Gesneriads, a Source of Resurrection and Double-Tolerant Species: Proposal of New Desiccation- and Freezing-Tolerant Plants and Their Physiological Adaptations


Abstract
:Simple Summary
Abstract
1. Introduction
2. Species Identification
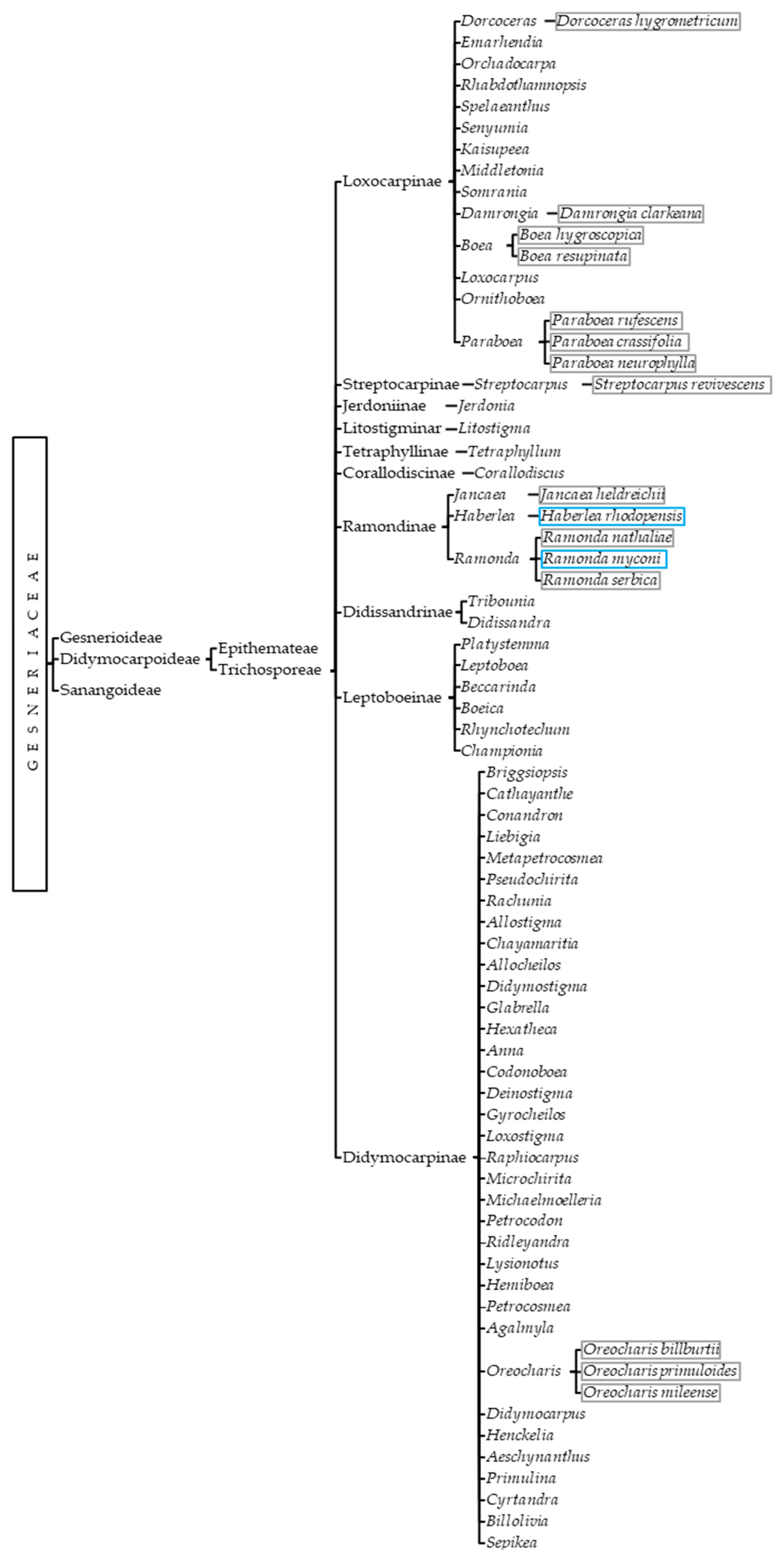
2.1. Phylogeny
- Sanangoideae, which did not officially become a subfamily until 2013, when it was defined as a monotypic family (Sanango racemosum) endemic to South America. It has a distinctive globose and slightly four-partite ovary with a depression in the internal structure and on top, from which the style arises surrounded by a large cupular disc [1].
- Gesnerioideae, which was initially named New World (NW) Gesneriaceae, since it was thought to contain only neotropical species. However, nowadays it also contains gesneriads from Asia and Australia, and it is considered a heterogeneous group with a well-established phylogeny [1,10]. It is generally characterized by the presence of seed endosperm, two equally sized cotyledons with limited growth, a nectary with separated glands, and an inferior ovary [11].
- Didymocarpoideae, which has been typically recognized as Old World (OW) Gesneriaceae; in this case, with the exception of Rhynchoglossum azureum, species of this subfamily are indeed found in Asia, Africa, and Europe [12,13]. The intrinsic morphological characteristics of the subfamily include the lack of endosperm, unequal cotyledon growth, ring-shaped nectary, and superior ovary [11].
