
Adonis fucensis (A. sect. Adonanthe, Ranunculaceae), a New Species from the Central Apennines (Italy)
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Morphometric Analyses
2.3. DNA Sequencing and AFLPseq Fingerprinting
3. Results
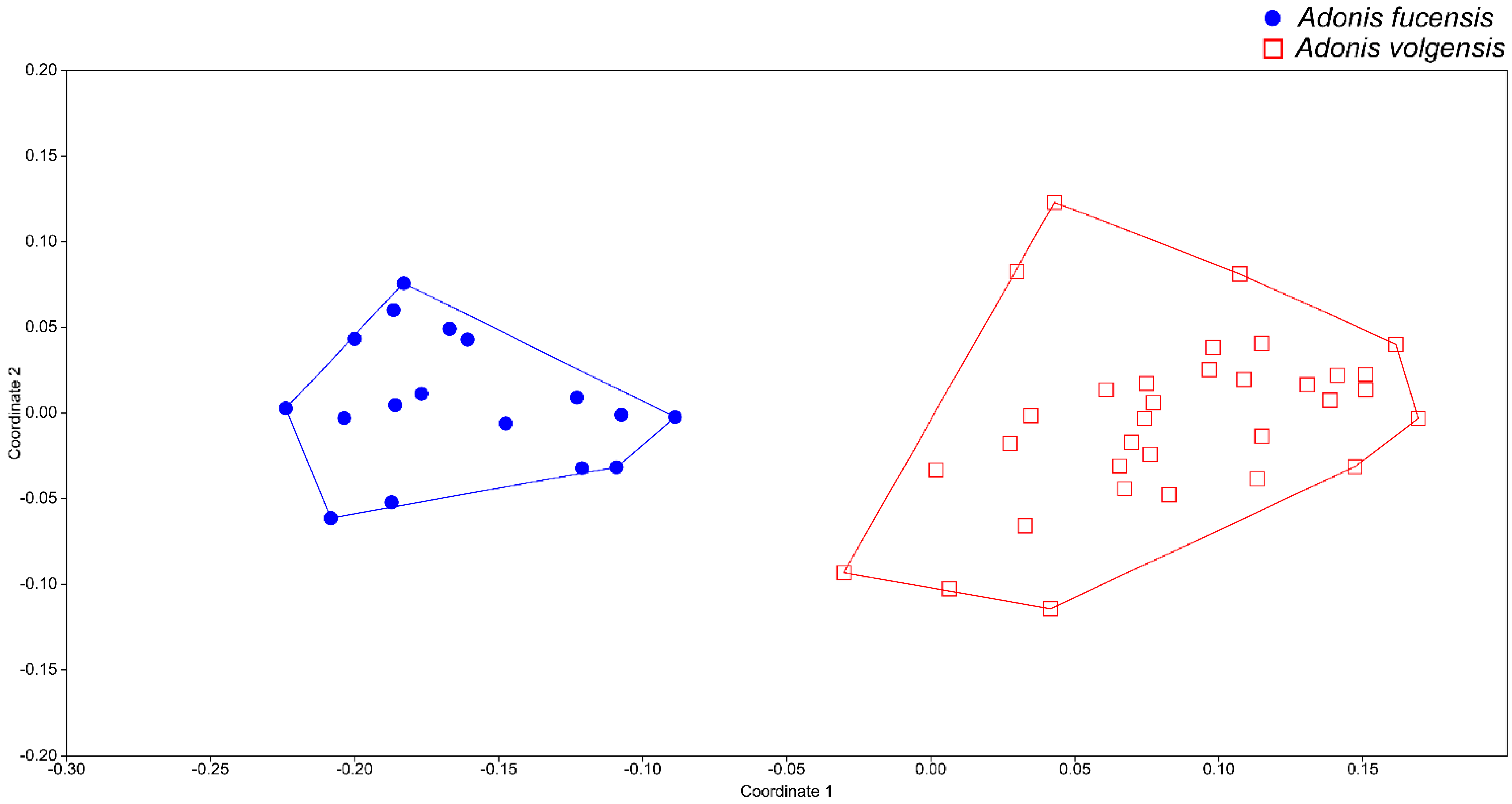
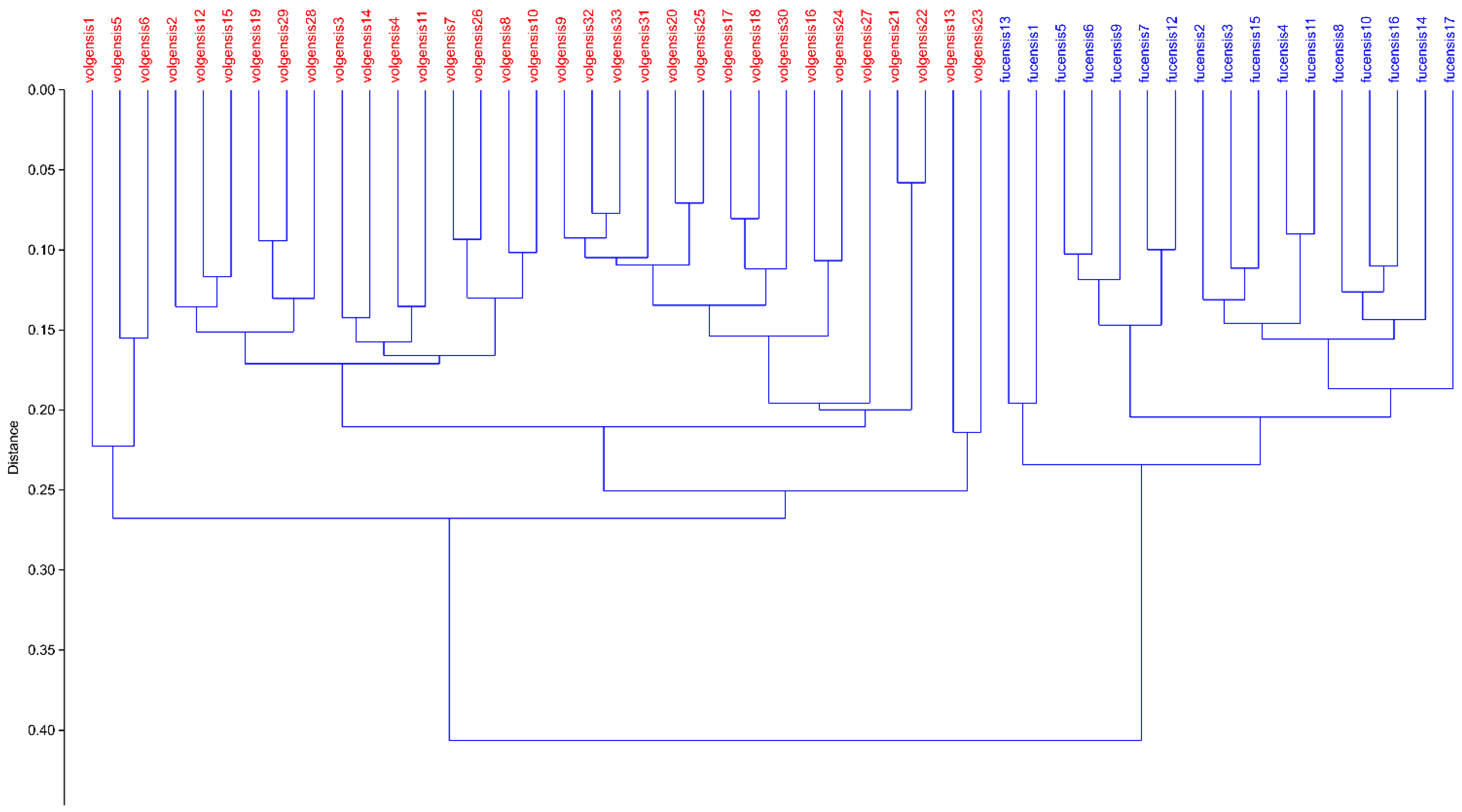
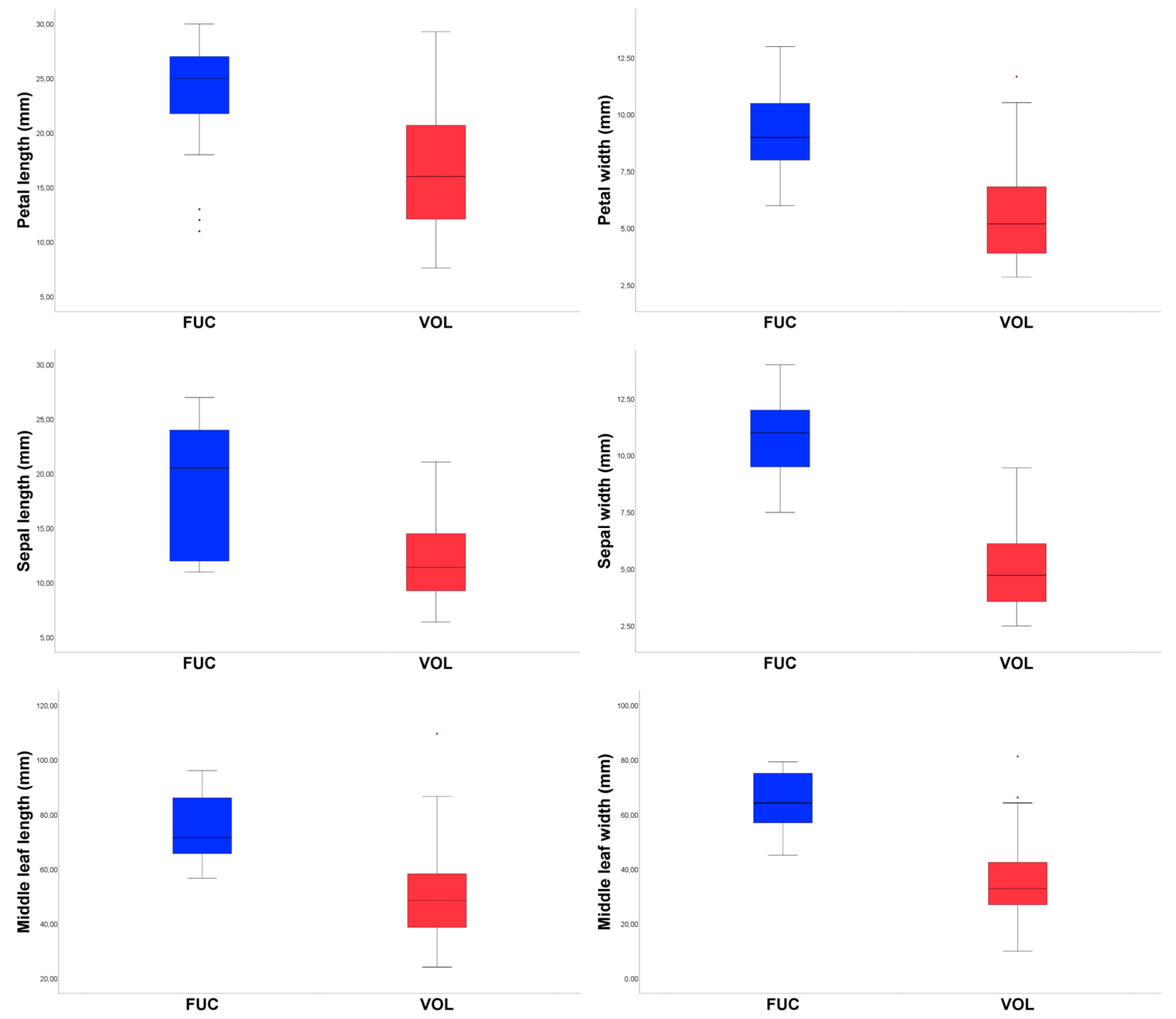
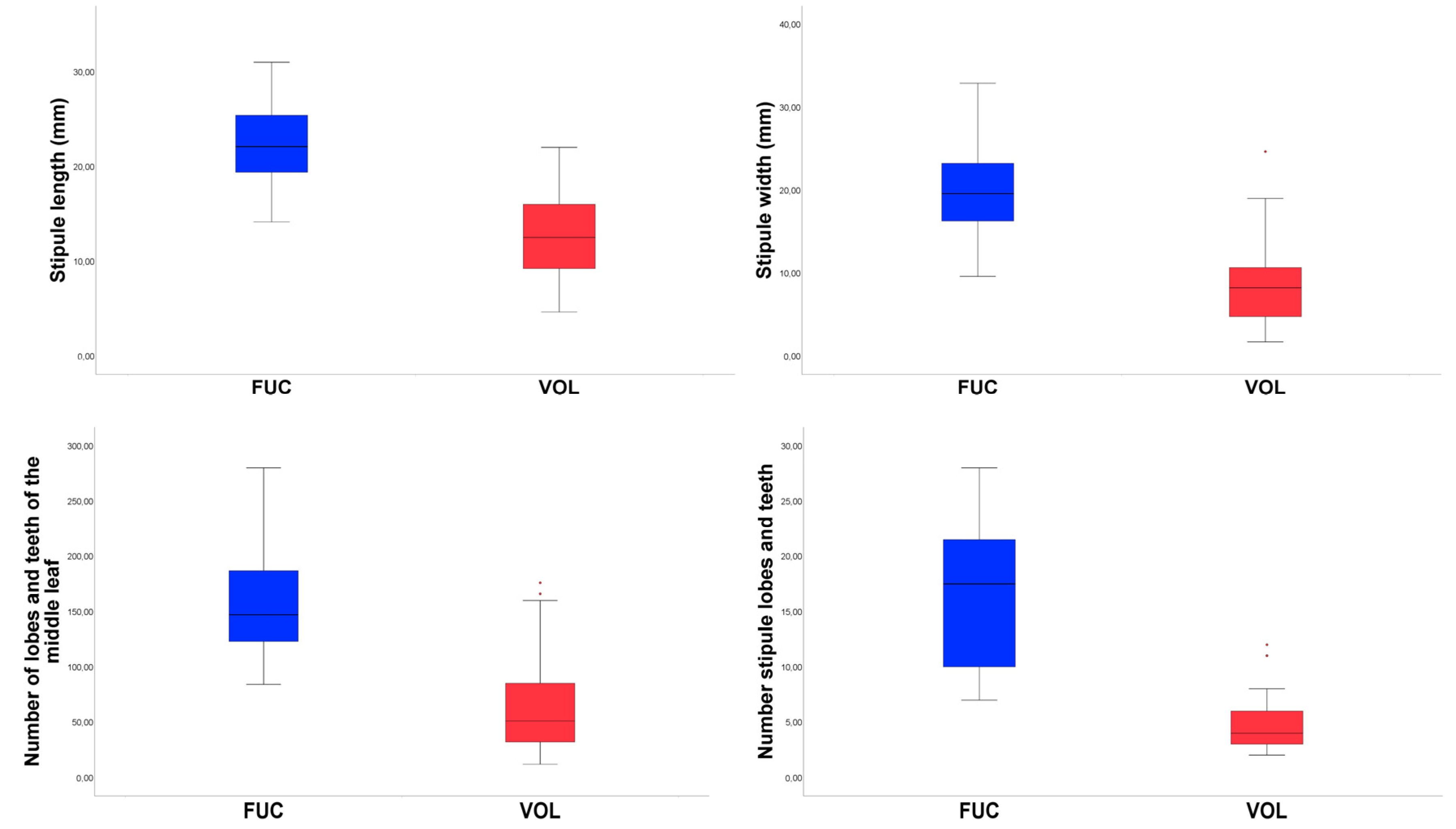
