
Investigation of Meat from Ostriches Raised and Slaughtered in Bavaria, Germany: Microbiological Quality and Antimicrobial Resistance

 ,
,  , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Detection of Trichinella spp.
2.3. General Microbiological Status
2.4. Detection of the Shiga Toxin-Producing Escherichia coli Genes stx1 and stx2 Including stx2f
2.5. Phenotypical Antimicrobial Susceptibility of Enterobacteriaceae and Enterococcus spp.
3. Results and Discussion
3.1. Detection of Trichinella spp.
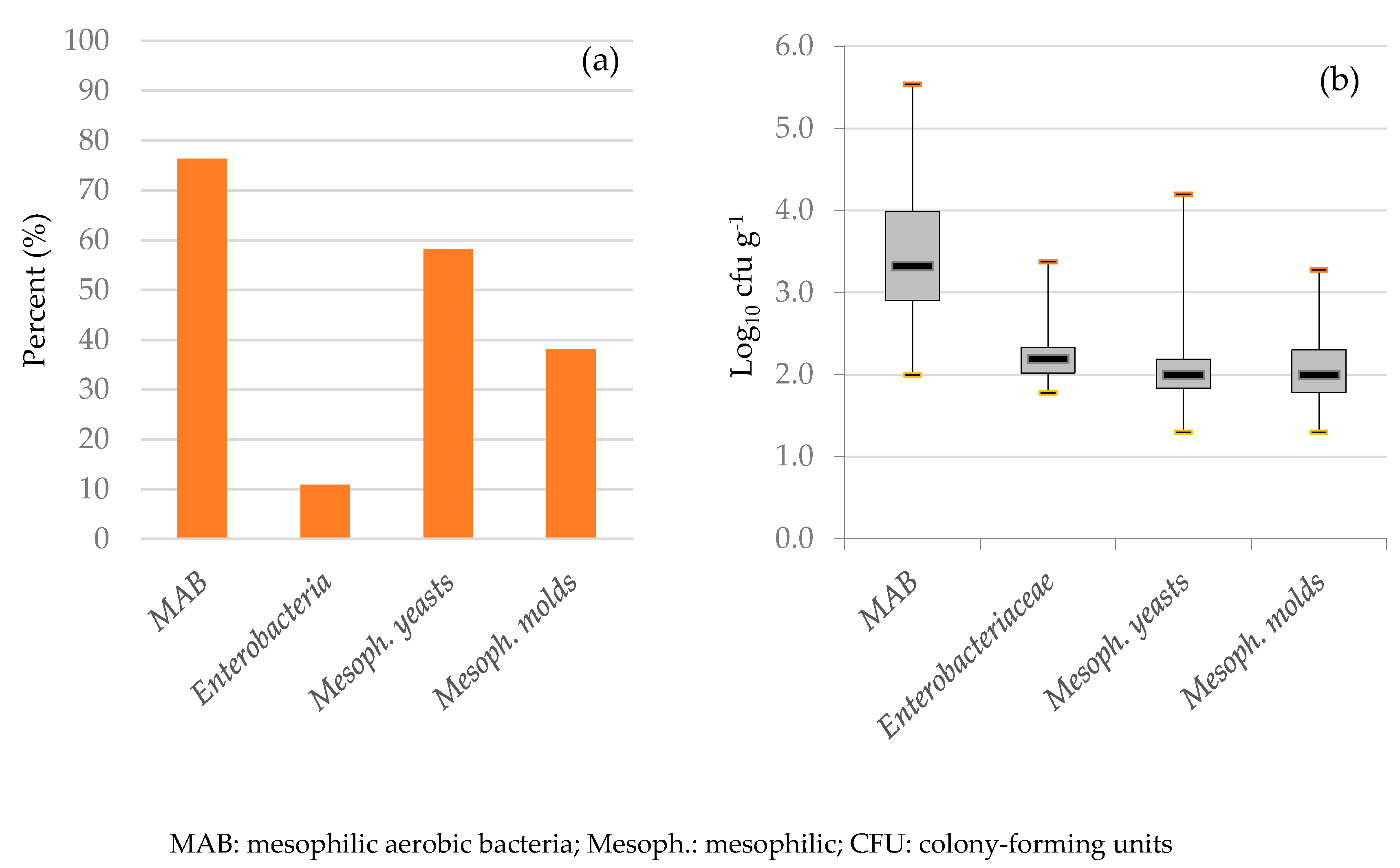
3.2. Microbiological Examination
3.3. Detection of the Shiga Toxin-Producing E. coli Genes stx1 and stx2 including stx2f
3.4. Phenotypical Antimicrobial Susceptibility
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Isolate No. | Antimicrobials—MIC Values (μg/mL) | |||||||||||||||||||||||
| AMP [0.25–8] | AMI [16–32] | FAZ [2–4] | CEP [2–4] | POD [2–8] | FOV [0.05–8] | CPT [0.12–4] | AXO [8–64] | CHL [2–32] | CIP [1–4] | CLI [0.5–4] | DAP [0.5–4] | ENRO [0.25–4] | ERY [0.25–4] | GEN [4–16] | ||||||||||
| 17K | 1 | >32 | >4 | >4 | >8 | >8 | 0.5 | >64 | 8 | 4 | >4 | >4 | 4 | >4 | 16 | |||||||||
| 18K | 1 | 32 | >4 | >4 | >8 | >8 | 0.5 | 32 | 8 | ≤1 | >4 | >4 | 1 | >4 | 8 | |||||||||
| 21K | 1 | 32 | >4 | >4 | >8 | >8 | 0.5 | >64 | 8 | ≤1 | 4 | >4 | 2 | >4 | 16 | |||||||||
| 25K | 1 | 32 | >4 | >4 | >8 | >8 | 0.5 | >64 | 8 | 4 | ≤0.5 | >4 | 4 | >4 | 8 | |||||||||
| Isolate No. | IMI [1–4] | LEVO [0.25–4] | LZD [1–8] | MAR [1–4] | MXF [0.25–4] | NIT [16–64] | OXA+ [0.25–2] | PRA [0.25–2] | PEN [0.05–8] | RIF [1,2] | TLA [0.015–0.25] | TET [0.25–8] | TGC [0.03–0.5] | VAN [0.25–16] | ||||||||||
| 17K | 2 | 4 | 2 | 4 | 2 | 64 | >2 | 2 | 4 | >2 | 0.03 | ≤0.25 | ≤0.03 | 0.5 | ||||||||||
| 18K | 2 | 2 | 2 | 2 | 0.5 | 64 | >2 | ≤0.25 | 4 | >2 | 0.03 | 0.5 | ≤0.03 | 0.5 | ||||||||||
| 21K | 4 | 2 | 2 | 4 | 0.5 | 64 | >2 | 0.5 | 4 | >2 | 0.03 | ≤0.25 | 0.06 | 0.5 | ||||||||||
| 25K | 2 | 4 | 2 | 4 | 2 | 64 | >2 | 2 | 8 | >2 | ≤0.015 | ≤0.25 | ≤0.03 | 0.5 | ||||||||||