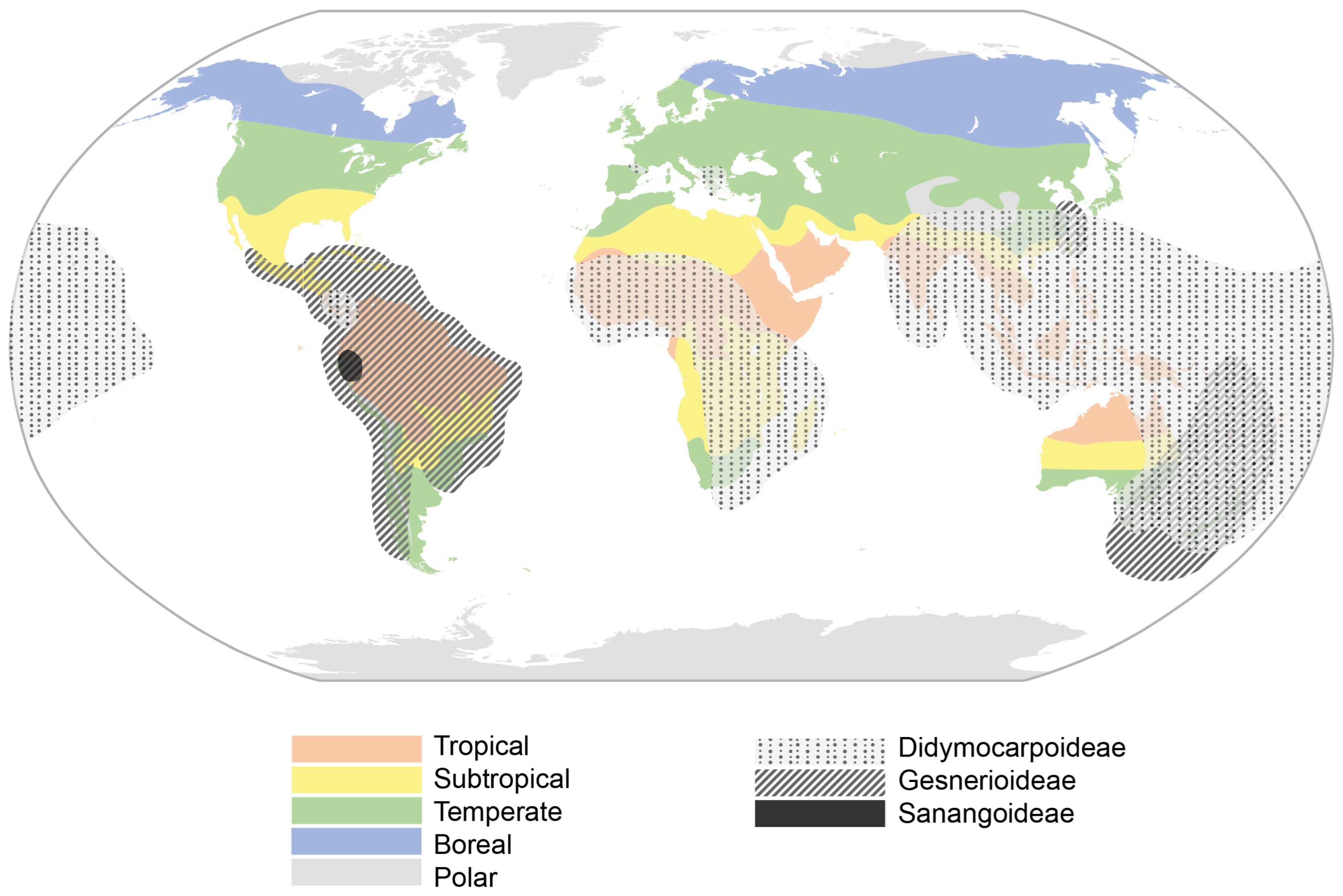
2.2. Geographic Distribution and Habitat
2.2.1. Origin and Geographic Evolution of Gesneriads
2.2.2. Current Distribution and Habitat
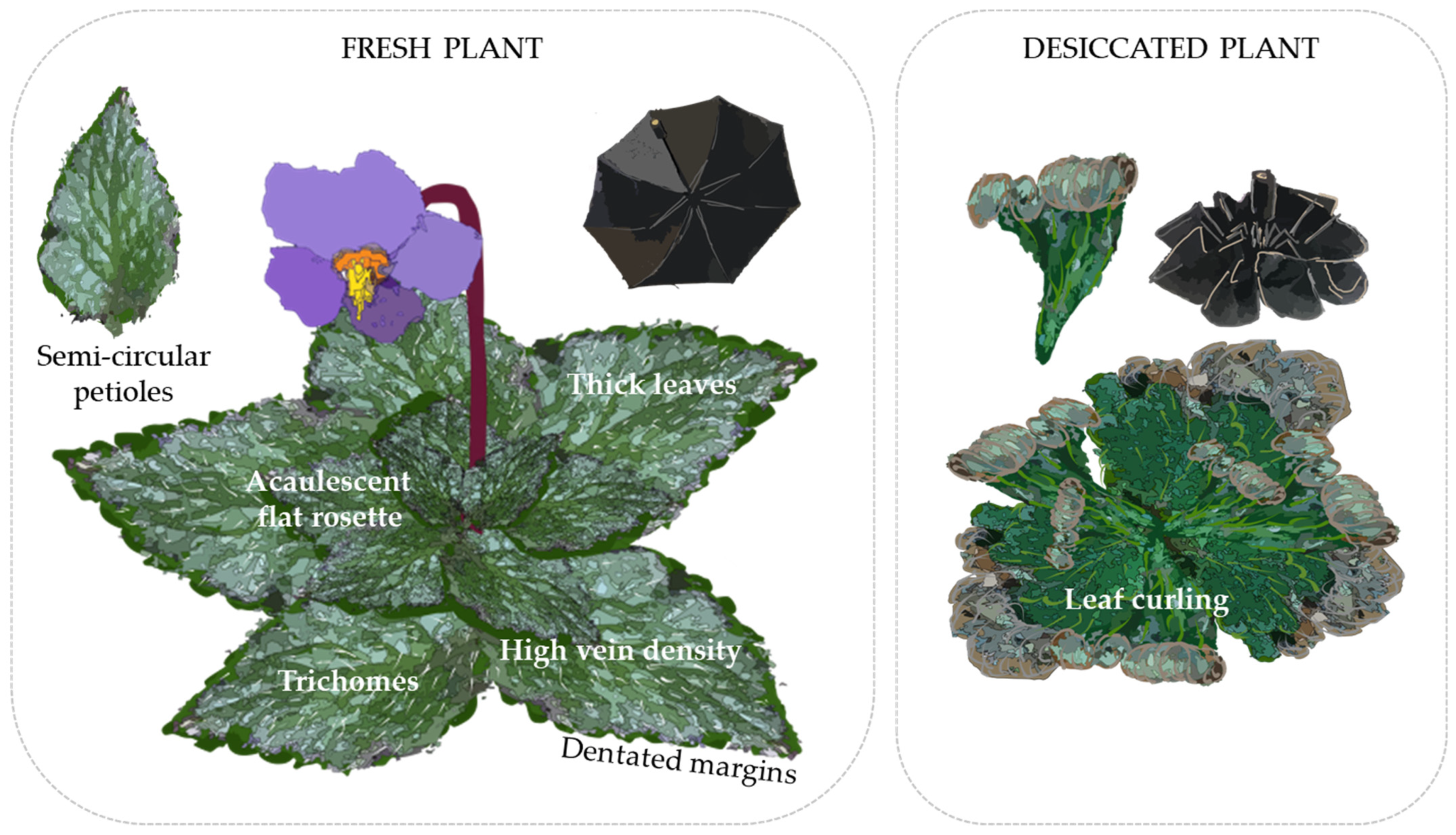
2.3. Morphological Characterization
2.3.1. Adaptation Pressures
2.3.2. Morphological Desiccation and Freezing Tolerance Traits
2.4. Tentative Desiccation/Double-Tolerant Species
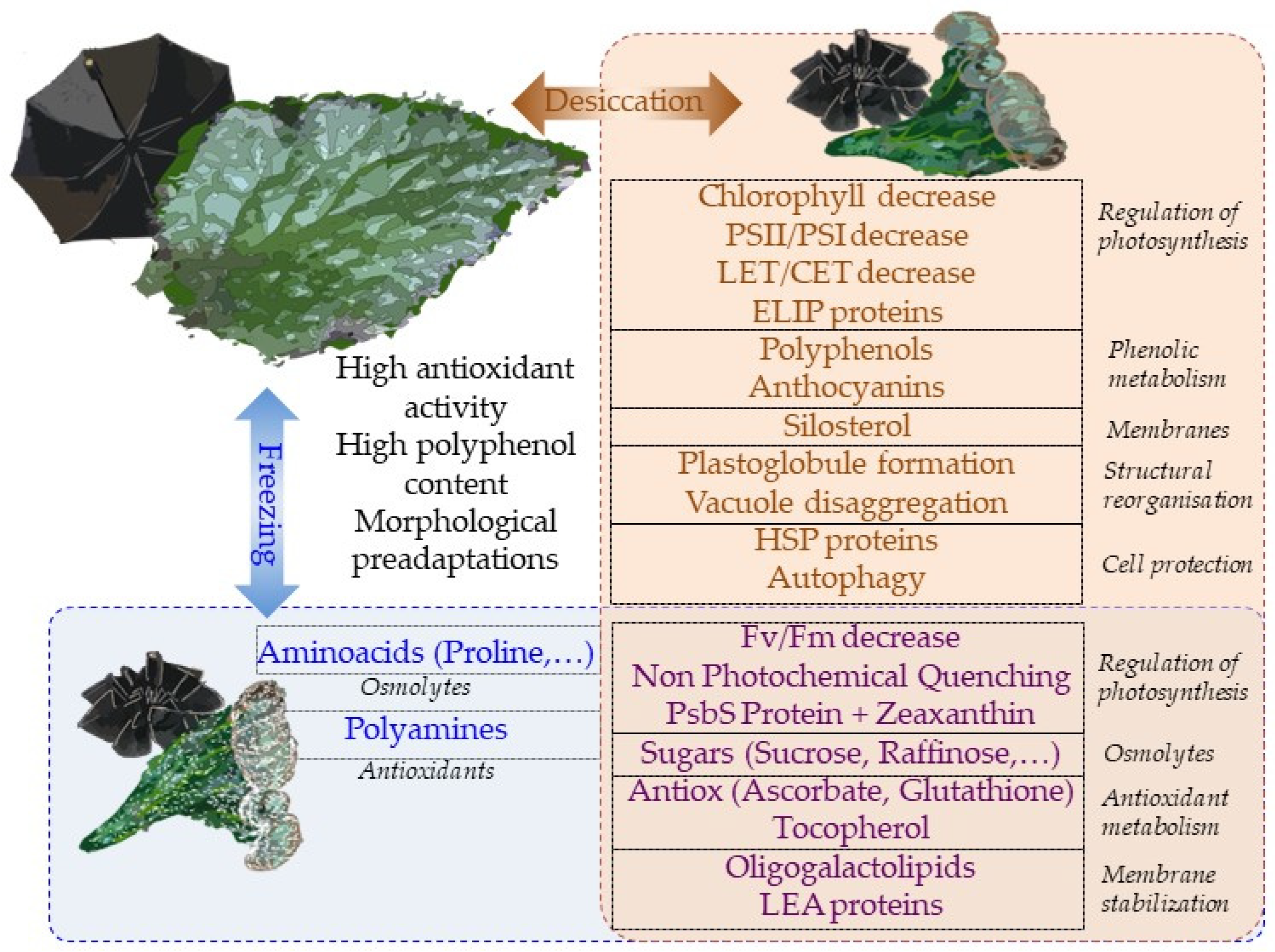
3. Physiological Adaptations for Desiccation and Freezing Tolerance
3.1. Desiccation Tolerance Strategies among Gesneriads
3.2. Avoiding Reactive Oxygen Species Formation
3.3. Avoiding Structural Damage
3.4. Cellular Protection
3.5. Freezing-Induced Desiccation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weber, A.; Clark, J.L.; Möller, M. A New Formal Classification of Gesneriaceae. Selbyana 2013, 31, 68–94. [Google Scholar]
- Marks, R.A.; Farrant, J.M.; Nicholas McLetchie, D.; VanBuren, R. Unexplored Dimensions of Variability in Vegetative Desiccation Tolerance. Am. J. Bot. 2021, 108, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Weber, A. Gesneriaceae. In The Families and Genera of Vascular Plants; Kulbitzki, K., Ed.; Springer: Berlin, Germany, 2004; Volume 7, pp. 63–158. [Google Scholar]
- Verhoeven, A.; García-Plazaola, J.I.; Fernández-Marín, B. Shared Mechanisms of Photoprotection in Photosynthetic Organisms Tolerant to Desiccation or to Low Temperature. Environ. Exp. Bot. 2018, 154, 66–79. [Google Scholar] [CrossRef]
- Tebele, S.M.; Marks, R.A.; Farrant, J.M. Two Decades of Desiccation Biology: A Systematic Review of the Best Studied Angiosperm Resurrection Plants. Plants 2021, 10, 2784. [Google Scholar] [CrossRef]
- Clark, J.L.; Funke, M.M.; Duffy, A.M.; Smith, J.F. Phylogeny of a Neotropical Clade in the Gesneriaceae: More Tales of Convergent Evolution. Int. J. Plant Sci. 2012, 173, 894–916. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Serrano, M.L.; Rolland, J.; Clark, J.L.; Salamin, N.; Perret, M. Hummingbird Pollination and the Diversification of Angiosperms: An Old and Successful Association in Gesneriaceae. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162816. [Google Scholar] [CrossRef] [Green Version]
- Roalson, E.H.; Roberts, W.R. Distinct Processes Drive Diversification in Different Clades of Gesneriaceae. Syst. Biol. 2016, 65, 662–684. [Google Scholar] [CrossRef] [Green Version]
- Ogutcen, E.; Christe, C.; Nishii, K.; Salamin, N.; Möller, M.; Perret, M. Phylogenomics of Gesneriaceae Using Targeted Capture of Nuclear Genes. Mol. Phylogenet. Evol. 2021, 157, 107068. [Google Scholar] [CrossRef]
- Möller, M.; Pfosser, M.; Jang, C.G.; Mayer, V.; Clark, A.; Hollingsworth, M.L.; Barfuss, M.H.J.; Wang, Y.Z.; Kiehn, M.; Weber, A. A Preliminary Phylogeny of the “didymocarpoid Gesneriaceae” Based on Three Molecular Data Sets: Incongruence with Available Tribal Classifications. Am. J. Bot. 2009, 96, 989–1010. [Google Scholar] [CrossRef]
- Burtt, B.L.; Wiehler, H. Classification of the Family Gesneriaceae. Gesneriana 1995, 1, 1–4. [Google Scholar]
- Xu, W.; Guo, J.; Pan, B.; Zhang, Q.; Liu, Y. Diversity and Distribution of Gesneriaceae in China. Guihaia 2017, 37, 1219–1226. [Google Scholar] [CrossRef]
- Möller, M.; Wei, Y.G.; Wen, F.; Clark, J.L.; Weber, A. You Win Some You Lose Some: Updated Generic Delimitations and Classification of Gesneriaceae-Implications for the Family in China. Guihaia 2016, 35, 44–60. [Google Scholar] [CrossRef]
- Weber, A.; Middleton, D.J.; Clark, J.L.; Möller, M. Keys to the Infrafamilial Taxa and Genera of Gesneriaceae. J. Indian Assoc. Angiosperm Taxon. 2020, 30, 5–47. [Google Scholar] [CrossRef]
- Tan, K.; Lu, T.; Ren, M.X. Biogeography and Evolution of Asian Gesneriaceae Based on Updated Taxonomy. PhytoKeys 2020, 157, 26. [Google Scholar] [CrossRef] [PubMed]
- Middleton, D.J.; Atkins, H.; Truong, L.H.; Nishii, K.; Möller, M. Billolivia, a New Genus of Gesneriaceae from Vietnam with Five New Species. Phytotaxa 2014, 161, 241–269. [Google Scholar] [CrossRef]
- Wen, F.; Xin, Z.B.; Fu, L.F.; Li, S.; Su, L.Y.; Maciejewski, S.; Huang, Z.J.; Van Do, T.; Wei, Y.G. Michaelmoelleria (Gesneriaceae), a New Lithophilous Dwelling Genus and Species with Zigzag Corolla Tube from Southern Vietnam. PhytoKeys 2020, 146, 107. [Google Scholar] [CrossRef] [PubMed]
- Middleton, D.J.; Nishii, K.; Puglisi, C.; Forrest, L.L.; Möller, M. Chayamaritia (Gesneriaceae: Didymocarpoideae), a New Genus from Southeast Asia on JSTOR. Plant Syst. Evol. 2015, 301, 1947–1966. [Google Scholar] [CrossRef]
- Möller, M.; Chen, W.-H.; Shui, Y.-M.; Atkins, H.; Middleton, D.J. A New Genus of Gesneriaceae in China and the Transfer of Briggsia Species to Other Genera. Gard. Bull. Singap. 2014, 66, 195–205. [Google Scholar]