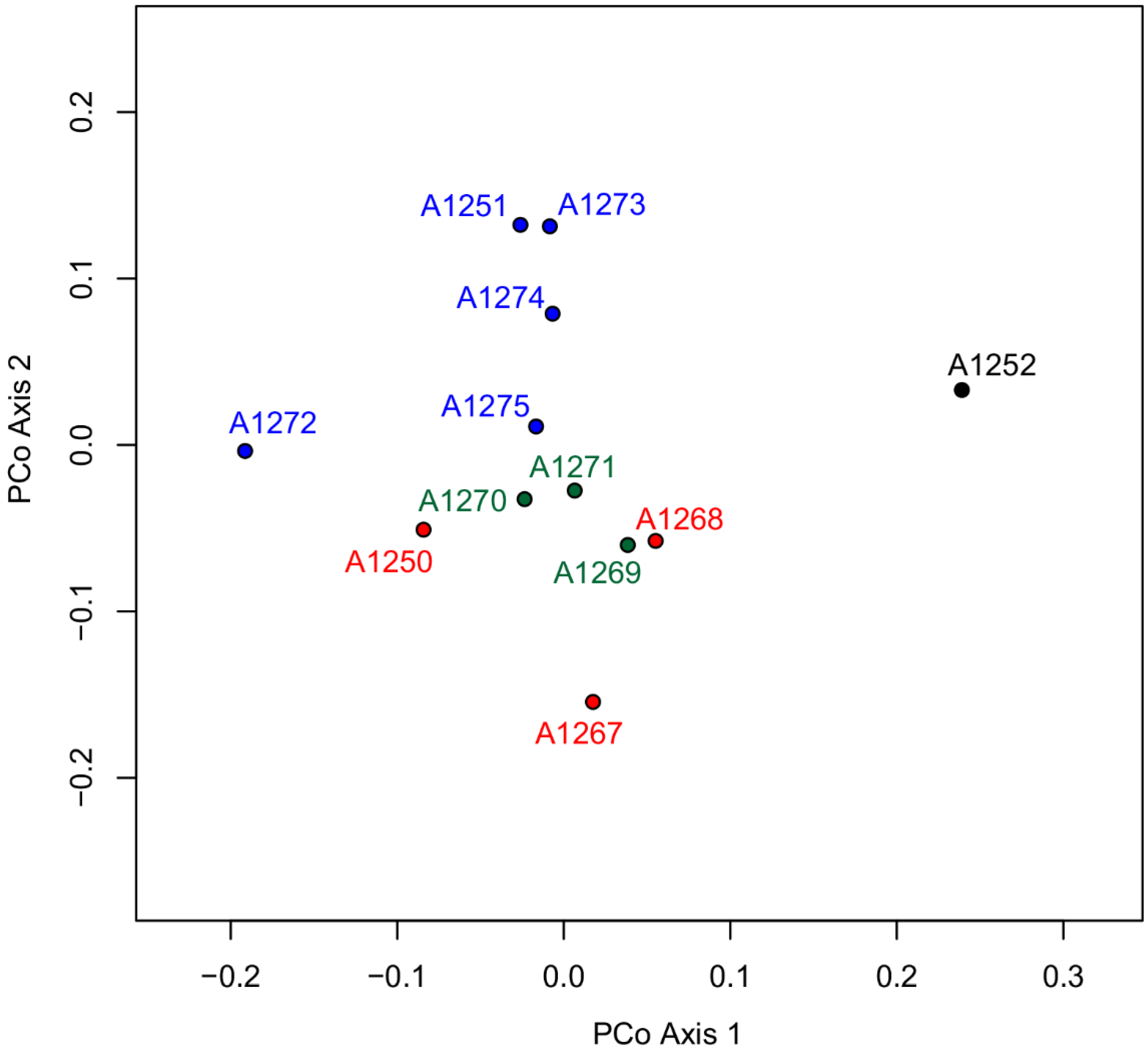
3.1. Morphometric Analyses
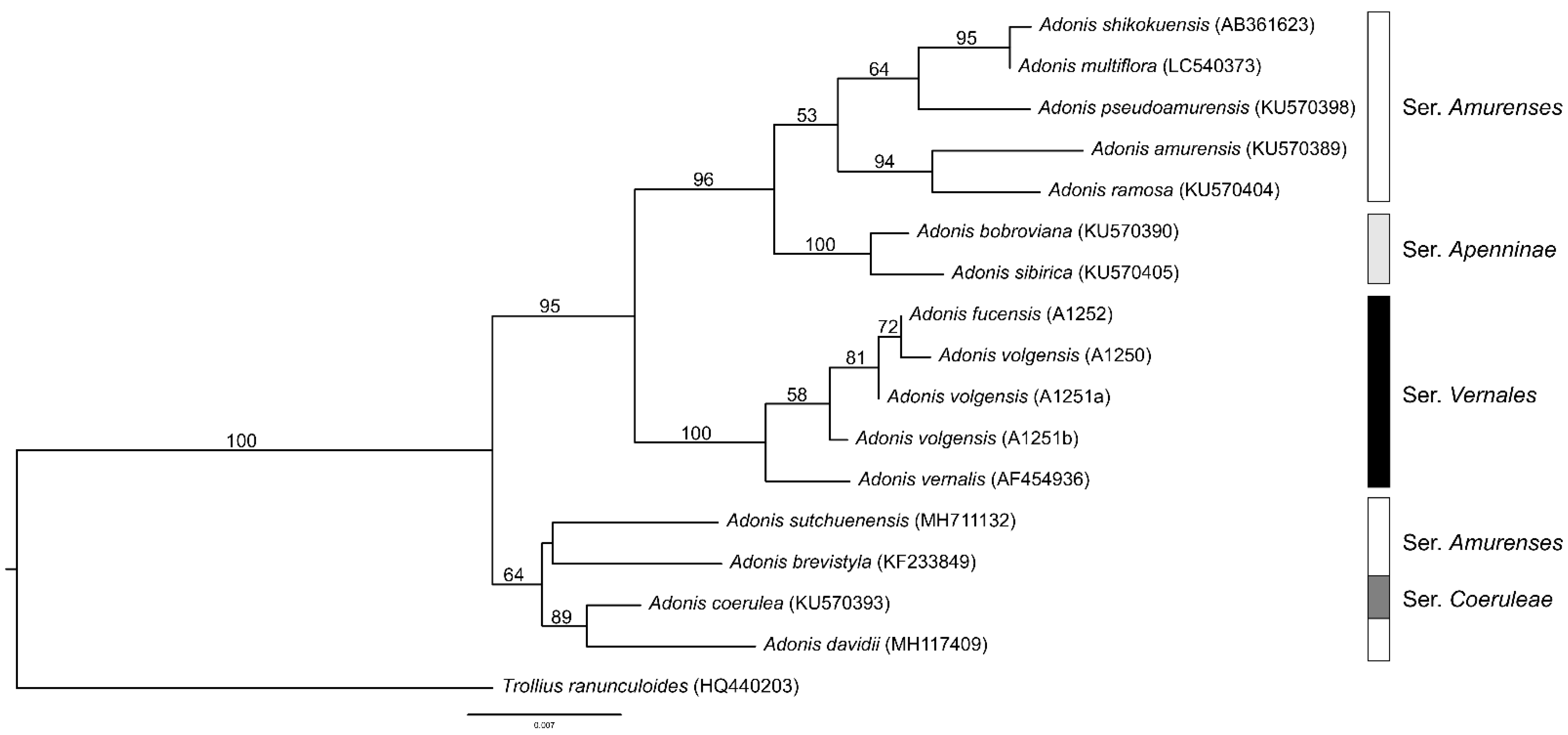
3.2. nrDNA Sequence Variation
3.3. AFLPseq Fingerprinting
4. Discussion
5. Taxonomic Treatment
Key to Adonis Species Belonging to sect. Adonanthe Distributed in Europe
- 1. Leaves pinnately compound …………………………………………………... A. apennina
- 1. Leaves palmately compound …………………………………………………….………… 2