| Antimicrobials—MIC Values (μg/mL) | |||||||
|---|---|---|---|---|---|---|---|
| Isolate No. | Species * | CTX [0.25–16] | CTL [0.016–1] | CAZ [0.5–32] | CAL [0.064–4] | FEP [0.25–16] | FEL [0.5–32] |
| 5F | K. aerogenes | ≥16 | ≥1.0 | 12 | ≥4 | <0.25 | 0.094 |
| 10F | En. cloacae | ≥16 | ≥1.0 | ≥32 | ≥4 | 2 | 0.75 |
| 11F | En. cloacae | ≥16 | ≥1.0 | ≥32 | ≥4 | 0.50 | 0.75 |
| 13F | En. cloacae | ≥16 | ≥1.0 | ≥32 | ≥4 | 2 | 0.75 |
| 14F | En. cloacae | ≥16 | ≥1.0 | ≥32 | ≥4 | 2 | 1 |
| 15F | En. cloacae | ≥16 | ≥1.0 | ≥32 | ≥4 | 0.50 | 1.5 |
| 16F | K. aerogenes | ≥16 | ≥1.0 | 1.5 | <0.064 | <0.25 | 0.125 |
| 21F | En. cloacae | ≥16 | ≥1.0 | ≥32 | ≥4 | 2 | 0.50 |
| 22F | En. cloacae | ≥16 | ≥1.0 | ≥32 | ≥4 | 1.5 | 0.50 |
| 28F | En. cloacae | ≥16 | ≥1.0 | ≥32 | ≥4 | 0.25 | 0.75 |
| 29bF | En. cloacae | ≥16 | ≥1.0 | ≥32 | ≥4 | <0.25 | 0.19 |
| 42F | P. septica | 3 | ≥1.0 | 0.125 | <0.064 | <0.25 | 0.094 |
| Antimicrobials—MIC Values (μg/mL) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Isolate No. | Species * | FOX [0.5–32] | AZI [0.12–16] | CHL [4–32] | TET [4–32] | AXO [0.25–64] | AUG02 [1/0.5–32/16] | CIP [0.015–4] | GEN [0.25–16] | NAL [0.5–32] | XNL [0.12–18] | FIS [16–256] | SXT [0.12/2.38–4/76] | AMP [1–32] | STR [2–64] |
| 5F | K. aerogenes | >32 | 4 | 4 | ≤4 | 32 | 32/16 | 0.03 | 2 | 2 | >8 | 64 | ≤0.12/2.38 | >32 | 8 |
| 10F | En. cloacae | >32 | 16 | 4 | ≤4 | >64 | >32/16 | 0.03 | ≤0.25 | 2 | >8 | 64 | ≤0.12/2.38 | >32 | ≤2 |
| 11F | En. cloacae | >32 | 16 | 8 | ≤4 | 4 | >32/16 | ≤0.015 | 0.5 | 2 | 4 | 64 | ≤0.12/2.38 | >32 | ≤2 |
| 13F | En. cloacae | >32 | 16 | 8 | ≤4 | >64 | >32/16 | 0.03 | 0.5 | 2 | >8 | 64 | ≤0.12/2.38 | >32 | 32 |
| 14F | En. cloacae | >32 | 16 | 4 | ≤4 | >64 | >32/16 | 0.03 | 0.5 | 2 | >8 | 64 | ≤0.12/2.38 | >32 | ≤2 |
| 15F | En. cloacae | >32 | >16 | 8 | ≤4 | 16 | >32/16 | 0.03 | 0.5 | 4 | 8 | 64 | ≤0.12/2.38 | >32 | ≤2 |
| 16F | K. aerogenes | >32 | 8 | 8 | ≤4 | 32 | >32/16 | 0.06 | 2 | 4 | >8 | 64 | ≤0.12/2.38 | >32 | 8 |