- Puglisi, C.; Middleton, D.J. A Revision of Microchirita (Gesneriaceae) in Thailand. Gard. Bull. Singap. 2017, 69, 211–284. [Google Scholar] [CrossRef]
- Puglisi, C.; Yao, T.L.; Milne, R.; Möller, M.; Middleton, D.J. Generic Recircumscription in the Loxocarpinae (Gesneriaceae), as Inferred by Phylogenetic and Morphological Data. Taxon 2016, 65, 277–292. [Google Scholar] [CrossRef] [Green Version]
- Middleton, D.J.; Khew, G.S.; Poopath, M.; Möller, M.; Puglisi, C. Rachunia cymbiformis, a New Genus and Species of Gesneriaceae from Thailand. Nord. J. Bot. 2018, 36, e01992. [Google Scholar] [CrossRef]
- Middleton, D.J.; Triboun, P. A New Species of Somrania (Gesneriaceae) from Thailand. Bull. Singap. 2013, 65, 181–184. [Google Scholar]
- Puglisi, C.; Middleton, D.J. A Revision of Damrongia (Gesneriaceae) in Thailand. Thai For. Bull. 2017, 45, 79–93. [Google Scholar] [CrossRef] [Green Version]
- Möller, M.; Nishii, K.; Atkins, H.J.; Kong, H.H.; Kang, M.; Wei, Y.G.; Wen, F.; Hong, X.; Middleton, D.J. An Expansion of the Genus Deinostigma (Gesneriaceae). Gard. Bull. Singap. 2016, 68, 145–172. [Google Scholar] [CrossRef]
- Wen, F.; Li, S.; Xin, Z.; Fu, L.F.; Hong, X.; Cai, L.; Qin, J.Q.; Pan, B.; Pan, F.Z.; Wei, Y.G. The Latest List of Gesneriaceae in China under the New Chinese Nomenclature. Guangxi Sci. 2019, 26, 37–63. [Google Scholar]
- Nishii, K.; Hughes, M.; Briggs, M.; Haston, E.; Christie, F.; DeVilliers, M.J.; Hanekom, T.; Roos, W.G.; Bellstedt, D.U.; Möller, M. Streptocarpus Redefined to Include All Afro-Malagasy Gesneriaceae: Molecular Phylogenies Prove Congruent with Geographical Distribution and Basic Chromosome Numbers and Uncover Remarkable Morphological Homoplasies. Taxon 2015, 64, 1243–1274. [Google Scholar] [CrossRef]
- Middleton, D.J.; Weber, A.; Yao, T.L.; Sontag, S.; Möller, M. The Current Status of the Species Hitherto Assigned to Henckelia (Gesneriaceae). Edinb. J. Bot. 2013, 70, 385–404. [Google Scholar] [CrossRef]
- Xin, D.; Zhiang, H.; Hongxin, W.; Xiaogang, W.; Tingyun, K. Effects of Dehydration and Rehydration on Photosynthesis of Detached Leaves of the Resurrective Plant Boea hygrometrica. Acta Bot. Sin. 2000, 42, 321–323. [Google Scholar]
- Wang, Y.; Liu, K.; Bi, D.; Zhou, S.; Shao, J. Molecular Phylogeography of East Asian Boea clarkeana (Gesneriaceae) in Relation to Habitat Restriction. PLoS ONE 2018, 13, e0199780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maria Sgherri, C.L.; Loggini, B.; Bochicchio, A.; Navari-Izzo, F. Antioxidant System in Boea hygroscopica: Changes in Response to Desiccation and Rehydration. Phytochemistry 1994, 37, 377–381. [Google Scholar] [CrossRef]
- Gray, B.; Tropical Herbarium, A.; Cook University, J.; Qld, S. Boea resupinata Zich & B.Gray (Gesneriaceae), a New Species from Cape York Peninsula, Queensland, Australia. Austrobaileya 2021, 11, 56–66. [Google Scholar]
- Zhao, H.; Liu, H.; Yu, H.; Hu, Y.; Gao, Y.; Li, Z.; Lin, Z. Cloning and Expression Pattern of a Dehydrin-LikeBDN1 Gene from Drought-Tolerant Boea crassifolia Hemsl. Chin. Sci. Bull. 2000, 45, 2072–2077. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Yang, S.J.; Zhang, S.B.; Zhang, J.L.; Cao, K.F. Cyclic Electron Flow Plays an Important Role in Photoprotection for the Resurrection Plant Paraboea rufescens under Drought Stress. Planta 2012, 235, 819–828. [Google Scholar] [CrossRef]
- Djilianov, D.L.; Moyankova, D.P.; Mladenov, P.V. The Mediterranean: A Cradle of the Resurrection Plants in Europe. Phytol. Balc. 2016, 22, 141–147. [Google Scholar]
- Rakić, T.; Lazarević, M.; Jovanović, Ž.S.; Radović, S.; Siljak-Yakovlev, S.; Stevanović, B.; Stevanović, V. Resurrection Plants of the Genus Ramonda: Prospective Survival Strategies-Unlock Further Capacity of Adaptation, or Embark on the Path of Evolution? Front. Plant Sci. 2014, 4, 550. [Google Scholar] [CrossRef] [Green Version]
- Muller, J.; Sprenger, N.; Bortlik, K.; Boiler, T.; Wiemken Muller, A.; Bortlik, N.; Wiemken, T.; Muller, J.; Sprenger, N.; Bortlik, K.; et al. Desiccation Increases Sucrose Levels in Ramonda and Haberlea, Two Genera of Resurrection Plants in the Gesneriaceae. Physiol. Plant 1997, 100, 153–158. [Google Scholar] [CrossRef]
- Gaff, D.F.; Oliver, M. The Evolution of Desiccation Tolerance in Angiosperm Plants: A rare yet common phenomenon. Funct. Plant Biol. 2013, 40, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Wang, D.; Yu, B.; Yu, X.; Li, W. Maintenance or Collapse: Responses of Extraplastidic Membrane Lipid Composition to Desiccation in the Resurrection Plant Paraisometrum mileense. PLoS ONE 2014, 9, e103430. [Google Scholar] [CrossRef]