- 2. Leaves glabrous, with narrowly linear, entire lobes, rarely few-dentate ........ A. vernalis
- 2. Leaves pubescent, rarely glabrescent, with linear-lanceolate to lanceolate, dentate lobes …………………………………………………………………………………………………….. 3
- 3. Middle cauline leaves triangular-ovate, rarely pentagonal, (24.12)38.70–58.35(109.66) mm long × (10.00)27.00–42.60(81.30) mm wide, with (12)32–85(176) lobes and teeth linear to narrowly lanceolate; stipule (4.64)9.31–15.88(22.00) mm long, with (2–)3–6(–12) lobes and teeth; sepal (6.40)9.26–14.51(21.07) mm long × (2.50)3.57–6.05(9.46) mm wide, petal (7.61)12.12–20.69(29.28) mm long × (2.85)3.95–6.83(11.68) mm wide ………………….………………………………………………………..………..…. A. volgensis
- 3. Middle cauline leaves pentagonal, (56.70)65.75–86.22(96.06) mm long × (45.16)57.01–75.06(79.30) mm wide, with (84)123–187(280) lobes and teeth lanceolate; stipule (14.13)19.83–25.23(31.00) with 10–22.4(–26) lobes and teeth; sepal (11.00)12.25–23.63(27.00) mm long × (7.50)9.63–12.00(14.00) mm wide, petal (11.00)21.75–27.00(30.00) mm long × (6.00)8.00–10.50(13.00) mm wide …………………………………………………… A. fucensis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- POWO. Plants of the World Online, Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 10 October 2022).