| 21F | En. cloacae | >32 | 16 | 4 | ≤4 | >64 | 32/16 | ≤0.015 | 0.5 | 2 | >8 | ≤16 | ≤0.12/2.38 | >32 | ≤2 |
| 22F | En. cloacae | >32 | 8 | 4 | ≤4 | >64 | 32/16 | 0.03 | ≤0.25 | 2 | >8 | 32 | ≤0.12/2.38 | >32 | ≤2 |
| 28F | En. cloacae | >32 | 16 | 8 | ≤4 | >64 | 32/16 | 0.03 | ≤0.25 | 2 | >8 | 64 | ≤0.12/2.38 | >32 | ≤2 |
| 29aF | E. coli | 32 | 8 | 8 | ≤4 | 1 | 32/16 | 0.03 | 0.5 | 2 | 2 | ≤16 | ≤0.12/2.38 | >32 | 4 |
| 29bF | En. cloacae | >32 | 16 | 8 | ≤4 | 64 | >32/16 | 0.03 | ≤0.25 | 4 | >8 | 64 | ≤0.12/2.38 | >32 | ≤2 |
| 42F | P. septica | >32 | 1 | 4 | ≤4 | 0.5 | 8/4 | 0.06 | ≤0.25 | 8 | 4 | 64 | ≤0.12/2.38 | 32 | ≤2 |
References
- Horbañczuk, J.; Sales, J.; Celeda, T.; Konecka, A.; Ziêba, G.; Kawka, P. Cholesterol content and fatty acid composition of ostrich meat as influenced by subspecies. Meat Sci. 1998, 50, 385–388. [Google Scholar] [CrossRef]
- Al-Khalaifa, H.; Al-Nasser, A. Ostrich meat: Production, quality parameters, and nutritional comparison to other types of meats. J. Appl. Poult. Res. 2014, 23, 784–790. [Google Scholar] [CrossRef]
- Horbańczuk, J.O.; Tomasik, C.; Cooper, R.G. Ostrich Farming in Poland—Its History and Current Situation after Accession to the European Union. Avian Biol. Res. 2008, 1, 65–71. [Google Scholar] [CrossRef]
- Kistner, C. Ostrich Production in Germany—Ecological way to economic success. Lohmann Inf. 2019, 53, 16–23. [Google Scholar]
- STMUV. Antwort auf Schriftliche Anfrage der Abgeordneten Rosi Steinberger BÜNDNIS 90/DIE GRÜNEN vom 09.08.2018. 17; Wahlperiode, Ed.; STMUV: Munich, Germany, 2018; p. 4. [Google Scholar]
- Hoffman, L.C.; Britz, T.J.; Schnetler, D.C. Prevalent organisms on ostrich carcasses found in a commercial abattoir. J. S. Afr. Vet. Assoc. 2010, 81, 151–155. [Google Scholar] [CrossRef]
- Gopo, J.M.; Banda, G.N. Occurrence of Salmonella on meat and products in an ostrich abattoir as determined with a DNA probe. S. Afr. J. Anim. Sci. 1997, 27, 1–6. [Google Scholar]
