perretti, M.; Nourshargh, S. Neutrophil Elastase (NE)-Deficient Mice Demonstrate a Nonredundant Role for NE in Neutrophil Migration, Generation of Proinflammatory Mediators, and Phagocytosis in Response to Zymosan Particles In Vivo. J. Immunol. 2004, 172, 4493–4502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heuberger, D.M.; Schuepbach, R.A. Protease-Activated Receptors (PARs): Mechanisms of Action and Potential Therapeutic Modulators in PAR-Driven Inflammatory Diseases. Thromb. J. 2019, 17, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, D.E.; Greene, C.M.; Carroll, T.P.; Taggart, C.C.; Gallagher, P.M.; O’Neill, S.J.; McElvaney, N.G. Interleukin-8 Up-Regulation by Neutrophil Elastase Is Mediated by MyD88/IRAK/TRAF-6 in Human Bronchial Epithelium. J. Biol. Chem. 2001, 276, 35494–35499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Berg, C.W.; Tambourgi, D.V.; Clark, H.W.; Hoong, S.J.; Spiller, O.B.; McGreal, E.P. Mechanism of Neutrophil Dysfunction: Neutrophil Serine Proteases Cleave and Inactivate the C5a Receptor. J. Immunol. 2014, 192, 1787–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Nathan, C.; Jin, W.; Sim, D.; Ashcroft, G.S.; Wahl, S.M.; Lacomis, L.; Erdjument-Bromage, H.; Tempst, P.; Wright, C.D.; et al. Conversion of Proepithelin to Epithelins: Roles of SLPI and Elastase in Host Defense and Wound Repair. Cell 2002, 111, 867–878. [Google Scholar] [CrossRef]
- Oltmanns, U.; Sukkar, M.B.; Xie, S.; John, M.; Chung, K.F. Induction of Human Airway Smooth Muscle Apoptosis by Neutrophils and Neutrophil Elastase. Am. J. Respir. Cell Mol. Biol. 2005, 32, 334–341. [Google Scholar] [CrossRef]
- Akula, S.; Thorpe, M.; Boinapally, V.; Hellman, L. Granule Associated Serine Proteases of Hematopoietic Cells—An Analysis of Their Appearance and Diversification during Vertebrate Evolution. PLoS ONE 2015, 10, e0143091. [Google Scholar] [CrossRef] [Green Version]
- Bazán, N.G. Effects of Ischemia and Electroconvulsive Shock on Free Fatty Acid Pool in the Brain. Biochim. Biophys. Acta 1970, 218, 1–10. [Google Scholar] [CrossRef]
- Shohami, E.; Shapira, Y.; Yadid, G.; Reisfeld, N.; Yedgar, S. Brain Phospholipase A2 Is Activated after Experimental Closed Head Injury in the Rat. J. Neurochem. 1989, 53, 1541–1546. [Google Scholar] [CrossRef]
- Dyall, S.C.; Balas, L.; Bazan, N.G.; Brenna, J.T.; Chiang, N.; da Costa Souza, F.; Dalli, J.; Durand, T.; Galano, J.-M.; Lein, P.J.; et al. Polyunsaturated Fatty Acids and Fatty Acid-Derived Lipid Mediators: Recent Advances in the Understanding of Their Biosynthesis, Structures, and Functions. Prog. Lipid Res. 2022, 86, 101165. [Google Scholar] [CrossRef] [PubMed]
- Flower, R.J. Prostaglandins, Bioassay and Inflammation. Br. J. Pharm. 2006, 147 (Suppl. 1), S182–S192. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Polyunsaturated Fatty Acids and Inflammation. Biochem. Soc. Trans. 2005, 33, 423–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serhan, C.N. Pro-Resolving Lipid Mediators Are Leads for Resolution Physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, E.-Y.; Cho, S.-H.; Kwon, S.-H.; Eom, C.-Y.; Jeong, M.S.; Lee, W.; Kim, S.-Y.; Heo, S.-J.; Ahn, G.; Lee, K.P.; et al. The Roles of NF-ΚB and ROS in Regulation of pro-Inflammatory Mediators of Inflammation Induction in LPS-Stimulated Zebrafish Embryos. Fish Shellfish Immunol. 2017, 68, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Thrikawala, S.; Niu, M.; Keller, N.P.; Rosowski, E.E. Cyclooxygenase Production of PGE2 Promotes Phagocyte Control of A. Fumigatus Hyphal Growth in Larval Zebrafish. PLoS Pathog. 2022, 18, e1010040. [Google Scholar] [CrossRef]