- Wang, W.T. Revision of Adonis (Ranunculaceae) I. Bull. Bot. Res. 1994, 14, 1–31. [Google Scholar]
- Wang, W.T. Revision of Adonis (Ranunculaceae) II. Bull. Bot. Res. 1994, 14, 105–138. [Google Scholar]
- Son, D.C.; Park, B.K.; Ko, S.C. Phylogenetic study of the section Adonanthe of genus Adonis L. (Ranunculaceae) based on ITS sequences. Korean J. Plant Taxon. 2016, 46, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Seidl, A.; Tremetsberger, K.; Pfanzelt, S.; Lindhuber, L.; Kropf, M.; Neuffer, B.; Blattner, F.R.; Király, G.; Smirnov, S.V.; Friesen, N.; et al. Genotyping-by-sequencing reveals range expansion of Adonis vernalis (Ranunculaceae) from Southeastern Europe into the zonal Euro-Siberian steppe. Sci. Rep. 2022, 12, 19074. [Google Scholar] [CrossRef] [PubMed]
- Peruzzi, L.; Conti, F.; Bartolucci, F. An inventory of vascular plants endemic to Italy. Phytotaxa 2014, 168, 1–75. [Google Scholar] [CrossRef]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Bartolucci, F.; Stinca, A.; Conti, F. Typification of the name Adonis distorta (Ranunculaceae). Phytotaxa 2021, 523, 264–268. [Google Scholar] [CrossRef]
- Conti, F.; Bartolucci, F. The vascular flora of Gran Sasso and Monti della Laga National Park (Central Italy). Phytotaxa 2016, 256, 1–119. [Google Scholar] [CrossRef]
- Conti, F.; Bartolucci, F. The Vascular Flora of the National Park of Abruzzo, Lazio and Molise (Central Italy): An Annotated Checklist; Geobotany Studies; Springer: Cham, Switzerland, 2015; 254p. [Google Scholar]
- Conti, F.; Bartolucci, F. La Flora Vascolare del Parco Nazionale d’Abruzzo, Lazio e Molise; FastEdit: Acquaviva Picena, Italy, 2022. [Google Scholar]
- Filibeck, G.; Cancellieri, L.; Sperandii, M.G.; Belonovskaya, E.A.; Sobolev, N.; Tsarevskaya, N.; Becker, T.; Berastegi, A.; Bückle, C.; Che, R.; et al. Biodiversity patterns of dry grasslands in the Central Apennines (Italy) along a precipitation gradient: Experiences from the 10th EDGG Field Workshop. Bull. Eurasian Dry Grassl. Group 2018, 36, 25–41. [Google Scholar] [CrossRef]
- Filibeck, G.; Cancellieri, L.; Rosati, L. Carta dei Tipi di Habitat di Interesse Comunitario (Direttiva 92/43/CEE) del Parco Nazionale d’Abruzzo, Lazio e Molise e dei siti Natura 2000: ZSC IT7110205, ZPS IT7120132, ZSC IT6050018, ZSC IT6050020, ZSC IT7212121—Note Illustrative; Università degli Studi della Tuscia: Viterbo, Italy, 2022; 114p. [Google Scholar]
- Conti, F.; Cancellieri, L.; Cangelmi, G.; Filibeck, G.; Rosati, L.; Bartolucci, F. New records of native and alien vascular plants from Abruzzo, Lazio and Molise National Park (Italy)—And additions to the flora of Abruzzo and Molise administrative regions. Ann. Bot. 2022, 12, 23–34. [Google Scholar] [CrossRef]
- Cancellieri, L.; Sperandii, M.G.; Filibeck, G. First record of the steppic relict Astragalus exscapus L. subsp. exscapus in the Apennines (Abruzzo, Italy), and biogeographic implications. Plant Biosyst. 2017, 151, 944–948. [Google Scholar] [CrossRef]
- Filibeck, G.; Cancellieri, L.; Bartolucci, F.; Becker, U.; Conti, F.; Maestri, S.; Mürz, M.; Schommer, E.; Sperandii, M.G.; Becker, T. Festuca valesiaca Schleich. ex Gaudin newly discovered in the Central Apennines (Italy): A further example of steppe relict in the Abruzzo “dry valleys”. Plant Biosyst. 2020, 154, 593–600. [Google Scholar] [CrossRef]
- Raab-Straube, E.V.; Hand, R.; Hörandl, E.; Nardi, E. Ranunculaceae. Available online: https://europlusmed.org/ (accessed on 27 November 2022).
- Akeroyd, J.R. Adonis L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1993; Volume 1, pp. 267–269. [Google Scholar]
- WFO. Adonis transsilvanica Simonovich. Available online: http://www.worldfloraonline.org/taxon/wfo-0000521284 (accessed on 15 December 2022).
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York Botanical Garden’s Virtual Herbarium. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 10 October 2022).
