
The Sheep as a Large Animal Model for the Investigation and Treatment of Human Disorders


Abstract
:Simple Summary
Abstract

1. Introduction
2. Criteria for Choosing a Suitable Experimental Animal Model for Translational Research
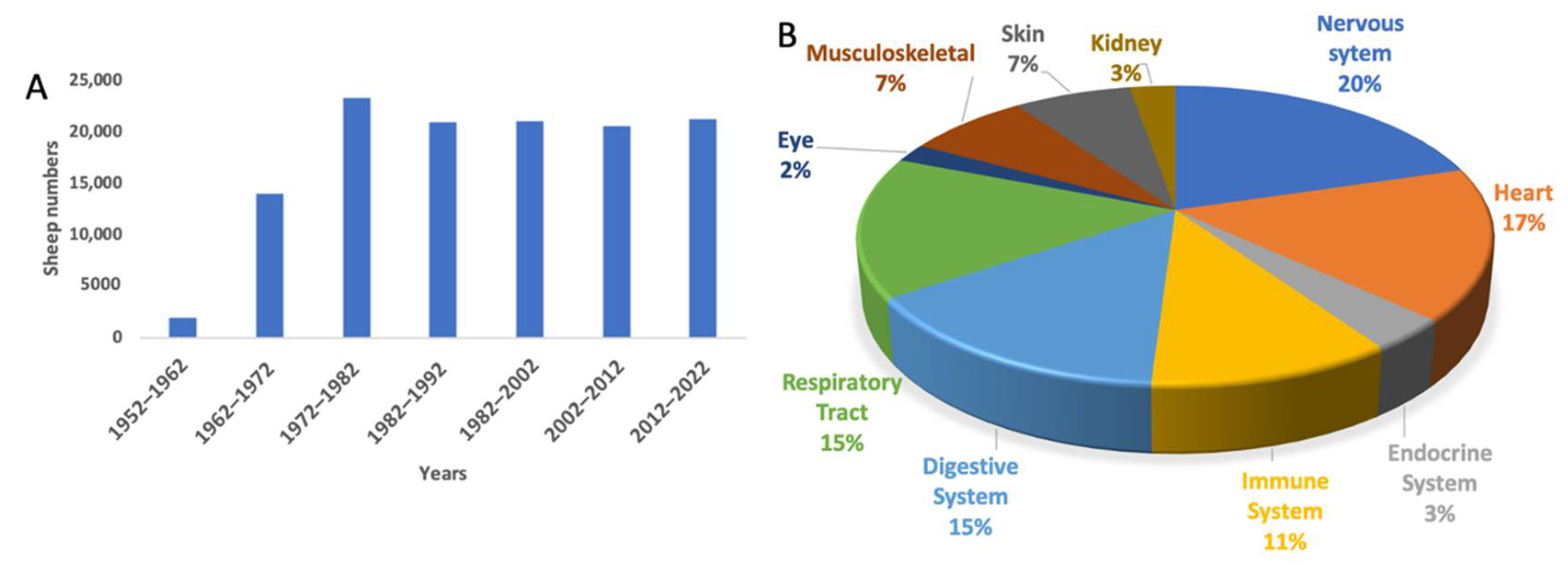
3. The Use of Sheep to Model Human Diseases in Biomedical Research
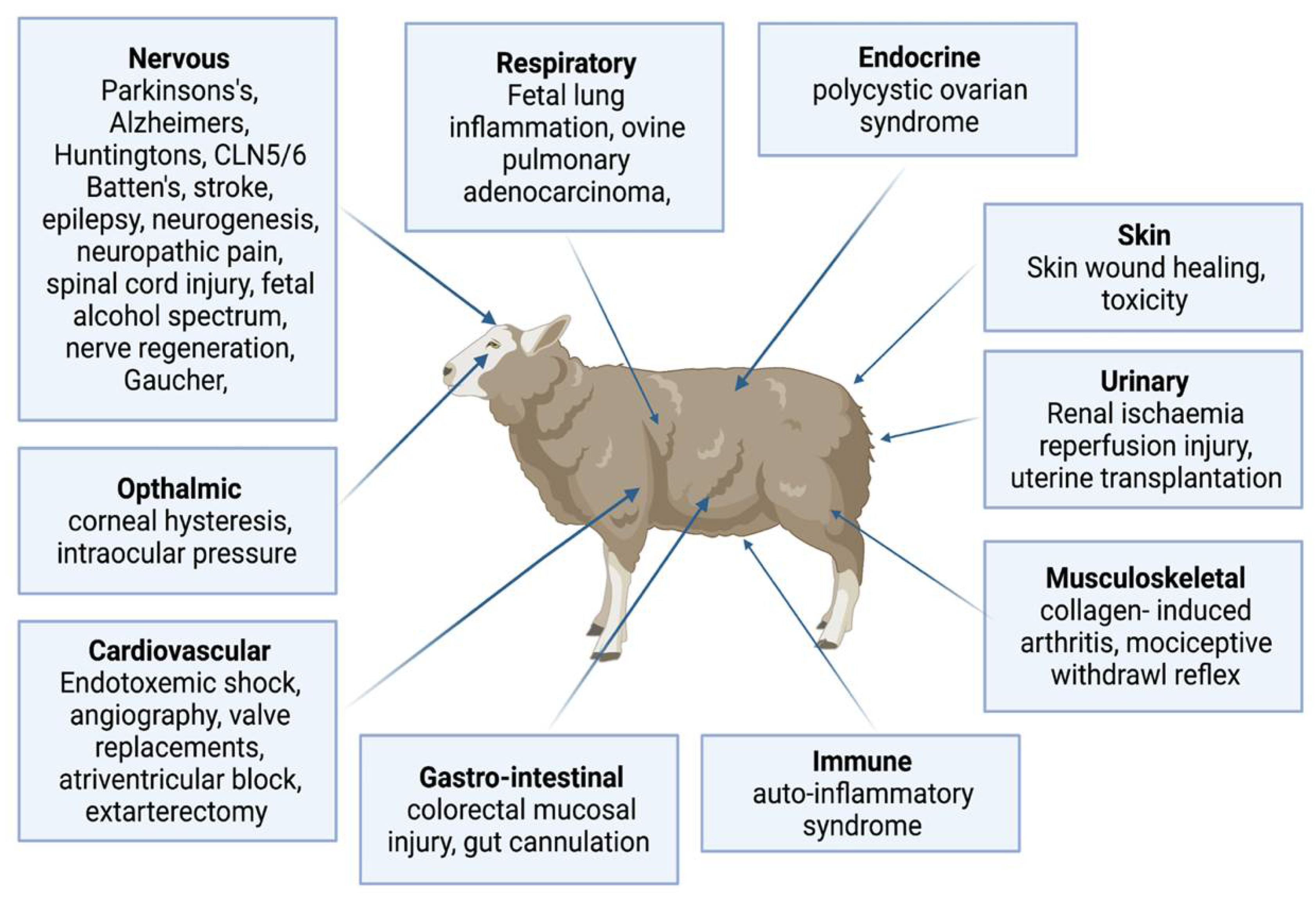
3.1. Central Nervous System Research
3.2. Cardiovascular Research
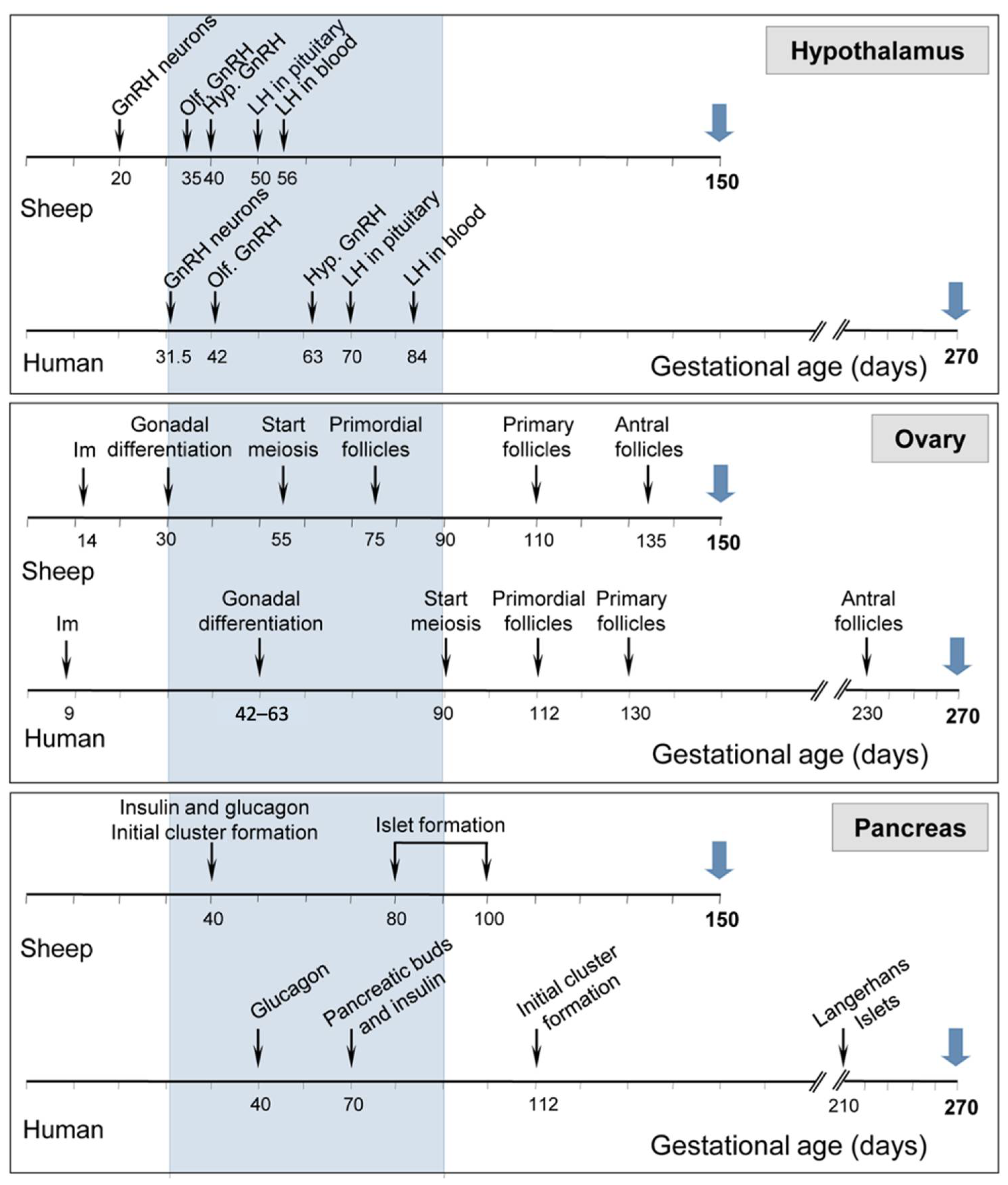
3.3. Endocrine and Reproductive Research
3.4. Immune System Research
3.5. Gastrointestinal Research
3.6. Respiratory System Research
3.7. Ophthalmic Research
3.8. Musculoskeletal Research
3.9. Skin Research
3.10. Renal Disease Research
4. Technical Advances
4.1. Vaccine Development and Testing
4.2. Therapeutic Interventions
5. Limitations and Future Directions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Eaton, S.L.; Wishart, T.M. Bridging the gap: Large animal models in neurodegenerative research. Mamm. Genome 2017, 28, 324–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leenaars, C.H.C.; Kouwenaar, C.; Stafleu, F.R.; Bleich, A.; Ritskes-Hoitinga, M.; De Vries, R.B.M.; Meijboom, F.L.B. Animal to human translation: A systematic scoping review of reported concordance rates. J. Transl. Med. 2019, 17, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyhan, A.A. Lost in translation: The valley of death across preclinical and clinical divide–identification of problems and overcoming obstacles. Transl. Med. Commun. 2019, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Volk, H.-D.; Stevens, M.M.; Mooney, D.J.; Grainger, D.W.; Duda, G.N. Key elements for nourishing the translational research environment. Sci. Transl. Med. 2015, 7, 282cm2. [Google Scholar] [CrossRef] [Green Version]
- Andrade, E.L.; Bento, A.F.; Cavalli, J.; Oliveira, S.K.; Schwanke, R.C.; Siqueira, J.M.; Freitas, C.S.; Marcon, R.; Calixto, J.B. Non-clinical studies in the process of new drug development—Part II: Good laboratory practice, metabolism, pharmacokinetics, safety and dose translation to clinical studies. Braz. J. Med. Biol. Res. 2016, 49, e5646. [Google Scholar] [CrossRef] [Green Version]
- Davidson, M.K.; Lindsey, J.R.; Davis, J.K. Requirement and selection of an animal model. Isr. J. Med. Sci. 1987, 23, 551–555. [Google Scholar]
- Pinnapureddy, A.R.; Stayner, C.; McEwan, J.; Baddeley, O.; Forman, J.; Eccles, M.R. Large animal models of rare genetic disorders: Sheep as phenotypically relevant models of human genetic disease. Orphanet J. Rare Dis. 2015, 10, 107. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, A.; Gonzalez, L.; Blikslager, A. Large animal models: The key to translational discovery in digestive disease research. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Marino, L.; Merskin, D. Intelligence, complexity, and individuality in sheep. Anim. Sentience 2019, 206, 1–26. [Google Scholar] [CrossRef]
- McMillen, C. The sheep—An ideal model for biomedical research? ANZCCART News 2001, 14, 12. [Google Scholar]
- De Las Heras Guillamón, M.; Borderías Clau, L. The sheep as a large animal experimental model in respiratory diseases research. Arch. Bronconeumol. 2010, 46, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.J.; Mitchell, N.L. The translational benefits of sheep as large animal models of human neurological disorders. Front. Vet. Sci. 2022, 9, 831838. [Google Scholar] [CrossRef] [PubMed]
- Van Norman, G.A. Limitations of animal studies for predicting toxicity in clinical trials: Is it time to rethink our current approach? JACC Basic Transl. Sci. 2019, 4, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Sartoretto, S.C.; Uzeda, M.J.; Miguel, F.B.; Nascimento, J.R.; Ascoli, F.; Calasans-Maia, M.D. Sheep as an experimental model for biomaterial implant evaluation. Acta. Ortop. Bras. 2016, 24, 262–266. [Google Scholar] [CrossRef] [Green Version]
- Hurtig, M.B.; Buschmann, M.D.; Fortier, L.A.; Hoemann, C.D.; Hunziker, E.B.; Jurvelin, J.S.; Mainil-Varlet, P.; McIlwraith, C.W.; Sah, R.L.; Whiteside, R.A. Preclinical studies for cartilage repair: Recommendations from the international cartilage repair society. Cartilage 2011, 2, 137–152. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Yeung, A.C.; Ikeno, F. The representative porcine model for human cardiovascular disease. J. Biomed. Biotechnol. 2011, 2011, 195483. [Google Scholar] [CrossRef]
- Kol, A.; Arzi, B.; Athanasiou, K.A.; Farmer, D.L.; Nolta, J.A.; Rebhun, R.B.; Chen, X.; Griffiths, L.G.; Verstraete, F.J.; Murphy, C.J.; et al. Companion animals: Translational scientist’s new best friends. Sci. Transl. Med. 2015, 7, 308ps321. [Google Scholar] [CrossRef] [Green Version]
- Ribitsch, I.; Baptista, P.M.; Lange-Consiglio, A.; Melotti, L.; Patruno, M.; Jenner, F.; Schnabl-Feichter, E.; Dutton, L.C.; Connolly, D.J.; van Steenbeek, F.G.; et al. Large animal models in regenerative medicine and tissue engineering: To do or not to do. Front. Bioeng. Biotechnol. 2020, 8, 972. [Google Scholar] [CrossRef]
- Turner, A.S. Animal model of osteoporosis- necessity and limitations. Eur. Cells Mater. 2001, 1, 66–81. [Google Scholar] [CrossRef]
- O’Kane, D.; Gibson, L.; May, C.N.; du Plessis, J.; Shulkes, A.; Baldwin, G.S.; Bolton, D.; Ischia, J.; Patel, O. Zinc preconditioning protects against renal ischaemia reperfusion injury in a preclinical sheep large animal model. BioMetals 2018, 31, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Partridge, B.; Rossmeisl, J.H., Jr. Companion animal models of neurological disease. J. Neurosci. Methods 2020, 331, 108484. [Google Scholar] [CrossRef]
- Phillips, K.A.; Bales, K.L.; Capitanio, J.P.; Conley, A.; Czoty, P.W.; t Hart, B.A.; Hopkins, W.D.; Hu, S.L.; Miller, L.A.; Nader, M.A.; et al. Why primate models matter. Am. J. Primatol. 2014, 76, 801–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swearengen, J.R. Choosing the right animal model for infectious disease research. Anim. Model. Exp. Med. 2018, 1, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K.; Goor, A.V.; Walker, K.E.; Hailstock, T.; Franklin, J.; Dai, C. Importance of the pig as a human biomedical model. Sci. Transl. Med. 2021, 24, 621. [Google Scholar] [CrossRef]
- Gonzalez, L.M.; Moeser, A.J.; Blikslager, A.T. Porcine models of digestive disease: The future of large animal translational research. Transl. Res. 2015, 166, 12–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, H.G.; Rashdan, N.A.; Whitelaw, C.B.; Corcoran, B.M.; Summers, K.M.; MacRae, V.E. Large animal models of cardiovascular disease. Cell Biochem. Funct. 2016, 34, 113–132. [Google Scholar] [CrossRef] [Green Version]
- Schuttler, D.; Tomsits, P.; Bleyer, C.; Vlcek, J.; Pauly, V.; Hesse, N.; Sinner, M.; Merkus, D.; Hamers, J.; Kaab, S.; et al. A practical guide to setting up pig models for cardiovascular catheterization, electrophysiological assessment and heart disease research. Lab Anim. 2022, 51, 46–67. [Google Scholar] [CrossRef]
- Geens, J.H.; Trenso, N.S.; Rega, F.R.; Verbekin, E.K.; Meyns, B.P. Ovine models for chronic failure. Int. J. Artif. Organs 2009, 32, 496–506. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.S. Experiences with sheep as an animal model for shoulder surgery: Strengths and shortcomings. J. Shoulder Elbow Surg. 2007, 16, S158–S163. [Google Scholar] [CrossRef]
- Bassols, A.; Costa, C.; Eckersall, P.D.; Osada, J.; Sabria, J.; Tibau, J. The pig as an animal model for human pathologies: A proteomics perspective. Proteom. Clin. Appl. 2014, 8, 715–731. [Google Scholar] [CrossRef]
- Hornak, M.; Oracova, E.; Hulinska, P.; Urbankova, L.; Rubes, J. Aneuploidy detection in pigs using comparative genomic hybridization: From the oocytes to blastocysts. PLoS ONE 2012, 7, e30335. [Google Scholar] [CrossRef] [Green Version]
- Szulanczyk-Mencel, K.; Rzasa, A.; Bielas, W. Relationships between ovarian cysts and morphological and hormonal state of ovarian cortex in sows. Anim. Reprod. Sci. 2010, 121, 273–278. [Google Scholar] [CrossRef]