- Adel, S.; Heydeck, D.; Kuhn, H.; Ufer, C. The Lipoxygenase Pathway in Zebrafish. Expression and Characterization of Zebrafish ALOX5 and Comparison with Its Human Ortholog. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2016, 1861, 1–11. [Google Scholar] [CrossRef]
- Chatzopoulou, A.; Heijmans, J.P.M.; Burgerhout, E.; Oskam, N.; Spaink, H.P.; Meijer, A.H.; Schaaf, M.J.M. Glucocorticoid-Induced Attenuation of the Inflammatory Response in Zebrafish. Endocrinology 2016, 157, 2772–2784. [Google Scholar] [CrossRef] [Green Version]
- Loynes, C.A.; Lee, J.A.; Robertson, A.L.; Steel, M.J.; Ellett, F.; Feng, Y.; Levy, B.D.; Whyte, M.K.B.; Renshaw, S.A. PGE2 Production at Sites of Tissue Injury Promotes an Anti-Inflammatory Neutrophil Phenotype and Determines the Outcome of Inflammation Resolution in Vivo. Sci. Adv. 2018, 4, eaar8320. [Google Scholar] [CrossRef] [Green Version]
- Hansen, T.V.; Vik, A.; Serhan, C.N. The Protectin Family of Specialized Pro-Resolving Mediators: Potent Immunoresolvents Enabling Innovative Approaches to Target Obesity and Diabetes. Front. Pharmacol. 2019, 9, 1582. [Google Scholar] [CrossRef]
- Schwab, J.M.; Chiang, N.; Arita, M.; Serhan, C.N. Resolvin E1 and Protectin D1 Activate Inflammation-Resolution Programmes. Nature 2007, 447, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Nguyen-Chi, M.; Luz-Crawford, P.; Balas, L.; Sipka, T.; Contreras-López, R.; Barthelaix, A.; Lutfalla, G.; Durand, T.; Jorgensen, C.; Djouad, F. Pro-resolving Mediator Protectin D1 Promotes Epimorphic Regeneration by Controlling Immune Cell Function in Vertebrates. Br. J. Pharm. 2020, 177, 4055–4073. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Langmead, C.J.; Riddy, D.M. New Advances in Targeting the Resolution of Inflammation: Implications for Specialized Pro-Resolving Mediator GPCR Drug Discovery. ACS Pharmacol. Transl. Sci. 2020, 3, 88–106. [Google Scholar] [CrossRef] [PubMed]
- Vago, J.P.; Nogueira, C.R.C.; Tavares, L.P.; Soriani, F.M.; Lopes, F.; Russo, R.C.; Pinho, V.; Teixeira, M.M.; Sousa, L.P. Annexin A1 Modulates Natural and Glucocorticoid-Induced Resolution of Inflammation by Enhancing Neutrophil Apoptosis. J. Leukoc. Biol. 2012, 92, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.A.; Vago, J.P.; Teixeira, M.M.; Sousa, L.P. Annexin A1 and the Resolution of Inflammation: Modulation of Neutrophil Recruitment, Apoptosis, and Clearance. J. Immunol. Res. 2016, 2016, 8239258. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, M.A.; Ribeiro, A.L.C.; Costa, B.R.C.; Vago, J.P.; Lima, K.M.; Carneiro, F.S.; Ortiz, M.M.O.; Lima, G.L.N.; Carmo, A.A.F.; Rocha, R.M.; et al. Plasmin and Plasminogen Induce Macrophage Reprogramming and Regulate Key Steps of Inflammation Resolution via Annexin A1. Blood 2017, 129, 2896–2907. [Google Scholar] [CrossRef] [Green Version]