- Carneiro, C.; Araújo, C.; Gonçalves, A.; Vinué, L.; Somalo, S.; Ruiz, E.; Uliyakina, I.; Rodrigues, J.; Igrejas, G.; Poeta, P.; et al. Detection of CTX-M-14 and TEM-52 extended-spectrum beta-lactamases in fecal Escherichia coli isolates of captive ostrich in Portugal. Foodborne Pathog. Dis. 2010, 7, 991–994. [Google Scholar] [CrossRef]
- Ley, E.C.; Morishita, T.Y.; Brisker, T.; Harr, B.S. Prevalence of Salmonella, Campylobacter, and Escherichia coli on ostrich carcasses and the susceptibility of ostrich-origin E. coli isolates to various antibiotics. Avian Dis. 2001, 45, 696–700. [Google Scholar] [CrossRef]
- Fioretti, D.; Moretti, A.; Marini, C.; Grelloni, V.; Antognoni, M. Trichinella spp. in ostrich meat: A public health risk? Parasite 2001, 8, S203–S205. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, H.; Scheef, J.; Morabito, S.; Caprioli, A.; Wieler, L.H.; Karch, H. A new Shiga toxin 2 variant (Stx2f) from Escherichia coli isolated from pigeons. Appl. Environ. Microbiol. 2000, 66, 1205–1208. [Google Scholar] [CrossRef] [Green Version]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Piérard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EU-RL VTEC. Identification and Characterization of Verocytotoxin-Producing Escherichia coli (VTEC) by Real Time PCR Amplification of the Main Virulence Genes and the Genes Associated with the Serogroups Mainly Associated with Severe Human Infections. Available online: https://www.iss.it/web/iss-en (accessed on 3 June 2021).
- Perelle, S.; Dilasser, F.; Grout, J.; Fach, P. Detection by 5’-nuclease PCR of Shiga-toxin producing Escherichia coli O26, O55, O91, O103, O111, O113, O145 and O157:H7, associated with the world’s most frequent clinical cases. Mol. Cell Probes 2004, 18, 185–192. [Google Scholar] [CrossRef] [PubMed]
- EU-RL VTEC. Identification of the Subtypes of Verocytotoxin encoding Genes (vtx) of Escherichia coli by Conventional PCR. Available online: https://www.iss.it/web/iss-en (accessed on 3 June 2021).