- Pabst, R. The pig as a model for immunology research. Cell Tissue Res. 2020, 380, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Evers, M.M.; Miniarikova, J.; Juhas, S.; Valles, A.; Bohuslavova, B.; Juhasova, J.; Skalnikova, H.K.; Vodicka, P.; Valekova, I.; Brouwers, C.; et al. AAV5-miHTT gene therapy demonstrates broad distribution and strong human mutant huntingtin lowering in a Huntington’s disease minipig model. Mol. Ther. 2018, 26, 2163–2177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallès, A.; Evers, M.M.; Stam, A.; Sogorb-Gonzalez, M.; Brouwers, C.; Vendrell-Tornero, C.; Acar-Broekmans, S.; Paerels, L.; Klima, J.; Bohuslavova, B.; et al. Widespread and sustained target engagement in Huntington’s disease minipigs upon intrastriatal microRNA-based gene therapy. Sci. Transl. Med. 2021, 13, eabb8920. [Google Scholar] [CrossRef] [PubMed]
- Sauleau, P.; Lapouble, E.; Val-Laillet, D.; Malbert, C.H. The pig model in brain imaging and neurosurgery. Animal 2009, 3, 1138–1151. [Google Scholar] [CrossRef] [Green Version]
- Ryan, M.C.; Sherman, P.; Rowland, L.M.; Wijtenburg, S.A.; Acheson, A.; Fieremans, E.; Veraart, J.; Novikov, D.S.; Hong, L.E.; Sladky, J.; et al. Miniature pig model of human adolescent brain white matter development. J. Neurosci. Methods 2018, 296, 99–108. [Google Scholar] [CrossRef]
- Bertoni, L.; Jacquet-Guibon, S.; Branly, T.; Legendre, F.; Desance, M.; Mespoulhes, C.; Melin, M.; Hartmann, D.J.; Schmutz, A.; Denoix, J.M.; et al. An experimentally induced osteoarthritis model in horses performed on both metacarpophalangeal and metatarsophalangeal joints: Technical, clinical, imaging, biochemical, macroscopic and microscopic characterization. PLoS ONE 2020, 15, e0235251. [Google Scholar] [CrossRef]
- Mcllwraith, C.W.; Frisbie, D.D.; Kawcak, C.E. The horse as a model of naturally occurring osteoarthritis. Bone Jt. J. 2012, 1, 297–309. [Google Scholar]
- Evans, C.H.; Ghivizzani, S.C.; Robbins, P.D. Arthritis gene therapy and its tortuous path into the clinic. Transl. Res. 2013, 161, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Frisbie, D.D.; Ghivizzani, S.C.; Robbins, P.D.; Evans, C.H.; Mcllwraith, C.W. Treatment of experimental equine osteoarthritis by invivo delivery of the equine interleukin-1 receptorantagonist gene. Gene Ther. 2002, 9, 12–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCoy, A.M. Animal models of osteoarthritis: Comparisons and key considerations. Vet. Pathol. 2015, 52, 803–818. [Google Scholar] [CrossRef] [PubMed]
- Casal, M.; Haskins, M. Large animal models and gene therapy. Eur. J. Hum. Genet. 2006, 14, 266–272. [Google Scholar] [CrossRef]
- Gurda, B.L.; Bradbury, A.M.; Vite, C.H. Canine and feline models of human genetic diseases and their contributions to advancing clinical therapies. Yale J. Biol. Med. 2017, 90, 417–431. [Google Scholar] [PubMed]
- Shelton, G.D.; Engvall, E. Canine and feline models of human inherited muscle diseases. Neuromuscul. Disord. 2005, 15, 127–138. [Google Scholar] [CrossRef]
- Houser, S.R.; Margulies, K.B.; Murphy, A.M.; Spinale, F.G.; Francis, G.S.; Prabhu, S.D.; Rockman, H.A.; Kass, D.A.; Molkentin, J.D.; Sussman, M.A.; et al. Animal models of heart failure: A scientific statement from the American Heart Association. Circ. Res. 2012, 111, 131–150. [Google Scholar] [CrossRef] [Green Version]
- Giuliano, A. Companion animal model in translational oncology; feline oral squamous cell carcinoma and canine oral melanoma. Biology 2021, 11, 54. [Google Scholar] [CrossRef]
- Cekanova, M.; Rathore, K. Animal models and therapeutic molecular targets of cancer: Utility and limitations. Drug Des. Devel. Ther. 2014, 8, 1911–1921. [Google Scholar] [CrossRef] [Green Version]
- Moran, C.J.; Ramesh, A.; Brama, P.A.; O’Byrne, J.M.; O’Brien, F.J.; Levingstone, T.J. The benefits and limitations of animal models for translational research in cartilage repair. J. Exp. Orthop. 2016, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Cauvin, A.J.; Peters, C.; Brennan, F. Advantages and limitations of commonly used nonhuman primate species in research and development of biopharmaceuticals. In The Nonhuman Primate in Nonclinical Drug Development and Safety Assessment; Bluemel, J., Korte, S., Schenck, E., Weinbauer, G.F., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 379–395. [Google Scholar]
- Carvalho, C.; Gaspar, A.; Knight, A.; Vicente, L. Ethical and scientific pitfalls concerning laboratory research with non-human primates, and possible solutions. Animals 2018, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- DeGrazia, D. Nonhuman primates, human need, and Eehical constraints. Hastings Cent. Rep. 2016, 46, 27–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorby-Adams, A.J.; Vink, R.; Turner, R.J. Large animal models of stroke and traumatic brain injury as translational tools. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R165–R190. [Google Scholar] [CrossRef] [PubMed]
- Genain, M.A.; Morlet, A.; Herrtage, M.; Muresian, H.; Anselme, F.; Latremouille, C.; Laborde, F.; Behr, L.; Borenstein, N. Comparative anatomy and angiography of the cardiac coronary venous system in four species: Human, ovine, porcine, and canine. J. Vet. Cardiol. 2018, 20, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Karwacki, Z.K.; Witkowska, M.P.; Karwacka, M.; Moryś, J. The technique of inhalation anaethesia in experimental investigation in the rat. Folia Morphol. 2006, 65, 414–418. [Google Scholar]
- Sengupta, P. The laboratory rat: Relating its age with human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar]
- Piumatti, M.; Palazzo, O.; La Rosa, C.; Crociara, P.; Parolisi, R.; Luzzati, F.; Levy, F.; Bonfanti, L. Non-newly generated, “immature" neurons in the sheep brain are not restricted to cerebral cortex. J. Neurosci. 2018, 38, 826–842. [Google Scholar] [CrossRef] [Green Version]
- Morton, A.J.; Howland, D.S. Large genetic animal models of Huntington’s Disease. J. Huntingt. Dis. 2013, 2, 3–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, D.B.; McKinley, M. The Sheep. Fact Sheet for ANZCCART. 2009. Available online: https://www.researchgate.net/publication/275037130 (accessed on 7 May 2020).
- Sund-Levander, M.; Forsberg, C.; Wahren, L.K. Normal oral, rectal, tympanic and axillary body temperature in adult men and women: A systematic literature review. Scand. J. Caring Sci. 2001, 16, 122–128. [Google Scholar] [CrossRef]
- Forkan, A.R.M.; Khalil, I.; Tari, Z.; Foufou, S.; Bouras, A. A context-aware approach for long-term behavioural change detection and abnormality prediction in ambient assisted living. Pattern Recog. 2015, 48, 628–641. [Google Scholar] [CrossRef]
- Kling, M.A. A review of respiratory system anatomy, physiology, and disease in the mouse, rat, hamster, and gerbil. Vet. Clin. N. Am. Exot. Anim. Pract. 2011, 14, 287–337, vi. [Google Scholar] [CrossRef]
- Norman, R.A.; Coleman, T.G.; Dent, A.C. Continuous monitoring of arterial pressure indicates sinoaortic denervated rats are not hypertensive. Hypertension 1981, 3, 119–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothbaum, D.A.; Shaw, D.J.; Angell, C.S.; Shock, N.W. Cardiac performance in the unanesthetized senescent male rat. J. Gerontol. 1973, 28, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Yelderman, M.; Quinn, M.D.; McKown, R.C.; Eberhart, R.C.; Dollar, M.L. Continuous thermodilution cardiac output measurement in sheep. J. Thorac. Cardiovasc. Surg. 1992, 104, 315–320. [Google Scholar] [CrossRef]
- Murray, S.J.; Black, B.L.; Reid, S.J.; Rudiger, S.R.; Simon Bawden, C.; Snell, R.G.; Waldvogel, H.J.; Faull, R.L.M. Chemical neuroanatomy of the substantia nigra in the ovine brain. J. Chem. Neuroanat. 2019, 97, 43–56. [Google Scholar] [CrossRef]