- Fernández-Marín, B.; Neuner, G.; Kuprian, E.; Laza, J.M.; García-Plazaola, J.I.; Verhoeven, A. First Evidence of Freezing Tolerance in a Resurrection Plant: Insights into Molecular Mobility and Zeaxanthin Synthesis in the Dark. Physiol. Plant 2018, 163, 472–489. [Google Scholar] [CrossRef] [PubMed]
- Benina, M.; Obata, T.; Mehterov, N.; Ivanov, I.; Petrov, V.; Toneva, V.; Fernie, A.R.; Gechev, T.S. Comparative Metabolic Profiling of Haberlea rhodopensis, Thellungiella halophyla, and Arabidopsis thaliana Exposed to Low Temperature. Front. Plant Sci. 2013, 4, 499. [Google Scholar] [CrossRef] [Green Version]
- Morley, R.J. Interplate Dispersal Paths for Megathermal Angiosperms. Perspect. Plant Ecol. Evol. Syst. 2003, 6, 5–20. [Google Scholar] [CrossRef]
- Perret, M.; Chautems, A.; de Araujo, A.O.; Salamin, N. Temporal and Spatial Origin of Gesneriaceae in the New World Inferred from Plastid DNA Sequences. Bot. J. Linn. Soc. 2013, 171, 61–79. [Google Scholar] [CrossRef] [Green Version]
- Woo, V.L.; Funke, M.M.; Smith, J.F.; Lockhart, P.J.; Garnock-Jones, P.J. New World Origins of Southwest Pacific Gesneriaceae: Multiple Movements Across and Within the South Pacific. Int. J. Plant Sci. 2011, 172, 434–457. [Google Scholar] [CrossRef] [Green Version]
- Petrova, G.; Moyankova, D.; Nishii, K.; Forrest, L.; Tsiripidis, I.; Drouzas, A.D.; Djilianov, D.; Möller, M. The European Paleoendemic Haberlea rhodopensis (Gesneriaceae) Has an Oligocene Origin and a Pleistocene Diversification and Occurs in a Long-Persisting Refugial Area in Southeastern Europe. Int. J. Plant Sci. 2015, 176, 499–514. [Google Scholar] [CrossRef]
- Hultén, E.; Fries, M. Atlas of North European Vascular Plants: North of the Tropic of Cancer; Koeltz Scientific Books: Königstein, Germany, 1986. [Google Scholar]
- Bettin, O.; Cornejo, C.; Edwards, P.J.; Holderegger, R. Phylogeography of the High Alpine Plant Senecio halleri (Asteraceae) in the European Alps: In Situ Glacial Survival with Postglacial Stepwise Dispersal into Peripheral Areas. Mol. Ecol. 2007, 16, 2517–2524. [Google Scholar] [CrossRef] [PubMed]
- Milne, R.; Abbott, R.J. The Origin and Evolution of Tertiary Relict Floras. Adv. Bot. Res. 2002, 38, 281–314. [Google Scholar] [CrossRef]
- Skog, L.E. A Study of the Tribe Gesneriaceae, with a Revision of Gesneria (Gesneriaceae-Gesnerioideae). Smithson. Contrib. Bot. 1976, 29, 1–182. [Google Scholar] [CrossRef]
- Martén-Rodríguez, S.; Fenster, C.B.; Agnarsson, I.; Skog, L.E.; Zimmer, E.A. Evolutionary Breakdown of Pollination Specialization in a Caribbean Plant Radiation. New Phytol. 2010, 188, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Perret, M.; Chautems, A.; Spichiger, R. Dispersal-Vicariance Analyses in the Tribe Sinningiaeae (Gesneriaceae): A Clue to Understanding Biogeographical History of the Brazilian Atlanric Forest. Ann. Mo. Bot. Gard. 2006, 93, 340–358. [Google Scholar] [CrossRef]
- Clements, R.; Sodhi, N.S.; Schilthuizen, M.; Hg, P.K.L. Limestone Karsts of Southeast Asia: Imperiled Arks of Biodiversity. Bioscience 2006, 56, 733–742. [Google Scholar] [CrossRef] [Green Version]
- Porembski, S.; Barthlott, W. Granitic and Gneissic Outcrops (Inselbergs) as Centers of Diversity for Desiccation-Tolerant Vascular Plants. Plant Ecol. 2000, 151, 19–28. [Google Scholar] [CrossRef]
- Alejo-Jacuinde, G.; Herrera-Estrella, L. Exploring the High Variability of Vegetative Desiccation Tolerance in Pteridophytes. Plants 2022, 11, 1222. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Huang, Y.; Wu, F.; Liu, W.; Ning, Y.; Huang, Z.; Tang, S.; Liang, Y. Plant Adaptability in Karst Regions. J. Plant Res. 2021, 134, 889–906. [Google Scholar] [CrossRef] [PubMed]
- Burke, A. Island–Matrix Relationships in Nama Karoo Inselberg Landscapes Part II: Are Some Inselbergs Better Sources than Others? Plant Ecol. 2002, 158, 41–48. [Google Scholar] [CrossRef]
- Smith, S.A.; Beaulieu, J.M. Life History Influences Rates of Climatic Niche Evolution in Flowering Plants. Proc. R. Soc. B Biol. Sci. 2009, 276, 4352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möller, M.; Cronk, Q.C.B. Evolution of Morphological Novelty: A Phylogenetic Analysis of Growth Patterns in Streptocarpus (Gesneriaceae). Evolution 2001, 55, 918–929. [Google Scholar] [CrossRef] [PubMed]
- Rafsanjani, A.; Brulé, V.; Western, T.L.; Pasini, D. Hydro-Responsive Curling of the Resurrection Plant Selaginella lepidophylla. Sci. Rep. 2015, 5, srep08064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; Springer: Heidelberg, Germany, 2003. [Google Scholar]
- Sklenář, P.; Kučerová, A.; Macková, J.; Romoleroux, K. Temperature Microclimates of Plants in a Tropical Alpine Environment: How Much Does Growth Form Matter? Arct. Antarct. Alp. Res. 2018, 48, 61–78. [Google Scholar] [CrossRef]