- Fricker, M.; Messelhäusser, U.; Busch, U.; Scherer, S.; Ehling-Schulz, M. Diagnostic real-time PCR assays for the detection of emetic Bacillus cereus strains in foods and recent food-borne outbreaks. Appl. Environ. Microbiol. 2007, 73, 1892–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallmann, L.; Burkhardt, F. Klinische Mikrobiologie; Georg Thieme Verlag: Stuttgart, Germany, 1974. [Google Scholar]
- Seeley, H.; Vandemark, H.; Lee, J. Microbes in Action; Freeman: New York, NY, USA, 1991. [Google Scholar]
- Bruker Daltonik GmbH. MALDI Biotyper 3.1 User Manual, 1st ed.; Bruker Daltonik GmbH: Bremen, Germany, 2012. [Google Scholar]
- Esteban-Cuesta, I.; Dorn-In, S.; Drees, N.; Hölzel, C.; Gottschalk, C.; Gareis, M.; Schwaiger, K. Antimicrobial resistance of Enterobacter cloacae complex isolates from the surface of muskmelons. Int. J. Food Microbiol. 2019, 301, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Dupouy-Camet, J. Trichinellosis: A worldwide zoonosis. Vet. Parasitol. 2001, 93, 191–200. [Google Scholar] [CrossRef]
- Zimmermann, W.J. Control II Surveillance in Swine and Other Animals by Muscle Examination. In Trichinella and Trichinosis; Campbell, W.C., Ed.; Springer US: Boston, MA, USA, 1983; pp. 515–528. [Google Scholar] [CrossRef]
- BfR. German Federal Institute for Risk Assessment—Bundesinstitut für Risikobewertung (BfR). In Deutschland gibt es aus Wissenschaftlicher Sicht Keine Region mit Einem Vernachlässigbaren Trichinella-Risiko. Stellungnahme Nr. 034/2007 des BfR vom 21. Mai 2007; BfR: Berlin, Germany, 2007. [Google Scholar]
- Capita, R.; Prieto, M.; Alonso-Calleja, C. Sampling Methods for Microbiological Analysis of Red Meat and Poultry Carcasses. J. Food Prot. 2004, 67, 1303–1308. [Google Scholar] [CrossRef]
- Karama, M.; de Jesus, A.E.; Veary, C.M. Microbial quality of ostrich carcasses produced at an export-approved South African abattoir. J. Food Prot. 2003, 66, 878–881. [Google Scholar] [CrossRef]
- Brenesselová, M.; Koréneková, B.; Mačanga, J.; Marcinčák, S.; Jevinová, P.; Pipová, M.; Turek, P. Effects of vacuum packaging conditions on the quality, biochemical changes and the durability of ostrich meat. Meat Sci. 2015, 101, 42–47. [Google Scholar] [CrossRef]
- Horbańczuk, O.K.; Jóźwik, A.; Wyrwisz, J.; Marchewka, J.; Wierzbicka, A. Physical Characteristics and Microbial Quality of Ostrich Meat in Relation to the Type of Packaging and Refrigerator Storage Time. Molecules 2021, 26, 3445. [Google Scholar] [CrossRef]
- OJ L 338 22.12.2005; Regulation (EC) No 2073/2005 of the European Commission of 15 November 2005 on microbiological Criteria for Foodstuffs. European Commission: Brussels, Belgium, 2005; p. 1.