- Roth, J.A.; Tuggle, C.K. Livestock models in translational medicine. ILAR J. 2015, 56, 1–6. [Google Scholar] [CrossRef] [Green Version]
- New Zealand Sheepbreeders’ Association. Sheep Breeds. Available online: http://www.nzsheep.co.nz/index.php?page=sheep-breeds (accessed on 16 December 2021).
- Camacho, P.; Fan, H.; Liu, Z.; He, J.Q. Large mammalian animal models of heart disease. J. Cardiovasc. Dev. Dis. 2016, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Back, S.A.; Riddle, A.; Dean, J.; Hohimer, A.R. The instrumented fetal sheep as a model of cerebral white matter injury in the premature infant. Neurotherapeutics 2012, 9, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Ella, A.; Barriere, D.A.; Adriaensen, H.; Palmer, D.N.; Melzer, T.R.; Mitchell, N.L.; Keller, M. The development of brain magnetic resonance approaches in large animal models for preclinical research. Anim. Front. 2019, 9, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Lee, S.D.; Park, M.Y.; Foley, L.; Purcell-Estabrook, E.; Kim, H.; Yoo, S.S. Functional and diffusion tensor magnetic resonance imaging of the sheep brain. BMC Vet. Res. 2015, 11, 262. [Google Scholar] [CrossRef] [Green Version]
- Baskin, D.S.; Browning, J.L.; Widmayer, M.A.; QingZhu, Z.; Grossman, R.G. Development of a model for Parkinson’s disease in sheep using unilateral intracarotid injection of MPTP via slow continuous infusion. Life Sci. 1994, 54, 471–479. [Google Scholar] [CrossRef]
- Jacobsen, J.C.; Bawden, C.S.; Rudiger, S.R.; McLaughlan, C.J.; Reid, S.J.; Waldvogel, H.J.; MacDonald, M.E.; Gusella, J.F.; Walker, S.K.; Kelly, J.M.; et al. An ovine transgenic Huntington’s disease model. Hum. Mol. Genet. 2010, 19, 1873–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morton, A.J.; Avanzo, L. Executive decision-making in the domestic sheep. PLoS ONE 2011, 6, e15752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vink, R. Large animal models of traumatic brain injury. J. Neurosci. Res. 2018, 96, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerdts, V.; Wilson, H.L.; Meurens, F.; van Drunen Littel-van den Hurk, S.; Wilson, D.; Walker, S.; Wheler, C.; Townsend, H.; Potter, A.A. Large animal models for vaccine development and testing. ILAR J. 2015, 56, 53–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hessler, M.; Pinto, B.B.; Arnemann, P.H.; Kampmeier, T.G.; Seidel, L.; Morelli, A.; Van Aken, H.; Westphal, M.; Rehberg, S.; Ertmer, C. Differential effects of selective and nonselective potassium channel inhibitors in ovine endotoxemic shock (macrocirculation) and in a rat model of septic shock (microcirculation). Shock 2019, 51, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Dekker, S.; van Geemen, D.; van den Bogaerdt, A.J.; Driessen-Mol, A.; Aikawa, E.; Smits, A. Sheep-specific immunohistochemical panel for the evaluation of regenerative and inflammatory processes in tissue-engineered heart valves. Front. Cardiovasc. Med. 2018, 5, 105. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Teng, F.; Xiao, J.; Bao, W.; Zhou, Q.; Sun, Q.; Meng, G. Implantation of a modified stented bovine pulmonary valve in a beating heart sheep model. J. Card. Surg. 2017, 32, 508–513. [Google Scholar] [CrossRef]
- Farraha, M.; Lu, J.; Trivic, I.; Barry, M.A.; Chong, J.; Kumar, S.; Kizana, E. Development of a sheep model of atrioventricular block for the application of novel therapies. PLoS ONE 2020, 15, e0229092. [Google Scholar] [CrossRef] [Green Version]
- Oner, K.; Bilgen, C.; Oran, I.; Kirazli, T.; Memis, A.; Ovul, I.; Yilmaz, O. “Extarterectomy” for tumors invading the carotid artery: An animal model. Skull Base 2007, 17, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Porter, D.T.; Moore, A.M.; Cobern, J.A.; Padmanabhan, V.; Goodman, R.L.; Coolen, L.M.; Lehman, M.N. Prenatal testosterone exposure alters GABAergic synaptic inputs to GnRH and KNDy neurons in a sheep model of polycystic ovarian syndrome. Endocrinology 2019, 160, 2529–2542. [Google Scholar] [CrossRef]
- Padmanabhan, V.; Veiga-Lopez, A. Sheep models of polycystic ovary syndrome phenotype. Mol. Cell. Endocrinol. 2013, 373, 8–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngamprasertwong, P.; Dong, M.; Niu, J.; Venkatasubramanian, R.; Vinks, A.A.; Sadhasivam, S. Propofol pharmacokinetics and estimation of fetal propofol exposure during mid-gestational fetal surgery: A maternal-fetal sheep model. PLoS ONE 2016, 11, e0146563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barry, J.S.; Anthony, R.V. The Pregnant Sheep as a Model for Human Pregnancy. Theriogenology 2008, 69, 55–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luján, L.; Perez, M.; Salazar, E.; Alvarez, N.; Gimeno, M.; Pinczowski, P.; Irusta, S.; Santamaria, J.; Insausti, N.; Cortes, Y.; et al. Autoimmune/autoinflammatory syndrome induced by adjuvants (ASIA syndrome) in commercial sheep. Immunol. Res. 2013, 56, 317–324. [Google Scholar] [CrossRef]
- Diogo, C.C.; Camassa, J.A.; Pereira, J.E.; Costa, L.M.D.; Filipe, V.; Couto, P.A.; Geuna, S.; Mauricio, A.C.; Varejao, A.S. The use of sheep as a model for studying peripheral nerve regeneration following nerve injury: Review of the literature. Neurol. Res. 2017, 39, 926–939. [Google Scholar] [CrossRef]
- Levy, F.; Batailler, M.; Meurisse, M.; Migaud, M. Adult neurogenesis in sheep: Characterization and contribution to reproduction and behavior. Front. Neurosci. 2017, 11, 570. [Google Scholar] [CrossRef]
- Low, V.F.; Faull, R.L.; Bennet, L.; Gunn, A.J.; Curtis, M.A. Neurogenesis and progenitor cell distribution in the subgranular zone and subventricular zone of the adult sheep brain. Neuroscience 2013, 244, 173–187. [Google Scholar] [CrossRef]
- Boltze, J.; Förschler, A.; Nitzsche, B.; Waldmin, D.; Hoffmann, A.; Boltze, C.M.; Dreyer, A.Y.; Goldammer, A.; Reischauer, A.; Hartig, W.; et al. Permanent middle cerebral artery occlusion in sheep: A novel large animal model of focal cerebral ischemia. J. Cereb. Blood Flow Metab. 2008, 28, 1951–1964. [Google Scholar] [CrossRef] [Green Version]
- Wells, A.J.; Vink, R.; Blumbergs, P.C.; Brophy, B.P.; Helps, S.C.; Knox, S.J.; Turner, R.J. A surgical model of permanent and transient middle cerebral artery stroke in the sheep. PLoS ONE 2012, 7, e42157. [Google Scholar] [CrossRef] [Green Version]
- Joyeux, L.; Engels, A.C.; Van Der Merwe, J.; Aertsen, M.; Patel, P.A.; Deprez, M.; Khatoun, A.; Pranpanus, S.; da Cunha, M.; De Vleeschauwer, S.; et al. Validation of the fetal lamb model of spina bifida. Sci. Rep. 2019, 9, 9327. [Google Scholar] [CrossRef]
- Lewis, S.B.; Finnie, J.W.; Blumbergs, P.C.; Scott, G.; Manavis, J.; Brown, C.; Reilly, P.L.; Jones, N.R.; McLean, A.J. A head impact model of early axonal injury in the sheep. J. Neurotrauma 1996, 13, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Reid, S.J.; McKean, N.E.; Henty, K.; Portelius, E.; Blennow, K.; Rudiger, S.R.; Bawden, C.S.; Handley, R.R.; Verma, P.J.; Faull, R.L.M.; et al. Alzheimer’s disease markers in the aged sheep (Ovis aries). Neurobiol. Aging 2017, 58, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Hammock, B.D.; Beale, A.M.; Work, T.; Gee, S.J.; Gunther, R.; Higgins, R.J.; Shinka, T.; Castagnoli, N. A sheep model for MPTP induced Parkinson-like symptoms. Life Sci. 1989, 45, 1601–1608. [Google Scholar] [CrossRef]
- Perentos, N.; Martins, A.Q.; Watson, T.C.; Bartsch, U.; Mitchell, N.L.; Palmer, D.N.; Jones, M.W.; Morton, A.J. Translational neurophysiology in sheep: Measuring sleep and neurological dysfunction in CLN5 Batten disease affected sheep. Brain 2015, 138, 862–874. [Google Scholar] [CrossRef] [PubMed]
- Bourke, C. Molybdenum deprivation, purine ingestion and an astrocyte-associated motor neurone syndrome in sheep: Assumed clinical effects of inosine. Aust. Vet. J. 2015, 93, 79–83. [Google Scholar] [CrossRef]