- Son, D.C.; Park, B.K.; Chang, K.S.; Choi, K.; Shin, C.H. Cladistic analysis of the section Adonanthe under genus Adonis L. (Ranunculaceae) from East Asia. J. Asia-Pac. Biodivers. 2017, 10, 232–236. [Google Scholar] [CrossRef]
- IBM. IBM SPSS Statistics for Windows, Version 25.0; IBM Corp.: Armonk, NY, USA, 2017. [Google Scholar]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Gower, J.C. A general coefficient of similarity and some of its properties. Biometrics 1971, 27, 857–874. [Google Scholar] [CrossRef]
- Doyle, J.; Dickson, E. Preservation of plant samples for DNA restriction endonuclease analysis. Taxon 1987, 36, 715–722. [Google Scholar] [CrossRef]
- Doyle, J.; Doyle, J. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Rydin, C.; Pedersen, K.R.; Friis, E.M. On the evolutionary history of Ephedra: Cretaceous fossils and extant molecules. Proc. Natl. Acad. Sci. USA 2004, 101, 16571–16576. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Gelfand, D.H., Sninsky, J.J., White, T.J., Innis, M.A., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Blattner, F.R. Direct amplification of entire ITS region from poorly preserved plant material using recombinant PCR. BioTechniques 1999, 27, 1180–1186. [Google Scholar] [CrossRef]
- Blattner, F.R.; Weising, K.; Bänfer, G.; Maschwitz, U.; Fiala, B. Molecular Analysis of Phylogenetic Relationships among Myrmecophytic Macaranga Species (Euphorbiaceae). Mol. Phylogenet. Evol. 2001, 19, 331–344. [Google Scholar] [CrossRef]
- Technelysium Pty Ltd. CHROMAS (Version 2.6.6). Available online: http://technelysium.com.au/wp/chromas (accessed on 15 October 2022).
- Swofford, D.L. PAUP* (Version PAUP* v.4.0a169). Phylogenetic Analysis Using Parsimony (*and Other Methods). Available online: http://phylosolutions.com/paup-test (accessed on 15 October 2022).
- Dorfner, M.; Ott, T.; Ott, P.; Oberprieler, C. Long-read genotyping with SLANG (Simple Long-read loci Assembly of Nanopore data for Genotyping). Appl. Plant Sci. 2022, 10, e11484. [Google Scholar] [CrossRef] [PubMed]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Friters, A.; Pot, J.; Paleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Barker, G. Landscape and Society: Prehistoric Central Italy (Studies in Archaeology); Academic Press: London, UK, 1981. [Google Scholar]
- Manzi, A. Storia Dell’ambiente nell’Appennino Centrale: La Trasformazione della Natura in Abruzzo Dall’ultima Glaciazione ai Giorni nostri; Meta: Treglio, Italy, 2012; 316p. [Google Scholar]
- Primi, R.; Filibeck, G.; Amici, A.; Bückle, C.; Cancellieri, L.; Di Filippo, A.; Gentile, C.; Guglielmino, A.; Latini, R.; Mancini, L.D.; et al. From Landsat to leafhoppers: A multidisciplinary approach for sustainable stocking assessment and ecological monitoring in mountain grasslands. Agric. Ecosyst. Environ. 2016, 234, 118–133. [Google Scholar] [CrossRef]
- Hejcman, M.; Hejcmanová, P.; Pavlů, V.; Beneš, J. Origin and history of grasslands in Central Europe—A review. Grass Forage Sci. 2013, 68, 345–363. [Google Scholar] [CrossRef]
- Grossi, G. Il territorio di Casali d’Aschi dall’antichita al medioevo. In Casali d’Aschi Ieri e Oggi; Grossi, G., Palozzi, L., Amendola, V., Melchiorre, A., Eds.; Edizioni dell’Urbe: Rome, Italy, 1983; pp. 11–57. [Google Scholar]
- Morretti, F.; Puppi, G.; Giuliani, C.; Conti, F. Heterostyly in Goniolimon italicum (Plumbaginaceae), endemic to Abruzzo (central Apennines, Italy). An. Jard. Bot. Madr. 2015, 72, e014. [Google Scholar] [CrossRef] [Green Version]
- Buzurović, U.; Tomović, G.; Niketić, M.; Bogdanović, S.; Aleksić, J.M. Phylogeographic and taxonomic considerations on Goniolimon tataricum (Plumbaginaceae) and its relatives from south-eastern Europe and the Apennine Peninsula. Plant Syst. Evol. 2020, 306, 29. [Google Scholar] [CrossRef]
- Anzalone, B. Su un nuovo Astragalus scoperto in Abruzzo. Webbia 1970, 24, 723–734. [Google Scholar] [CrossRef]
- Bachman, S.; Moat, J.; Hill, A.W.; de la Torre, J.; Scott, B. Supporting Red List threat assessments with GeoCAT: Geospatial conservation assessment tool. ZooKeys 2011, 150, 117–126. [Google Scholar] [CrossRef]