- Gill, C.; Jones, T.; Bryant, J.; Brereton, D. The microbiological conditions of the carcasses of six species after dressing at a small abattoir. Food Microbiol. 2000, 17, 233–239. [Google Scholar] [CrossRef]
- EFSA. European Food Safety Authority. The European Union One Health 2019 Zoonoses Report. EFSA J 2019, 17, 5926. [Google Scholar] [CrossRef] [Green Version]
- EFSA. European Food Safety Authority. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, 6406. [Google Scholar] [CrossRef]
- Vanhooser, S.; Welsh, R. Isolation of Salmonella Species from Ratites. J. Vet. Diagn. Investig. 1995, 7, 268–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solheim, H.T.; Sekse, C.; Urdahl, A.M.; Wasteson, Y.; Nesse, L.L. Biofilm as an environment for dissemination of stx genes by transduction. Appl. Environ. Microbiol. 2013, 79, 896–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macori, G.; McCarthy, S.C.; Burgess, C.M.; Fanning, S.; Duffy, G. Investigation of the Causes of Shigatoxigenic Escherichia coli PCR Positive and Culture Negative Samples. Microorganisms 2020, 8, 587. [Google Scholar] [CrossRef] [Green Version]
- Schwaiger, K.; Hölzel, C.; Bauer, J. Detection of the Macrolide-Efflux Protein A Gene mef(A) in Enterococcus faecalis. Microb. Drug Resist. 2011, 17, 429–432. [Google Scholar] [CrossRef]
- Klare, I.; Konstabel, C.; Badstübner, D.; Werner, G.; Witte, W. Occurrence and spread of antibiotic resistances in Enterococcus faecium. Int. J. Food Microbiol. 2003, 88, 269–290. [Google Scholar] [CrossRef]
- Franklin, A.; Acar, J.; Anthony, F.; Gupta, R.; Nicholls, T.; Tamura, Y.; Thompson, S.; Threlfall, E.J.; Vose, D.; van Vuuren, M.; et al. Antimicrobial resistance: Harmonisation of national antimicrobial resistance monitoring and surveillance programmes in animals and in animal-derived food. Rev. Sci. Et Tech.-Off. Int. Des Epizoot. 2001, 20, 859–870. [Google Scholar] [CrossRef]
- Peters, J.; Mac, K.; Wichmann-Schauer, H.; Klein, G.; Ellerbroek, L. Species distribution and antibiotic resistance patterns of enterococci isolated from food of animal origin in Germany. Int. J. Food Microbiol. 2003, 88, 311–314. [Google Scholar] [CrossRef]
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; León-Sampedro, R.; Del Campo, R.; Coque, T.M. Antimicrobial Resistance in Enterococcus spp. of animal origin. Microbiol. Spectr. 2018, 6, 6.4.24. [Google Scholar] [CrossRef]
- Fluckey, W.M.; Loneragan, G.H.; Warner, R.D.; Echeverry, A.; Brashears, M.M. Diversity and Susceptibility of Enterococcus Isolated from Cattle before and after Harvest. J. Food Prot. 2009, 72, 766–774. [Google Scholar] [CrossRef] [PubMed]
- ECDC. European Centre for Disease Prevention and Control. Antimicrobial resistance in the EU/EEA (EARS-Net)—Annual Epidemiological Report 2019; ECDC: Solna, Sweden, 2020. [Google Scholar]
- Bortolaia, V.; Espinosa-Gongora, C.; Guardabassi, L. Human health risks associated with antimicrobial-resistant enterococci and Staphylococcus aureus on poultry meat. Clin. Microbiol. Infect. 2016, 22, 130–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakoulas, G.; Rose, W.; Nonejuie, P.; Olson, J.; Pogliano, J.; Humphries, R.; Nizet, V. Ceftaroline restores daptomycin activity against daptomycin-nonsusceptible vancomycin-resistant Enterococcus faecium. Antimicrob. Agents Chemother. 2014, 58, 1494–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Critically Important Antimicrobials for human Medicine: 6th Revision. Available online: https://www.who.int/publications/i/item/9789241515528 (accessed on 15 January 2021).
- Thomson, K.S. Controversies about extended-spectrum and AmpC beta-lactamases. Emerg. Infect. Dis. 2001, 7, 333–336. [Google Scholar] [CrossRef]
- Löscher, W.; Richter, A.; Potschka, H. III Anhang—36 Zugelassene Arzneimittel für Geflügel. In Pharmakotherapie bei Haus-und Nutztieren; Enke: Stuttgart, Germany, 2014. [Google Scholar]
- Qualität und Sicherheit GmbH. Tierhaltung. Geflügelhalter/Brütereien im QS-System. Leitfäden. Available online: https://www.q-s.de/futter-tiere-fleisch/landwirtschaft-gefluegelhalter.html#leitfaeden (accessed on 18 May 2022).