- Mitchell, N.L.; Russell, K.N.; Wellby, M.P.; Wicky, H.E.; Schoderboeck, L.; Barrell, G.K.; Melzer, T.R.; Gray, S.J.; Hughes, S.M.; Palmer, D.N. Longitudinal in vivo monitoring of the CNS demonstrates the efficacy of gene therapy in a sheep model of CLN5 Batten disease. Mol. Ther. 2018, 26, 2366–2378. [Google Scholar] [CrossRef] [Green Version]
- Opdam, H.I.; Federico, P.; Jackson, G.D.; Buchanan, J.; Abbott, D.F.; Fabinyi, G.C.A.; Syngeniotis, A.; Vosmansky, M.; Archer, J.S.; WEllard, R.M.; et al. A sheep model for the study of focal epilepsy with concurrent intracranial EEG and functional MRI. Epilepsia 2002, 43, 779–787. [Google Scholar] [CrossRef] [Green Version]
- Wilkes, D.; Li, G.; Angeles, C.F.; Patterson, J.T.; Huang, L.Y. A large animal neuropathic pain model in sheep: A strategy for improving the predictability of preclinical models for therapeutic development. J. Pain Res. 2012, 5, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Broom, M.F.; Zhou, C.; Broom, J.E.; Barwell, K.J.; Jolly, R.D.; Hill, D.F. Ovine neuronal ceroid lipofuscinosis: A large animal model syntenic with the human neuronal ceroid lipofuscinosis variant CLN6. J. Med. Genet. 1998, 35, 717–721. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.; Fredericks, D.C.; Safayi, S.; DeVries-Watson, N.A.; Holland, M.T.; Nagel, S.J.; Gillies, G.T.; Howard, M.A., III. Ovine hemisection model of spinal cord injury. J. Investig. Surg. 2019, 34, 380–392. [Google Scholar] [CrossRef]
- Balaraman, S.; Lunde, E.R.; Sawant, O.; Cudd, T.A.; Washburn, S.E.; Miranda, R.C. Maternal and neonatal plasma microRNA biomarkers for fetal alcohol exposure in an ovine model. Alcohol. Clin. Exp. Res. 2014, 38, 1390–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karageorgos, L.; Lancaster, M.J.; Nimmo, J.S.; Hopwood, J.J. Gaucher disease in sheep. J. Inherit. Metab. Dis. 2010, 34, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.W.G.; Sandoz, B.; Dutschke, J.K.; Finnie, J.W.; Turner, R.J.; Blumbergs, P.C.; Manavis, J.; Vink, R. Biomechanical studies in an ovine model of non-accidental head injury. J. Biomech. 2014, 47, 2578–2583. [Google Scholar] [CrossRef] [PubMed]
- Vincent, K.L.; Vargas, G.; Bourne, N.; Galvan-Turner, V.; Saada, J.I.; Lee, G.H.; Sbrana, E.; Motamedi, M. Image-based noninvasive evaluation of colorectal mucosal injury in sheep after topical application of microbicides. Sex. Transm. Dis. 2013, 40, 854–859. [Google Scholar] [CrossRef] [Green Version]
- Inglis, G.D.; Kastrlic, J.P.; Uwiera, R.R.E. Catheterization of intestinal loops in ruminants does not adversely affect loop function. Comp. Med. 2010, 60, 469–478. [Google Scholar]
- Kemp, M.W.; Kannan, P.S.; Saito, M.; Newnham, J.P.; Cox, T.; Jobe, A.H.; Kramer, B.W.; Kallapur, S.G. Selective exposure of the fetal lung and skin/amnion (but not gastro-intestinal tract) to LPS elicits acute systemic inflammation in fetal sheep. PLoS ONE 2013, 8, e63355. [Google Scholar] [CrossRef] [Green Version]
- Gray, M.E.; Sullivan, P.; Marland, J.R.K.; Greenhalgh, S.N.; Meehan, J.; Gregson, R.; Clutton, R.E.; Cousens, C.; Griffiths, D.J.; Murray, A.; et al. A novel translational ovine pulmonary adenocarcinoma model for human lung cancer. Front. Oncol. 2019, 9, 534. [Google Scholar] [CrossRef]
- Greene, C.A.; Misra, S.L.; Lee, H.; McKelvie, J.; Kapadia, K.; McFarlane, R.; McGhee, C.N.J.; Green, C.R.; Sherwin, T. The sheep cornea: Structural and clinical characteristics. Curr. Eye Res. 2018, 43, 1432–1438. [Google Scholar] [CrossRef]
- Mohammadi, S.F.; Mazouri, A.; Jabbarvand, M.; Rahman, A.N.; Mohammadi, A. Sheep practice eye for ophthalmic surgery training in skills laboratory. J. Cataract Refract. Surg. 2011, 37, 987–991. [Google Scholar] [CrossRef]
- Kerdraon, Y.A.; Downie, J.A.; Suaning, G.J.; Capon, M.R.; Coroneo, M.T.; Lovell, N.H. Development and surgical implantation of a vision prosthesis model into the ovine eye. Clin. Exp. Opthal. 2002, 30, 36–40. [Google Scholar] [CrossRef]
- Williams, K.A.; Standfield, S.D.; Mills, R.A.; Takano, T.; Larkin, D.F.; Krishna, R.; Russ, G.R.; Coster, D.J. A new model of orthotopic penetrating corneal transplantation in the sheep: Graft survival, phenotypes of graft-infiltrating cells and local cytokine production. Aust. N. Z. J. Ophthalmol. 1999, 27, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Abdalmula, A.; Washington, E.A.; House, J.V.; Dooley, L.M.; Blacklaws, B.A.; Ghosh, P.; Bailey, S.R.; Kimpton, W.G. Clinical and histopathological characterization of a large animal (ovine) model of collagen-induced arthritis. Vet. Immunol. Immunopathol. 2014, 159, 83–90. [Google Scholar] [CrossRef]
- Rohrbach, H.; Zeiter, S.; Andersen, O.K.; Wieling, R.; Spadavecchia, C. Quantitative assessment of the nociceptive withdrawal reflex in healthy, non-medicated experimental sheep. Physiol. Behav. 2014, 129, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Oheim, R.; Beil, F.T.; Kohne, T.; Wehner, T.; Barvencik, F.; Ignatius, A.; Amling, M.; Clarke, I.J.; Pogoda, P. Sheep model for osteoporosis: Sustainability and biomechanical relevance of low turnover osteoporosis induced by hypothalamic-pituitary disconnection. J. Orthop. Res. 2013, 31, 1067–1074. [Google Scholar] [CrossRef]
- Mageed, M.; Berner, D.; Julke, H.; Hohaus, C.; Brehm, W.; Gerlach, K. Is sheep lumbar spine a suitable alternative model for human spinal researches? Morphometrical comparison study. Lab. Anim. Res. 2013, 29, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.K.; Pinzon, C.; Huggins, S.; Pryor, J.H.; Falck, A.; Herman, F.; Oldeschulte, J.; Chavez, M.B.; Foster, B.L.; White, S.H.; et al. Genetic engineering a large animal model of human hypophosphatasia in sheep. Sci. Rep. 2018, 8, 16945. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, A.; Pelletier, M.H.; Yu, Y.; Christou, C.; Walsh, W.R. A sheep model for cancellous bone healing. Front. Surg. 2014, 1, 37. [Google Scholar] [CrossRef] [Green Version]
- Harms, C.; Helms, K.; Taschner, T.; Stratos, I.; Ignatius, A.; Gerber, T.; Lenz, S.; Rammelt, S.; Vollmar, B.; Mittlmeier, T. Osteogenic capacity of nanocrystalline bone cement in a weight-bearing defect at the ovine tibial metaphysis. Int. J. Nanomed. 2012, 7, 2883–2889. [Google Scholar] [CrossRef] [Green Version]
- Badis, D.; Omar, B. The effectiveness of platelet-rich plasma on the skin wound healing process: A comparative experimental study in sheep. Vet. World 2018, 11, 800–808. [Google Scholar] [CrossRef]
- Martinello, T.; Gomiero, C.; Perazzi, A.; Iacopetti, I.; Gemignani, F.; DeBenedictis, G.M.; Ferro, S.; Zuin, M.; Martines, E.; Brun, P.; et al. Allogeneic mesenchymal stem cells improve the wound healing process of sheep skin. BMC Vet. Res. 2018, 14, 202. [Google Scholar] [CrossRef] [Green Version]
- Favre-Inhofer, A.; Carbonnel, M.; Revaux, A.; Sandra, O.; Mougenot, V.; Bosc, R.; Gelin, V.; Rafii, A.; Hersant, B.; Vialard, F.; et al. Critical steps for initiating an animal uterine transplantation model in sheep: Experience from a case series. Int. J. Surg. 2018, 60, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Luis-Lima, S.; Mas-Sanmartin, C.; Rodriguez-Rodriguez, A.E.; Porrini, E.; Ortiz, A.; Gaspari, F.; Diaz-Martin, L.; Asberg, A.; Jenssen, T.; Jimenez-Sosa, A.; et al. A simplified iohexol-based method to measure renal function in sheep models of renal disease. Biology 2020, 9, 259. [Google Scholar] [CrossRef] [PubMed]
- Bujok, J.; Walski, T.; Czerski, A.; Galecka, K.; Grzeszczuk-Kuc, K.; Zawadzki, W.; Witkiewicz, W.; Komorowska, M. Sheep model of haemodialysis treatment. Lab. Anim. 2018, 52, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.E. The cytoarchitectural study of the sheep cortex. J. Comp. Neurol. 1942, 76, 1–55. [Google Scholar] [CrossRef]
- Sun, T.; Hevner, R.F. Growth and folding of the mammalian cerebral cortex: From molecules to malformations. Nat. Rev. Neurosci. 2014, 15, 217–232. [Google Scholar] [CrossRef] [Green Version]
- McKenzie, J.S.; Chai, S.; Giles, M.E.; McKinley, M.J. Dopaminergic system in the basal ganglia of sheep. In The Basal Ganglia VII; Springer: Boston, MA, USA, 2002; pp. 107–116. [Google Scholar]