- Lim, L.H.K.; Solito, E.; Russo-Marie, F.; Flower, R.J.; Perretti, M. Promoting Detachment of Neutrophils Adherent to Murine Postcapillary Venules to Control Inflammation: Effect of Lipocortin 1. Proc. Natl. Acad. Sci. USA 1998, 95, 14535–14539. [Google Scholar] [CrossRef] [Green Version]
- Rubinstein, N.; Ilarregui, J.M.; Toscano, M.A.; Rabinovich, G.A. The Role of Galectins in the Initiation, Amplification and Resolution of the Inflammatory Response. Tissue Antigens 2004, 64, 1–12. [Google Scholar] [CrossRef]
- Advedissian, T.; Deshayes, F.; Poirier, F.; Grandjean, C.; Viguier, M. Les Galectines: Des Lectines Pas Comme Les Autres. Med. Sci. 2015, 31, 499–505. [Google Scholar] [CrossRef] [Green Version]
- Almkvist, J.; Karlsson, A. Galectins as Inflammatory Mediators. Glycoconj. J. 2002, 19, 575–581. [Google Scholar] [CrossRef]
- Yaseen, H.; Butenko, S.; Polishuk-Zotkin, I.; Schif-Zuck, S.; Pérez-Sáez, J.M.; Rabinovich, G.A.; Ariel, A. Galectin-1 Facilitates Macrophage Reprogramming and Resolution of Inflammation Through IFN-β. Front. Pharmacol. 2020, 11, 901. [Google Scholar] [CrossRef] [PubMed]
- Quenum Zangbede, F.O.; Chauhan, A.; Sharma, J.; Mishra, B.B. Galectin-3 in M2 Macrophages Plays a Protective Role in Resolution of Neuropathology in Brain Parasitic Infection by Regulating Neutrophil Turnover. J. Neurosci. 2018, 38, 6737–6750. [Google Scholar] [CrossRef] [Green Version]
- Farber, S.A.; De Rose, R.A.; Olson, E.S.; Halpern, M.E. The Zebrafish Annexin Gene Family. Genome Res. 2003, 13, 1082–1096. [Google Scholar] [CrossRef] [Green Version]
- Vasta, G.R.; Ahmed, H.; Du, S.-J.; Henrikson, D. Galectins in Teleost Fish: Zebrafish (Danio Rerio) as a Model Species to Address Their Biological Roles in Development and Innate Immunity. Glycoconj. J. 2004, 21, 503–521. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Nita-Lazar, M.; González-Montalbán, N.; Wang, J.; Mancini, J.; Ravindran, C.; Ahmed, H.; Vasta, G.R. Manipulating Galectin Expression in Zebrafish (Danio Rerio). In Galectins; Stowell, S.R., Cummings, R.D., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2015; Volume 1207, pp. 327–341. ISBN 978-1-4939-1395-4. [Google Scholar]
- Ahmed, H.; Du, S.-J.; Vasta, G.R. Knockdown of a Galectin-1-like Protein in Zebrafish (Danio Rerio) Causes Defects in Skeletal Muscle Development. Glycoconj. J. 2009, 26, 277–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leite, C.E.; de Oliveira Maboni, L.; Cruz, F.F.; Rosemberg, D.B.; Zimmermann, F.F.; Pereira, T.C.B.; Bogo, M.R.; Bonan, C.D.; Campos, M.M.; Morrone, F.B.; et al. Involvement of Purinergic System in Inflammation and Toxicity Induced by Copper in Zebrafish Larvae. Toxicol. Appl. Pharmacol. 2013, 272, 681–689. [Google Scholar] [CrossRef]
- Powell, D.; Tauzin, S.; Hind, L.E.; Deng, Q.; Beebe, D.J.; Huttenlocher, A. Chemokine Signaling and the Regulation of Bidirectional Leukocyte Migration in Interstitial Tissues. Cell Rep. 2017, 19, 1572–1585. [Google Scholar] [CrossRef] [Green Version]