- TÄHAV. Neuerungen für Geflügelpraxis (Huhn, Pute). BLTK_TÄHAV_Huhn_Pute_Stand 11.03.2018: 2018. Available online: https://www.bltk.de/fileadmin/user_upload/Tieraerzte/TAEHAV/TAEHAV_Huhn_Pute_0_.pdf (accessed on 7 June 2022).
- Korbel, R.; Baas, N.; Bergmann, S.; Erhard, W.; Engelhard, S.; Engelhard, W.; Hörndl, S.; Kienzle, E.; Rückschloss, S.; Schubert, S.; et al. Handbuch Sachkundeseminar Straußenhaltung und Straußenschlachtung. Available online: https://sachkundeseminar-straussenhaltung.de (accessed on 7 June 2022).
- Korbel, R.; Schubert, M.; Erhard, M.; Wöhr, C.; Bergmann, S.; Rückschloss, S.; Thiel, S.; Engelhardt, H.; Engelhardt, S. Betrachtungen und Empfehlungen zur artgemäßen und tierschutzgerechten Haltung von Straußenvögeln in Deutschland. Tierarztl Prax Ausg G Grosstiere Nutztiere 2015, 43, 232–244. [Google Scholar] [CrossRef] [PubMed]

| Target Gene | Primer/ Probe | Sequence (5′–3′) | Fragment Size (bp) | Reference |
|---|---|---|---|---|
| stx1/ stx2 | stx1+2-F | TTTGTYACTGTSACAGCWGAAGCYTTACG | 131/128 | [13,14] |
| stx1+2-R | CCCCAGTTCARWGTRAGRTCMACRTC | |||
| stx1-Probe | FAM-CTGGATGATCTCAGTGGGCGTTCTTATGTAA-BHQ1 | |||
| stx2-Probe | TEXASRED-TCGTCAGGCACTGTCTGAAACTGCTCC-BHQ2 | |||
| stx2f | stx2f-F | TGGGCGTCATTCACTGGTTG | 424 | Primers: [12,15] |
| stx2f-R | TAATGGCCGCCCTGTCTCC | |||
| stx2f-Probe | Cy5.5-ATTCCGACCGGCGCTGTCTGAG-BBQ-650 | Probe: this study | ||
| IAC * | IAC-F | GCAGCCACTGGTAACAGGAT | 118 | [13,16] |
| IAC-R | GCAGAGCGCAGATACCAAAT | |||
| IAC-Probe | HEX-AGAGCGAGGTATGTAGGCGG-BHQ1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beindorf, P.-M.; Kovalenko, O.; Ulrich, S.; Geißler, H.; Korbel, R.; Schwaiger, K.; Dorn-In, S.; Esteban-Cuesta, I. Investigation of Meat from Ostriches Raised and Slaughtered in Bavaria, Germany: Microbiological Quality and Antimicrobial Resistance. Biology 2022, 11, 985. https://doi.org/10.3390/biology11070985
Beindorf P-M, Kovalenko O, Ulrich S, Geißler H, Korbel R, Schwaiger K, Dorn-In S, Esteban-Cuesta I. Investigation of Meat from Ostriches Raised and Slaughtered in Bavaria, Germany: Microbiological Quality and Antimicrobial Resistance. Biology. 2022; 11(7):985. https://doi.org/10.3390/biology11070985
Chicago/Turabian StyleBeindorf, Philipp-Michael, Oksana Kovalenko, Sebastian Ulrich, Hanna Geißler, Rüdiger Korbel, Karin Schwaiger, Samart Dorn-In, and Irene Esteban-Cuesta. 2022. "Investigation of Meat from Ostriches Raised and Slaughtered in Bavaria, Germany: Microbiological Quality and Antimicrobial Resistance" Biology 11, no. 7: 985. https://doi.org/10.3390/biology11070985