- Werk, K.S.; Ehleringer, J.; Forseth, I.N.; Cook, C.S. Photosynthetic Characteristics of Sonoran Desert Winter Annuals. Oecologia 1983, 59, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.; Larcher, W. Frost Survival of Plants: Responses and Adaptation to Freezing Stress; Springer: Berlin, Germany, 1987. [Google Scholar]
- Alpert, P. Constraints of Tolerance: Why Are Desiccation-Tolerant Organisms so Small or Rare? J. Exp. Biol. 2006, 209, 1575–1584. [Google Scholar] [CrossRef] [Green Version]
- Asami, P.; Mundree, S.; Williams, B. Saving for a Rainy Day: Control of Energy Needs in Resurrection Plants. Plant Sci. 2018, 271, 62–66. [Google Scholar] [CrossRef]
- Squeo, F.A.; Rada, F.; Azocar, A.; Goldstein, G. Freezing Tolerance and Avoidance in High Tropical Andean Plants: Is It Equally Represented in Species with Different Plant Height? Oecologia 1991, 86, 378–382. [Google Scholar] [CrossRef]
- Cruz-Maldonado, N.; Weemstra, M.; Jiménez, L.; Roumet, C.; Angeles, G.; Barois, I.; de los Santos, M.; Morales-Martinez, M.A.; Palestina, R.A.; Rey, H.; et al. Aboveground-Trait Variations in 11 (Sub)Alpine Plants along a 1000-m Elevation Gradient in Tropical Mexico. Alp. Bot. 2021, 131, 187–200. [Google Scholar] [CrossRef]
- Leon-Garcia, I.V.; Lasso, E. High Heat Tolerance in Plants from the Andean Highlands: Implications for Paramos in a Warmer World. PLoS ONE 2019, 14, e0224218. [Google Scholar] [CrossRef] [Green Version]
- Scheepens, J.F.; Frei, E.S.; Stöcklin, J. Genotypic and Environmental Variation in Specific Leaf Area in a Widespread Alpine Plant after Transplantation to Different Altitudes. Oecologia 2010, 164, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Beck, E.; Schulze, E.D.; Senser, M.; Scheibe, R. Equilibrium Freezing of Leaf Water and Extracellular Ice Formation in Afroalpine ‘Giant Rosette’ Plants. Planta 1984, 162, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Melcher, P.J.; Goldstein, G.; Meinzer, F.C.; Minyard, B.; Giambelluca, T.W.; Loope, L.L. Determinants of Thermal Balance in the Hawaiian Giant Rosette Plant, Argyroxiphium sandwicense. Oecologia 1994, 98, 412–418. [Google Scholar] [CrossRef]
- McKown, A.D.; Cochard, H.; Sack, L. Decoding Leaf Hydraulics with a Spatially Explicit Model: Principles of Venation Architecture and Implications for Its Evolution. Am. Nat. 2010, 175, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Niklas, K.J. A Mechanical Perspective on Foliage Leaf Form and Function. New Phytol. 1999, 143, 19–31. [Google Scholar] [CrossRef]
- Kehr, J.; Buhtz, A. Long Distance Transport and Movement of RNA through the Phloem. J. Exp. Bot. 2008, 59, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sack, L.; Frole, K. Leaf Structural Diversity Is Related to Hydraulic Capacity in Tropical Rain Forest Trees. Ecology 2006, 87, 483–491. [Google Scholar] [CrossRef]
- Givnish, T. Comparative Studies of Leaf from: Assessing the Relative Roles of Selective Preassures and Phylogenetic Constraints. New Phytol. 1987, 106, 131–160. [Google Scholar] [CrossRef]
- Fonseca, C.R.; Overton, J.M.C.; Collins, B.; Westoby, M. Shifts in Trait-Combinations along Rainfall and Phosphorus Gradients. J. Ecol. 2000, 88, 964–977. [Google Scholar] [CrossRef]
- Davis, S.D.; Sperry, J.S.; Hacke, U.G. The Relationship between Xylem Conduit Diameter and Cavitation Caused by Freezing. Am. J. Bot. 1999, 86, 1367–1372. [Google Scholar] [CrossRef] [Green Version]
- Baas, P.; Ewers, F.W.; Davis, S.D.; Wheeler, E.A. Evolution of Xylem Physiology. In The Evolution of Plant Physiology; Hemsley, A.R., Poole, I., Eds.; Academic Press: Oxford, UK, 2004; pp. 273–295. ISBN 9780123395528. [Google Scholar]
- Scoffoni, C.; Rawls, M.; Mckown, A.; Cochard, H.; Sack, L. Decline of Leaf Hydraulic Conductance with Dehydration: Relationship to Leaf Size and Venation Architecture. Plant Physiol. 2011, 156, 843. [Google Scholar] [CrossRef] [Green Version]
- Sack, L.; Dietrich, E.M.; Streeter, C.M.; Sánchez-Gómez, D.; Holbrook, N.M. Leaf Palmate Venation and Vascular Redundancy Confer Tolerance of Hydraulic Disruption. Proc. Natl. Acad. Sci. USA 2008, 105, 1567–1572. [Google Scholar] [CrossRef] [Green Version]
- Ichie, T.; Inoue, Y.; Takahashi, N.; Kamiya, K.; Kenzo, T. Ecological Distribution of Leaf Stomata and Trichomes among Tree Species in a Malaysian Lowland Tropical Rain Forest. J. Plant Res. 2016, 129, 625–635. [Google Scholar] [CrossRef]
- Dahlin, R.M.; Brick, M.A.; Barry, J.; Dahlin, O.R.M.; Brick, M.A.; Ogg, J.B. Characterization and Density of Trichomes on Three Common Bean Cultivars. Econ. Bot. 1992, 46, 299–304. [Google Scholar] [CrossRef]