- Wittmann, C.; Reischl, M.; Shah, A.H.; Kronfuss, E.; Mikut, R.; Liebel, U.; Grabher, C. A Zebrafish Drug-Repurposing Screen Reveals SGC-Dependent and SGC-Independent Pro-Inflammatory Activities of Nitric Oxide. PLoS ONE 2015, 10, e0137286. [Google Scholar] [CrossRef]
- Matrone, G.; Jung, S.Y.; Choi, J.M.; Jain, A.; Leung, H.-C.E.; Rajapakshe, K.; Coarfa, C.; Rodor, J.; Denvir, M.A.; Baker, A.H.; et al. Nuclear S-Nitrosylation Impacts Tissue Regeneration in Zebrafish. Nat. Commun. 2021, 12, 6282. [Google Scholar] [CrossRef]
- Cha, S.-H.; Hwang, Y.; Kim, K.-N.; Jun, H.-S. Palmitate Induces Nitric Oxide Production and Inflammatory Cytokine Expression in Zebrafish. Fish Shellfish Immunol. 2018, 79, 163–167. [Google Scholar] [CrossRef]
- Leite, C.E.; da Cruz Teixeira, A.; Cruz, F.F.; Concatto, S.C.; Amaral, J.H.; Bonan, C.D.; Campos, M.M.; Morrone, F.B.; Battastini, A.M.O. Analytical Method for Determination of Nitric Oxide in Zebrafish Larvae: Toxicological and Pharmacological Applications. Anal. Biochem. 2012, 421, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Oehlers, S.H.; Flores, M.V.; Hall, C.J.; Crosier, K.E.; Crosier, P.S. Retinoic Acid Suppresses Intestinal Mucus Production and Exacerbates Experimental Enterocolitis. Dis. Model. Mech. 2012, 5, 457–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paudel, Y.N.; Khan, S.U.; Othman, I.; Shaikh, M.F. Naturally Occurring HMGB1 Inhibitor, Glycyrrhizin, Modulates Chronic Seizures-Induced Memory Dysfunction in Zebrafish Model. ACS Chem. Neurosci. 2021, 12, 3288–3302. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Jiang, X.; Paudel, Y.N.; Gao, X.; Gao, D.; Zhang, P.; Sheng, W.; Shang, X.; Liu, K.; Zhang, X.; et al. Co-Treatment with Natural HMGB1 Inhibitor Glycyrrhizin Exerts Neuroprotection and Reverses Parkinson’s Disease like Pathology in Zebrafish. J. Ethnopharmacol. 2022, 292, 115234. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Sun, C.; Chen, Z.; Yang, D.; Zhou, Z.; Peng, X.; Tang, C. Alcohol Induces Zebrafish Skeletal Muscle Atrophy through HMGB1/TLR4/NF-ΚB Signaling. Life 2022, 12, 1211. [Google Scholar] [CrossRef]
- Progatzky, F.; Cook, H.T.; Lamb, J.R.; Bugeon, L.; Dallman, M.J. Mucosal Inflammation at the Respiratory Interface: A Zebrafish Model. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 310, L551–L561. [Google Scholar] [CrossRef] [Green Version]
- Hatzold, J.; Beleggia, F.; Herzig, H.; Altmüller, J.; Nürnberg, P.; Bloch, W.; Wollnik, B.; Hammerschmidt, M. Tumor Suppression in Basal Keratinocytes via Dual Non-Cell-Autonomous Functions of a Na,K-ATPase Beta Subunit. eLife 2016, 5, e14277. [Google Scholar] [CrossRef]
- Karra, R.; Knecht, A.K.; Kikuchi, K.; Poss, K.D. Myocardial NF-ΚB Activation Is Essential for Zebrafish Heart Regeneration. Proc. Natl. Acad. Sci. USA 2015, 112, 13255–13260. [Google Scholar] [CrossRef] [Green Version]
- Goody, M.; Jurczyszak, D.; Kim, C.; Henry, C. Influenza A Virus Infection Damages Zebrafish Skeletal Muscle and Exacerbates Disease in Zebrafish Modeling Duchenne Muscular Dystrophy. PLoS Curr. 2017, 9, ecurrents.md.8a7e35c50fa2b48156799d3c39788175. [Google Scholar] [CrossRef]
- Kienle, K.; Lämmermann, T. Neutrophil Swarming: An Essential Process of the Neutrophil Tissue Response. Immunol. Rev. 2016, 273, 76–93. [Google Scholar] [CrossRef]