- IUCN. IUCN Guidelines for Using the IUCN Red List Categories and Criteria. Version 15.1. Available online: https://www.iucnredlist.org/resources/redlistguidelines (accessed on 23 September 2022).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Description of the Character | Type |
|---|---|---|
| H | Height (cm) | quantitative continuous |
| MLL | Middle leaf length (mm) | quantitative continuous |
| MLW | Middle leaf width (mm) | quantitative continuous |
| SML | Shape of middle leaf (0: pentagonal; 1: triangular-ovate) | qualitative |
| NLML | Number of lobes and teeth of the middle leaf | quantitative discrete |
| STL | Shape of terminal lobe (0: lanceolate; 1: narrowly lanceolate; 2: linear) | qualitative |
| ATL | Angle of terminal apex lobe (°) | quantitative continuous |
| LMW | Subterminal lobe max width (mm) | quantitative continuous |
| LWB | Subterminal lobe width at base (mm) | quantitative continuous |
| LMW/LWB | Ratio lobe max width/lobe width at base | calculated ratio |
| SL | Stipule length (mm) | quantitative continuous |
| SW | Stipule width (mm) | quantitative continuous |
| NSL | Number stipule lobes and teeth | quantitative discrete |
| SLD | Sepal length (mm) | quantitative continuous |
| SWD | Sepal width (mm) | quantitative continuous |
| PLD | Petal length (mm) | quantitative continuous |
| PWD | Petal width (mm) | quantitative continuous |
| NP | Number of petals | quantitative discrete |
| Adonis volgensis | Adonis fucensis | |||
|---|---|---|---|---|
| Height (cm) | 22.80 ± 7.21 | (10.49)17.42–27.39(39.61) | 12.06 ± 2.46 | (8)10.25–13.75(16) |
| Middle leaf length (mm) | 50.36 ± 16.86 | (24.12)38.70–58.35(109.66) | 74.28 ± 12.39 | (56.70)65.75–86.22(96.06) |
| Middle leaf width (mm) | 35.01 ± 13.75 | (10.00)27.00–42.60(81.30) | 64.31 ± 10.83 | (45.16)57.01–75.06(79.30) |
| Shape of middle leaf | triangular-ovate, rarely pentagonal | pentagonal | ||
| Number of lobes and teeth of the middle leaf | (12)32–85(176) | (84)123–187(280) | ||
| Shape of terminal lobe | linear to narrowly lanceolate | lanceolate | ||
| Angle of terminal apex lobe (°) | 31.95 ± 11.21 | (15.04)23.73–37.55(64.96) | 50.45 ± 11.76 | (31.73)43.36–54.32(74.4) |
| Subterminal lobe max width (mm) | 1.13 ± 0.39 | (0.53)0.84–1.34(2.39) | 1.76 ± 0.48 | (1.14)1.41–1.91(2.83) |
| Subterminal lobe width at base (mm) | 0.94 ± 0.28 | (0.51)0.70–1.10(1.75) | 1.36 ± 0.38 | (0.90)1.08–1.49(2.29) |
| Ratio lobe max width/lobe width at base | 1.21 ± 0.27 | (0.91)1.02–1.32(2.52) | 1.29 ± 0.11 | (1.13)1.21–1.37(1.57) |
| Stipule length (mm) | 12.63 ± 4.36 | (4.64)9.31–15.88(22.00) | 22.41 ± 4.53 | (14.13)19.83–25.23(31.00) |
| Stipule width (mm) | 8.45 ± 4.82 | (1.69)4.85–10.60(24.63) | 22.06 ± 5.56 | (9.59)16.40–23.24(32.87) |
| Number of stipule lobes and teeth | (2–)3–6(–12) | (7)10–21(–28) | ||
| Sepal length (mm) | 12.23 ± 3.67 | (6.40)9.26–14.51(21.07) | 18.81 ± 5.80 | (11.00)12.25–23.63(27.00) |
| Sepal width (mm) | 4.98 ± 1.71 | (2.50)3.57–6.05(9.46) | 10.61 ± 1.82 | (7.50)9.63–12.00(14.00) |
| Petal length (mm) | 17.2 ± 6.04 | (7.61)12.12–20.69(29.28) | 23.48 ± 5.54 | (11.00)21.75–27.00(30.00) |
| Petal width (mm) | 5.59 ± 2.15 | (2.85)3.95–6.83(11.68) | 9.37 ± 1.96 | (6.00)8.00–10.50(13.00) |
| Number of petals | (10)12–13(16) | (9)12–14(18) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conti, F.; Oberprieler, C.; Dorfner, M.; Schabel, E.; Nicoară, R.; Bartolucci, F. Adonis fucensis (A. sect. Adonanthe, Ranunculaceae), a New Species from the Central Apennines (Italy). Biology 2023, 12, 118. https://doi.org/10.3390/biology12010118
Conti F, Oberprieler C, Dorfner M, Schabel E, Nicoară R, Bartolucci F. Adonis fucensis (A. sect. Adonanthe, Ranunculaceae), a New Species from the Central Apennines (Italy). Biology. 2023; 12(1):118. https://doi.org/10.3390/biology12010118
Chicago/Turabian StyleConti, Fabio, Christoph Oberprieler, Marco Dorfner, Erik Schabel, Roxana Nicoară, and Fabrizio Bartolucci. 2023. "Adonis fucensis (A. sect. Adonanthe, Ranunculaceae), a New Species from the Central Apennines (Italy)" Biology 12, no. 1: 118. https://doi.org/10.3390/biology12010118