- Benz, B.W.; Martin, C.E. Foliar Trichomes, Boundary Layers, and Gas Exchange in 12 Species of Epiphytic Tillandsia (Bromeliaceae). J. Plant Physiol. 2006, 163, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Tattini, M. Multiple Functional Roles of Flavonoids in Photoprotection on JSTOR. New Phytol. 2010, 186, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Alcalá, G.; Gotor, C.; Meyer, A.J.; Fricker, M.; Vega, J.M.; Romero, L.C. Glutathione Biosynthesis in Arabidopsis Trichome Cells. Proc. Natl. Acad. Sci. USA 2000, 97, 11108–11113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauser, M.T. Molecular Basis of Natural Variation and Environmental Control of Trichome Patterning. Front. Plant Sci. 2014, 5, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prozherina, N.; Freiwald, V.; Rousi, M.; Oksanen, E. Interactive Effect of Springtime Frost and Elevated Ozone on Early Growth, Foliar Injuries and Leaf Structure of Birch (Betula pendula). New Phytol. 2003, 159, 623–636. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Peiffer, M.; de Moraes, C.M.; Felton, G.W. Roles of Ethylene and Jasmonic Acid in Systemic Induced Defense in Tomato (Solanum lycopersicum) against Helicoverpa zea. Planta 2014, 239, 577–589. [Google Scholar] [CrossRef]
- Ning, P.; Wang, J.; Zhou, Y.; Gao, L.; Wang, J.; Gong, C. Adaptional Evolution of Trichome in Caragana korshinskii to Natural Drought Stress on the Loess Plateau, China. Ecol. Evol. 2016, 6, 3786–3795. [Google Scholar] [CrossRef] [Green Version]
- Yan, A.; Pan, J.; An, L.; Gan, Y.; Feng, H. The Responses of Trichome Mutants to Enhanced Ultraviolet-B Radiation in Arabidopsis thaliana. J. Photochem. Photobiol. B 2012, 113, 29–35. [Google Scholar] [CrossRef]
- Schuepp, P.H. Leaf Boundary Layers. New Phytol. 1993, 125, 477–507. [Google Scholar] [CrossRef]
- Lacey, M. From Fossils to Physiology: Testing the Functional Significance of Leaf Shape; University of Washington: Washington, DC, USA, 2019. [Google Scholar]
- Willson, C.J.; Manos, P.S.; Jackson, R.B. Hydraulic Traits Are Influenced by Phylogenetic History in the Drought-Resistant, Invasive Genus Juniperus (Cupressaceae). Am. J. Bot. 2008, 95, 299–314. [Google Scholar] [CrossRef]
- Moore, J.P.; Lindsey, G.G.; Farrant, J.M.; Brandt, W.F. An Overview of the Biology of the Desiccation-Tolerant Resurrection Plant Myrothamnus flabellifolia. Ann. Bot. 2007, 99, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Kampowski, T.; Demandt, S.; Poppinga, S.; Speck, T. Kinematical, Structural and Mechanical Adaptations to Desiccation in Poikilohydric Ramonda myconi (Gesneriaceae). Front. Plant Sci. 2018, 9, 1701. [Google Scholar] [CrossRef] [Green Version]
- Vieira, E.A.; Silva, K.R.; Oriani, A.; Moro, C.F.; Braga, M.R. Mechanisms of Desiccation Tolerance in the Bromeliad Pitcairnia burchellii Mez: Biochemical Adjustments and Structural Changes. Plant Physiol. Biochem. 2017, 121, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heilmeier, H.; Hartung, W. Chamaegigas Intrepidus DINTER: An Aquatic Poikilohydric Angiosperm That Is Perfectly Adapted to Its Complex and Extreme Environmental Conditions. In Plant Desiccation Tolerance. Ecological Studies; Lüttge, U., Beck, E., Bartel, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 215, pp. 233–254. [Google Scholar]
- Gaff, D.F. Desiccation Tolerant ‘Resurrection’ Grasses from Kenya and West Africa. Oecologia 1986, 70, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Tang, M.J.; Hu, Y.L.; Lin, Z.P. Isolation and Characterization of a Dehydrin-like Gene from Drought-Tolerant Boea crassifolia. Plant Sci. 2004, 166, 1167–1175. [Google Scholar] [CrossRef]
- Fernández-Marín, B.; Nadal, M.; Gago, J.; Fernie, A.R.; López-Pozo, M.; Artetxe, U.; García-Plazaola, J.I.; Verhoeven, A. Born to Revive: Molecular and Physiological Mechanisms of Double Tolerance in a Paleotropical and Resurrection Plant. New Phytol. 2020, 226, 741–759. [Google Scholar] [CrossRef] [PubMed]
- Jovanović, Ž.; Rakić, T.; Stevanović, B.; Radović, S. Characterization of Oxidative and Antioxidative Events during Dehydration and Rehydration of Resurrection Plant Ramonda nathaliae. Plant Growth Regul. 2011, 64, 231–240. [Google Scholar] [CrossRef]
- Liu, J.; Moyankova, D.; Djilianov, D.; Deng, X. Common and Specific Mechanisms of Desiccation Tolerance in Two Gesneriaceae Resurrection Plants. Multiomics Evidences. Front. Plant Sci. 2019, 10, 1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drazic, G.; Mihailovic, N.; Stevanovic, B. Chlorophyll Metabolism in Leaves of Higher Poikilohydric Plants Ramonda serbica Panč, and Ramonda nathaliae Panč, et Petrov. during Dehydration and Rehydration. J. Plant Physiol. 1999, 154, 379–384. [Google Scholar] [CrossRef]
- Liu, J.; Moyankova, D.; Lin, C.T.; Mladenov, P.; Sun, R.Z.; Djilianov, D.; Deng, X. Transcriptome Reprogramming during Severe Dehydration Contributes to Physiological and Metabolic Changes in the Resurrection Plant Haberlea rhodopensis. BMC Plant Biol. 2018, 18, 351. [Google Scholar] [CrossRef]