- Poplimont, H.; Georgantzoglou, A.; Boulch, M.; Walker, H.A.; Coombs, C.; Papaleonidopoulou, F.; Sarris, M. Neutrophil Swarming in Damaged Tissue Is Orchestrated by Connexins and Cooperative Calcium Alarm Signals. Curr. Biol. 2020, 30, 2761–2776.e7. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Mantovani, A. Macrophage Plasticity and Polarization: In Vivo Veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Ginhoux, F.; Schultze, J.L.; Murray, P.J.; Ochando, J.; Biswas, S.K. New Insights into the Multidimensional Concept of Macrophage Ontogeny, Activation and Function. Nat. Immunol. 2016, 17, 34–40. [Google Scholar] [CrossRef]
- Forlenza, M.; Fink, I.R.; Raes, G.; Wiegertjes, G.F. Heterogeneity of Macrophage Activation in Fish. Dev. Comp. Immunol. 2011, 35, 1246–1255. [Google Scholar] [CrossRef]
- Wentzel, A.S.; Petit, J.; van Veen, W.G.; Fink, I.R.; Scheer, M.H.; Piazzon, M.C.; Forlenza, M.; Spaink, H.P.; Wiegertjes, G.F. Transcriptome Sequencing Supports a Conservation of Macrophage Polarization in Fish. Sci. Rep. 2020, 10, 13470. [Google Scholar] [CrossRef]
- Wiegertjes, G.F.; Wentzel, A.S.; Spaink, H.P.; Elks, P.M.; Fink, I.R. Polarization of Immune Responses in Fish: The ‘Macrophages First’ Point of View. Mol. Immunol. 2016, 69, 146–156. [Google Scholar] [CrossRef]
- Rougeot, J.; Torraca, V.; Zakrzewska, A.; Kanwal, Z.; Jansen, H.J.; Sommer, F.; Spaink, H.P.; Meijer, A.H. RNAseq Profiling of Leukocyte Populations in Zebrafish Larvae Reveals a Cxcl11 Chemokine Gene as a Marker of Macrophage Polarization During Mycobacterial Infection. Front. Immunol. 2019, 10, 832. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.; Elks, P.M. Hypoxia Induces Macrophage Tnfa Expression via Cyclooxygenase and Prostaglandin E2 in Vivo. Front. Immunol. 2019, 10, 2321. [Google Scholar] [CrossRef]
- Miskolci, V.; Squirrell, J.; Rindy, J.; Vincent, W.; Sauer, J.D.; Gibson, A.; Eliceiri, K.W.; Huttenlocher, A. Distinct Inflammatory and Wound Healing Responses to Complex Caudal Fin Injuries of Larval Zebrafish. eLife 2019, 8, e45976. [Google Scholar] [CrossRef]
- Greenlee-Wacker, M.C. Clearance of Apoptotic Neutrophils and Resolution of Inflammation. Immunol. Rev. 2016, 273, 357–370. [Google Scholar] [CrossRef] [Green Version]
- Mathias, J.R.; Perrin, B.J.; Liu, T.-X.; Kanki, J.; Look, A.T.; Huttenlocher, A. Resolution of Inflammation by Retrograde Chemotaxis of Neutrophils in Transgenic Zebrafish. J. Leukoc. Biol. 2006, 80, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Ellett, F.; Elks, P.M.; Robertson, A.L.; Ogryzko, N.V.; Renshaw, S.A. Defining the Phenotype of Neutrophils Following Reverse Migration in Zebrafish. J. Leukoc. Biol. 2015, 98, 975–981. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Morejón, A.; García-Redondo, A.B.; Reuter, H.; Marques, I.J.; Bates, T.; Galardi-Castilla, M.; Große, A.; Manig, S.; Langa, X.; Ernst, A.; et al. Wilms Tumor 1b Expression Defines a Pro-Regenerative Macrophage Subtype and Is Required for Organ Regeneration in the Zebrafish. Cell Rep. 2019, 28, 1296–1306.e6. [Google Scholar] [CrossRef] [Green Version]