- Feigin, V.L.; Nichols, E.; Alam, T.; Bannick, M.S.; Beghi, E.; Blake, N.; Culpepper, W.J.; Dorsey, E.R.; Elbaz, A.; Ellenbogen, R.G.; et al. Global, regional, and national burden of neurological disorders, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 459–480. [Google Scholar] [CrossRef] [Green Version]
- Bennett, J.E.; Stevens, G.A.; Mathers, C.D.; Bonita, R.; Rehm, J.; Kruk, M.E.; Riley, L.M.; Dain, K.; Kengne, A.P.; Chalkidou, K.; et al. NCD Countdown 2030: Worldwide trends in non-communicable disease mortality and progress towards Sustainable Development Goal target 3.4. Lancet 2018, 392, 1072–1088. [Google Scholar] [CrossRef] [Green Version]
- Mak, I.W.Y.; Evaniew, N.; Ghert, M. Lost in translation: Animal models and clinical trials in cancer treatment. Am. J. Transl. Stud. 2014, 6, 114–118. [Google Scholar]
- Vandamme, T.F. Rodent models for human diseases. Eur. J. Pharmacol. 2015, 759, 84–89. [Google Scholar] [CrossRef]
- Birbeck, G.L.; Meyer, A.C.; Ogunniyi, A. Nervous system disorders across the life course in resource-limited settings. Nature 2015, 527, S167–S171. [Google Scholar] [CrossRef]
- Collier, T.J.; Kanaan, N.M.; Kordower, J.H. Aging and Parkinson’s disease: Different sides of the same coin? Mov. Disord. 2017, 32, 983–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kip, E.; Parr-Brownlie, L.C. Reducing neuroinflammation via therapeutic compounds and lifestyle to prevent or delay progression of Parkinson’s disease. Ageing Res. Rev. 2022, 78, 101618. [Google Scholar] [CrossRef] [PubMed]
- Callixte, K.T.; Clet, T.B.; Jacques, D.; Faustin, Y.; Francois, D.J.; Maturin, T.T. The pattern of neurological diseases in elderly people in outpatient consultations in Sub-Saharan Africa. BMC Res. Notes 2015, 8, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peruffo, A.; Corain, L.; Bombardi, C.; Centelleghe, C.; Grisan, E.; Graic, J.M.; Bontempi, P.; Grandis, A.; Cozzi, B. The motor cortex of the sheep: Laminar organization, projections and diffusion tensor imaging of the intracranial pyramidal and extrapyramidal tracts. Brain Struct. Funct. 2019, 224, 1933–1946. [Google Scholar] [CrossRef] [PubMed]
- John, S.E.; Lovell, T.J.H.; Opie, N.L.; Wilson, S.; Scordas, T.C.; Wong, Y.T.; Rind, G.S.; Ronayne, S.; Bauquier, S.H.; May, C.N.; et al. The ovine motor cortex: A review of functional mapping and cytoarchitecture. Neurosci. Biobehav. Rev. 2017, 80, 306–315. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, A.; Sinnott, B.; Kuchel, T.R.; Perumal, S.R.; Fraser, C.K.; Hemsley, K.M.; Morton, A.J. Neurological examination of sheep (Ovis aries) with unilateral and bilateral quinolinic acid lesions of the striatum assessed by magnetic resonance imaging. J. Neurol. Exp. Neurosci. 2019, 5, 56–67. [Google Scholar] [CrossRef] [Green Version]
- Oswald, M.J.; Palmer, D.N.; Kay, G.W.; Barwell, K.J.; Cooper, J.D. Location and connectivity determine GABAergic interneuron survival in the brains of South Hampshire sheep with CLN6 neuronal ceroid lipofuscinosis. Neurobiol. Dis. 2008, 32, 50–65. [Google Scholar] [CrossRef] [Green Version]
- Massano, J.; Bhatia, K.P. Clinical approach to Parkinson’s disease: Features, diagnosis, and principles of management. Cold Spring Harb. Perspect. Med. 2012, 2, a008870. [Google Scholar] [CrossRef]
- Emborg, M.E. Nonhuman primate models of Parkinson’s disease. ILAR 2007, 48, 339–355. [Google Scholar] [CrossRef] [Green Version]
- Lentz, L.; Zhao, Y.; Kelly, M.T.; Schindeldecker, W.; Goetz, S.; Nelson, D.E.; Raike, R.S. Motor behaviors in the sheep evoked by electrical stimulation of the subthalamic nucleus. Exp. Neurol. 2015, 273, 69–82. [Google Scholar] [CrossRef]
- McColgan, P.; Tabrizi, S.J. Huntington’s disease: A clinical review. Eur. J. Neurol. 2018, 25, 24–34. [Google Scholar] [CrossRef]
- Pfister, E.L.; DiNardo, N.; Mondo, E.; Borel, F.; Conroy, F.; Fraser, C.; Gernoux, G.; Han, X.; Hu, D.; Johnson, E.; et al. Artificial miRNAs reduce human mutant Huntingtin throughout the striatum in a transgenic sheep model of Huntington’s disease. Hum. Gene Ther. 2018, 29, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Bondi, M.W.; Edmonds, E.C.; Salmon, D.P. Alzheimer’s disease: Past, present, and future. J. Int. Neuropsychol. Soc. 2017, 23, 818–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, S.J.; Shuttleworth, E.C.; Freidenberg, D.L. Neuropsychological differences between the dementias of Alzheimer’s and Parkinson’s diseases. Arch. Neurol. 1989, 46, 1287–1291. [Google Scholar] [CrossRef] [PubMed]
- Breijyeh, Z.; Karaman, R. Comprehensive review on Alzheimer’s disease: Causes and treatment. Molecules 2020, 25, 5789. [Google Scholar] [CrossRef]
- Johnson, T.B.; Cain, J.T.; White, K.A.; Ramirez-Montealegre, D.; Pearce, D.A.; Weimer, J.M. Therapeutic landscape for Batten disease: Current treatments and future prospects. Nat. Rev. Neurol. 2019, 15, 161–178. [Google Scholar] [CrossRef]
- Linterman, K.S.; Palmer, D.N.; Kay, G.W.; Barry, L.A.; Mitchell, N.L.; McFarlane, R.G.; Black, M.A.; Sands, M.S.; Hughes, S.M. Lentiviral-mediated gene transfer to the sheep brain: Implications for gene therapy in Batten disease. Hum. Gene Ther. 2011, 22, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Weber, K.; Pearce, D.A. Large animal models for Batten disease: A review. J. Child Neurol. 2013, 28, 1123–1127. [Google Scholar] [CrossRef]
- Entrican, G.; Wattegedera, S.R.; Griffiths, D.J. Exploiting ovine immunology to improve the relevance of biomedical models. Mol. Immunol. 2015, 66, 68–77. [Google Scholar] [CrossRef]
- Jolly, R.D.; Martinus, R.D.; Palmer, D.N. Sheep and other animals with ceroid lipofuscinoses: Their relevance to Batten disease. Am. J. Med. Genet. 1992, 42, 609–614. [Google Scholar] [CrossRef]
- Knolle, F.; Goncalves, R.P.; Morton, A.J. Sheep recognize familiar and unfamiliar human faces from two-dimensional images. R. Soc. Open Sci. 2017, 4, 171228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veissier, I.; Boissy, A.; Desire, L.; Greiveldinger, L. Animals’ emotions: Studies in sheep using appraisal theories. Anim. Welfare 2009, 18, 347–354. [Google Scholar]
- Rashid, S.T.; Salacinski, H.J.; Hamilton, G.; Seifalian, A.M. The use of animal models in developing the discipline of cardiovascular tissue engineering: A review. Biomaterials 2004, 25, 1627–1637. [Google Scholar] [CrossRef]
- DiVincenti, L., Jr.; Westcott, R.; Lee, C. Sheep (Ovis aries) as a model for cardiovascular surgery and management before, during, and after cardiopulmonary bypass. J. Am. Assoc. Lab. Anim. Sci. 2014, 53, 439–448. [Google Scholar]
- Katz, M.G.; Kendle, A.P.; Fargnoli, A.S.; Mihalko, K.L.; Bridges, C.R. Sheep (Ovis aries) as a model for cardiovascular surgery and management before, during, and after cardiopulmonary bypass. J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 7–8. [Google Scholar]
- Ngamprasertwong, P.; Michelfelder, E.C.; Arbabi, S.; Choi, Y.S.; Statile, C.; Ding, L.; Boat, A.; Eghtesady, P.; Holland, K.; Sadhasivam, S. Anesthetic techniques for fetal surgery: Effects of maternal anesthesia on intraoperative fetal outcomes in a sheep model. Anesthesiology 2013, 118, 796–808. [Google Scholar] [CrossRef] [Green Version]
- Wani, S.A.; Praharaj, M.R.; Sahu, A.R.; Khan, R.I.N.; Saxena, S.; Rajak, K.K.; Muthuchelvan, D.; Sahoo, A.; Mishra, B.; Singh, R.K.; et al. Systems biology behind immunoprotection of both sheep and goats after Sungri/96 PPRV vaccination. mSystems 2021, 30, e00820. [Google Scholar] [CrossRef]
- Hein, W.R.; Griebel, P.J. A road less travelled: Large animalmodels in immunological research. Nat. Rev. Immunol. 2002, 3, 79–84. [Google Scholar] [CrossRef]
- Hecht, S.J.; Stedman, K.E.; Carlson, J.O.; DeMartini, J.C. Distribution of endogenous type B and type D sheep retrovirus sequences in ungulates and other mammals. Proc. Natl. Acad. Sci. USA 1996, 93, 3297–3302. [Google Scholar] [CrossRef] [Green Version]