- Georgieva, K.; Mihailova, G.; Velitchkova, M.; Popova, A. Recovery of Photosynthetic Activity of Resurrection Plant Haberlea rhodopensis from Drought-and Freezing-Induced Desiccation. Photosynthetica 2020, 58, 911–921. [Google Scholar] [CrossRef]
- Mladenov, P.; Finazzi, G.; Bligny, R.; Moyankova, D.; Zasheva, D.; Boisson, A.M.; Brugière, S.; Krasteva, V.; Alipieva, K.; Simova, S.; et al. In Vivo Spectroscopy and NMR Metabolite Fingerprinting Approaches to Connect the Dynamics of Photosynthetic and Metabolic Phenotypes in Resurrection Plant Haberlea rhodopensis during Desiccation and Recovery. Front. Plant Sci. 2015, 6, 564. [Google Scholar] [CrossRef] [Green Version]
- Asada, K. Production and Scavenging of Reactive Oxygen Species in Chloroplasts and Their Functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Oliver, M.J.; Farrant, J.M.; Hilhorst, H.W.M.; Mundree, S.; Williams, B.; Bewley, J.D. Desiccation Tolerance: Avoiding Cellular Damage during Drying and Rehydration. Annu. Rev. Plant Biol. 2020, 71, 435–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgieva, K.; Rapparini, F.; Bertazza, G.; Mihailova, G.; Sárvári, É.; Solti, Á.; Keresztes, Á. Alterations in the Sugar Metabolism and in the Vacuolar System of Mesophyll Cells Contribute to the Desiccation Tolerance of Haberlea rhodopensis Ecotypes. Protoplasma 2017, 254, 193–201. [Google Scholar] [CrossRef]
- Cañigueral, S.; Salvía, M.J.; Vila, R.; Iglesias, J.; Virgili, A.; Parella, T. New Polyphenol Glycosides from Ramonda myconi. J. Nat. Prod. 1996, 59, 419–422. [Google Scholar] [CrossRef]
- Jensen, S.R. Caffeoyl Phenylethanoid Glycosides in Sanango racemosum and in the Gesneriaceae. Phytochemistry 1996, 43, 777–783. [Google Scholar] [CrossRef]
- Feng, W.S.; Li, Y.J.; Zheng, X.K.; Wang, Y.Z.; Su, F.Y.; Pei, Y.Y. Two New C-Glycosylflavones from Boea hygrometrica. J. Asian Nat. Prod. Res. 2011, 13, 618–623. [Google Scholar] [CrossRef]
- Georgieva, K.; Dagnon, S.; Gesheva, E.; Bojilov, D.; Mihailova, G.; Doncheva, S. Antioxidant Defense during Desiccation of the Resurrection Plant Haberlea rhodopensis. Plant Physiol. Biochem. 2017, 114, 51–59. [Google Scholar] [CrossRef]
- Gechev, T.S.; Benina, M.; Obata, T.; Tohge, T.; Sujeeth, N.; Minkov, I.; Hille, J.; Temanni, M.R.; Marriott, A.S.; Bergström, E.; et al. Molecular Mechanisms of Desiccation Tolerance in the Resurrection Glacial Relic Haberlea rhodopensis. Cell. Mol. Life Sci. 2013, 70, 689–709. [Google Scholar] [CrossRef]
- Mladenov, P.V.; Zasheva, D.; Planchon, S.; Leclercq, C.; Falconet, D.; Moyet, L.; Brugière, S.; Moyankova, D.; Tchorbadjieva, M.; Ferro, M.; et al. Proteomics Evidence of a Systemic Response to Desiccation in the Resurrection Plant Haberlea rhodopensis. SSRN Electron. J. 2022, 23, 8520. [Google Scholar] [CrossRef]
- Quartacci, M.F.; Glišić, O.; Stevanović, B.; Navari-Izzo, F. Plasma Membrane Lipids in the Resurrection Plant Ramonda serbica Following Dehydration and Rehydration. J. Exp. Bot. 2002, 53, 2159–2166. [Google Scholar] [CrossRef] [Green Version]
- Moyankova, D.; Mladenov, P.; Berkov, S.; Peshev, D.; Georgieva, D.; Djilianov, D. Metabolic Profiling of the Resurrection Plant Haberlea rhodopensis during Desiccation and Recovery. Physiol. Plant 2014, 152, 675–687. [Google Scholar] [CrossRef]
- Navari-Izzo, F.; Ricci, F.; Vazzana, C.; Quartacci, M.F. Unusual Composition of Thylakoid Membranes of the Resurrection Plant Boea hygroscopica: Changes in Lipids upon Dehydration and Rehydration. Physiol. Plant 1995, 94, 135–142. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, B.; Phillips, J.; Zhang, Z.N.; Du, H.; Xu, T.; Huang, L.C.; Zhang, X.F.; Xu, G.H.; Li, W.L.; et al. Global Transcriptome Analysis Reveals Acclimation-Primed Processes Involved in the Acquisition of Desiccation Tolerance in Boea hygrometrica. Plant Cell Physiol. 2015, 56, 1429–1441. [Google Scholar] [CrossRef] [Green Version]
- Pinnola, A.; Bassi, R. Molecular Mechanisms Involved in Plant Photoprotection. Biochem. Soc. Trans. 2018, 46, 467–482. [Google Scholar] [CrossRef] [PubMed]
- Mihailova, G.; Vasileva, I.; Gigova, L.; Gesheva, E.; Simova-Stoilova, L.; Georgieva, K. Antioxidant Defense during Recovery of Resurrection Haberlea rhodopensis from Drought- and Freezing-Induced Desiccation. Plants 2022, 11, 175. [Google Scholar] [CrossRef] [PubMed]
- Mitra, J.; Xu, G.; Wang, B.; Li, M.; Deng, X. Understanding Desiccation Tolerance Using the Resurrection Plant Boea hygrometrica as a Model System. Front. Plant Sci. 2013, 4, 446. [Google Scholar] [CrossRef] [Green Version]