- Denans, N.; Tran, N.T.T.; Swall, M.E.; Diaz, D.C.; Blanck, J.; Piotrowski, T. An Anti-Inflammatory Activation Sequence Governs Macrophage Transcriptional Dynamics during Tissue Injury in Zebrafish. Nat. Commun. 2022, 13, 5356. [Google Scholar] [CrossRef] [PubMed]
- Bohaud, C.; Johansen, M.D.; Jorgensen, C.; Ipseiz, N.; Kremer, L.; Djouad, F. The Role of Macrophages During Zebrafish Injury and Tissue Regeneration Under Infectious and Non-Infectious Conditions. Front. Immunol. 2021, 12, 707824. [Google Scholar] [CrossRef]
- Iribarne, M. Inflammation Induces Zebrafish Regeneration. Neural Regen. Res. 2021, 16, 1693. [Google Scholar] [CrossRef]
- Hasegawa, T.; Nakajima, T.; Ishida, T.; Kudo, A.; Kawakami, A. A Diffusible Signal Derived from Hematopoietic Cells Supports the Survival and Proliferation of Regenerative Cells during Zebrafish Fin Fold Regeneration. Dev. Biol. 2015, 399, 80–90. [Google Scholar] [CrossRef] [Green Version]
- Fang, P.; Pan, H.-C.; Lin, S.L.; Zhang, W.-Q.; Rauvala, H.; Schachner, M.; Shen, Y.-Q. HMGB1 Contributes to Regeneration After Spinal Cord Injury in Adult Zebrafish. Mol. Neurobiol. 2014, 49, 472–483. [Google Scholar] [CrossRef]
- Natarajan, N.; Abbas, Y.; Bryant, D.M.; Gonzalez-Rosa, J.M.; Sharpe, M.; Uygur, A.; Cocco-Delgado, L.H.; Ho, N.N.; Gerard, N.P.; Gerard, C.J.; et al. Complement Receptor C5aR1 Plays an Evolutionarily Conserved Role in Successful Cardiac Regeneration. Circulation 2018, 137, 2152–2165. [Google Scholar] [CrossRef]
- Mesureur, J.; Feliciano, J.R.; Wagner, N.; Gomes, M.C.; Zhang, L.; Blanco-Gonzalez, M.; van der Vaart, M.; O’Callaghan, D.; Meijer, A.H.; Vergunst, A.C. Macrophages, but Not Neutrophils, Are Critical for Proliferation of Burkholderia Cenocepacia and Ensuing Host-Damaging Inflammation. PLoS Pathog. 2017, 13, e1006437. [Google Scholar] [CrossRef]
- Cronan, M.R.; Hughes, E.J.; Brewer, W.J.; Viswanathan, G.; Hunt, E.G.; Singh, B.; Mehra, S.; Oehlers, S.H.; Gregory, S.G.; Kaushal, D.; et al. A Non-Canonical Type 2 Immune Response Coordinates Tuberculous Granuloma Formation and Epithelialization. Cell 2021, 184, 1757–1774.e14. [Google Scholar] [CrossRef] [PubMed]
- Farr, D.; Nag, D.; Chazin, W.J.; Harrison, S.; Thummel, R.; Luo, X.; Raychaudhuri, S.; Withey, J.H. Neutrophil-Associated Responses to Vibrio Cholerae Infection in a Natural Host Model. Infect. Immun. 2022, 90, e0046621. [Google Scholar] [CrossRef] [PubMed]
- Briolat, V.; Jouneau, L.; Carvalho, R.; Palha, N.; Langevin, C.; Herbomel, P.; Schwartz, O.; Spaink, H.P.; Levraud, J.-P.; Boudinot, P. Contrasted Innate Responses to Two Viruses in Zebrafish: Insights into the Ancestral Repertoire of Vertebrate IFN-Stimulated Genes. J. Immunol. 2014, 192, 4328–4341. [Google Scholar] [CrossRef] [Green Version]