- Flores, T.J.; Nguyen, V.B.; Widdop, R.E.; Sutherland, M.R.; Polglase, G.R.; Abud, H.E.; Black, M.J. Morphology and function of the lamb ileum following preterm birth. Front. Pediatr. 2018, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Medjekal, S.; Ghadbane, M. Sheep digestive physiology and constituents of feeds. In Sheep Farming—An Approach to Feed, Growth and Health; Monteiro, A., Ed.; IntechOpen Limited: London, UK, 2021. [Google Scholar]
- Sedgmen, B.J.; Lofthouse, S.A.; Scheerlinck, J.Y.; Meeusen, E.N.T. Cellular and molecular characterisation of the ovine rectal mucosal environment. Vet. Immunol. Immunopathol. 2002, 86, 215–220. [Google Scholar] [CrossRef]
- Youssef, G.; Wallace, W.A.; Dagleish, M.P.; Cousens, C.; Griffiths, D.J. Ovine pulmonary adenocarcinoma: A large animal model for human lung cancer. ILAR J. 2015, 56, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Collie, D.; Govan, J.; Wright, S.; Thornton, E.; Tennant, P.; Smith, S.; Doherty, C.; McLachlan, G. A lung segmental model of chronic Pseudomonas infection in sheep. PLoS ONE 2013, 8, e67677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martini, L.; Fini, M.; Giavaresi, G.; Giardino, R. Sheep model in orthopedic research: A literature review. Comp. Med. 2001, 51, 292–299. [Google Scholar] [PubMed]
- Casaroli, G.; Galbusera, F.; Jonas, R.; Schlager, B.; Wilke, H.J.; Villa, T. A novel finite element model of the ovine lumbar intervertebral disc with anisotropic hyperelastic material properties. PLoS ONE 2017, 12, e0177088. [Google Scholar] [CrossRef]
- Wilke, H.J.; Kettler, A.; Wenger, K.H.; Claes, L.E. Anatomy of the sheep spine and its comparison to the human spine. Anat. Rec. 1997, 247, 542–555. [Google Scholar] [CrossRef]
- Iacopetti, I.; Perazzi, A.; Martinello, T.; Gemignani, F.; Patruno, M. Hyaluronic acid, Manuka honey and Acemannan gel: Wound-specific applications for skin lesions. Res. Vet. Sci. 2020, 129, 82–89. [Google Scholar] [CrossRef]
- Liebsch, C.; Bucan, V.; Menger, B.; Kohne, F.; Waldmann, K.H.; Vaslaitis, D.; Vogt, P.M.; Strauss, S.; Kuhbier, J.W. Preliminary investigations of spider silk in wounds in vivo—Implications for an innovative wound dressing. Burns 2018, 44, 1829–1838. [Google Scholar] [CrossRef]
- Martines, E.; Brun, P.; Cavazzana, R.; Cordaro, L.; Zuin, M.; Martinello, T.; Gomiero, C.; Perazzi, A.; Melotti, L.; Maccatrozzo, L.; et al. Wound healing improvement in large animals using an indirect helium plasma treatment. Clin. Plasma Med. 2020, 17–18, 100095. [Google Scholar] [CrossRef]
- Melotti, L.; Martinello, T.; Perazzi, A.; Martines, E.; Zuin, M.; Modenese, D.; Cordaro, L.; Ferro, S.; Maccatrozzo, L.; Iacopetti, I.; et al. Could cold plasma act synergistically with allogeneic mesenchymal stem cells to improve wound skin regeneration in a large size animal model? Res. Vet. Sci. 2021, 136, 97–110. [Google Scholar] [CrossRef]
- Breyer, M.D.; Susztak, K. Developing treatments for chronic kidney disease in the 21st century. Semin. Nephrol. 2016, 36, 436–447. [Google Scholar] [CrossRef] [Green Version]
- Alvites, R.D.; Branquinho, M.V.; Sousa, A.C.; Lopes, B.; Sousa, P.; Mendonca, C.; Atayde, L.M.; Mauricio, A.C. Small Ruminants and Its Use in Regenerative Medicine: Recent Works and Future Perspectives. Biology 2021, 10, 249. [Google Scholar] [CrossRef] [PubMed]
- Ambros, J.T.; Herrero-Fresneda, I.; Borau, O.G.; Boira, J.M. Ischemic preconditioning in solid organ transplantation: From experimental to clinics. Transpl. Int. 2007, 20, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Diao, Y.; Chen, G.; Tanaka, A.; Eastwood, G.M.; Bellomo, R. Remote ischemic conditioning for kidney protection: A meta-analysis. J. Crit. Care 2016, 33, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Luna, M.J.; Ramirez-Martinez, L.A.; Sarmiento Silva, R.E.; Cruz Lazo, C.; Perez Torres, A.; Sanchez-Betancourt, J.I. Evidence of respiratory syncytial virus and parainfluenza-3 virus in Mexican sheep. Virusdisease 2017, 28, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Cornick, N.A.; Booher, S.L.; Casey, T.A.; Moon, H.W. Persistent colonization of sheep by Escherichia coli O157:H7 and other E. coli pathotypes. Appl. Environ. Microbiol. 2000, 66, 4926–4934. [Google Scholar] [CrossRef] [Green Version]
- Aragon-Aranda, B.; de Miguel, M.J.; Martinez-Gomez, E.; Zuniga-Ripa, A.; Salvador-Bescos, M.; Moriyon, I.; Iriarte, M.; Munoz, P.M.; Conde-Alvarez, R. Rev1 wbdR tagged vaccines against Brucella ovis. Vet. Res. 2019, 50, 95. [Google Scholar] [CrossRef] [Green Version]
- Gerdts, V.; van Drunen Littel-van den Hurk, S.; Potter, A. Protection of neonates and infants by maternal immunization. Expert Rev. Vaccines 2016, 15, 1347–1349. [Google Scholar] [CrossRef] [Green Version]
- Pelekanos, M.; Leinenga, G.; Odabaee, M.; Odabaee, M.; Saifzadeh, S.; Steck, R.; Gotz, J. Establishing sheep as an experimental species to validate ultrasound-mediated blood-brain barrier opening for potential therapeutic interventions. Theranostics 2018, 8, 2583–2602. [Google Scholar] [CrossRef]
- Pernot, M.; Aubry, J.F.; Tanter, M.; Boch, A.L.; Marquet, F.; Kujas, M.; Seilhean, D.; Fink, M. In vivo transcranial brain surgery with an ultrasonic time reversal mirror. J. Neurosurg. 2007, 106, 1061–10066. [Google Scholar] [CrossRef] [Green Version]
- Cohen, Z.R.; Zaubermann, J.; Harnof, S.; Mardor, Y.; Nass, D.; Zadicario, E.; Hananel, A.; Castel, D.; Faibel, M.; Ram, Z. Magnetic resonance imaging-guided focused ultrasound for thermal ablation in the brain. Neurosurgery 2007, 60, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Kohler, F.; Gkogkidis, C.A.; Bentler, C.; Wang, X.; Gierthmuehlen, M.; Fischer, J.; Stolle, C.; Reindl, L.M.; Rickert, J.; Stieglitz, T.; et al. Closed-loop interaction with the cerebral cortex: A review of wireless implant technology. Brain Comput. Interfaces 2017, 4, 146–154. [Google Scholar] [CrossRef]
- Banstola, A.; Reynolds, J.N.J. Mapping sheep to human brain: The need for a sheep brain atlas. Front. Vet. Sci. 2022, 9, 961413. [Google Scholar] [CrossRef] [PubMed]
- Liyanage, K.A.; Steward, C.; Moffat, B.A.; Opie, N.L.; Rind, G.S.; John, S.E.; Ronayne, S.; May, C.N.; O’Brien, T.J.; Milne, M.E.; et al. Development and implementation of a Corriedale ovine brain atlas for use in atlas-based segmentation. PLoS ONE 2016, 11, e0155974. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Criteria |
|---|
| Ethical regulations and public scrutiny Cost and availability |
| Anatomical and physiological comparability |
| Adaptability to experimental manipulation |
| Genetic similarity, where applicable |
| Behavioural phenotype |
| Resistance to infections and disease progression |
| Investigation of spontaneous (natural) or induced (experimental) response |
| Transferability of information between species |
| Physiological Variables | Rodents | Sheep | Human | Reference |
|---|---|---|---|---|
| Adult body weight (kg) | 0.3–4.0 | 60–70 | 70–80 | [54,55] |
| Lifespan (y) | 2.5–3.5 | 10–30 | 70–80 | [25,56,57] |
| Brain weight (kg) | 0.013 | 0.13–0.14 | 1.3–1.4 | [58] |
| Rectal temperature (°C) | 38–39 | 38–39.5 | 36.7–37.5 | [55,59,60] |
| Respiratory rate (breaths/min) | 66–114 | 15–40 | 9–20 | [55,59,61] |
| Respiratory dead space (mL) | 0.6–1.25 | 100 | 150 | [55,59] |
| Tidal volume (mL/kg) | 0.6–1.25 | 4–9 | 7 | [59,62] |
| Heart rate (beat/min) | 330–480 | 50–88 | 50–100 | [55,59,61] |
| Maximum heart rate (beats/min) | 370–580 | 260–280 | 140–150 | [59,62] |
| Mean arterial pressure (mm Hg) | 119–122 | 70 | 70–105 | [59,63] |
| Cardiac output (L/min) | 0.12–0.13 | 1.5–13.2 | 4–8 | [59,64,65] |
| Stroke volume (mL/beat) | 0.41–0.43 | 74 | 60–100 | [59,64] |
| Extracellular fluid volume (mL/kg) | 300 | 246 | 260 | [59] |
| Plasma volume (mL/kg) | 30.8–36.7 | 37 | 43 | [55,59] |