- Mihailova, G.; Kocheva, K.; Goltsev, V.; Kalaji, H.M.; Georgieva, K. Application of a Diffusion Model to Measure Ion Leakage of Resurrection Plant Leaves Undergoing Desiccation. Plant Physiol. Biochem. 2018, 125, 185–192. [Google Scholar] [CrossRef]
- Farrant, J.M.; Cooper, K.; Nell, H. Desiccation Tolerance. In Plant Stress Physiology; CABI: Boston, MA, USA, 2000; pp. 248–265. ISBN 9781845939953. [Google Scholar]
- Georgieva, K.; Sárvári, É.; Keresztes, Á. Protection of Thylakoids against Combined Light and Drought by a Lumenal Substance in the Resurrection Plant Haberlea rhodopensis. Ann. Bot. 2010, 105, 117–126. [Google Scholar] [CrossRef]
- Djilianov, D.; Ivanov, S.; Moyankova, D.; Miteva, L.; Kirova, E.; Alexieva, V.; Joudi, M.; Peshev, D.; van den Ende, W. Sugar Ratios, Glutathione Redox Status and Phenols in the Resurrection Species Haberlea rhodopensis and the Closely Related Non-Resurrection Species Chirita eberhardtii. Plant Biol. 2011, 13, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-Mediated Abiotic Stress-Induced Programmed Cell Death in Plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [Green Version]
- Kappen, V.L. Sucht an Blättern Einiger Farne Und von Ramonda myconi. Flora 1966, 156, 427. [Google Scholar]
- Georgieva, K.; Mihailova, G.; Gigova, L.; Dagnon, S.; Simova-Stoilova, L.; Velitchkova, M. The Role of Antioxidant Defense in Freezing Tolerance of Resurrection Plant Haberlea rhodopensis. Physiol. Mol. Biol. Plants 2021, 27, 1119–1133. [Google Scholar] [CrossRef] [PubMed]
- Mihailova, G.; Solti, Á.; Sárvári, É.; Keresztes, Á.; Rapparini, F.; Velitchkova, M.; Simova-Stoilova, L.; Aleksandrov, V.; Georgieva, K. Freezing Tolerance of Photosynthetic Apparatus in the Homoiochlorophyllous Resurrection Plant Haberlea rhodopensis. Environ. Exp. Bot. 2020, 178, 104157. [Google Scholar] [CrossRef]
- Mihailova, G.; Christov, N.K.; Sárvári, É.; Solti, Á.; Hembrom, R.; Solymosi, K.; Keresztes, Á.; Velitchkova, M.; Popova, A.V.; Simova-Stoilova, L.; et al. Reactivation of the Photosynthetic Apparatus of Resurrection Plant Haberlea rhodopensis during the Early Phase of Recovery from Drought- and Freezing-Induced Desiccation. Plants 2022, 11, 2185. [Google Scholar] [CrossRef]
- Georgieva, K.; Popova, A.V.; Mihailova, G.; Ivanov, A.G.; Velitchkova, M. Limiting Steps and the Contribution of Alternative Electron Flow Pathways in the Recovery of the Photosynthetic Functions after Freezing-Induced Desiccation of Haberlea rhodopensis. Photosynthetica 2022, 60, 136–146. [Google Scholar] [CrossRef]
- Kuroki, S.; Tsenkova, R.; Moyankova, D.; Muncan, J.; Morita, H.; Atanassova, S.; Djilianov, D. Water Molecular Structure Underpins Extreme Desiccation Tolerance of the Resurrection Plant Haberlea rhodopensis. Sci. Rep. 2019, 9, 3049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, A.; O′Leary, B.; Signorelli, S.; Falconet, D.; Moyankova, D.; Whelan, J.; Djilianov, D.; Murcha, M.W. Mitochondrial Activity and Biogenesis during Resurrection of Haberlea rhodopensis. New Phytol. 2022, 236, 943–957. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subtribe | Documented DT Species | Tentative Double-Tolerant/DT Species |
|---|---|---|
| Streptocarpinae | Streptocarpus revivescens | Streptocarpus rexii Streptocarpus meyeri Streptocarpus baudertii Streptocarpus montigena Streptocarpus rhodesianus |
| Corallodiscinae | - | Corallodiscus kingianus Corallodiscus cooperi Corallodiscus bhutanicus |
| Didymocarpinae | Oreocharis billburttii Oreocharis primuloides Oreocharis mileensis | Oreocharis pankaiyuae Oreocharis mairei Oreocharis ovatilobata Oreocharis flavovirens Oreocharis muscicola Oreocharis blepharophylla Oreocharis delavayi Oreocharis ninglangensis Oreocharis crispata Oreocharis magnidens Oreocharis stewardii |
| Didymocarpinae | - | Henckelia incana Henckelia gambleana Henckelia fischeri Henckelia bracteata Henckelia wayanadensis Henckelia innominata |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Legardón, A.; García-Plazaola, J.I. Gesneriads, a Source of Resurrection and Double-Tolerant Species: Proposal of New Desiccation- and Freezing-Tolerant Plants and Their Physiological Adaptations. Biology 2023, 12, 107. https://doi.org/10.3390/biology12010107
Legardón A, García-Plazaola JI. Gesneriads, a Source of Resurrection and Double-Tolerant Species: Proposal of New Desiccation- and Freezing-Tolerant Plants and Their Physiological Adaptations. Biology. 2023; 12(1):107. https://doi.org/10.3390/biology12010107
Chicago/Turabian StyleLegardón, Ane, and José Ignacio García-Plazaola. 2023. "Gesneriads, a Source of Resurrection and Double-Tolerant Species: Proposal of New Desiccation- and Freezing-Tolerant Plants and Their Physiological Adaptations" Biology 12, no. 1: 107. https://doi.org/10.3390/biology12010107