- Levraud, J.-P.; Boudinot, P.; Colin, I.; Benmansour, A.; Peyrieras, N.; Herbomel, P.; Lutfalla, G. Identification of the Zebrafish IFN Receptor: Implications for the Origin of the Vertebrate IFN System. J. Immunol. 2007, 178, 4385–4394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Muñoz, A.; Roca, F.J.; Sepulcre, M.P.; Meseguer, J.; Mulero, V. Zebrafish Larvae Are Unable to Mount a Protective Antiviral Response against Waterborne Infection by Spring Viremia of Carp Virus. Dev. Comp. Immunol. 2010, 34, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Espín-Palazón, R.; Martínez-López, A.; Roca, F.J.; López-Muñoz, A.; Tyrkalska, S.D.; Candel, S.; García-Moreno, D.; Falco, A.; Meseguer, J.; Estepa, A.; et al. TNFα Impairs Rhabdoviral Clearance by Inhibiting the Host Autophagic Antiviral Response. PLoS Pathog. 2016, 12, e1005699. [Google Scholar] [CrossRef] [Green Version]
- Phelan, P.E.; Mellon, M.T.; Kim, C.H. Functional Characterization of Full-Length TLR3, IRAK-4, and TRAF6 in Zebrafish (Danio Rerio). Mol. Immunol. 2005, 42, 1057–1071. [Google Scholar] [CrossRef]
- Novoa, B.; Romero, A.; Mulero, V.; Rodríguez, I.; Fernández, I.; Figueras, A. Zebrafish (Danio Rerio) as a Model for the Study of Vaccination against Viral Haemorrhagic Septicemia Virus (VHSV). Vaccine 2006, 24, 5806–5816. [Google Scholar] [CrossRef]
- Gong, X.-Y.; Zhang, Q.-M.; Zhao, X.; Li, Y.-L.; Qu, Z.-L.; Li, Z.; Dan, C.; Gui, J.-F.; Zhang, Y.-B. LGP2 Is Essential for Zebrafish Survival through Dual Regulation of IFN Antiviral Response. iScience 2022, 25, 104821. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, Q.; Wang, J.; Tian, J.; Song, Y.; Xie, H.; Chang, M.; Nie, P.; Gao, Q.; Zou, J. Transcriptomic Responses of S100 Family to Bacterial and Viral Infection in Zebrafish. Fish Shellfish Immunol. 2019, 94, 685–696. [Google Scholar] [CrossRef]
- Nag, D.; Farr, D.; Raychaudhuri, S.; Withey, J.H. An Adult Zebrafish Model for Adherent-Invasive Escherichia Coli Indicates Protection from AIEC Infection by Probiotic E. Coli Nissle. iScience 2022, 25, 104572. [Google Scholar] [CrossRef] [PubMed]

| Tool Name & Category | Target Component | Application/Target Type | References |
|---|---|---|---|
| Transgenic lines | |||
| Tg(6xNF-kBre:eGFP)nc1 | NFκB activation | Monitoring of the temporal and spatial patterns of NFκB activation | [73] |
| Tg(asc:asc-eGFP) | Inflammasome activation | Visualization of cells with inflammasome activated, CRISPR knock-in line in which the endogenous ASC is fused to eGFP | [74,75] |
| TgBAC(mpx:GFP)i114xTg(lyz:h2az2a-mCherry,cmlc2:GFP) | Neutrophil Extracellular Traps (NETs) | Visualization of neutrophils and neutrophil derived DNA-associated proteins | [76] |
| Tg(ifnφ1:mCherry) | Cells producing IFNφ1 | Reporter line, ifnφ1 promoter | [77] |
| Tg(cryaa:DsRed/mxa:mCherry-F) | Cells stimulated by IFNs | MXA promoter used to drive the specific expression of membrane-targeted mCherry in cells responding to type I IFN | [78] |
| Tg(actb2:GCaMP3) | Ca2+ | Monitoring of Ca2+, using a line in which the GCaMP3.1 indicator is expressed under the control of ubiquitous β-actin promoter | [67,79] |
| Tg(ins:hmgb1-eGFP) | HMGB1 | Dynamics of HMGB1 in β-cells | [80] |
| Tg(lyz:cxcr1-FT) FT: Fluorescent timer | CXCR1 in neutrophils | Constitutive and ligand-induced dynamics of CXCR1 in neutrophils | [81] |
| Tg(lyz:cxcr2-FT) FT: Fluorescent timer | CXCR2 in neutrophils | Constitutive and ligand-induced dynamics of CXCR2 in neutrophils | [81] |
| Tg(cxcl18b:eGFP) | CXCL18b | Reporter line, cxcl18b promoter | [82] |
| Tg(sdf-1a:DsRed2) | CXCL12a/SDF1a | Reporter line, cxcl12a promoter | [83] |