| Blood volume (mL/kg) | 56–71 | 49 | 70 | [55,59] |
| Haemoglobin (g/100 mL) | 11–19 g/dL | 9–15 | 14–16 | [55,59] |
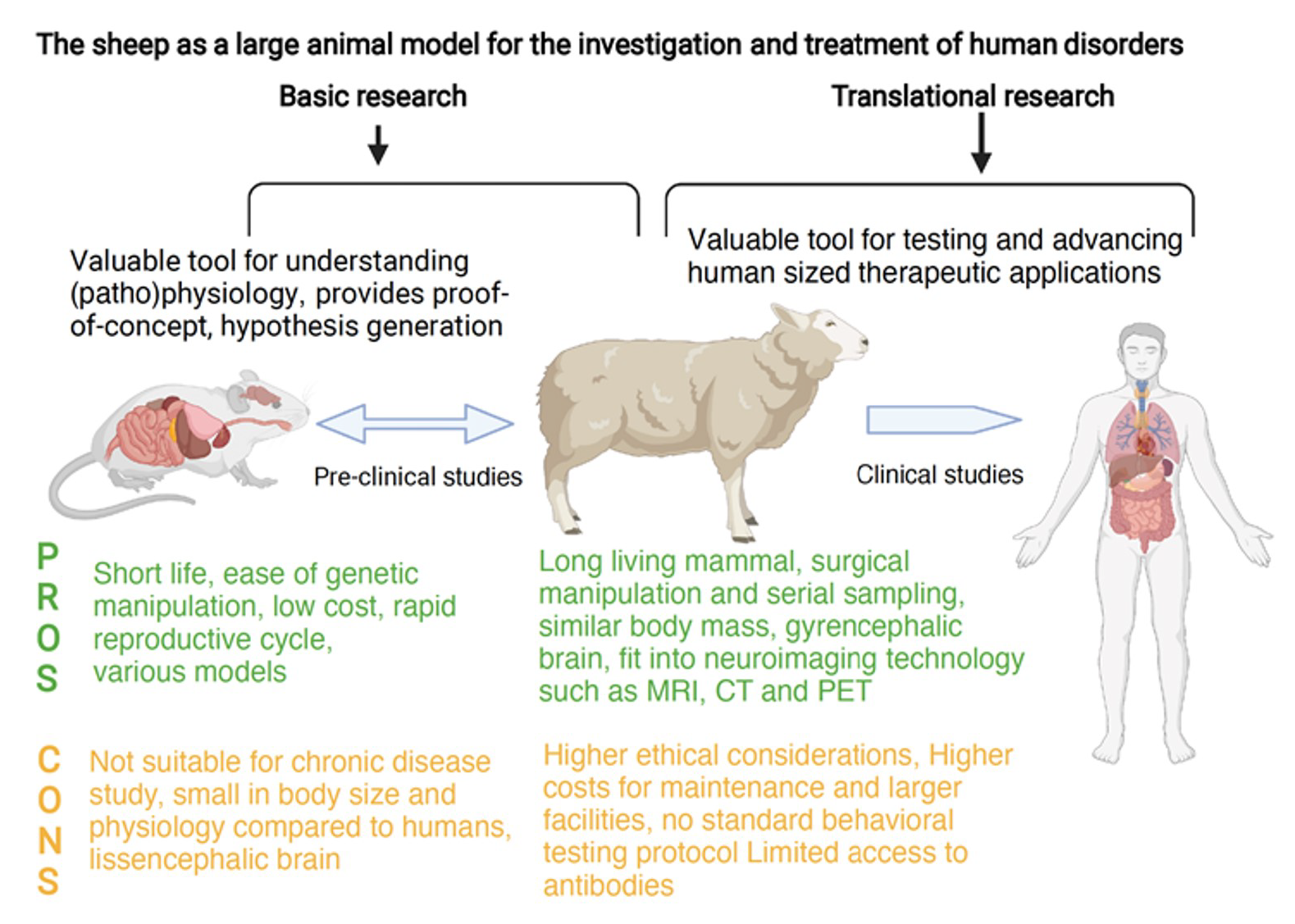
| Factors To Consider | Pros | Reference |
|---|---|---|
| Accessibility for research | Greater acceptability to animal ethics committees cf. companion animals | [26,53] |
| Easily available, reasonably outbreed, less expensive to buy cf. other large species | [10,29,53,70] | |
| Long living mammal (10–20 years) with body size, weight and brain comparable to the human | [19,29,57,58,70,71,72,73] | |
| Easy management, Surgical manipulation | [14,29,70,74,75] | |
| Body system specific advantages | Brain: higher degree of white matter and gyrencephalic structure, strong cerebellar tentorium cortical organization | [19,57,58,70,72,73] |
| Excellent animal model to study HD due to HTT gene | [69] | |
| Long bones with dimensions suitable for the deployment of implant systems | [14,29,58] | |
| Excellent animal model to study osteoporosis due to long bones, early brain development | [14,29,70] | |
| Suitable to study the main physiological systems: cardiovascular, orthopedic, endocrine, respiratory, renal, nervous and reproductive systems, host organisms for virus infection | [10,14,29,58,70] | |
| Others | Environmental enrichment not required as they live in their natural pasture | [58] |
| Increased clinical translation and more accurate indication (as the dosage, drug distribution, and safety of potential therapies trialed) | [66] | |
| Ability to use clinically relevant technology to provide clinically translational measures, such as MRI, CT, and PET; ability to use clinical equipment such as anesthesia, physiological monitoring, surgical equipment | [53,76] | |
| Strong, can carry a backpack with transmitting devices | [58] | |
| Natural model for human infections such as parainfluenza, E.Coli. | [77] | |
| Cons | ||
| Accessibility for research | Higher ethical considerations than small animals | [29,53] |
| Higher costs for maintenance and larger facilities required to perform procedures than small animals | [10,29,53,70] | |
| Body system-specific advantages | Longer gestation time compared with small animals; Uniparous in breeding, difficulty scaling up the number of offspring | [19,29,57,58,70,71,72,73] |
| Not suitable for studying neurobiology of fine motor control and binocular eye movements | [14,29,70,74,75] | |
| Transgenic selection and production of transgenic strains limited | [69] | |
| Poor availability of physiological databases for mapping to humans including atlases | [14,20,70] | |
| Others | Vital sign monitoring needed by a veterinary practitioner during all surgical procedures | [19,57,58,70,72,73] |
| Unlike, rodents, no standard behavioral testing protocol | [14,29,58] | |
| Limited access to antibodies | [10,14,29,58,70] |
| Body System | Disease or Syndrome Model | Strain/ Sex | Age/ Body Weight | Description/Observation | Reference |
|---|---|---|---|---|---|
| Cardiovascular | Ovine endo-toxemic shock (macrocirculation) | Sheep/F | Adult | The macro- and microvascular effects of selective and nonselective potassium channel inhibitors studied in ovine endotoxemic shock. | [78] |
| Angiography of the cardiac coronary venous system | Lacaune/M | 68 ± 5.3 kg | The general organization of the coronary venous circulation evaluated from clinical angiographic studies. | [54] | |
| Tissue-Engineered(TE) heart valves replacement | Swifter/F | 1 y | Immunological markers and expression of proteins specific to sheep validated for the immunohistochemical analysis of tissue-engineered heart valve after implantation in a sheep model. | [79] | |
| Beating heart sheep model | Sheep | 42 ± 5 kg | Structural and functional outcomes of trans-ventricular implantation of a modified stented bovine pulmonary valve assessed. | [80] | |
| Atrioventricular block model | Castrated, merino cross/M | 1 y/ 63.1 ± 5.6 kg | Atrioventricular block by radiofrequency ablation of the His bundle and implantation of the pacemaker into the right ventricular apex developed, characterized and validated in adult sheep. | [81] | |
| Extarterectomy model | Sheep/M | 11/2 y/ 39–48 kg | Tumor removal by extarterectomy technique and its long term effect of on the vascular wall and arterial blood flow investigated in male sheep. | [82] | |
| Chronic heart failure | Sheep | NA | Testing and optimizing of surgical therapies for chronic heart failure. | [28] | |
| Endocrine | Polycystic ovarian syndrome | Scottish Greyface/F | Adult | Prenatal testosterone (T)-treated female sheep showed reproductive deficits comparable to women with polycystic ovarian syndrome (PCOS). | [83,84] |
| Maternal-Fetal sheep model | Dorse/F | Adult pregnant | a maternal-fetal pharmacokinetic model of propofol in pregnant ewes successfully developed. | [85] | |
| Pregnancy model | sheep | Adult pregnant | Placental development, oxygen and nutrient transfer between maternal-fetal interaction are similar to human pregnancy. | [86] | |
| Immune | Autoimmune/autoinflammatory syndrome induced by adjuvants (ASIA syndrome) | Spanish castrated lamb/M | 3 months | Repetitive inoculation of aluminium-containing adjuvants through vaccination showed acute and chronic neurological episode resulting in ASIA syndrome that can be used to model similar disease affecting both human and animals. | [87] |
| Nervous | Peripheral nerve regeneration | Sheep | NA | Similar size and regeneration behavior of nerves supports the use of sheep as a model for studying peripheral nerve regeneration following nerve injury. | [88] |
| Neurogenesis | Sheep | NA | Distinctive feature of hypothalamic, olfactory and hippocampal neurogenesis in adult sheep and its contribution to reproduction, odour processing and maternal behavioral revealed. | [89] | |
| Neurogenesis | Romney/ Suffolk | 3–6 y/ 53.1–59.8 | Cell proliferation in the subventricular and subgranular zone of adult sheep is comparable with human and has the same distinct layers. | [90] | |
| Transient stroke | Merino/ M and F | 18–36 months/ 65 ± 7 kg | A survival model of sheep transient middle cerebral artery occlusion and the temporal profile of intracranial pressure change following transient stroke developed in sheep. | [53,76] | |
| Occlusion (permanent middle cerebral artery) stroke model | Outbreed adult hornless merino /M | Adult/ 42–65 kg | The permanent middle cerebral artery (MCA) occlusion results into cerebral ischemia and produces reproducible neurologic dysfunctions and can be modified by altering the occlusions in MCA. | [91] | |
| Acute proximal middle cerebral artery ischemic stroke | Merino/M and F | 18–24 months/50.1 ± 5.8 kg | A surgical model of permanent and transient MCA stroke in the sheep developed. | [92] | |
| Spina bifida | Lamb | NA | The congenital anomaly of CNS (spina bifida phenotype) with and without myelotomy comprehensively and reliably characterised in fetal lamb. | [93] | |
| Axonal injury | Merino/F | 2 y | The physiological and pathological changes resulting from traumatic injury using immunostaining as a marker of early axonal injury developed to established a head impact model of axonal injury in sheep. | [94] | |
| Transgenic Huntington’s disease | Sheep/F | 1 and 7 months | Six transgenic founder sheep generated, expressing full length human HTT with a poly- glutamine region of 73 residues. | [75] | |
| Alzheimer’s disease | Sheep | 8–14 y | The processing of amyloid protein, total tau and neurofilament markers in the aged sheep comparable to those found in sheep. | [95] | |
| Parkinson’s diseases | Columbia-Suffolk cross bred/F | 1–3 y/ 40–70 kg | Infusion of MPTP and MPP+ through jugular cannula produces parkinsonian-like behavior in sheep. | [96] | |
| Parkinson’s diseases | Rambouilette, ranch bred/F | 1–3 y/ 40–55 kg | Unilateral acute (over 30 min) and chronic (over 1 week) intracarotid injection of MPTP (0.4–5.0 mg/kg) via slow continuous infusion produces Parkinsonian-like behavior in sheep. | [74] | |