| Tg(cxcr4b:mCherry) | CXCR4b | Reporter line, cxcr4b promoter | [84] |
| Morpholinos (MO) | |||
| MO-tnfα | TNFα | Cytokine | [85] |
| MO-il1b | IL1β | Cytokine | [49,52] |
| MO-tnfrsf1b/tnfrsf1a | TNFα receptors | Cytokine receptor | [86] |
| MO-hmgb1a | HMGB1a | Alarmin | [87,88] |
| MO-cxcl8a | CXCL8a | Chemokine | [89,90] |
| MO-cxcr1 | CXCR1 | Chemokine receptor | [91] |
| MO-cxcr4b | CXCR4b | Chemokine receptor | [92,93] |
| MO-ccl25a | CCL25a | Chemokine | [94] |
| MO-asc | ASC | Inflammasome | [74,75,95] |
| MO-caspa, MO-caspb | CASPa, CASPb | Inflammasome | [75,96,97,98] |
| MO-gsdmea, MO-gsdmea | Gasdermins | Inflammasome | [75,99] |
| MO-nod1, MO-nod2 | NOD1, NOD2 | Inflammasome | [100,101] |
| MO-C3a, MO-C3aR | C3a, C3aR | Complement | [102,103] |
| MO-s100a10a | S100A10a | Alarmin | [104] |
| Mutants and CRISPR Mutants | |||
| tlr2 mutant | TLR2 | Receptor | [105] |
| myd88 mutant | MYD88 | Adaptor protein | [105] |
| il4/13a or il4/13b mutant | IL4/13a & IL4/13b | Interleukin | [106] |
| il10 | IL10 | Interleukin | [106] |
| anxa1c mutant | ANXA1C | Annexin | [107] |
| ifnφ3ip7/ip7 mutant | IFNφ3 | Interferon | [78] |
| cxcr1 mutant | CXCR1 | Chemokine receptor | [81] |
| cxcr2 mutant | CXCR2 | Chemokine receptor | [81] |
| asc mutants | ASC | Inflammasome | [99,108] |
| caspa mutant | CASPa | Inflammasome | [74,75] |
| caspb mutant | CASPb | Inflammasome | [109] |
| s100i2 CRISPR | S100I2 | Alarmin | [110] |
| Recombinant proteins/antibodies | |||
| Recombinant IFNφ1, IFNφ2, IFNφ4 | IFNφ1, IFNφ2, IFNφ4 | Production of recombinant IFNφ and injection in zebrafish for functional studies | [111] |
| Asc antibody | Asc | Inflammasome | [74] |
| Overexpression plasmids | |||
| IFNφ1-pTol2S263C IFNφ2-pTol2S263C IFNφ3-pTol2S263C IFNφ4-pTol2S263C | IFNφ1 IFNφ2 IFNφ3 IFNφ4 | Overexpression of the four IFNφ genes in zebrafish, using the pTol2S263C vector | [111] |
| Fluorescent sensors | |||
| CellRox & acetyl-pentafluorobenzene sulphonyl fluorescein | ROS | ROS-specific probes that fluoresce in the presence of ROS | [53,112] |
| Fluo-3-AM | Ca2+ | Ca-specific fluorescent probe for the detection of intracellular Ca2+ concentration | [53] |
| HyPer mRNA | H2O2 | Genetically encoded ratiometric H2O2 sensor to monitor intracellular H2O2 concentration | [113] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leiba, J.; Özbilgiç, R.; Hernández, L.; Demou, M.; Lutfalla, G.; Yatime, L.; Nguyen-Chi, M. Molecular Actors of Inflammation and Their Signaling Pathways: Mechanistic Insights from Zebrafish. Biology 2023, 12, 153. https://doi.org/10.3390/biology12020153
Leiba J, Özbilgiç R, Hernández L, Demou M, Lutfalla G, Yatime L, Nguyen-Chi M. Molecular Actors of Inflammation and Their Signaling Pathways: Mechanistic Insights from Zebrafish. Biology. 2023; 12(2):153. https://doi.org/10.3390/biology12020153
Chicago/Turabian StyleLeiba, Jade, Resul Özbilgiç, Liz Hernández, Maria Demou, Georges Lutfalla, Laure Yatime, and Mai Nguyen-Chi. 2023. "Molecular Actors of Inflammation and Their Signaling Pathways: Mechanistic Insights from Zebrafish" Biology 12, no. 2: 153. https://doi.org/10.3390/biology12020153