| Sleep and neurological dysfunction | Borderdale | 14.3 ± 0.5 months | Electroencephalography study performed in CLN5 batten disease-affected sheep to characterize the sleep and neurological dysfunction. | [97] | |
| Motor neuron syndrome | Border Leicester dominant cross/ | 18 months/ 34–45 kg | Molybdenum deprivation, purine ingestion and an astrocyte-associated motor neurone syndrome produced 18–27 months later in sheep. | [98] | |
| CLN5 Batten disease | Borderdale | 2 to 3 months | Efficacy of CLN5 gene therapy on the CNS monitored in a sheep model of CLN5 batten disease. | [99] | |
| Focal epilepsy | Merino/F | Adult | Focal epilepsy in sheep generated with injection of penicillin into the right prefrontal cortex and studied with the use of fMRI and iEEG. | [100] | |
| Neuropathic pain model | Polypay sheep/F | 75 kg | A neuropathic pain model established by tight ligation and axotomy of the common peroneal nerve and analgesic effect of morphine studied in sheep. | [101] | |
| Ovine ceroid lipofuscinosis (CLN6) | South Hampshire lamb/M and F | 9–12 month | Linkage between ovine ceroid lipofuscinoses (CLN6) and microsatellite markers OAR 7 q13–15 established. | [102] | |
| Spinal cord injury | Suffolk | 2 y | A model of spinal cord injury established by hemi sectioning of the spinal cord (left side) and injury quantified by a gait analysis of pre an post injury in ovine. | [103] | |
| Fetal Alcohol Spectrum Disorder | Sheep/F | NA | Pregnant sheep were exposed to binge alcohol consumption for a three-trimester period and plasma MiRNA profile was assessed from pregnant and from newborn. | [104] | |
| Acute neuronopathic Gaucher disease | Lamb | 2 h after birth | Acute neuronopathic Gaucher disease model developed in lamb by mutation in the β-glucocerebrosidase gene C381Y, which is equivalent to human C342Y. | [105] | |
| Non-accidental head injury | Lamb | 5–9 days old/ 5–12 kg | Axonal injury, neuronal reaction, and albumin extravasation examined in the hemispheric white matter, brainstem and at the intracranio cervical junction after manual shaking of head in lamb. | [106] | |
| Gastro-intestinal | Colorectal Mucosal Injury | Yearling virginal/F | NA | Due to similar gross and microscopic between sheep and human rectum Optical coherence tomography imaging and colonoscopy used to visualize morphological abnormalities and scoring of microbicide-induced injury in sheep model. | [107] |
| Intestinal loop model (gut cannulation) | Canadian Arcott/F | 8–10 months | Surgical method of catherization of intestinal loops without affecting health or loop function developed to elucidate the host response to various treatments within the small intestine of ruminants. | [108] | |
| Respiratory | Fetal lung inflammation | Merino/F | NA | Acute systemic inflammatory response of the 10 mg e.coli LPS into amniotic fluid showed three major fetal surfaces exposed to inflammatory mediators in pregnancy (the lung, gastro-intestinal tract and skin/amnion). | [109] |
| Ovine pulmonary adenocarcinoma | Sheep/F | 39–65 kg | Sheep model of naturally occurring lung cancer, ovine pulmonary adenocarcinoma caused by jaagsiekte sheep retrovirus has similar histological characteristics of human lung adenocarcinomas. | [110] | |
| Ocular | Ophthalmic model | Coopworth × Texel/M | 10–12 months/ 50–60 kg | The anatomical and clinical characteristics of sheep such as ocular response, central corneal thickness, topographic maps, intraocular pressure, corneal hysteresis, and corneal resistance factor confirmed the suitability of sheep as a model for ophthalmic experiments. | [111] |
| Opthalmic surgery training | Afshar | 1 y | Sheep eye for ophthalmic surgery training in skills laboratory found similar ultrasonic graphic and physical biometric description. | [112] | |
| Vision prosthesis model | NA | Development and surgical implantation of a vision prosthesis model into the ovine eye. | [113] | ||
| Corneal transplantation | Merino | Adult/44–68 kg | Corneal graft rejection in the sheep is macroscopically and histologically similar to human corneal graft rejection. | [114] | |
| Musculoskeletal | Collagen-induced arthritis | Merino (F) | 2 y | Bovine collagen type II injected into the hock joint and the histopathological scoring system was developed to established a collagen induced arthritis model in sheep. | [115] |
| Nociceptive withdrawal reflex | Swiss alpine | 2–3 y/ 63.1 ± 6.1 kg | Nociceptive withdrawal reflex induced by electrical stimulation in the thoracic and pelvic limb used as a tool to evaluate nociception in conscious non-medicated sheep. | [116] | |
| Osteoporosis | Corriedale/F | 2 y | Osteoporosis induced by hypothalamic-pituitary disconnection in sheep model to determine sustainability of bone loss and its biomechanical relevance. | [117] | |
| Spinal orthopedic model | Merino/F | 2 y/62.5 ± 5.3 kg | CT scan from sheep L1 to L6 showed similar vertebral endplates and spinal canal to humans supporting sheep as a model for human orthopedic spinal research. | [118] | |
| Hypophosphatasia | Sheep | NA | Using CRISPR/Cas9,a single point mutation in the tissue nonspecific alkaline phosphatase (TNSALP) gene (ALPL) made to generated hypophosphatasia- rare human bone disease. | [119] | |
| Cancellous bone healing | Sheep | 18 months and 5 y | A sheep model for cancellous bone healing surgically created to assess early healing and biological changes of the medial distal femoral and proximal tibial epipheses bilaterally. | [120] | |
| Artificial bone substitute | Mecklenburg | 63 kg | Artificial bone substitute (Nanobone) used in bone defect model of ovine tibial metaphysis appears to be highly potent bone substitute with osteoconductive properties. | [121] | |
| Skin | Skin wound healing model | Sheep/M | 6 months/ 20–25 kg | Topical administration of platelet-rich plasma improved skin wound healing process in sheep making a good model for regenerative medicine research. | [122] |
| Second intention healing model | Bergamasca/F | NA | Peripheral blood-derived MSCs improve the quality of wound healing by accelerating granulation, reepithelization and neovascularization both for superficial injuries and deep lesions. | [123] | |
| Urinary | Renal ischaemia–reperfusion injury | Sheep/F | 35–40 kg | The reno-protective effects of zinc preconditioning in renal ischaemia reperfusion injury was assessed in a sheep model and found to be beneficial for human applications. | [20] |
| Uterine transplantation | Biparous (Prealpes and Romane)/F | 35–70 months/ 57–91 kg | Uterine transplantation method set-up using end to end anastomosis of external iliac vessels in sheep and complications were highlighted. | [124] | |
| Renal function | Segurena/M and F | 2–6 y/50 kg | Simplified Iohexol-based method was developed to measure renal function using glomerular filtration rate as a model of renal diseases in sheep. | [125] | |
| Haemodialysis treatment | Merino/M | 16–18 months | The two-step bilateral nephrectomy was conducted to develop ovine model of haemodialysis treatment to measure dialysis adequacy and urea reduction ratio for each haemodialysis treatment. | [126] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banstola, A.; Reynolds, J.N.J. The Sheep as a Large Animal Model for the Investigation and Treatment of Human Disorders. Biology 2022, 11, 1251. https://doi.org/10.3390/biology11091251
Banstola A, Reynolds JNJ. The Sheep as a Large Animal Model for the Investigation and Treatment of Human Disorders. Biology. 2022; 11(9):1251. https://doi.org/10.3390/biology11091251
Chicago/Turabian StyleBanstola, Ashik, and John N. J. Reynolds. 2022. "The Sheep as a Large Animal Model for the Investigation and Treatment of Human Disorders" Biology 11, no. 9: 1251. https://doi.org/10.3390/biology11091251